CTLA-4 Blockade, during HIV Virus-Like Particles Immunization, Alters HIV-Specific B-Cell Responses

 and
and
Abstract
1. Introduction
2. Materials and Methods:
2.1. Cells and Virus-Like Particle (VLP) Production
2.2. VLP Envelope (Env) Conformation Analysis
2.3. C57BL/6J Mice Immunization and Specimen Collection
2.4. AID-Cre Mice Immunization
2.5. Flow Cytometry Analysis
2.6. Antigen-Specific Follicular T-Cell Assay
2.7. Serum IgG ELISA
2.8. Quantitative IgG Subtype ELISA
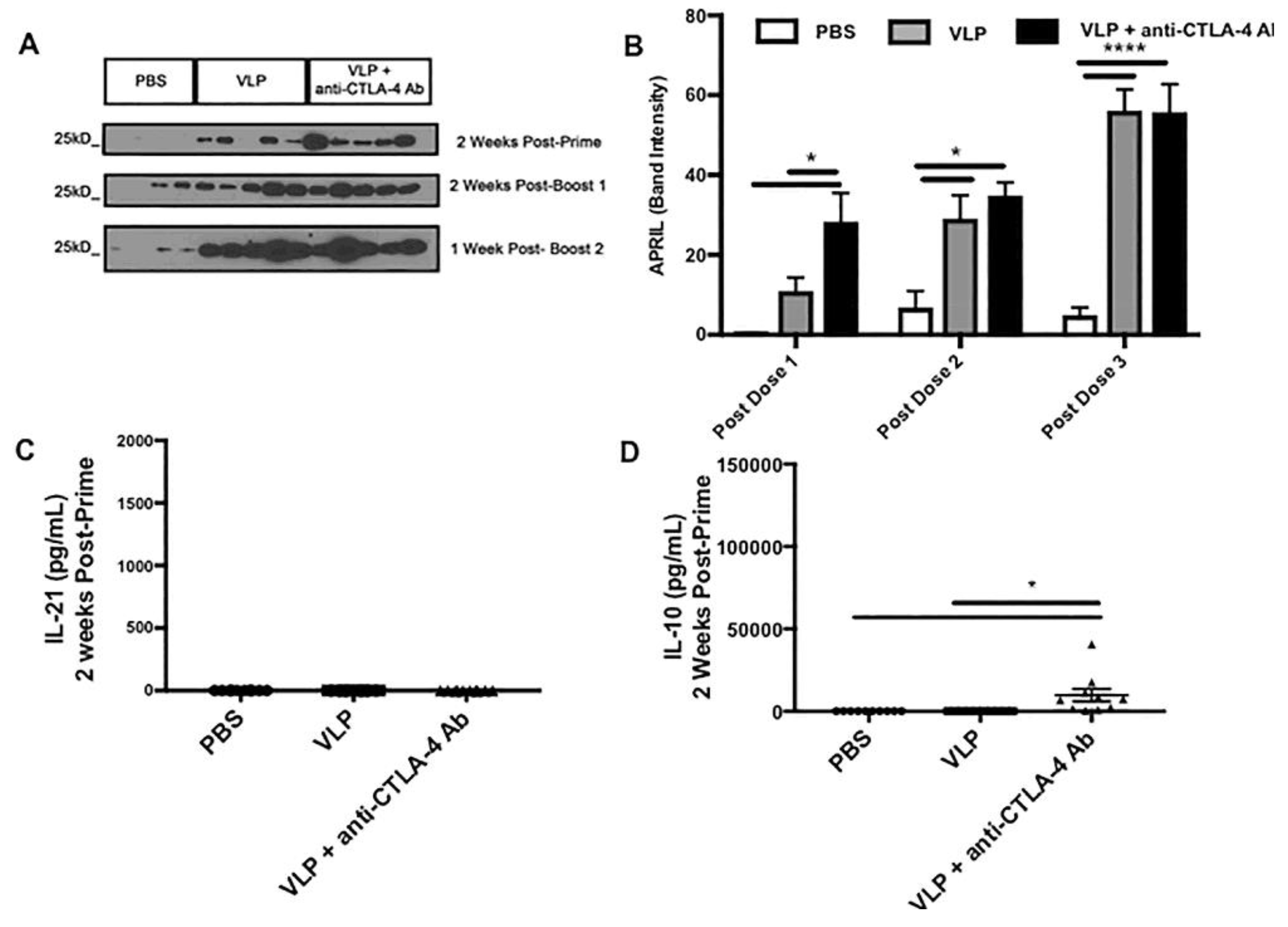
2.9. Western Blot for A Proliferation-Inducing Ligand (APRIL)
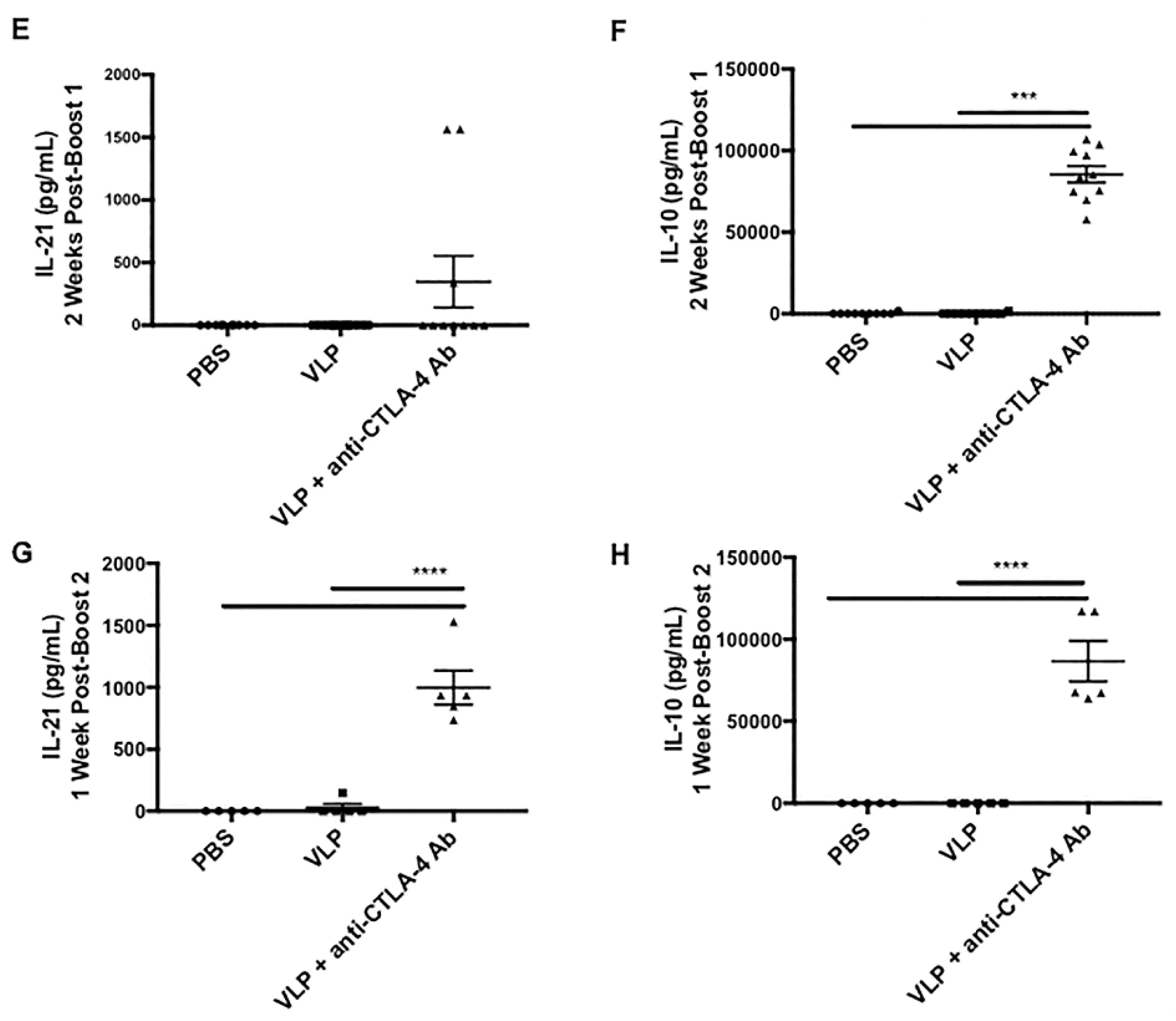
2.10. Serum Cytokine Analysis
2.11. HIV Neutralization Analysis
2.12. Avidity Analysis
2.13. Antibody Dependent Cellular Cytotoxicity
2.14. Statistical Analysis
3. Results
3.1. CTLA-4 Blockade with VLP Vaccination Results in Increased CD4+ T-Cell Activation
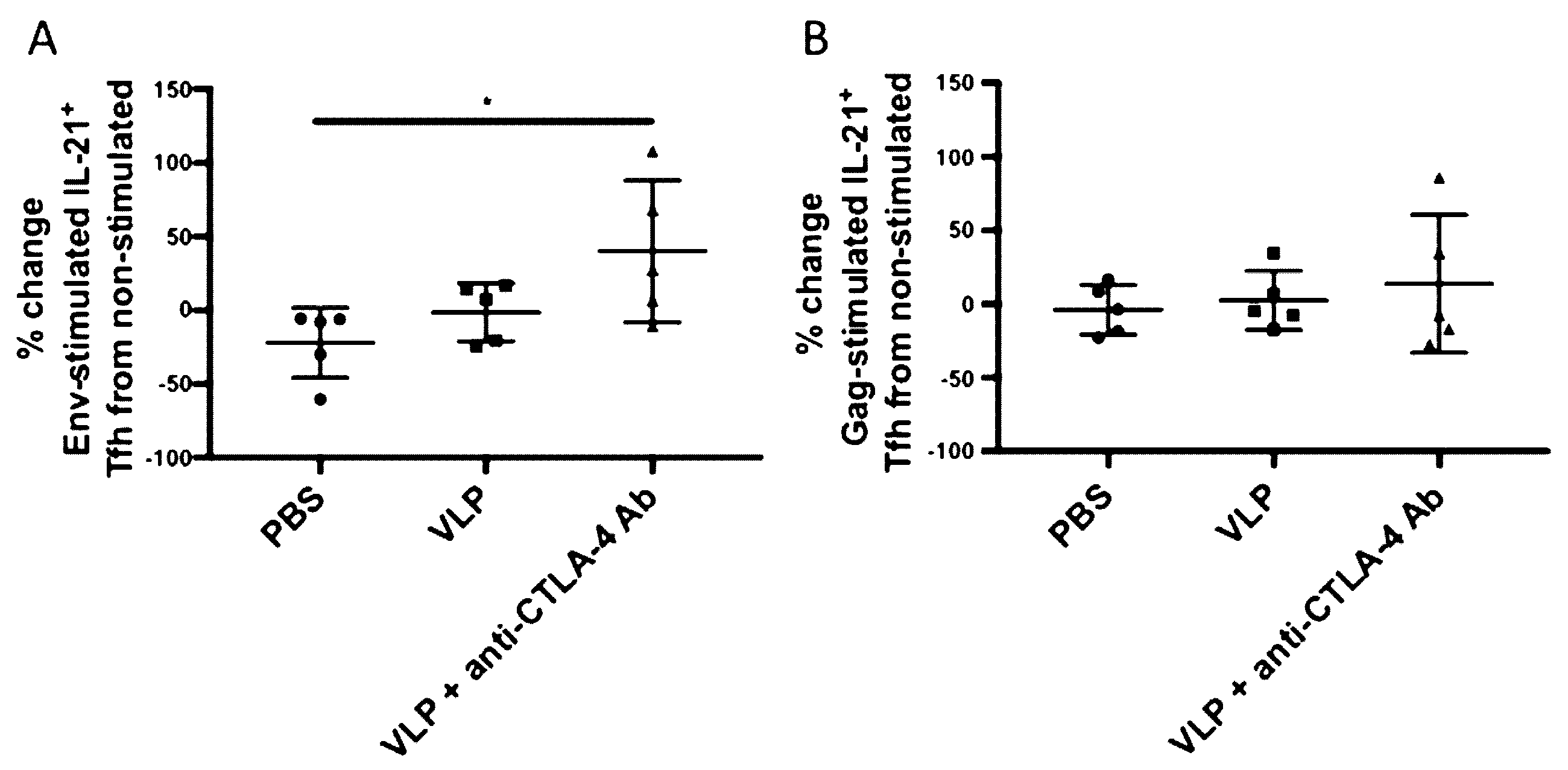
3.2. CTLA-4 Blockade Induces Increased HIV Env-Specific Tfh Cells
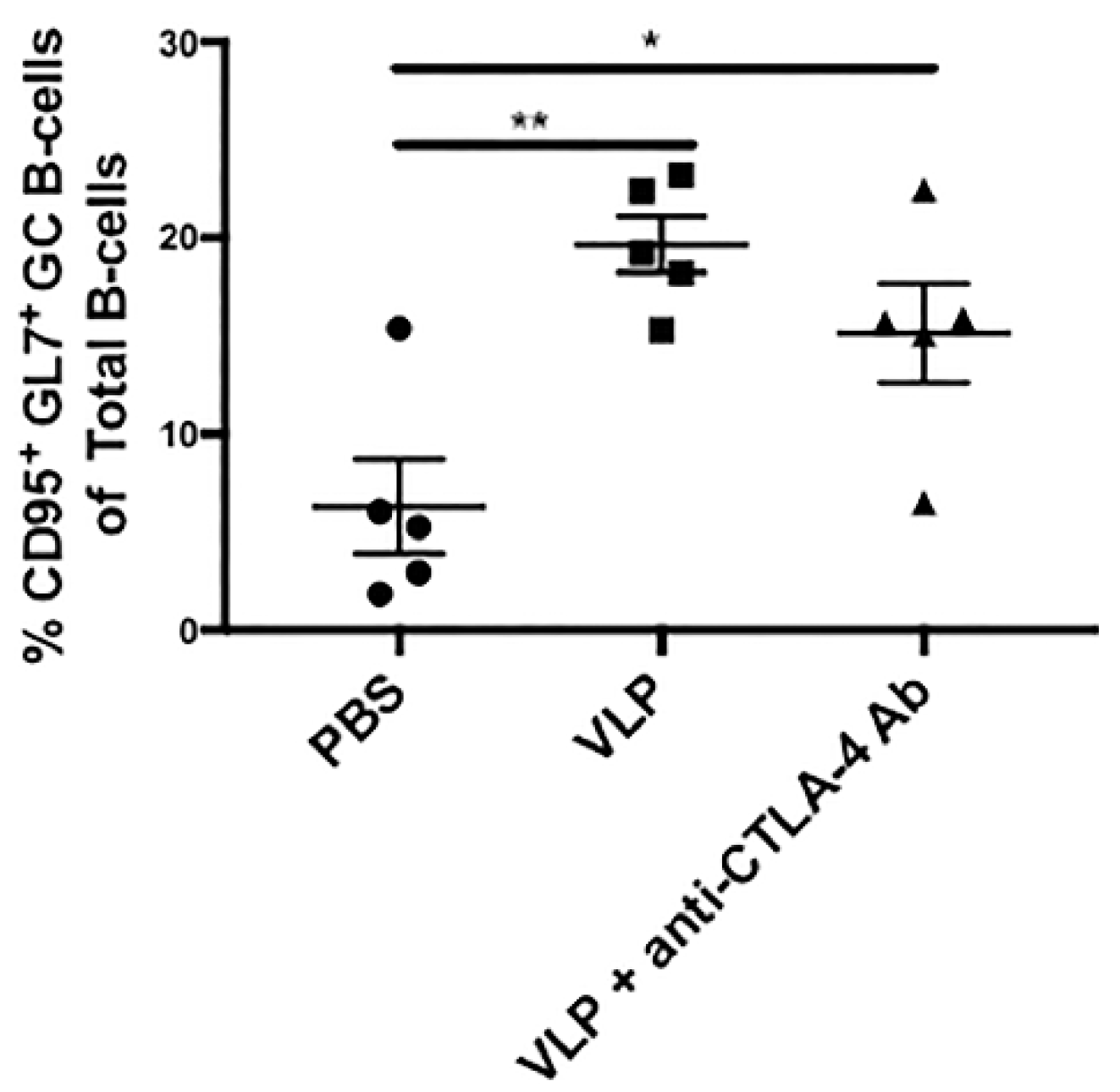
3.3. VLP Immunization Independent of CTLA-4 Blockade Promotes Increased Germinal Center B-Cells
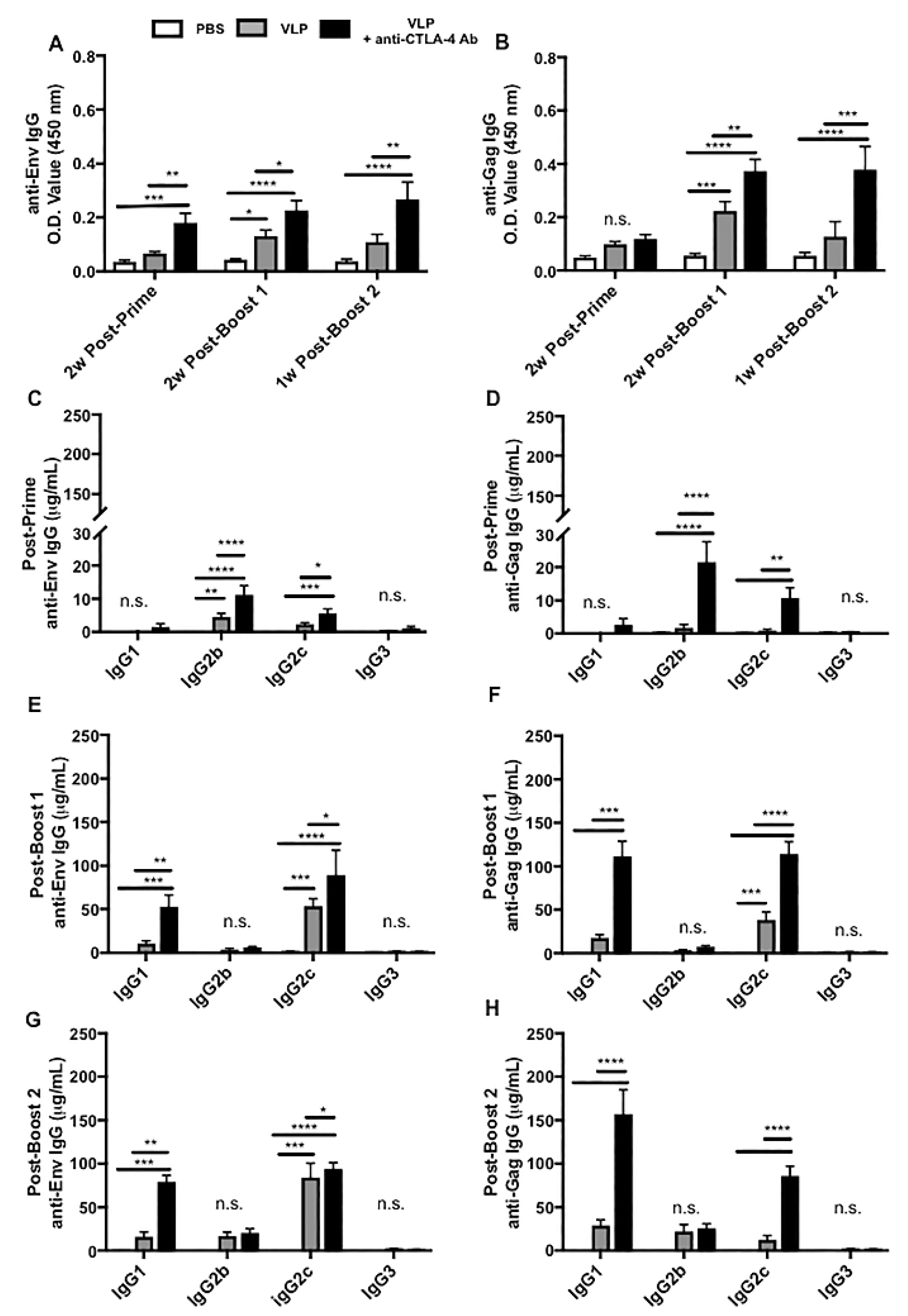
3.4. CTLA-4 Blockade Results in Increased HIV-Specific Antibody Responses, Polyclonal B-Cell Activation, and Class Switch Recombination Favoring IgG1 and IgG2c
3.5. CTLA-4 Blockade Accelerates the Induction of APRIL in the Sera
3.6. CTLA-4 Blockade during VLP Immunization Increases Serum Levels of IL-10 and IL-21
3.7. CTLA-4 Blockade Does Not Improve HIV Neutralizing Antibody Responses with VLP Immunization
3.8. CTLA-4 Blockade Induces High Avidity HIV-Specific Antibodies
3.9. CTLA-4 Blockade Increases Antibody-Dependent Cellular Cytotoxicity (ADCC) Activity
3.10. VLP Immunization, Independent of CTLA-4 Blockade, Induces Increased AID Activated Memory B-Cells and Memory T-Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Disclaimer
References
- World Health Organization. Global Health Sector Strategy on HIV 2016–2021 Towards Ending AIDS; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Pitisuttithum, P.; Gilbert, P.; Gurwith, M.; Heyward, W.; Martin, M.; van Griensven, F.; Hu, D.; Tappero, J.W.; Choopanya, K. Randomized, double-blind, placebo-controlled efficacy trial of a bivalent recombinant glycoprotein 120 HIV-1 vaccine among injection drug users in Bangkok, Thailand. J. Infect. Dis. 2006, 194, 1661–1671. [Google Scholar] [CrossRef]
- Flynn, N.M.; Forthal, D.N.; Harro, C.D.; Judson, F.N.; Mayer, K.H.; Para, M.F. Placebo-controlled phase 3 trial of a recombinant glycoprotein 120 vaccine to prevent HIV-1 infection. J. Infect. Dis. 2005, 191, 654–665. [Google Scholar] [PubMed]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kaewkungwal, J.; Chiu, J.; Paris, R.; Premsri, N.; Namwat, C.M. Vaccination with ALVAC and AIDSVAX to Prevent HIV-1 Infection in Thailand. N. Engl. J. Med. 2009, 361, 2209–2220. [Google Scholar] [CrossRef] [PubMed]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; del Rio, C. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): A double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 2008, 372, 1881–1893. [Google Scholar] [CrossRef]
- Gray, G.E.; Allen, M.; Moodie, Z.; Churchyard, G.; Bekker, L.G.; Nchabeleng, M.; Mlisana, K.; Metch, B.; de Bruyn, G.; Latka, H.; et al. Safety and efficacy of the HVTN 503/Phambili Study of a clade-B-based HIV-1 vaccine in South Africa: A double-blind, randomised, placebo-controlled test-of-concept phase 2b study. Lancet Infect. Dis. 2011, 11, 507–515. [Google Scholar] [CrossRef]
- Hammer, S.M.; Sobieszczyk, M.E.; Janes, H.; Karuna, S.T.; Mulligan, M.J.; Grove, D.; Koblin, A.; Buchbinder, S.P.; Keefer, M.C.; Tomaras, G.D.; et al. Efficacy trial of a DNA/rAd5 HIV-1 preventive vaccine. N. Engl. J. Med. 2013, 369, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- Karnasuta, C.; Akapirat, S.; Madnote, S.; Savadsuk, H.; Nicos, K. Comparison of Antibody Responses Induced by RV144, VAX003, and VAX004 Vaccination Regimens. AIDS Res. Hum. Retrovir. 2017, 33, 410–423. [Google Scholar] [CrossRef]
- Balasubramanian, P.; Williams, C.; Shapiro, M.B.; Sinangil, F.; Hioe, C.E. Functional Antibody Response Against V1V2 and V3 of HIV gp120 in the VAX003 and VAX004 Vaccine Trials. Sci. Rep. 2018, 8, 542. [Google Scholar] [CrossRef]
- Chung, A.W.; Ghebremichael, M.; Robinson, H.; Brown, E.; Choi, I.; Dugast, A.-S.; Schoen, M.K.; Rolland, M.; Suscovich, T.J. Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines. Sci. Transl. Med. 2014, 6, 228ra38. [Google Scholar] [CrossRef]
- Yates, N.L.; Liao, H.-X.; Fong, Y.; decamp, A.; Vandergrift, N.A.; Williams, W.T.; Alam, S.M.; Ferrari, G.; Yang, Y.Z.; Seaton, K.E.; et al. Vaccine-induced Env V1-V2 IgG3 correlates with lower HIV-1 infection risk and declines soon after vaccination. Sci. Transl. Med. 2014, 6, 228ra39. [Google Scholar] [CrossRef]
- Wing, J.B.; Ise, W.; Kurosaki, T.; Sakaguchi, S. Regulatory T cells control antigen-specific expansion of Tfh cell number and humoral immune responses via the coreceptor CTLA-4. Immunity 2014, 41, 1013–1025. [Google Scholar] [CrossRef]
- Sage, P.T.; Paterson, A.M.; Lovitch, S.B.; Sharpe, A.H. The coinhibitory receptor CTLA-4 controls B cell responses by modulating T follicular helper, T follicular regulatory, and T regulatory cells. Immunity 2014, 41, 1026–1039. [Google Scholar] [CrossRef] [PubMed]
- Sage, P.T.; Ron-Harel, N.; Juneja, V.R.; Sen, D.R.; Maleri, S.; Sungnak, W.; Kuchroo, V.K.; Haining, W.N.; Chevrier, N.; Haigis, M.; et al. Suppression by T FR cells leads to durable and selective inhibition of B cell effector function. Nat. Immunol. 2016, 17, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Geng, J.; Wang, Y.-H.; Wei, H.; Walters, B.; Li, W.; Luo, X.; Stevens, A.; Pittman, M.; Li, B.; et al. Foxp1 Negatively Regulates T Follicular Helper Cell Differentiation and Germinal Center Responses by Controlling Cell Migration and CTLA-4. J. Immunol. 2018, 200, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.; Heuts, F.; Ovcinnikovs, V. CTLA-4 controls follicular helper T-cell differentiation by regulating the strength of CD28 engagement. Proc. Natl. Acad. Sci. USA 2015, 112, 524–529. [Google Scholar] [CrossRef]
- Laidlaw, B.J.; Lu, Y.; Amezquita, R.A.; Weinstein, J.S.; Vander Heiden, J.A.; Gupta, N.T.; Kleinstein, S.H.; Kaech, S.M.; Craft, J. Interleukin-10 from CD4 + follicular regulatory T cells promotes the germinal center response. Sci. Immunol. 2017, 2, eaan4767. [Google Scholar] [CrossRef]
- Baumjohann, D.; Preite, S.; Reboldi, A.; Ronchi, F.; MarkAnsel, K.; Lanzavecchia, A.; Sallusto, F. Persistent Antigen and Germinal Center B Cells Sustain T Follicular Helper Cell Responses and Phenotype. Immunity 2013, 38, 596–605. [Google Scholar] [CrossRef]
- O’Connell, R.J.; Excler, J.-L. HIV vaccine efficacy and immune correlates of risk. Curr. HIV Res. 2013, 11, 450–463. [Google Scholar] [CrossRef]
- Karnasuta, C.; Paris, R.M.; Cox, J.H.; Nitayaphan, S.; Pitisuttithum, P.; Brown, A.E.; Gurunathan, S.; Tartaglia, J.; Heyward, W.L. Antibody-dependent cell-mediated cytotoxic responses in participants enrolled in a phase I/II ALVAC-HIV/AIDSVAX® B/E prime-boost HIV-1 vaccine trial in Thailand. Vaccine 2005, 23, 2522–2529. [Google Scholar] [CrossRef]
- Bonsignori, M.; Pollara, J.; Moody, M.A.; Alpert, M.D.; Chen, X.; Hwang, K.K.; Gilbert, P.B.; Huang, Y.; Gurley, T.C.; Kozink, D.M.; et al. Antibody-Dependent Cellular Cytotoxicity-Mediating Antibodies from an HIV-1 Vaccine Efficacy Trial Target Multiple Epitopes and Preferentially Use the VH1 Gene Family. J. Virol. 2012, 86, 11521–11532. [Google Scholar] [CrossRef]
- Keler, T.; Halk, E.; Vitale, L.; O’Neill, T.; Blanset, D.; Lee, S.; Srinivasan, M.; Graziano, R.F.; Davis, T.; Lonberg, N.; et al. Activity and Safety of CTLA-4 Blockade Combined with Vaccines in Cynomolgus Macaques. J. Immunol. 2003, 171, 6251–6259. [Google Scholar] [CrossRef] [PubMed]
- Sakai, J.; Akkoyunlu, M. The role of BAFF system molecules in host response to pathogens. Clin. Microbiol. Rev. 2017, 30, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Xu, S.; Lam, K.-P. Deficiency in TNFRSF13B (TACI) expands T-follicular helper and germinal center B cells via increased ICOS-ligand expression but impairs plasma cell survival. Proc. Natl. Acad. Sci. USA 2012, 109, 15401–15406. [Google Scholar] [CrossRef] [PubMed]
- Goenka, R.; Matthews, A.H.; Zhang, B.; O’Neill, P.J.; Scholz, J.L.; Migone, T.S.; Leonard, W.J.; Stohl, W.; Hershberg, U.; Cancro, M.P.; et al. Local BLyS production by T follicular cells mediates retention of high affinity B cells during affinity maturation. J. Exp. Med. 2014, 211, 45–56. [Google Scholar] [CrossRef]
- Hua, C.; Audo, R.; Yeremenko, N. A proliferation inducing ligand (APRIL) promotes IL-10 production and regulatory functions of human B cells. J. Autoimmun. 2016, 73, 64–72. [Google Scholar] [CrossRef]
- Yao, Q.; Kuhlmann, F.M.; Eller, R.O.N.; Compans, R.W.; Chen, C. Production and Characterization of Simian–Human Immunodeficiency Virus-Like Particles. AIDS Res. Hum. Retrovir. 2000, 16, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Doan, L.X.; Li, M.; Chen, C.; Yao, Q. Virus-like particles as HIV-1 vaccines. Rev. Med. Virol. 2005, 15, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Poteet, E.; Lewis, P.; Li, F.; Zhang, S.; Gu, J. A Novel Prime and Boost Regimen of HIV Virus-Like Particles with TLR4 Adjuvant MPLA Induces Th1 Oriented Immune Responses against HIV. PLoS ONE 2015, 10, e0136862. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, S.; Li, M.; Chen, C.; Yao, Q. Incorporation of CD40 ligand into SHIV virus-like particles (VLP) enhances SHIV-VLP-induced dendritic cell activation and boosts immune responses against HIV. Vaccine 2010, 28, 5114–5127. [Google Scholar] [CrossRef][Green Version]
- Zhang, R.; Li, M.; Chen, C.; Yao, Q. SHIV virus-like particles bind and activate human dendritic cells. Vaccine 2004, 23, 139–147. [Google Scholar] [CrossRef]
- Poteet, E.; Lewis, P.; Chen, C.; Ho, S.O.; Do, T.; Chiang, S.M.; Labranche, C.; Montefiori, D.; Fujii, G.; Yao, Q.; et al. Toll-like receptor 3 adjuvant in combination with virus-like particles elicit a humoral response against HIV. Vaccine 2016, 34, 5886–5894. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Zhang, R.; Guo, L.; Li, M.; Chen, C. Th Cell-Independent Immune Responses to Chimeric Hemagglutinin/Simian Human Immunodeficiency Virus-Like Particles Vaccine. J. Immunol. 2004, 173, 1951–1958. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cubas, R.; Li, M.; Chen, C.; Yao, Q. Virus-like particle vaccine activates conventional B2 cells and promotes B cell differentiation to IgG2a producing plasma cells. Mol. Immunol. 2009, 46, 1988–2001. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Doan, L.X.; Zhang, R.; Bharadwaj, U.; Li, M.; Chen, C. Thymosin-alpha1 modulates dendritic cell differentiation and functional maturation from human peripheral blood CD14+ monocytes. Immunol. Lett. 2007, 110, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Cubas, R.; Zhang, S.; Kwon, S.; Sevick-Muraca, E.M.; Li, M.; Chen, C.; Yao, Q. Virus-like particle (VLP) lymphatic trafficking and immune response generation after immunization by different routes. J. Immunother. 2009, 32, 118–128. [Google Scholar] [CrossRef]
- Hammonds, J.; Chen, X.; Zhang, X.; Lee, F.; Spearman, P. Advances in methods for the production, purification, and characterization of HIV-1 Gag-Env pseudovirion vaccines. Vaccine 2007, 25, 8036–8048. [Google Scholar] [CrossRef]
- Dogan, I.; Bertocci, B.; Vilmont, V. Multiple layers of B cell memory with different effector functions. Nat. Immunol. 2009, 10, 1292–1299. [Google Scholar] [CrossRef]
- Montefiori, D.C. HIV Protocols. In Methods in Molecular Biology, 2nd ed.; Prasad, V.R., Kalpana, G.V., Eds.; Humana Press: Totowa, NJ, USA, 2009; Volume 485, pp. 395–405. [Google Scholar]
- Levett, P.N.; Sonnenberg, K.; Sidaway, F. Use of immunoglobulin G avidity assays for differentiation of primary from previous infections with West Nile virus. J. Clin. Microbiol. 2005, 43, 5873–5875. [Google Scholar] [CrossRef]
- Shepherd, S.J.; Mcallister, G.; Kean, J. Development of an avidity assay for detection of recent HIV infections. J. Virol. Methods 2015, 217, 42–49. [Google Scholar] [CrossRef]
- Pollara, J.; Bonsignori, M.; Moody, M.A. HIV-1 Vaccine-Induced C1 and V2 Env-Specific Antibodies Synergize for Increased Antiviral Activities. J. Virol. 2014, 88, 7715–7726. [Google Scholar] [CrossRef]
- Trkola, A.; Matthews, J.; Gordon, C.; Ketas, T.; Moore, J.P. A cell line-based neutralization assay for primary human immunodeficiency virus type 1 isolates that use either the CCR5 or the CXCR4 coreceptor. J. Virol. 1999, 73, 8966–8974. [Google Scholar] [CrossRef]
- Clémenceau, B.; Vivien, R.; Pellat, C.; Foss, M.; Thibault, G.; Vié, H. The human natural killer cytotoxic cell line NK-92, once armed with a murine CD16 receptor, represents a convenient cellular tool for the screening of mouse mAbs according to their ADCC potential. MAbs 2013, 5, 587–594. [Google Scholar] [CrossRef]
- Nurieva, R.I.; Chung, Y.; Hwang, D.; Yang, X.O.; Dong, C. Generation of T Follicular Helper Cells Is Mediated by Interleukin-21 but Independent of T Helper 1, 2, or 17 Cell Lineages. Immunity 2008, 29, 138–149. [Google Scholar] [CrossRef]
- Locci, M.; Havenar-daughton, C.; Landais, E. Tfh Cells Are Highly Functional and Correlate with Broadly Neutralizing HIV Antibody Responses. Immunity 2013, 39, 1–12. [Google Scholar] [CrossRef]
- Trovato, M.; Apice, L.D. HIV Vaccination: A Roadmap among Advancements and Concerns. Int. J. Mol. Sci. 2018, 19, 1241. [Google Scholar] [CrossRef]
- Streeck, H.; D’Souza, M.P.; Littman, D.R.; Crotty, S. Harnessing CD4+ T cell responses in HIV vaccine development. Nat. Med. 2013, 19, 143–149. [Google Scholar] [CrossRef]
- Klasse, P.J. How to assess the binding strength of antibodies elicited by vaccination against HIV and other viruses. Expert Rev. Vaccines 2016, 15, 295–311. [Google Scholar] [CrossRef]
- Havenar-Daughton, C.; Lee, J.H.; Crotty, S. Tfh cells and HIV bnAbs, an immunodominance model of the HIV neutralizing antibody generation problem. Immunol. Rev. 2017, 275, 49–61. [Google Scholar] [CrossRef]
- Cirelli, K.M.; Carnathan, D.G.; Nogal, B.; Martin, J.T.; Rodriguez, O.L.; Upadhyay, A.A.; Enemuo, C.A.; Gebru, E.H.; Choe, Y.; Viviano, F.; et al. Slow Delivery Immunization Enhances HIV Neutralizing Antibody and Germinal Center Responses via Modulation of Immunodominance. Cell 2019, 177, 1153–1171. [Google Scholar] [CrossRef]
- Harper, J.; Gordon, S.; Chan, C.N.; Wang, H.; Lindemuth, E.; Galardi, C. CTLA-4 and PD-1 Dual Blockade Induces SIV Reactivation without Control of Rebound after Antiretroviral Therapy Interruption. Nat. Med. 2020, 26, 519–528. [Google Scholar] [CrossRef]
- Cecchinato, V.; Tryniszewska, E.; Ma, Z.M.; Vaccari, M.; Genoveffa, F. Immune Activation Driven by CTLA-4 Blockade Augments Viral Replication at Mucosal Sites in Simian Immunodeficiency Virus Infection. J. Immunol. 2008, 180, 5439–5447. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Tang, F.; Liu, M.; Su, J.; Zhang, Y.; Devenport, M.; Lazarski, C.A.; Zhang, P.; Wang, X. A reappraisal of CTLA-4 checkpoint blockade in cancer immunotherapy. Cell Res. 2017, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.; Gibson, H.; Aufiero, B. Differential CTLA-4 expression in human CD4+ versus CD8+ T cells is associated with increased NFAT1 and inhibition of CD4+ proliferation. Genes Immun. 2014, 15, 25–32. [Google Scholar] [CrossRef]
- Peggs, K.S.; Quezada, S.A.; Chambers, C.A.; Korman, A.J.; Allison, J.P. Blockade of CTLA-4 on both effector and regulatory T cell compartments contributes to the antitumor activity of anti–CTLA-4 antibodies. J. Exp. Med. 2009, 206, 1717–1725. [Google Scholar] [CrossRef]
- Crotty, S. T Follicular Helper Cell Differentiation, Function, and Roles in Disease. Immunity 2014, 41, 529–542. [Google Scholar] [CrossRef]
- Van Gils, M.J.; Sanders, R.W. Broadly neutralizing antibodies against HIV-1: Templates for a vaccine. Virology 2013, 435, 46–56. [Google Scholar] [CrossRef]
- Corti, D.; Lanzavecchia, A. Broadly Neutralizing Antiviral Antibodies. Annu. Rev. Immunol. 2013, 31, 705–742. [Google Scholar] [CrossRef]
- Kwong, P.D.; Mascola, J.R.; Nabel, G.J. Broadly neutralizing antibodies and the search for an HIV-1 vaccine: The end of the beginning. Nat. Rev. Immunol. 2013, 13, 693–701. [Google Scholar] [CrossRef]
- Van Gils, M.J.; Sanders, R.W. In vivo protection by broadly neutralizing HIV antibodies. Trends Microbiol. 2014, 22, 550–551. [Google Scholar] [CrossRef]
- Xu, L.; Pegu, A.; Rao, E.; Doria-Rose, N.; Beninga, J.; McKee, K.; Lord, D.M.; Wei, R.R.; Deng, G.; Louder, M.; et al. Trispecific broadly neutralizing HIV antibodies mediate potent SHIV protection in macaques. Science 2017, 358, 85–90. [Google Scholar] [CrossRef]
- Caskey, M.; Klein, F.; Nussenzweig, M.C. Broadly Neutralizing Antibodies for HIV-1 Prevention or Immunotherapy. N. Engl. J. Med. 2016, 375, 2019–2021. [Google Scholar] [CrossRef]
- Stephenson, K.E.; Barouch, D.H. Broadly Neutralizing Antibodies for HIV Eradication. Curr. HIV/AIDS Rep. 2016, 13, 31–37. [Google Scholar] [CrossRef]
- Wu, X.; Yang, Z.-Y.; Li, Y. Rational design of envelope identifies broadly neutralizing human monoclonal antibodies to HIV-1. Science 2010, 329, 856–861. [Google Scholar] [CrossRef]
- Saito, T.; Sadoshima, J. T Follicular Regulatory Cells in the Regulation of B Cell Responses. Trends Immunol. 2016, 116, 1477–1490. [Google Scholar]
- Sage, P.T.; Sharpe, A.H. T follicular regulatory cells. Immunol. Rev. 2016, 271, 246–259. [Google Scholar] [CrossRef]
- Linterman, M.A.; Denton, A.E. Treg cells and CTLA-4: The ball and chain of the germinal center response. Immunity 2014, 41, 876–878. [Google Scholar] [CrossRef][Green Version]
- Mikell, I.; Sather, D.N.; Kalams, S.A.; Altfeld, M.; Alter, G.; Stamatatos, L. Characteristics of the earliest cross-neutralizing antibody response to HIV-1. PLoS Pathog. 2011, 7, e1001251. [Google Scholar] [CrossRef]
- Pissani, F.; Streeck, H. Emerging concepts on T follicular helper cell dynamics in HIV infection. Trends Immunol. 2014, 35, 1–9. [Google Scholar] [CrossRef]
- Speiser, D.E.; Liénard, D.; Rufer, N.; Rubio-Godoy, V.; Rimoldi, D.; Lejeune, F.; Krieg, A.M.; Cerottini, J.-C.; Romero, P. Rapid and strong human CD8+T cell responses to vaccination with peptide, IFA, and CpG oligodeoxynucleotide 7909. J. Clin. Investig. 2005, 115, 739–746. [Google Scholar] [CrossRef]
- Hammonds, J.; Chen, X.; Fouts, T. Induction of neutralizing antibodies against human immunodeficiency virus type 1 primary isolates by Gag-Env pseudovirion immunization. J. Virol. 2005, 79, 14804–14814. [Google Scholar] [CrossRef]
- Huang, J.; Kang, B.H.; Ishida, E. Identification of a CD4-Binding-Site Antibody to HIV that Evolved Near-Pan Neutralization Breadth. Immunity 2016, 45, 1108–1121. [Google Scholar] [CrossRef]
- Walker, L.M.; Huber, M.; Doores, K.J. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef]
- Richard, J.; Prévost, J.; Baxter, A.E.; von Bredow, B.; Ding, S.; Medjahed, H.; Delgado, G.G.; Brassard, N.; Stürzel, C.M.; Kirchhoff, F.; et al. Uninfected Bystander Cells Impact the Measurement of HIV-. MBio 2018, 9, e00358-18. [Google Scholar] [CrossRef]
- Hraber, P.; Seaman, M.S.; Bailer, R.T.; Mascola, J.R.; Montefiori, D.C.; Korber, B.T. Prevalence of broadly neutralizing antibody responses during chronic HIV-1 infection. Aids 2014, 28, 163–169. [Google Scholar] [CrossRef]
- Yu, L.; Guan, Y. Immunologic basis for long HCDR3s in broadly neutralizing antibodies against HIV-1. Front. Immunol. 2014, 5, 28–32. [Google Scholar] [CrossRef]
- Easterhoff, D.; Moody, M.A.; Fera, D. Boosting of HIV envelope CD4 binding site antibodies with long variable heavy third complementarity determining region in the randomized double blind RV305 HIV-1 vaccine trial. PLoS Pathog. 2017, 13, e1006182. [Google Scholar] [CrossRef]
- Policicchio, B.B.; Pandrea, I.; Apetrei, C. Animal Models for HIV Cure Research. Front. Immunol. 2016, 7, 1–15. [Google Scholar] [CrossRef]
- Tager, A.M.; Pensiero, M.; Allen, T.M. Recent advances in humanized mice: Accelerating the development of an HIV vaccine. J. Infect. Dis. 2013, 208 (Suppl. 2), S121–S124. [Google Scholar] [CrossRef]
- Jardine, J.G.; Ota, T.; Sok, D.; Pauthner, M.; Kulp, D.W.; Kalyuzhniy, O.; Skog, P.D.; Thinnes, T.C.; Bhullar, D.; Briney, B.; et al. Priming a broadly neutralizing antibody response to HIV-1 using a germline-targeting immunogen. Science 2015, 349, 1–12. [Google Scholar] [CrossRef]
- Nkolola, J.P.; Bricault, C.A.; Cheung, A. Characterization and immunogenicity of a novel mosaic M HIV-1 gp140 trimer. J. Virol. 2014, 88, 9538–9552. [Google Scholar] [CrossRef]
- Li, Y.; Svehla, K.; Mathy, N.L.; Voss, G.; Mascola, J.R.; Wyatt, R. Characterization of Antibody Responses Elicited by Human Immunodeficiency Virus Type 1 Primary Isolate Trimeric and Monomeric Envelope Glycoproteins in Selected Adjuvants Characterization of Antibody Responses Elicited by Human Immunodeficiency Virus Type. J. Virol. 2006, 80, 1414–1426. [Google Scholar] [CrossRef] [PubMed]
- Bradley, T.; Kuraoka, M.; Yeh, C.H.; Tian, M.; Chen, H.; Cain, D.W. Immune checkpoint modulation enhances HIV-1 antibody induction. Nat. Commun. 2020, 11, 948. [Google Scholar] [CrossRef] [PubMed]
- Tijani, M.K.; Reddy, S.B.; Langer, C.; Beeson, J.G.; Wahlgren, M.; Nwuba, R.I.; Persson, K.E.M. Factors influencing the induction of high affinity antibodies to falciparum merozoite antigens and how affinity changes over time. Sci. Rep. 2018, 8, 9026. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.K.; Lee, J.H.; Menis, S. Precursor Frequency and Affinity Determine B Cell Competitive Fitness in Germinal Centers, Tested with Germline-Targeting HIV Vaccine Immunogens. Immunity 2017, 48, 1–14. [Google Scholar] [CrossRef]
- Shah, H.B.; Joshi, S.K.; Rampuria, P. BAFF- and APRIL-Dependent Maintenance of Antibody Titers after Immunization with T-Dependent Antigen and CD1d-Binding Ligand. J. Immunol. 2013, 191, 1154–1163. [Google Scholar] [CrossRef]
- Hryniewicz, A.; Boasso, A.; Edghill-smith, Y. CTLA-4 blockade decreases TGF-b, IDO, and viral RNA expression in tissues of SIV mac251 -infected macaques. Immunity 2006, 108, 3834–3843. [Google Scholar]
- Hirao, L.A.; Hokey, D.A.; Morrow, M.P.; Jure-Kunkel, M.N.; Weiner, D.B. Immune modulation through 4-1BB enhances SIV vaccine protection in non-human primates against SIVmac251 challenge. PLoS ONE 2011, 6, e24250. [Google Scholar] [CrossRef]
- Vaccari, M.; Boasso, A.; Fenizia, C. Fatal Pancreatitis in Simian Immunodeficiency Virus SIV mac251 -Infected Macaques Treated with 2=,3=-Dideoxyinosine and Stavudine following Cytotoxic-T-Lymphocyte-Associated Antigen 4 and Indoleamine 2,3-Dioxygenase Blockade. J. Virol. 2011, 86, 108–113. [Google Scholar] [CrossRef]










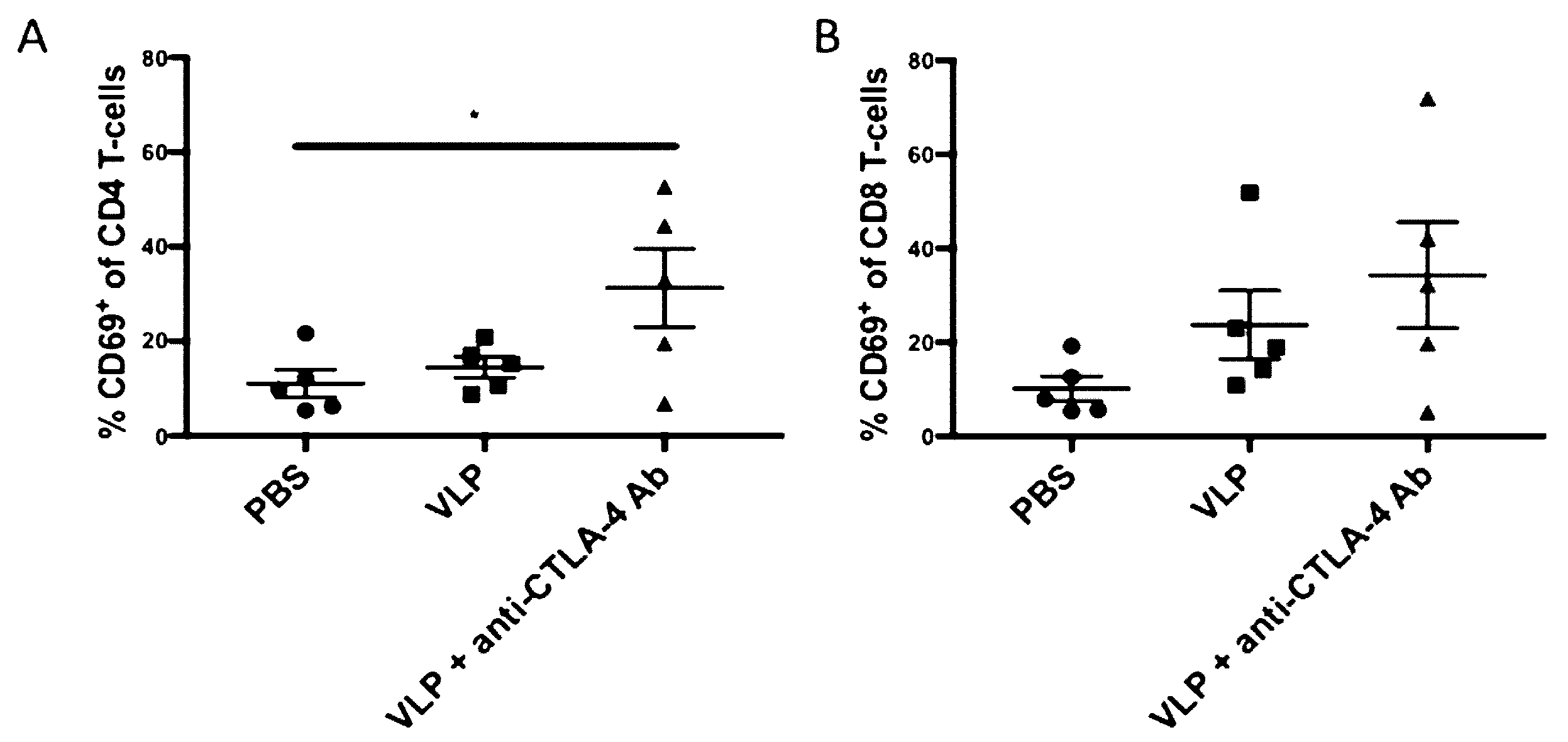{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
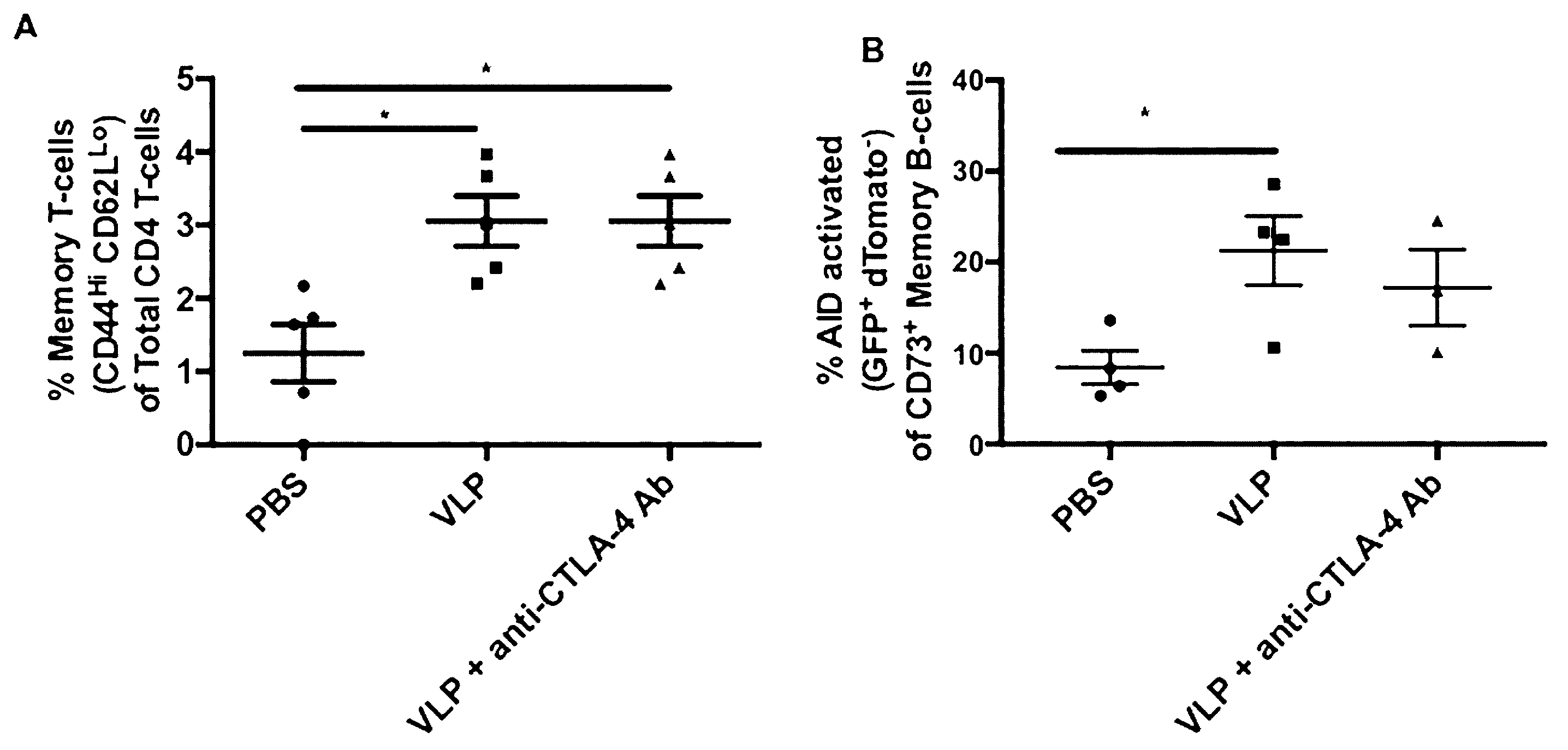{kind=link}
| Specimen ID | Specimen Collection Time | SVA-MLV Negative Control | HIV-MN.3 Clade B Tier 1a | HIV-BaL Clade B Tier 1b |
|---|---|---|---|---|
| PBS 1 | 10 days post-boost 1 | 59 | 180 | 49 |
| PBS 2 | <20 | <20 | <20 | |
| PBS 3 | 48 | 134 | 44 | |
| PBS 4 | 72 | 220 | 39 | |
| PBS 5 | 22 | 22 | 30 | |
| PBS 6 | 7 days post-boost 2 | 220 | 302 | 474 |
| PBS 7 | 25 | <20 | 37 | |
| PBS 8 | <20 | 55 | 23 | |
| PBS 9 | <20 | 25 | <20 | |
| PBS 10 | <20 | 55 | 21 | |
| VLP 1 | 10 days post-boost 1 | 22 | 185 | 57 |
| VLP 2 | <20 | 217 | 21 | |
| VLP 3 | <20 | 66 | 34 | |
| VLP 4 | <20 | 129 | 26 | |
| VLP 5 | 27 | 104 | 39 | |
| VLP 6 | 7 days post-boost 2 | <20 | 233 | <20 |
| VLP 7 | <20 | 121 | 34 | |
| VLP 8 | <20 | 198 | <20 | |
| VLP 9 | 65 | 255 | 78 | |
| VLP 10 | 24 | 137 | 44 | |
| VLP + anti-CTLA-4 Ab 1 | 10 days post-boost 1 | <20 | 55 | 25 |
| VLP + anti-CTLA-4 Ab 2 | <20 | <20 | 22 | |
| VLP + anti-CTLA-4 Ab 3 | <20 | 77 | 35 | |
| VLP + anti-CTLA-4 Ab 4 | 22 | 243 | 27 | |
| VLP + anti-CTLA-4 Ab 5 | 77 | 148 | 90 | |
| VLP + anti-CTLA-4 Ab 6 | 7 days post-boost 2 | <20 | <20 | <20 |
| VLP + anti-CTLA-4 Ab 7 | <20 | <20 | <20 | |
| VLP + anti-CTLA-4 Ab 8 | <20 | 203 | <20 | |
| VLP + anti-CTLA-4 Ab 9 | <20 | 248 | <20 | |
| VLP + anti-CTLA-4 Ab 10 | <20 | 284 | <20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lewis, P.E.; Poteet, E.C.; Liu, D.; Chen, C.; LaBranche, C.C.; Stanfield-Oakley, S.A.; Montefiori, D.C.; Ferrari, G.; Yao, Q. CTLA-4 Blockade, during HIV Virus-Like Particles Immunization, Alters HIV-Specific B-Cell Responses. Vaccines 2020, 8, 284. https://doi.org/10.3390/vaccines8020284
Lewis PE, Poteet EC, Liu D, Chen C, LaBranche CC, Stanfield-Oakley SA, Montefiori DC, Ferrari G, Yao Q. CTLA-4 Blockade, during HIV Virus-Like Particles Immunization, Alters HIV-Specific B-Cell Responses. Vaccines. 2020; 8(2):284. https://doi.org/10.3390/vaccines8020284
Chicago/Turabian StyleLewis, Phoebe E., Ethan C. Poteet, Dongliang Liu, Changyi Chen, Celia C. LaBranche, Sherry A. Stanfield-Oakley, David C. Montefiori, Guido Ferrari, and Qizhi Yao. 2020. "CTLA-4 Blockade, during HIV Virus-Like Particles Immunization, Alters HIV-Specific B-Cell Responses" Vaccines 8, no. 2: 284. https://doi.org/10.3390/vaccines8020284
APA StyleLewis, P. E., Poteet, E. C., Liu, D., Chen, C., LaBranche, C. C., Stanfield-Oakley, S. A., Montefiori, D. C., Ferrari, G., & Yao, Q. (2020). CTLA-4 Blockade, during HIV Virus-Like Particles Immunization, Alters HIV-Specific B-Cell Responses. Vaccines, 8(2), 284. https://doi.org/10.3390/vaccines8020284