Time to Micromanage the Pathogen-Host-Vector Interface: Considerations for Vaccine Development



Abstract
1. Introduction
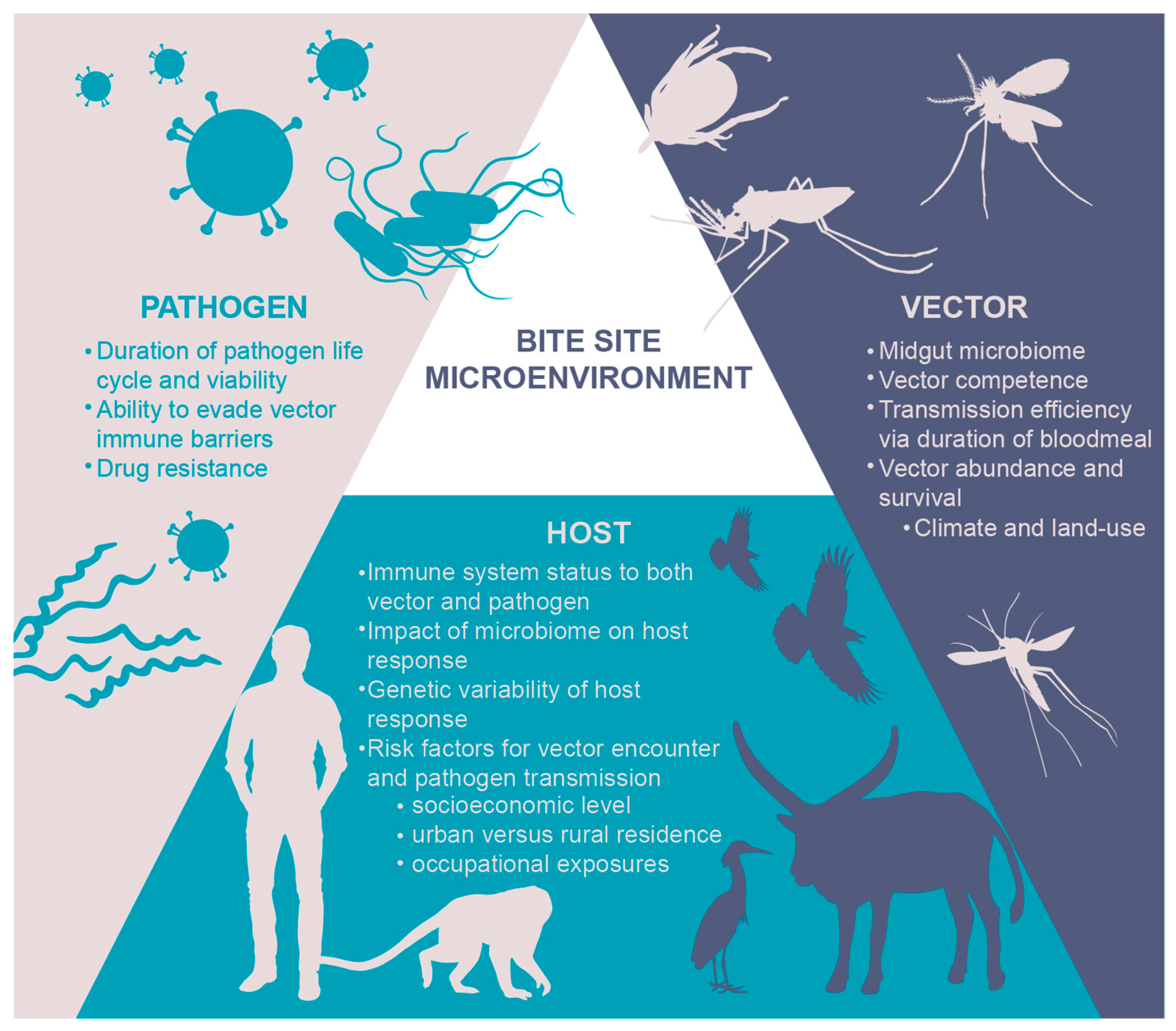
2. The Micro-Environment: Why “the Bite Site” Matters
2.1. A Skin-Deep Immunology Review
2.2. Immunogen Discovery
2.3. Lessons Learned from Other Microenvironments
3. The Microbiome: Impact of Both Host and Vector Flora on the Pathogen
4. The Micro-Needle: Rethinking Delivery
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. A Global Brief on Vector-Borne Diseases. Available online: http://www.who.int/campaigns/world-health-day/2014/global-brief/en/ (accessed on 31 March 2017).
- Rosenberg, R. Vital Signs: Trends in Reported Vectorborne Disease Cases—United States and Territories, 2004–2016. MMWR Morb. Mortal. Wkly. Rep. 2018, 67, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Mellanby, K. Man’s reaction to mosquito bites. Nature 1946, 158, 554. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.F.; Stewart, S.J.; Wikel, S.K. Resistance to Tick-Borne Francisella Tularensis by Tick-Sensitized Rabbits: Allergic Klendusity*. Am. J. Trop. Med. Hyg. 1979, 28, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Leitner, W.W.; Wali, T.; Costero-Saint Denis, A. Is Arthropod Saliva the Achilles’ Heel of Vector-Borne Diseases? Front. Immunol. 2013, 4, 255. [Google Scholar] [CrossRef] [PubMed]
- Manning, J.E.; Morens, D.M.; Kamhawi, S.; Valenzuela, J.G.; Memoli, M. Mosquito saliva: The hope for a universal arbovirus vaccine? J. Infect. Dis. 2018, 218, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.G.; Coler, R.N.; Mondal, D.; Kamhawi, S.; Valenzuela, J.G. Leishmania vaccine development: Exploiting the host-vector-parasite interface. Expert Rev. Vaccines 2016, 15, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Wikel, S.K. Tick-host-pathogen systems immunobiology: An interactive trio. Front. Biosci. Landmark Ed. 2018, 23, 265–283. [Google Scholar] [CrossRef]
- Vector-Borne Diseases. Available online: http://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 29 August 2018).
- Gotuzzo, E.; Yactayo, S.; Córdova, E. Efficacy and Duration of Immunity after Yellow Fever Vaccination: Systematic Review on the Need for a Booster Every 10 Years. Am. J. Trop. Med. Hyg. 2013, 89, 434–444. [Google Scholar] [CrossRef]
- Seven-Year Efficacy of RTS,S/AS01 Malaria Vaccine among Young African Children. NEJM. Available online: https://www.nejm.org/doi/full/10.1056/NEJMoa1515257 (accessed on 27 August 2018).
- WHO. Revised SAGE Recommendation on Use of Dengue Vaccine. Available online: http://www.who.int/immunization/diseases/dengue/revised_SAGE_recommendations_dengue_vaccines_apr2018/en/ (accessed on 27 August 2018).
- Smalley, C.; Erasmus, J.H.; Chesson, C.B.; Beasley, D.W.C. Status of research and development of vaccines for chikungunya. Vaccine 2016, 34, 2976–2981. [Google Scholar] [CrossRef]
- Osman, M.; Mistry, A.; Keding, A.; Gabe, R.; Cook, E.; Forrester, S.; Wiggins, R.; Marco, S.D.; Colloca, S.; Siani, L.; et al. A third generation vaccine for human visceral leishmaniasis and post kala azar dermal leishmaniasis: First-in-human trial of ChAd63-KH. PLoS Negl. Trop. Dis. 2017, 11, e0005527. [Google Scholar] [CrossRef]
- Nuttall, P.A. Pathogen-tick-host interactions: Borrelia burgdorferi and TBE virus. Zentralblatt Für Bakteriol. 1999, 289, 492–505. [Google Scholar] [CrossRef]
- Chmelař, J.; Kotál, J.; Kopecký, J.; Pedra, J.H.F.; Kotsyfakis, M. All For One and One For All on the Tick–Host Battlefield. Trends Parasitol. 2016, 32, 368–377. [Google Scholar] [CrossRef]
- Millington, P.F.; Wilkinson, R. Skin; Cambridge University Press: Cambridge, UK, 1983; ISBN 978-0-521-24122-9. [Google Scholar]
- Walker, J.A.; McKenzie, A.N.J. TH2 cell development and function. Nat. Rev. Immunol. 2018, 18, 121–133. [Google Scholar] [CrossRef]
- Pasparakis, M.; Haase, I.; Nestle, F.O. Mechanisms regulating skin immunity and inflammation. Nat. Rev. Immunol. 2014, 14, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Richmond, J.M.; Harris, J.E. Immunology and skin in health and disease. Cold Spring Harb. Perspect. Med. 2014, 4, a015339. [Google Scholar] [CrossRef]
- Yazdi, A.S.; Röcken, M.; Ghoreschi, K. Cutaneous immunology: Basics and new concepts. Semin. Immunopathol. 2016, 38, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, S.F.; Pitcher, L.A.; Mempel, T.; Schuerpf, F.; Carroll, M.C. B cell acquisition of antigen in vivo. Curr. Opin. Immunol. 2009, 21, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Lämmermann, T.; Sixt, M. The microanatomy of T-cell responses. Immunol. Rev. 2008, 221, 26–43. [Google Scholar] [CrossRef] [PubMed]
- Clement, C.C.; Rotzschke, O.; Santambrogio, L. The lymph as a pool of self antigens. Trends Immunol. 2011, 32, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Mellor, A.L.; Munn, D.H. Creating immune privilege: Active local suppression that benefits friends, but protects foes. Nat. Rev. Immunol. 2008, 8, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, F.; Traoré, B.; Gomes, R.; Faye, O.; Gilmore, D.C.; Keita, S.; Traoré, P.; Teixeira, C.; Coulibaly, C.A.; Samake, S.; et al. Delayed-Type Hypersensitivity to Sand Fly Saliva in Humans from a Leishmaniasis-Endemic Area of Mali Is TH1-Mediated and Persists to Midlife. J. Invest. Dermatol. 2013, 133, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Depinay, N.; Hacini, F.; Beghdadi, W.; Peronet, R.; Mécheri, S. Mast Cell-Dependent Down-Regulation of Antigen-Specific Immune Responses by Mosquito Bites. J. Immunol. 2006, 176, 4141–4146. [Google Scholar] [CrossRef]
- Scott, J.E.; ElKhal, A.; Freyschmidt, E.-J.; MacArthur, D.H.; McDonald, D.; Howell, M.D.; Leung, D.Y.M.; Laouar, A.; Manjunath, N.; Bianchi, T.; et al. Impaired immune response to vaccinia virus inoculated at the site of cutaneous allergic inflammation. J. Allergy Clin. Immunol. 2007, 120, 1382–1388. [Google Scholar] [CrossRef]
- Sakakibara, Y.; Wada, T.; Muraoka, M.; Matsuda, Y.; Toma, T.; Yachie, A. Basophil activation by mosquito extracts in patients with hypersensitivity to mosquito bites. Cancer Sci. 2015, 106, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, E.; Tritt, M.; Hay, V.; Shevach, E.M.; Belkaid, Y.; Piccirillo, C.A. CCR5-dependent homing of naturally occurring CD4+ regulatory T cells to sites of Leishmania major infection favors pathogen persistence. J. Exp. Med. 2006, 203, 2451–2460. [Google Scholar] [CrossRef]
- Peters, N.; Sacks, D. Immune privilege in sites of chronic infection: Leishmania and regulatory T cells. Immunol. Rev. 2006, 213, 159–179. [Google Scholar] [CrossRef] [PubMed]
- Guilbride, D.L.; Guilbride, P.D.L.; Gawlinski, P. Malaria’s deadly secret: A skin stage. Trends Parasitol. 2012, 28, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Seder, R.A.; Chang, L.-J.; Enama, M.E.; Zephir, K.L.; Sarwar, U.N.; Gordon, I.J.; Holman, L.A.; James, E.R.; Billingsley, P.F.; Gunasekera, A.; et al. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 2013, 341, 1359–1365. [Google Scholar] [CrossRef]
- Pombo, D.J.; Lawrence, G.; Hirunpetcharat, C.; Rzepczyk, C.; Bryden, M.; Cloonan, N.; Anderson, K.; Mahakunkijcharoen, Y.; Martin, L.B.; Wilson, D.; et al. Immunity to malaria after administration of ultra-low doses of red cells infected with Plasmodium falciparum. Lancet Lond. Engl. 2002, 360, 610–617. [Google Scholar] [CrossRef]
- Leitner, W.W.; Bergmann-Leitner, E.S.; Angov, E. Comparison of Plasmodium berghei challenge models for the evaluation of pre-erythrocytic malaria vaccines and their effect on perceived vaccine efficacy. Malar. J. 2010, 9, 145. [Google Scholar] [CrossRef]
- Styer, L.M.; Kent, K.A.; Albright, R.G.; Bennett, C.J.; Kramer, L.D.; Bernard, K.A. Mosquitoes Inoculate High Doses of West Nile Virus as They Probe and Feed on Live Hosts. PLoS Pathog. 2007, 3, e132. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Tammariello, R.F.; Spielman, A. Nonvascular delivery of St. Louis encephalitis and Venezuelan equine encephalitis viruses by infected mosquitoes (Diptera: Culicidae) feeding on a vertebrate host. J. Med. Entomol. 1995, 32, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Spielman, A. Nonvascular delivery of Rift Valley fever virus by infected mosquitoes. Am. J. Trop. Med. Hyg. 1992, 47, 190–194. [Google Scholar] [CrossRef]
- Dudley, D.M.; Newman, C.M.; Lalli, J.; Stewart, L.M.; Koenig, M.R.; Weiler, A.M.; Semler, M.R.; Barry, G.L.; Zarbock, K.R.; Mohns, M.S.; et al. Infection via mosquito bite alters Zika virus tissue tropism and replication kinetics in rhesus macaques. Nat. Commun. 2017, 8, 2096. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.J.; Watson, A.M.; Colpitts, T.M.; Dragovic, S.M.; Li, Z.; Wang, P.; Feitosa, F.; Shepherd, D.T.; Ryman, K.D.; Klimstra, W.B.; et al. Mosquito Saliva Serine Protease Enhances Dissemination of Dengue Virus into the Mammalian Host. J. Virol. 2014, 88, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Kamhawi, S.; Belkaid, Y.; Modi, G.; Rowton, E.; Sacks, D. Protection against cutaneous leishmaniasis resulting from bites of uninfected sand flies. Science 2000, 290, 1351–1354. [Google Scholar] [CrossRef]
- Oliveira, F.; Rowton, E.; Aslan, H.; Gomes, R.; Castrovinci, P.A.; Alvarenga, P.H.; Abdeladhim, M.; Teixeira, C.; Meneses, C.; Kleeman, L.T.; et al. A sand fly salivary protein vaccine shows efficacy against vector-transmitted cutaneous leishmaniasis in nonhuman primates. Sci. Transl. Med. 2015, 7, 290ra90. [Google Scholar] [CrossRef]
- Cox, J.; Mota, J.; Sukupolvi-Petty, S.; Diamond, M.S.; Rico-Hesse, R. Mosquito Bite Delivery of Dengue Virus Enhances Immunogenicity and Pathogenesis in Humanized Mice. J. Virol. 2012, 86, 7637–7649. [Google Scholar] [CrossRef]
- Briant, L.; Desprès, P.; Choumet, V.; Missé, D. Role of skin immune cells on the host susceptibility to mosquito-borne viruses. Virology 2014, 464–465, 26–32. [Google Scholar] [CrossRef]
- Surasombatpattana, P.; Patramool, S.; Luplertlop, N.; Yssel, H.; Missé, D. Aedes aegypti saliva enhances dengue virus infection of human keratinocytes by suppressing innate immune responses. J. Invest. Dermatol. 2012, 132, 2103–2105. [Google Scholar] [CrossRef]
- Schuijt, T.J.; Coumou, J.; Narasimhan, S.; Dai, J.; DePonte, K.; Wouters, D.; Brouwer, M.; Oei, A.; Roelofs, J.J.T.H.; van Dam, A.P.; et al. A tick mannose-binding lectin inhibits the vertebrate complement cascade to enhance transmission of the Lyme disease agent. Cell Host Microbe 2011, 10, 136–146. [Google Scholar] [CrossRef]
- Vogt, M.B.; Lahon, A.; Arya, R.P.; Kneubehl, A.R.; Clinton, J.L.S.; Paust, S.; Rico-Hesse, R. Mosquito saliva alone has profound effects on the human immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006439. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wang, P.; Adusumilli, S.; Booth, C.J.; Narasimhan, S.; Anguita, J.; Fikrig, E. Antibodies against a tick protein, Salp15, protect mice from the Lyme disease agent. Cell Host Microbe 2009, 6, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.A.; Glasner, D.R.; Shah, S.; Michlmayr, D.; Kramer, L.D.; Harris, E. Mosquito Saliva Increases Endothelial Permeability in the Skin, Immune Cell Migration, and Dengue Pathogenesis during Antibody-Dependent Enhancement. PLoS Pathog. 2016, 12, e1005676. [Google Scholar] [CrossRef]
- Pingen, M.; Bryden, S.R.; Pondeville, E.; Schnettler, E.; Kohl, A.; Merits, A.; Fazakerley, J.K.; Graham, G.J.; McKimmie, C.S. Host Inflammatory Response to Mosquito Bites Enhances the Severity of Arbovirus Infection. Immunity 2016, 44, 1455–1469. [Google Scholar] [CrossRef]
- Schneider, B.S.; Soong, L.; Coffey, L.L.; Stevenson, H.L.; McGee, C.E.; Higgs, S. Aedes aegypti Saliva Alters Leukocyte Recruitment and Cytokine Signaling by Antigen-Presenting Cells during West Nile Virus Infection. PLoS ONE 2010, 5, e11704. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Simons, F.E.R. Mosquito allergy: Immune mechanisms and recombinant salivary allergens. Int. Arch. Allergy Immunol. 2004, 133, 198–209. [Google Scholar] [CrossRef]
- McCracken, M.K.; Christofferson, R.C.; Grasperge, B.J.; Calvo, E.; Chisenhall, D.M.; Mores, C.N. Aedes aegypti salivary protein “aegyptin” co-inoculation modulates dengue virus infection in the vertebrate host. Virology 2014, 468–470, 133–139. [Google Scholar] [CrossRef]
- Jin, L.; Guo, X.; Shen, C.; Hao, X.; Sun, P.; Li, P.; Xu, T.; Hu, C.; Rose, O.; Zhou, H.; et al. Salivary factor LTRIN from Aedes aegypti facilitates the transmission of Zika virus by interfering with the lymphotoxin-β receptor. Nat. Immunol. 2018, 19, 342–353. [Google Scholar] [CrossRef]
- Anguita, J.; Ramamoorthi, N.; Hovius, J.W.R.; Das, S.; Thomas, V.; Persinski, R.; Conze, D.; Askenase, P.W.; Rincón, M.; Kantor, F.S.; et al. Salp15, an Ixodes scapularis Salivary Protein, Inhibits CD4+ T Cell Activation. Immunity 2002, 16, 849–859. [Google Scholar] [CrossRef]
- Chmelař, J.; Kotál, J.; Karim, S.; Kopacek, P.; Francischetti, I.M.B.; Pedra, J.H.F.; Kotsyfakis, M. Sialomes and mialomes: A systems biology view of tick tissues and tick-host interactions. Trends Parasitol. 2016, 32, 242–254. [Google Scholar] [CrossRef]
- Kazimírová, M.; Štibrániová, I. Tick salivary compounds: Their role in modulation of host defences and pathogen transmission. Front. Cell. Infect. Microbiol. 2013, 3, 43. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M.; Makoul, G.T.; Levine, J.; Robinson, D.R.; Spielman, A. Antihemostatic, antiinflammatory, and immunosuppressive properties of the saliva of a tick, Ixodes dammini. J. Exp. Med. 1985, 161, 332–344. [Google Scholar] [CrossRef]
- Gwakisa, P.; Yoshihara, K.; Long To, T.; Gotoh, H.; Amano, F.; Momotani, E. Salivary gland extract of Rhipicephalus appendiculatus ticks inhibits in vitro transcription and secretion of cytokines and production of nitric oxide by LPS-stimulated JA-4 cells. Vet. Parasitol. 2001, 99, 53–61. [Google Scholar] [CrossRef]
- Chen, G.; Severo, M.S.; Sohail, M.; Sakhon, O.S.; Wikel, S.K.; Kotsyfakis, M.; Pedra, J.H. Ixodes scapularis saliva mitigates inflammatory cytokine secretion during Anaplasma phagocytophilum stimulation of immune cells. Parasit. Vectors 2012, 5, 229. [Google Scholar] [CrossRef] [PubMed]
- Kuthejlová, M.; Kopecký, J.; Stepánová, G.; Macela, A. Tick salivary gland extract inhibits killing of Borrelia afzelii spirochetes by mouse macrophages. Infect. Immun. 2001, 69, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Kotál, J.; Langhansová, H.; Lieskovská, J.; Andersen, J.F.; Francischetti, I.M.; Chavakis, T.; Kopecký, J.; Pedra, J.H.; Kotsyfakis, M.; Chmelař, J. Modulation of host immunity by tick saliva. J. Proteomics 2015, 128, 58–68. [Google Scholar] [CrossRef]
- Wanasen, N.; Nussenzveig, R.H.; Champagne, D.E.; Soong, L.; Higgs, S. Differential modulation of murine host immune response by salivary gland extracts from the mosquitoes Aedes aegypti and Culex quinquefasciatus. Med. Vet. Entomol. 2004, 18, 191–199. [Google Scholar] [CrossRef]
- Abdeladhim, M.; Kamhawi, S.; Valenzuela, J.G. What’s behind a sand fly bite? The profound effect of sand fly saliva on host hemostasis, inflammation and immunity. Infect. Genet. Evol. 2014, 28, 691–703. [Google Scholar] [CrossRef]
- Conway, M.J.; Colpitts, T.M.; Fikrig, E. Role of the Vector in Arbovirus Transmission. Annu. Rev. Virol. 2014, 1, 71–88. [Google Scholar] [CrossRef]
- Schneider, B.S.; Soong, L.; Zeidner, N.S.; Higgs, S. Aedes aegypti Salivary Gland Extracts Modulate Anti-Viral and TH1/TH2 Cytokine Responses to Sindbis Virus Infection. Viral Immunol. 2004, 17, 565–573. [Google Scholar] [CrossRef]
- Johnston, L.J.; Halliday, G.M.; King, N.J. Langerhans cells migrate to local lymph nodes following cutaneous infection with an arbovirus. J. Invest. Dermatol. 2000, 114, 560–568. [Google Scholar] [CrossRef]
- Bizzarro, B.; Barros, M.S.; Maciel, C.; Gueroni, D.I.; Lino, C.N.; Campopiano, J.; Kotsyfakis, M.; Amarante-Mendes, G.P.; Calvo, E.; Capurro, M.L.; et al. Effects of Aedes aegypti salivary components on dendritic cell and lymphocyte biology. Parasit. Vectors 2013, 6, 329. [Google Scholar] [CrossRef] [PubMed]
- Skallová, A.; Iezzi, G.; Ampenberger, F.; Kopf, M.; Kopecký, J. Tick Saliva Inhibits Dendritic Cell Migration, Maturation, and Function while Promoting Development of Th2 Responses. J. Immunol. 2008, 180, 6186–6192. [Google Scholar] [CrossRef]
- Bournazos, S.; Ravetch, J.V. Fcγ Receptor Function and the Design of Vaccination Strategies. Immunity 2017, 47, 224–233. [Google Scholar] [CrossRef]
- Ariotti, S.; Hogenbirk, M.A.; Dijkgraaf, F.E.; Visser, L.L.; Hoekstra, M.E.; Song, J.-Y.; Jacobs, H.; Haanen, J.B.; Schumacher, T.N. Skin-resident memory CD8+ T cells trigger a state of tissue-wide pathogen alert. Science 2014, 346, 101–105. [Google Scholar] [CrossRef]
- Kumar, B.V.; Ma, W.; Miron, M.; Granot, T.; Guyer, R.S.; Carpenter, D.J.; Senda, T.; Sun, X.; Ho, S.-H.; Lerner, H.; et al. Human Tissue-Resident Memory T Cells Are Defined by Core Transcriptional and Functional Signatures in Lymphoid and Mucosal Sites. Cell Rep. 2017, 20, 2921–2934. [Google Scholar] [CrossRef] [PubMed]
- Schenkel, J.M.; Masopust, D. Tissue-Resident Memory T Cells. Immunity 2014, 41, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Zaid, A.; Hor, J.L.; Christo, S.N.; Groom, J.R.; Heath, W.R.; Mackay, L.K.; Mueller, S.N. Chemokine Receptor-Dependent Control of Skin Tissue-Resident Memory T Cell Formation. J. Immunol. Baltim. 2017, 199, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Zaid, A.; Hor, J.L.; Christo, S.N.; Prier, J.E.; Davies, B.; Alexandre, Y.O.; Gregory, J.L.; Russell, T.A.; Gebhardt, T.; et al. Local proliferation maintains a stable pool of tissue-resident memory T cells after antiviral recall responses. Nat. Immunol. 2018, 19, 183–191. [Google Scholar] [CrossRef]
- Shin, H. Formation and function of tissue-resident memory T cells during viral infection. Curr. Opin. Virol. 2018, 28, 61–67. [Google Scholar] [CrossRef]
- Ribeiro, J.M.C.; Martin-Martin, I.; Arcà, B.; Calvo, E. A Deep Insight into the Sialome of Male and Female Aedes aegypti Mosquitoes. PloS One 2016, 11, e0151400. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, J.G. High-throughput approaches to study salivary proteins and genes from vectors of disease. Insect Biochem. Mol. Biol. 2002, 32, 1199–1209. [Google Scholar] [CrossRef]
- Arcà, B.; MC Ribeiro, J. Saliva of hematophagous insects: A multifaceted toolkit. Curr. Opin. Insect Sci. 2018, 29, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Neelakanta, G.; Sultana, H. Transmission-Blocking Vaccines: Focus on Anti-Vector Vaccines against Tick-Borne Diseases. Arch. Immunol. Ther. Exp. 2015, 63, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Choumet, V.; Carmi-Leroy, A.; Laurent, C.; Lenormand, P.; Rousselle, J.-C.; Namane, A.; Roth, C.; Brey, P.T. The salivary glands and saliva of Anopheles gambiae as an essential step in the Plasmodium life cycle: A global proteomic study. Proteomics 2007, 7, 3384–3394. [Google Scholar] [CrossRef] [PubMed]
- Chisenhall, D.M.; Christofferson, R.C.; McCracken, M.K.; Johnson, A.-M.F.; Londono-Renteria, B.; Mores, C.N. Infection with dengue-2 virus alters proteins in naturally expectorated saliva of Aedes aegypti mosquitoes. Parasit. Vectors 2014, 7, 252. [Google Scholar] [CrossRef]
- Wasinpiyamongkol, L.; Patramool, S.; Luplertlop, N.; Surasombatpattana, P.; Doucoure, S.; Mouchet, F.; Séveno, M.; Remoue, F.; Demettre, E.; Brizard, J.-P.; et al. Blood-feeding and immunogenic Aedes aegypti saliva proteins. Proteomics 2010, 10, 1906–1916. [Google Scholar] [CrossRef]
- Oktarianti, R.; Senjarini, K.; Hayano, T.; Fatchiyah, F. Aulanni’am Proteomic analysis of immunogenic proteins from salivary glands of Aedes aegypti. J. Infect. Public Health 2015, 8, 575–582. [Google Scholar] [CrossRef]
- Bliss, E.; Heywood, W.E.; Benatti, M.; Sebire, N.J.; Mills, K. An optimised method for the proteomic profiling of full thickness human skin. Biol. Proced. Online 2016, 18, 15. [Google Scholar] [CrossRef]
- Surasombatpattana, P.; Ekchariyawat, P.; Hamel, R.; Patramool, S.; Thongrungkiat, S.; Denizot, M.; Delaunay, P.; Thomas, F.; Luplertlop, N.; Yssel, H.; et al. Aedes aegypti saliva contains a prominent 34-kDa protein that strongly enhances dengue virus replication in human keratinocytes. J. Invest. Dermatol. 2014, 134, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Elanga Ndille, E.; Doucoure, S.; Poinsignon, A.; Mouchet, F.; Cornelie, S.; D’Ortenzio, E.; DeHecq, J.S.; Remoue, F. Human IgG Antibody Response to Aedes Nterm-34kDa Salivary Peptide, an Epidemiological Tool to Assess Vector Control in Chikungunya and Dengue Transmission Area. PLoS Negl. Trop. Dis. 2016, 10, e0005109. [Google Scholar] [CrossRef] [PubMed]
- Calvo, E.; Tokumasu, F.; Marinotti, O.; Villeval, J.-L.; Ribeiro, J.M.C.; Francischetti, I.M.B. Aegyptin, a Novel Mosquito Salivary Gland Protein, Specifically Binds to Collagen and Prevents Its Interaction with Platelet Glycoprotein VI, Integrin α2β1, and von Willebrand Factor. J. Biol. Chem. 2007, 282, 26928–26938. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.J.; Londono-Renteria, B.; Troupin, A.; Watson, A.M.; Klimstra, W.B.; Fikrig, E.; Colpitts, T.M. Aedes aegypti D7 Saliva Protein Inhibits Dengue Virus Infection. PLoS Negl. Trop. Dis. 2016, 10, e0004941. [Google Scholar] [CrossRef] [PubMed]
- Schleicher, T.R.; Yang, J.; Freudzon, M.; Rembisz, A.; Craft, S.; Hamilton, M.; Graham, M.; Mlambo, G.; Tripathi, A.K.; Li, Y.; et al. A mosquito salivary gland protein partially inhibits Plasmodium sporozoite cell traversal and transmission. Nat. Commun. 2018, 9, 2908. [Google Scholar] [CrossRef]
- Tomás-Cortázar, J.; Martín-Ruiz, I.; Barriales, D.; Pascual-Itoiz, M.Á.; de Juan, V.G.; Caro-Maldonado, A.; Merino, N.; Marina, A.; Blanco, F.J.; Flores, J.M.; et al. The immunosuppressive effect of the tick protein, Salp15, is long-lasting and persists in a murine model of hematopoietic transplant. Sci. Rep. 2017, 7, 10740. [Google Scholar] [CrossRef]
- Hackenberg, M.; Langenberger, D.; Schwarz, A.; Erhart, J.; Kotsyfakis, M. In silico target network analysis of de novo-discovered, tick saliva-specific microRNAs reveals important combinatorial effects in their interference with vertebrate host physiology. RNA 2017, 23, 1259–1269. [Google Scholar] [CrossRef]
- Maharaj, P.D.; Widen, S.G.; Huang, J.; Wood, T.G.; Thangamani, S. Discovery of Mosquito Saliva MicroRNAs during CHIKV Infection. PLoS Negl. Trop. Dis. 2015, 9, e0003386. [Google Scholar] [CrossRef]
- Drury, R.E.; O’Connor, D.; Pollard, A.J. The Clinical Application of MicroRNAs in Infectious Disease. Front. Immunol. 2017, 8, 1182. [Google Scholar] [CrossRef]
- Heiss, B.L.; Maximova, O.A.; Thach, D.C.; Speicher, J.M.; Pletnev, A.G. MicroRNA Targeting of Neurotropic Flavivirus: Effective Control of Virus Escape and Reversion to Neurovirulent Phenotype. J. Virol. 2012, 86, 5647–5659. [Google Scholar] [CrossRef]
- Woodland, D.L.; Kohlmeier, J.E. Migration, maintenance and recall of memory T cells in peripheral tissues. Nat. Rev. Immunol. 2009, 9, 153–161. [Google Scholar] [CrossRef]
- Gaide, O.; Emerson, R.O.; Jiang, X.; Gulati, N.; Nizza, S.; Desmarais, C.; Robins, H.; Krueger, J.G.; Clark, R.A.; Kupper, T.S. Common clonal origin of central and resident memory T cells following skin immunization. Nat. Med. 2015, 21, 647–653. [Google Scholar] [CrossRef]
- Jiang, X.; Clark, R.A.; Liu, L.; Wagers, A.J.; Fuhlbrigge, R.C.; Kupper, T.S. Skin infection generates non-migratory memory CD8+ TRM cells providing global skin immunity. Nature 2012, 483, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Muruganandah, V.; Sathkumara, H.D.; Navarro, S.; Kupz, A. A Systematic Review: The Role of Resident Memory T Cells in Infectious Diseases and Their Relevance for Vaccine Development. Front. Immunol. 2018, 9, 1574. [Google Scholar] [CrossRef] [PubMed]
- Laidlaw, B.J.; Zhang, N.; Marshall, H.D.; Staron, M.M.; Guan, T.; Hu, Y.; Cauley, L.S.; Craft, J.; Kaech, S.M. CD4+ T cell help guides formation of CD103+ lung-resident memory CD8+ T cells during influenza viral infection. Immunity 2014, 41, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Sckisel, G.D.; Tietze, J.K.; Zamora, A.E.; Hsiao, H.-H.; Priest, S.O.; Wilkins, D.E.C.; Lanier, L.L.; Blazar, B.R.; Baumgarth, N.; Murphy, W.J. Influenza infection results in local expansion of memory CD8+ T cells with antigen non-specific phenotype and function. Clin. Exp. Immunol. 2014, 175, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Blanc, C.; Hans, S.; Tran, T.; Granier, C.; Saldman, A.; Anson, M.; Oudard, S.; Tartour, E. Targeting Resident Memory T Cells for Cancer Immunotherapy. Front. Immunol. 2018, 9, 1722. [Google Scholar] [CrossRef] [PubMed]
- Hopp, C.S.; Sinnis, P. The innate and adaptive response to mosquito saliva and Plasmodium sporozoites in the skin. Ann. NY Acad. Sci. 2015, 1342, 37–43. [Google Scholar] [CrossRef] [PubMed]
- McMaster, S.R.; Wilson, J.J.; Wang, H.; Kohlmeier, J.E. Airway-Resident Memory CD8 T Cells Provide Antigen-Specific Protection against Respiratory Virus Challenge through Rapid IFN-γ Production. J. Immunol. 2015, 195, 203–209. [Google Scholar] [CrossRef]
- Park, C.; Kupper, T.S. The emerging role of resident memory T cells in protective immunity and inflammatory disease. Nat. Med. 2015, 21, 688–697. [Google Scholar] [CrossRef]
- Malik, B.T.; Byrne, K.T.; Vella, J.L.; Zhang, P.; Shabaneh, T.B.; Steinberg, S.M.; Molodtsov, A.K.; Bowers, J.S.; Angeles, C.V.; Paulos, C.M.; et al. Resident memory T cells in the skin mediate durable immunity to melanoma. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Enamorado, M.; Iborra, S.; Priego, E.; Cueto, F.J.; Quintana, J.A.; Martínez-Cano, S.; Mejías-Pérez, E.; Esteban, M.; Melero, I.; Hidalgo, A.; et al. Enhanced anti-tumour immunity requires the interplay between resident and circulating memory CD8+ T cells. Nat. Commun. 2017, 8, 16073. [Google Scholar] [CrossRef]
- Quezada, S.A.; Simpson, T.R.; Peggs, K.S.; Merghoub, T.; Vider, J.; Fan, X.; Blasberg, R.; Yagita, H.; Muranski, P.; Antony, P.A.; et al. Tumor-reactive CD4+ T cells develop cytotoxic activity and eradicate large established melanoma after transfer into lymphopenic hosts. J. Exp. Med. 2010, 207, 637–650. [Google Scholar] [CrossRef]
- Mitchell, R.A.; Altszuler, R.; Frevert, U.; Nardin, E.H. Skin scarification with Plasmodium falciparum peptide vaccine using synthetic TLR agonists as adjuvants elicits malaria sporozoite neutralizing immunity. Sci. Rep. 2016, 6, 32575. [Google Scholar] [CrossRef]
- Rice, A.D.; Adams, M.M.; Lindsey, S.F.; Swetnam, D.M.; Manning, B.R.; Smith, A.J.; Burrage, A.M.; Wallace, G.; MacNeill, A.L.; Moyer, R.W. Protective properties of vaccinia virus-based vaccines: Skin scarification promotes a nonspecific immune response that protects against orthopoxvirus disease. J. Virol. 2014, 88, 7753–7763. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhong, Q.; Tian, T.; Dubin, K.; Athale, S.K.; Kupper, T.S. Epidermal injury and infection during poxvirus immunization is crucial for the generation of highly protective T cell-mediated immunity. Nat. Med. 2010, 16, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Labuda, M.; Kozuch, O. Amplification of arbovirus transmission by mosquito intradermal probing and interrupted feeding. Acta Virol. 1989, 33, 63–67. [Google Scholar] [PubMed]
- Schiffer, J.T.; Abu-Raddad, L.; Mark, K.E.; Zhu, J.; Selke, S.; Koelle, D.M.; Wald, A.; Corey, L. Mucosal host immune response predicts the severity and duration of herpes simplex virus-2 genital tract shedding episodes. Proc. Natl. Acad. Sci. 2010, 107, 18973–18978. [Google Scholar] [CrossRef] [PubMed]
- Gebhardt, T.; Whitney, P.G.; Zaid, A.; Mackay, L.K.; Brooks, A.G.; Heath, W.R.; Carbone, F.R.; Mueller, S.N. Different patterns of peripheral migration by memory CD4+ and CD8+ T cells. Nature 2011, 477, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.M.; Lebratti, T.J.; Richner, J.M.; Jiang, X.; Fernandez, E.; Zhao, H.; Fremont, D.H.; Diamond, M.S.; Shin, H. Cellular and Humoral Immunity Protect against Vaginal Zika Virus Infection in Mice. J. Virol. 2018. [Google Scholar] [CrossRef]
- Davies, B.; Prier, J.E.; Jones, C.M.; Gebhardt, T.; Carbone, F.R.; Mackay, L.K. Cutting Edge: Tissue-Resident Memory T Cells Generated by Multiple Immunizations or Localized Deposition Provide Enhanced Immunity. J. Immunol. 2017, 198, 2233–2237. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D.S. Battling the Bite: Tradeoffs in Immunity to Insect-Borne Pathogens. Immunity 2016, 44, 1251–1252. [Google Scholar] [CrossRef] [PubMed]
- Donovan, M.J.; Messmore, A.S.; Scrafford, D.A.; Sacks, D.L.; Kamhawi, S.; McDowell, M.A. Uninfected Mosquito Bites Confer Protection against Infection with Malaria Parasites. Infect. Immun. 2007, 75, 2523–2530. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, B.; Targett, G. The mysteries of immunity to malaria. The Lancet 2011, 377, 1729–1730. [Google Scholar] [CrossRef]
- Trager, W. Acquired Immunity to Ticks. J. Parasitol. 1939, 25, 57–81. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. The influence of the intestinal microbiome on vaccine responses. Vaccine 2018, 36, 4433–4439. [Google Scholar] [CrossRef]
- Da Fonseca, D.M.; Hand, T.W.; Han, S.-J.; Gerner, M.Y.; Glatman Zaretsky, A.; Byrd, A.L.; Harrison, O.J.; Ortiz, A.M.; Quinones, M.; Trinchieri, G.; et al. Microbiota-Dependent Sequelae of Acute Infection Compromise Tissue-Specific Immunity. Cell 2015, 163, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Rosshart, S.P.; Vassallo, B.G.; Angeletti, D.; Hutchinson, D.S.; Morgan, A.P.; Takeda, K.; Hickman, H.D.; McCulloch, J.A.; Badger, J.H.; Ajami, N.J.; et al. Wild Mouse Gut Microbiota Promotes Host Fitness and Improves Disease Resistance. Cell 2017, 171, 1015–1028.e13. [Google Scholar] [CrossRef] [PubMed]
- Grassly, N.C.; Fraser, C.; Wenger, J.; Deshpande, J.M.; Sutter, R.W.; Heymann, D.L.; Aylward, R.B. New strategies for the elimination of polio from India. Science 2006, 314, 1150–1153. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.M.; Kaper, J.B.; Herrington, D.; Ketley, J.; Losonsky, G.; Tacket, C.O.; Tall, B.; Cryz, S. Safety, immunogenicity, and efficacy of recombinant live oral cholera vaccines, CVD 103 and CVD 103-HgR. Lancet Lond. Engl. 1988, 2, 467–470. [Google Scholar] [CrossRef]
- Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.; et al. Compartmentalized Control of Skin Immunity by Resident Commensals. Science 2012, 337, 1115–1119. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Piccirillo, C.A.; Mendez, S.; Shevach, E.M.; Sacks, D.L. CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 2002, 420, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Ives, A.; Ronet, C.; Prevel, F.; Ruzzante, G.; Fuertes-Marraco, S.; Schutz, F.; Zangger, H.; Revaz-Breton, M.; Lye, L.-F.; Hickerson, S.M.; et al. Leishmania RNA virus controls the severity of mucocutaneous leishmaniasis. Science 2011, 331, 775–778. [Google Scholar] [CrossRef]
- Mueller, S.N.; Matloubian, M.; Clemens, D.M.; Sharpe, A.H.; Freeman, G.J.; Gangappa, S.; Larsen, C.P.; Ahmed, R. Viral targeting of fibroblastic reticular cells contributes to immunosuppression and persistence during chronic infection. Proc. Natl. Acad. Sci. 2007, 104, 15430–15435. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota-host interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.N.; Rocha, M.N.; Sucupira, P.H.F.; Carvalho, F.D.; Moreira, L.A. Wolbachia significantly impacts the vector competence of Aedes aegypti for Mayaro virus. Sci. Rep. 2018, 8, 6889. [Google Scholar] [CrossRef] [PubMed]
- Dey, R.; Joshi, A.B.; Oliveira, F.; Pereira, L.; Guimarães-Costa, A.B.; Serafim, T.D.; de Castro, W.; Coutinho-Abreu, I.V.; Bhattacharya, P.; Townsend, S.; et al. Gut Microbes Egested during Bites of Infected Sand Flies Augment Severity of Leishmaniasis via Inflammasome-Derived IL-1β. Cell Host Microbe 2018, 23, 134–143.e6. [Google Scholar] [CrossRef]
- Gendrin, M.; Rodgers, F.H.; Yerbanga, R.S.; Ouédraogo, J.B.; Basáñez, M.-G.; Cohuet, A.; Christophides, G.K. Antibiotics in ingested human blood affect the mosquito microbiota and capacity to transmit malaria. Nat. Commun. 2015, 6, 5921. [Google Scholar] [CrossRef]
- Zheng, Z.; Diaz-Arévalo, D.; Guan, H.; Zeng, M. Noninvasive vaccination against infectious diseases. Hum. Vaccines Immunother. 2018, 14, 1717–1733. [Google Scholar] [CrossRef]
- Marshall, S.; Sahm, L.J.; Moore, A.C. The success of microneedle-mediated vaccine delivery into skin. Hum. Vaccines Immunother. 2016, 12, 2975–2983. [Google Scholar] [CrossRef]
- Gurera, D.; Bhushan, B.; Kumar, N. Lessons from mosquitoes’ painless piercing. J. Mech. Behav. Biomed. Mater. 2018, 84, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Norman, J.J.; Arya, J.M.; McClain, M.A.; Frew, P.M.; Meltzer, M.I.; Prausnitz, M.R. Microneedle patches: Usability and acceptability for self-vaccination against influenza. Vaccine 2014, 32, 1856–1862. [Google Scholar] [CrossRef] [PubMed]
- Carey, J.B.; Pearson, F.E.; Vrdoljak, A.; McGrath, M.G.; Crean, A.M.; Walsh, P.T.; Doody, T.; O’Mahony, C.; Hill, A.V.S.; Moore, A.C. Microneedle Array Design Determines the Induction of Protective Memory CD8+ T Cell Responses Induced by a Recombinant Live Malaria Vaccine in Mice. PLoS ONE 2011, 6, e22442. [Google Scholar] [CrossRef] [PubMed]
- Prow, T.W.; Chen, X.; Prow, N.A.; Fernando, G.J.P.; Tan, C.S.E.; Raphael, A.P.; Chang, D.; Ruutu, M.P.; Jenkins, D.W.K.; Pyke, A.; et al. Nanopatch-Targeted Skin Vaccination against West Nile Virus and Chikungunya Virus in Mice. Small 2010, 6, 1776–1784. [Google Scholar] [CrossRef]
- Moreno, E.; Schwartz, J.; Calvo, A.; Blanco, L.; Larrea, E.; Irache, J.M.; Sanmartín, C.; Coulman, S.A.; Soto, M.; Birchall, J.C.; et al. Skin vaccination using microneedles coated with a plasmid DNA cocktail encoding nucleosomal histones of Leishmania spp. Int. J. Pharm. 2017, 533, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Van De Wall, S.; Walczak, M.; Van Rooij, N.; Hoogeboom, B.-N.; Meijerhof, T.; Nijman, H.W.; Daemen, T. Tattoo Delivery of a Semliki Forest Virus-Based Vaccine Encoding Human Papillomavirus E6 and E7. Vaccines 2015, 3, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, N.G.; Paine, M.; Mosley, R.; Henry, S.; McAllister, D.V.; Kalluri, H.; Pewin, W.; Frew, P.M.; Yu, T.; Thornburg, N.J.; et al. The safety, immunogenicity, and acceptability of inactivated influenza vaccine delivered by microneedle patch (TIV-MNP 2015): A randomised, partly blinded, placebo-controlled, phase 1 trial. Lancet Lond. Engl. 2017, 390, 649–658. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Pathogen | Primary Vector(s) | Vaccine Name | Platform | Immunogen | Adjuvant | Status 2 | Sponsor |
|---|---|---|---|---|---|---|---|
| Dengue virus 2,3 serotypes 1–4 | Aedes aegypti mosquito, Aedes albopictus mosquito | CYD-TDV | Recombinant viral vector (YFV backbone) | PrM+E of DENV1-4 | None | Registered by WHO in select populations * | Sanofi-Pasteur |
| TDV | Recombinant viral vector | PrM+E | None | Phase 3 | Takeda | ||
| TV003/TV005 | Recombinant viral vector (DENV backbone) | Whole virus/PrM+E | None | Phase 3 | NIAID | ||
| TDENV-PIV | Inactivated whole target organism | Whole virus | Alum or AS03 | Phase 2 | USAMRMC | ||
| V180 | Subunit | PrM+E | Alhydrogel | Phase 1 | Merck | ||
| Zika virus 2 | Aedes aegypti mosquito | GLS-5700 | DNA | PrM+E | None | Phase 1 | GeneOne Life Science/Inovio |
| MV-Zika | Recombinant viral vector | PrM+E | None | Phase 1 | Themis Bioscience | ||
| AGS-v | Synthetic peptide | Mosquito saliva peptide | IFA-51 | Phase 1 | NIAID | ||
| mRNA-1325 | mRNA | PrM+E | None | Phase 2 | Moderna Therapeutics | ||
| VRC-ZKADNA085-00-VP | DNA | PrM+E | None | Phase 1 | NIAID | ||
| VRC-ZKADNA090-00-VP | DNA | PrM+E | None | Phase 2 | NIAID | ||
| ZIKA PIV | Inactivated whole target organism | Whole virus | Alum | Phase 1 | NIAID | ||
| PIZV or TAK-426 | Inactivated whole target organism | Whole virus | Alum | Phase 1 | Takeda | ||
| VLA1601 | Inactivated whole target organism | Whole virus | Alum | Phase 1 | Valneva Austria GmbH | ||
| Chikungunya virus 2 | Aedes aegypti mosquito, Aedes albopictus mosquito | PXVX0317 CHIKV-VLP | Virus-like particle | E1, E2 and capsid proteins | With and without Alhydrogel | Phase 2 | NIAID now transferred to PaxVax |
| MV-CHIK | Recombinant viral vector | Phase 2 | Themis Bioscience | ||||
| VAL 181388 | mRNA | N.A. | N.A. | Phase 1 | Moderna Therapeutics | ||
| CHIK001 or ChAdOx1 | Recombinant viral vector | Phase 1 | University of Oxford | ||||
| VLA1533 | Live, attenuated virus | Whole virus | None | Phase 1 | Valneva SE | ||
| BBV87 | Inactivated whole target organism | Phase 1 | Bharath Biotech | ||||
| CHIKV 181/25 | Inactivated whole target organism | Phase 1 | USAMRMC transferred to Indian Immuno-logicals | ||||
| Yellow Fever virus | Aedes aegypti mosquito | Yellow Fever Vaccines (YFV) sold as YF-VAX in USA, STAMARIL elsewhere | Live, attenuated virus of 17D lineage | Licensed worldwide | Sanofi-Pastuer | ||
| West Nile Virus | Culex spp. mosquito | WN/DEN4Δ30 | Recombinant viral vector | Whole live, attenuated virus | Phase 1 | Johns Hopkins University | |
| HydroVax-001 | Inactivated WNV | Alum | Phase 1 | NIAID | |||
| VRC-WNVDNA020-00-VP | DNA | PrM and E proteins of NY99 strain with CMV/R promoter | Phase 1 | NIAID | |||
| Japanese Encephalitis virus | Culex spp. mosquito | CD.JEVAX® | Primary hamster kidney cell-derived, live, attenuated vaccine based on SA 14-14-2 strain | PrM+E | Licensed in China since 1988 as JEVAX | Chengdu Institute | |
| IMOJEV®, JE-CV®, ChimeriVax-JE® | Live, attenuated YFV with SA 14-14-2 live attenuated JEV produced in Vero cells | PrM+E | Licensed as early as 2010 in Australia and other Asian countries | Sanofi Pasteur | |||
| Ixiaro®, JESPECT®, JEEV® | Inactivated Vero cell-derived | Whole virus | Alum | Licensed in USA and Europe since 2009 | Valneva Austria GmbH | ||
| Equine Encephalitis Viruses (Eastern, Western, and Venezuelan) | Culiseta melanura mosquito (but human bridge vectors are likely Aedes, Culex, or Coquillettidia spp. mosquitos) | TSI-GSD 210, Lot 3-1-92 | Inactivated WEE | Whole virus | Phase 2 | USAMRMC | |
| TSI-GSD 104, Lot 2-1-89 | Inactivated EEE | Whole virus | |||||
| C-84, TSI-GSD 205, Lot 7 | Inactivated VEE | Whole virus | |||||
| TC-83, NDBR-102 | Live, attenuated VEE | ||||||
| pWRG/VEE | DNA | pWRG/VEE | Phase 1 | Ichor Medical Systems | |||
| Plasmodium falciparum2 | Anopheles spp. mosquito | RTS,S/ASO4 (Mosquirix®) | Recombinant subunit | CSP | AS01 | Approved by EMA for children 6–17 months of age | Glaxo-SmithKline Inc. |
| ChAd63/MVA ME-TRAP | Recombinant subunit | TRAP + ME epitopes (CS, LSA1, LSA3, STARP, EXP1, pb9) | None | Phase 2B | University of Oxford | ||
| ChAd63/-METRAP | Recombinant subunit | ME+TRAP | None | Phase 1 | University of Oxford | ||
| ChAd63 RH5 +/− MVA RH5 | Recombinant subunit | RH5 | None | Phase 1 | University of Oxford | ||
| PfsSPZ | Inactivated whole organism | Whole sporozoite | None | Phase 2 | Sanaria | ||
| PfCelTOS FMP012 | Recombinant subunit | CelTOS protein | AS01B or GLA-SE | Phase 1 | USAMRMC | ||
| Pfs25-EPA+Pfs230-EPA | Pfs25M or Pf230D1M conjugated to EPA, respectively | Pfs25M, Pfs230D1M | AS01 or Alhydrogel | Phase 1 | NIAID | ||
| R21 | Recombinant subunit | CSP less-HepBsA | AS01B or Matrix-M1 | Phase 1/2 | University of Oxford | ||
| GMZ2 | Recombinant subunit | GLURP, MSP3 | Aluminium hydroxide, GLA-SE | Phase 2b | Statens Serum Institute | ||
| PRIMVAC (placental malaria) | Recombinant protein | VAR2CSA fragment | Alhydrogel or GLA-SE | Phase 1 | INSERM | ||
| SE36 | Recombinant subunit | N-terminal SERA5 | Alhydrogel | Phase 1 | NobelPharma Co Ltd, Japan | ||
| Plasmodium vivax | Anopheles spp. mosquito | ChAd63/MVA PvDBP | Recombinant viral vector | PvDBP_RII | Phase 1 | University of Oxford | |
| Borrelia burgdorferi | Ixodes scapularis, blacklegged or deer tick | Multivalent OspA Lyme Borreliosis vaccine | Recombinant peptide | 6 antigens of rOspA | Alum | Phase 1/2 | Baxalta (Shire) |
| VLA15 | Recombinant peptide | Multivalent OspA | Alum | Phase 1 | Valneva Austria GmbH | ||
| Tick-borne Encephalitis | Hard ticks of Ixodidae family | FSME-Immun (Junior) | Neudorfl strain of European subtype | Aluminum hydroxide | Licensed in Europe in 1976 | ||
| Encepur-Adults (-Children) | K23 virus strain | Aluminum hydroxide | Licensed in Europe in 1994 | ||||
| TBE-Moscow | Sofjin strain of Far-Eastern viral subtype | Aluminum hydroxide | Licensed in Russia in 1982 (and in 1999 for children >3 years) | ||||
| EnceVir | Far-Eastern strain 205 | Aluminum hydroxide | Licensed in Russia |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manning, J.E.; Cantaert, T. Time to Micromanage the Pathogen-Host-Vector Interface: Considerations for Vaccine Development. Vaccines 2019, 7, 10. https://doi.org/10.3390/vaccines7010010
Manning JE, Cantaert T. Time to Micromanage the Pathogen-Host-Vector Interface: Considerations for Vaccine Development. Vaccines. 2019; 7(1):10. https://doi.org/10.3390/vaccines7010010
Chicago/Turabian StyleManning, Jessica E., and Tineke Cantaert. 2019. "Time to Micromanage the Pathogen-Host-Vector Interface: Considerations for Vaccine Development" Vaccines 7, no. 1: 10. https://doi.org/10.3390/vaccines7010010
APA StyleManning, J. E., & Cantaert, T. (2019). Time to Micromanage the Pathogen-Host-Vector Interface: Considerations for Vaccine Development. Vaccines, 7(1), 10. https://doi.org/10.3390/vaccines7010010