
Live-Attenuated Bacterial Vectors: Tools for Vaccine and Therapeutic Agent Delivery

Abstract
:1. Introduction
2. Live-Attenuated Bacterial Vaccine Vectors for Infectious Diseases
2.1. Attenuation of Bacterial Vectors
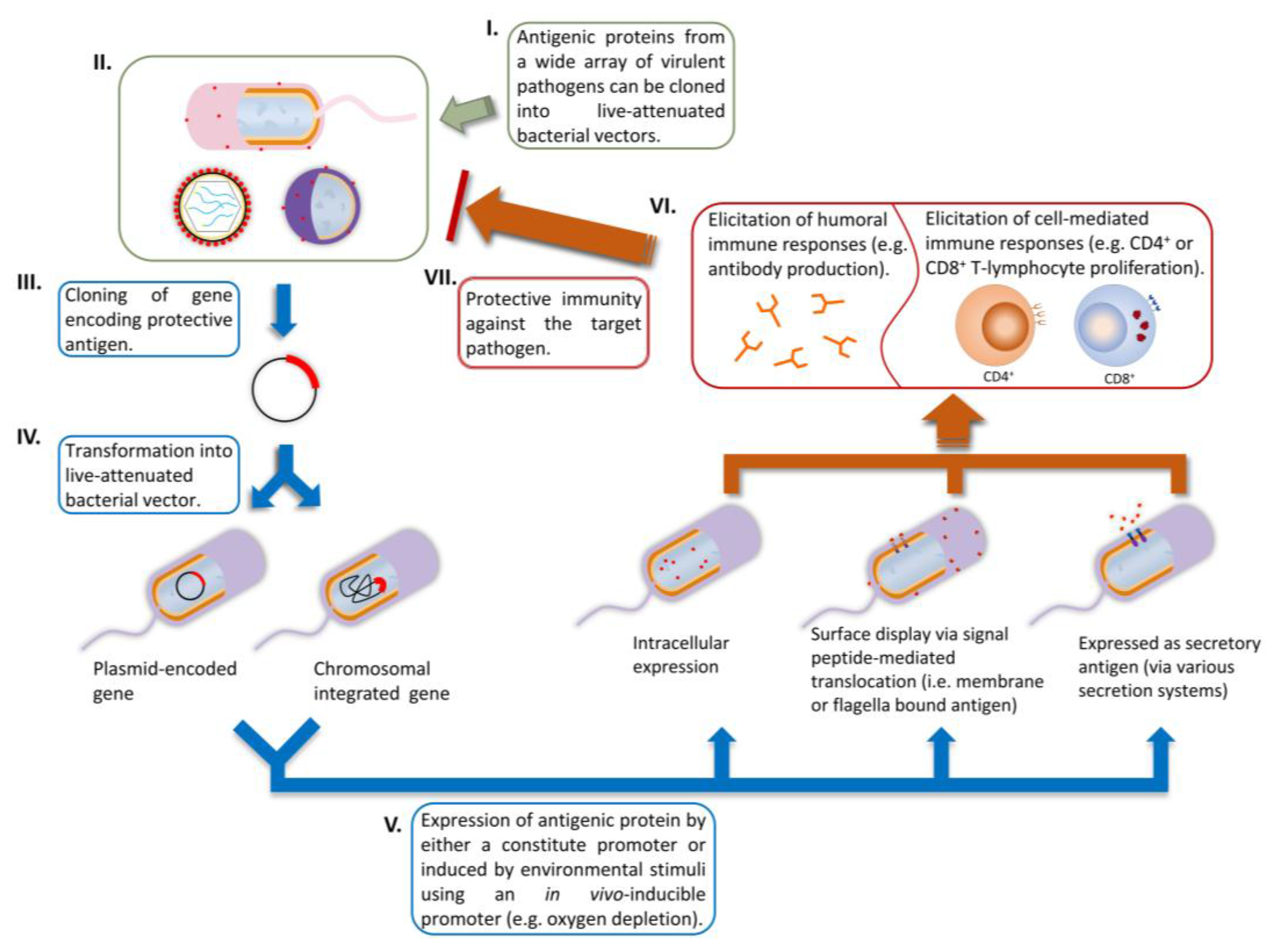
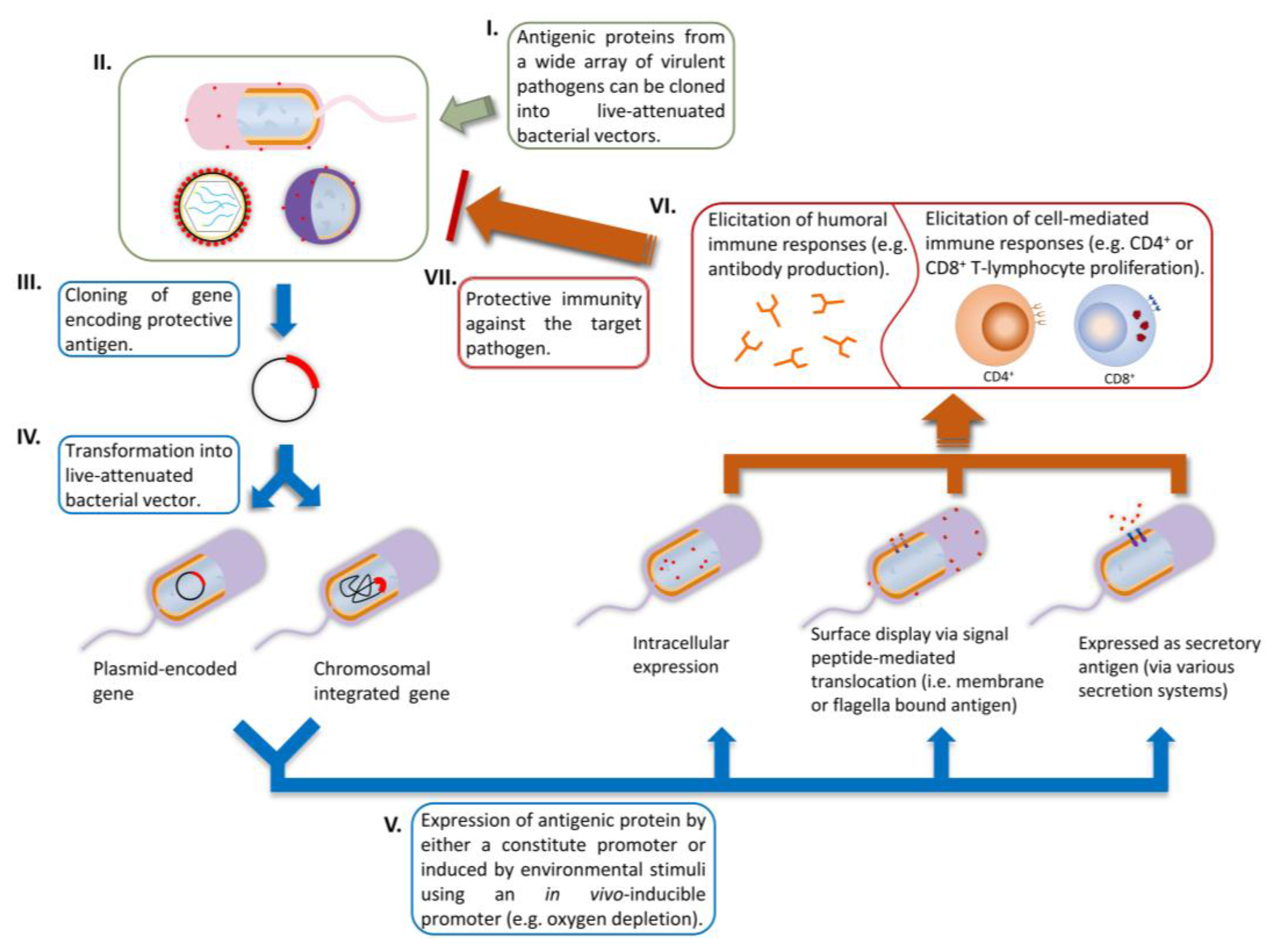
2.2. Genetic Stability and Protein Expression of Heterologous Genes
2.3. Controlling Antigen Expression and Antigen Compartmentalization
2.4. Current Achievements and Perspectives

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vector | Target Host Cell | Advantages | Limitations/Concerns | Ref. |
|---|---|---|---|---|
| Enteric pathogens | ||||
| Listeria monocytogenes | Intestinal epithelial cells and non-phagocytic cells such as hepatocytes are primary invasion sites before systemic dissemination. Intracellular pathogen. | Ability to present homologous and heterologous antigens to both endogenous and exogenous antigen-presenting pathways, thus eliciting CD4+ and CD8+ T-lymphocyte responses. | Wild-type Listeria can cause serious and potentially lethal disease, especially in immunocompromised individuals. Severe attenuation to ensure safety could over-attenuate and lead to poor immunogenicity. Possible reversion to pathogenic state. | [64] |
| Salmonella spp. | M cells as primary invasion site, and taken up by phagocytic cells for systemic dissemination. Intracellular pathogen. | Among the first bacteria used as vaccine-carrier to deliver heterologous antigens, well-established protocol for genetic manipulation. Stimulate humoral immune responses and induce serum IgG and secretory IgA antibody. Elicits both cytotoxic and memory T-lymphocyte responses. | Pre-existing immunity could decrease immunogenicity. Possible reversion to pathogenic wild-type. | [14,29,60,65,66,67] |
| Vibrio cholerae | M cells and intestinal epithelial cells. Extracellular pathogen. | Ability to adhere to M cells and other epithelial cells without further invasion (decreased pathogenicity). Ideal for delivering antigens from luminal pathogens rather than systemic infections. | Unable to elicit systemic and potent cell-mediated immune responses. Possible reversion to pathogenic state. | [68] |
| Commensal organisms | ||||
| Lactobacillus spp. | Colonize gastrointestinal and uro-genital mucosa. Normal flora. | Non-pathogenic bacteria. Ability to stimulate antigen-specific immune responses via intranasal and oral routes. Special interest for the development of sexually transmitted diseases vaccines. | Unable to elicit cell-mediated immune responses. | [69,70,71] |
| Staphylococcus spp. | Colonize oral, nasal and uro-genital mucosa. Normal flora. | Food grade bacteria with intrinsic safety profiles. Stable colonization by a single intranasal or oral inoculation for more than two months. Ability to stimulate systemic immune responses against heterologous antigens. Strains such as S. carnosus have low extracellular proteolytic activity, which facilitates stable display of heterologous antigens. | Possible cause of pyelonephritis and endocarditis. Pre-existing immunity could decrease immunogenicity. | [72,73,74] |
| Vector | Mutation/Attenuation | Heterologous Antigen | Inoculation Route | Target Host | Outcome | Ref. |
|---|---|---|---|---|---|---|
| Listeria monocytogenes BMB72 | ΔactA ΔactB | Influenza A nucleoprotein | Oral and transcutaneous | Human | All volunteers who received the vector vaccine developed detectible mucosal immune responses to listerial antigens, but not to the heterologous influenza antigen. | [75,76] |
| Bordetella pertussis BPZE1 | Lacking dnt gene and producing inactive pertussis toxin and reduced tracheal cytotoxin. | SP70 derived from enterovirus 71 | Intranasal | Mouse | Strong and sustained systemic anti-SP70 antibody response was observed in nasally immunized mice. | [77] |
| Listeria monocytogenes XFL-7 | prfA-defective | HPV-16 E7 antigen | Intravenous | Human | HPV-16 E7-specific T lymphocyte responses were elicited | [78] |
| Salmonella Typhi | ΔssaV ΔaroC | Escherichia coli heat labile toxin (LT-B) | Oral | Human | Humoral immune responses to LT-B and S. Typhi lipopolysaccharide were observed in 67 and 97% of subjects. | [79] |
| Salmonella Typhi Ty21a | ΔgalE with undefined attenuating mutations | OprF-Oprl derived from Pseudomonas aeruginosa | Oral and intranasal | Human | A significant elevated IgA and IgG antibody levels in the lower airways was observed. | [80] |
| Salmonella Typhi Ty21a | ΔgalE with undefined attenuating mutations | Urease or HP0231 derived from Helicobactor pylori | Oral | Human | T cell-mediated immunity against H. pylori was elicited in immunized subjects. | [81] |
| Salmonella Typhi Ty21a | ΔgalE with undefined attenuating mutations | O-Ps derived from shigella dysenteriae | Oral | Human | Protective immunity was elicited against challenge assay with S. dysenteriae | [82] |
| Vibrio cholera | ΔCTA | Cholera toxin-B | Oral and intranasal | Mouse and rabbit | Cholera toxin has >80% identity to E. coli (ETEC) heat-labile protein. Neutralizing antibody responses against ETEC heat-labile toxicity was observed in vaccinated mice and rabbits. | [83] |
3. Live-Attenuated Bacterial Vectors for Cancer Treatment
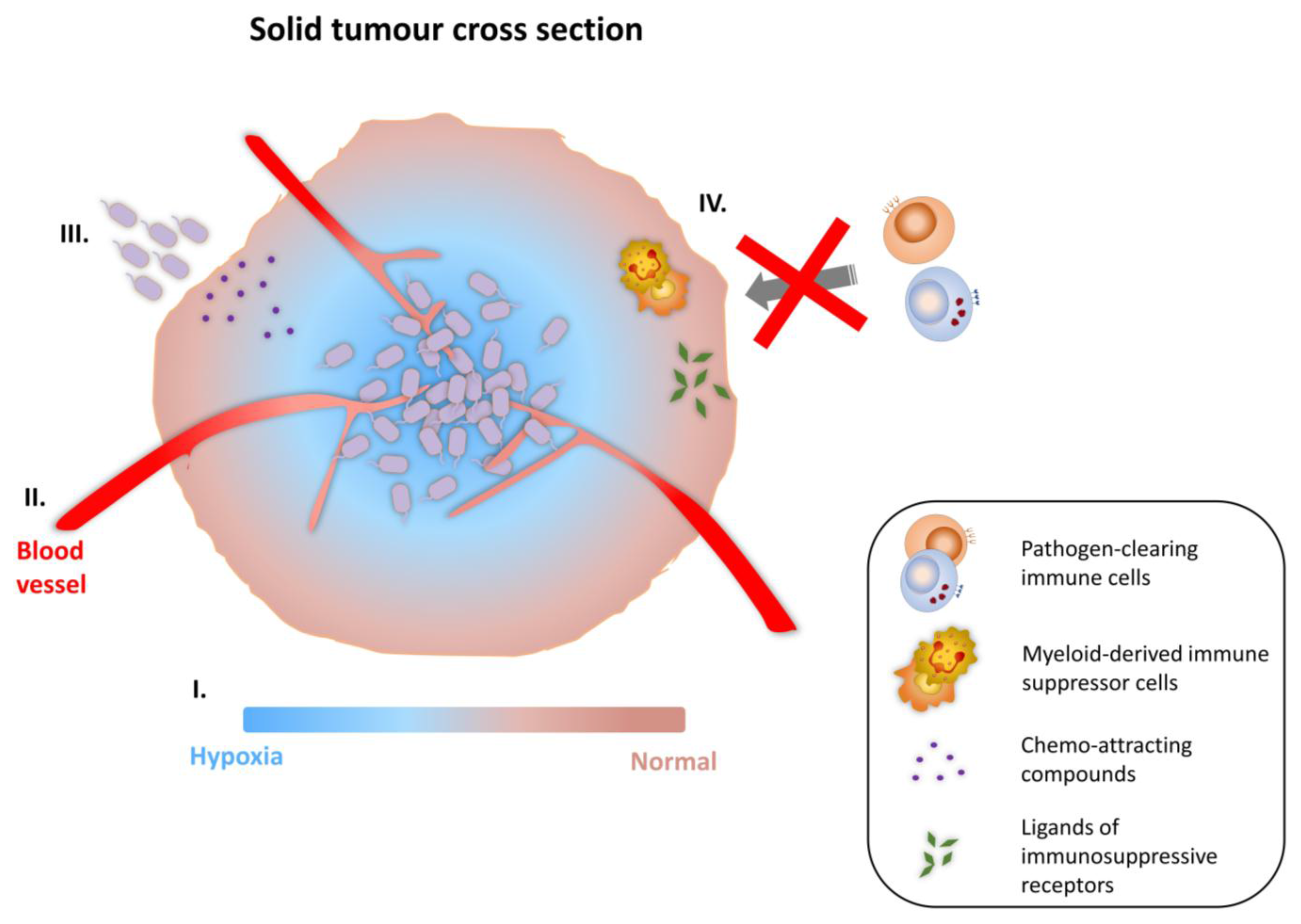
3.1. Tumor-Targeting Ability of Bacterial Vectors
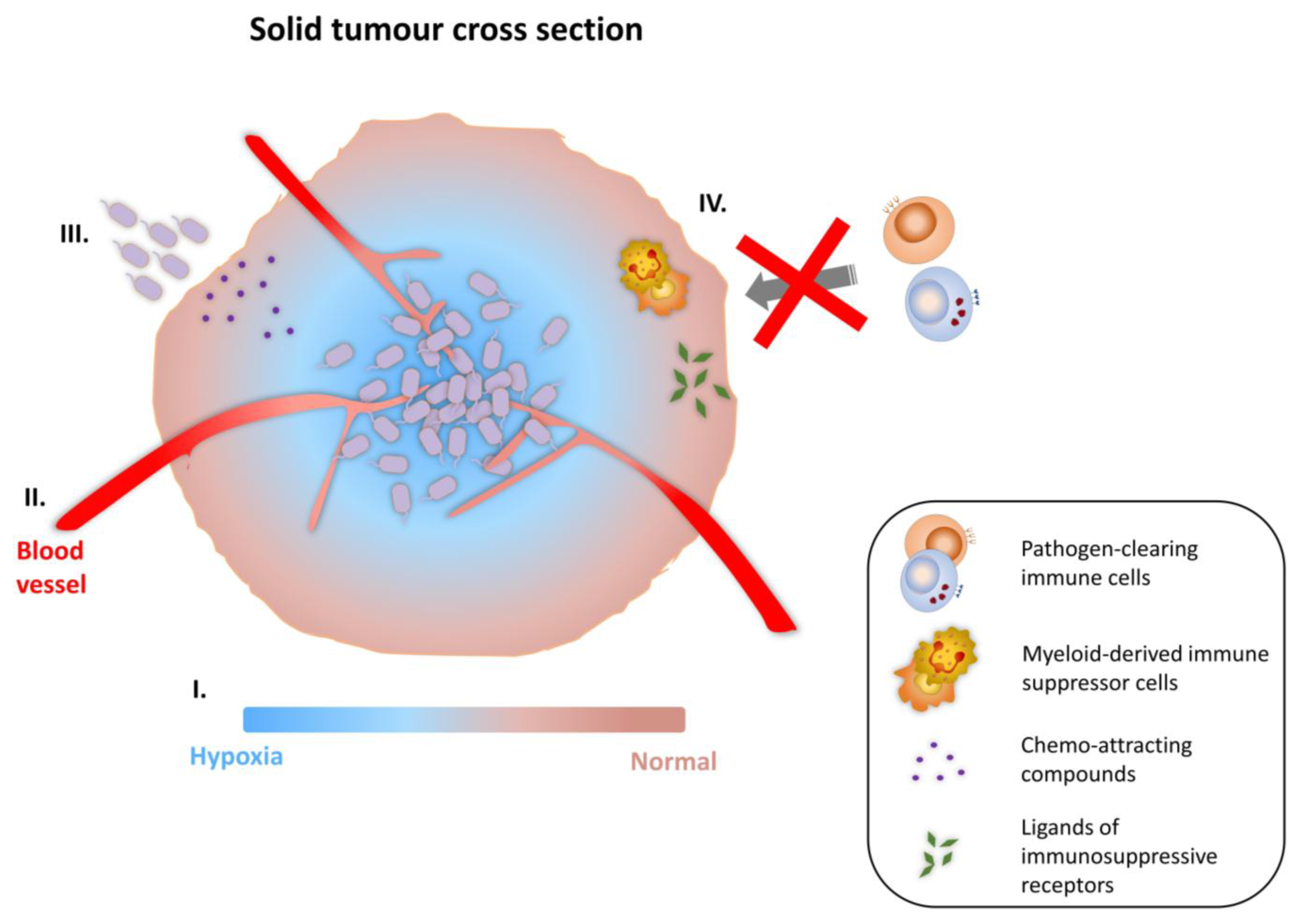
3.2. Strategies for Cancer Therapy
| Mode of Action | Therapeutic Approach | Example of Passenger Gene (or Antigen) | Delivery Vector | References |
|---|---|---|---|---|
| Direct cell killing or tumor growth retardation | Intrinsic oncolytic vector | None | Clostridium spp. Salmonella spp. | [91,92,93,94,95,96] |
| Anti-angiogenic molecule | Vascular endothelial growth factor receptor 2 molecule (fetal liver kinase-1) | L. monocytogenes S. Typhimurium | [97,98] | |
| Endostatin | B. longum B. adolescentis | [99,100,101,102] | ||
| S. choleraesuis | [103] | |||
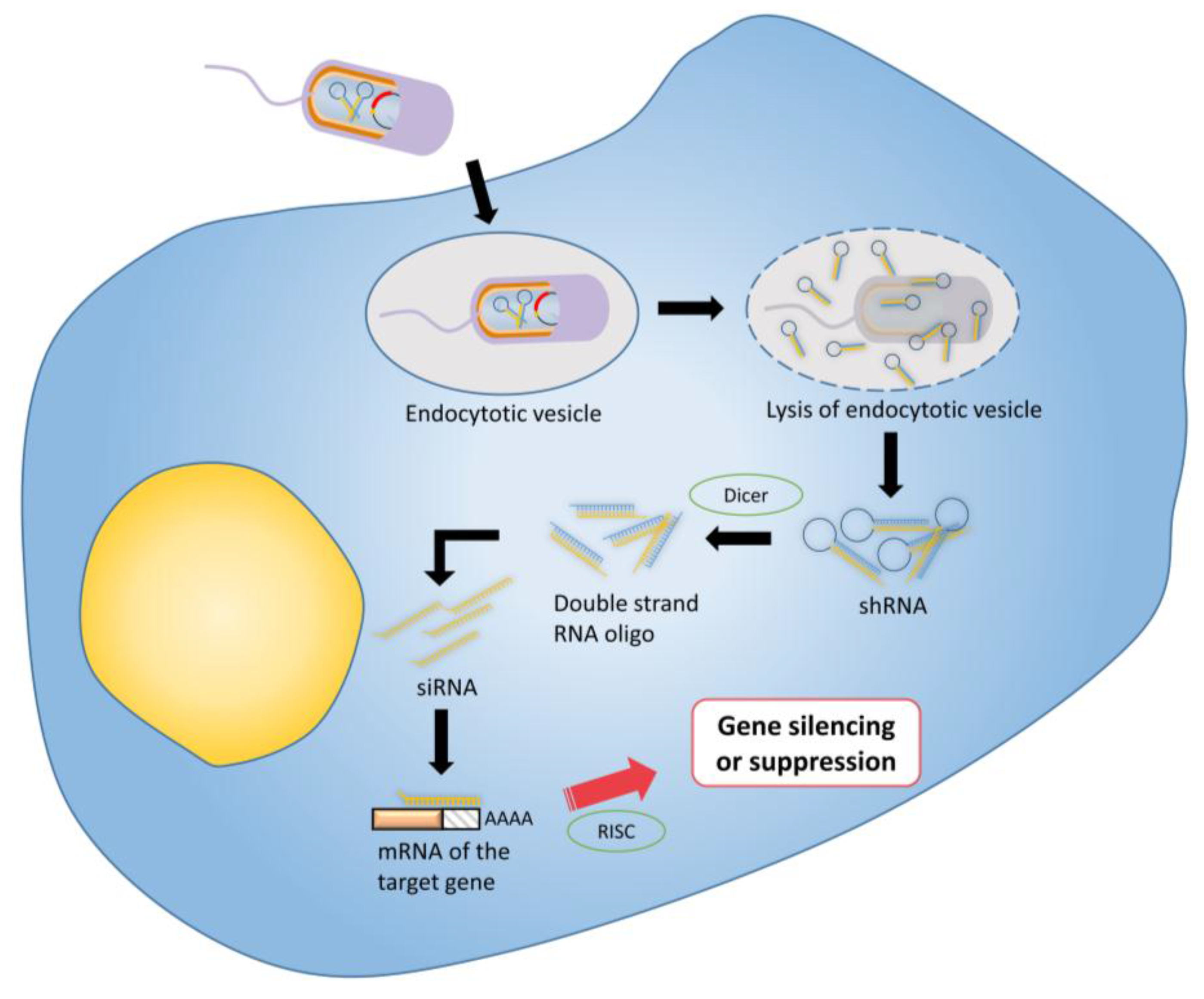
| RNA interference | Anti-bcl2 shRNA | S. Typhimurium | [104] | |
| Anti-MDR1 siRNA | S. Typhi | [105] | ||
| Cell death inducer | Fas ligand | S. Typhimurium | [106] | |
| HylE cytolynsin | S. Typhimurium | [107] | ||
| TNF-related factor apoptosis ligand (TRAIL) | S. Typhimurium B. longum | [101,108,109] | ||
| Apoptin | S. Typhimurium | [110] | ||
| Pro-drug activating enzyme (+drug) | Herpes Simplex Virus thymidine kinase (HSVtk) + ganciclovir | S. Typhimurium | [111,112] | |
| B. infantis | [113] |
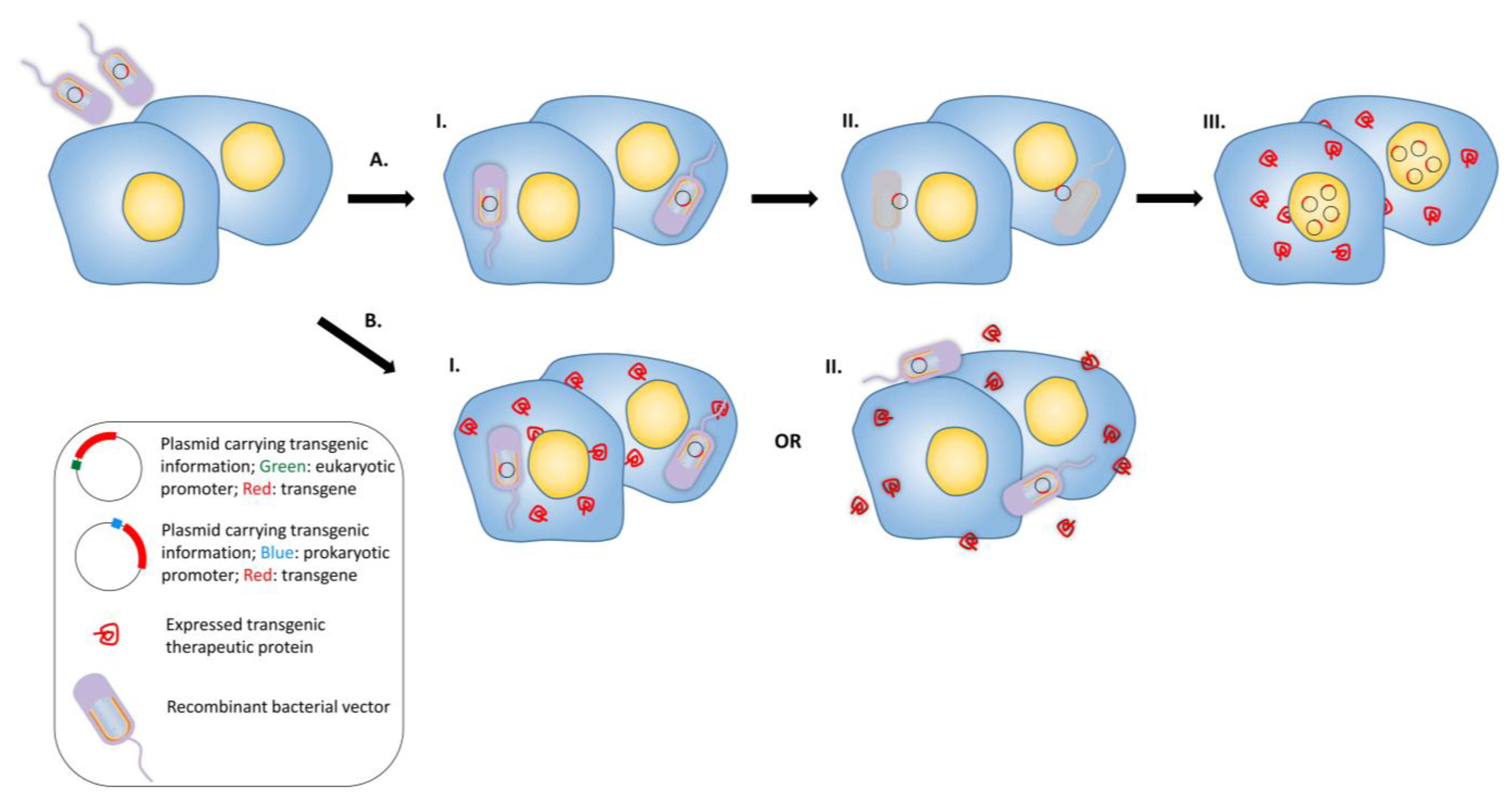
3.3. Anti-Angiogenesis
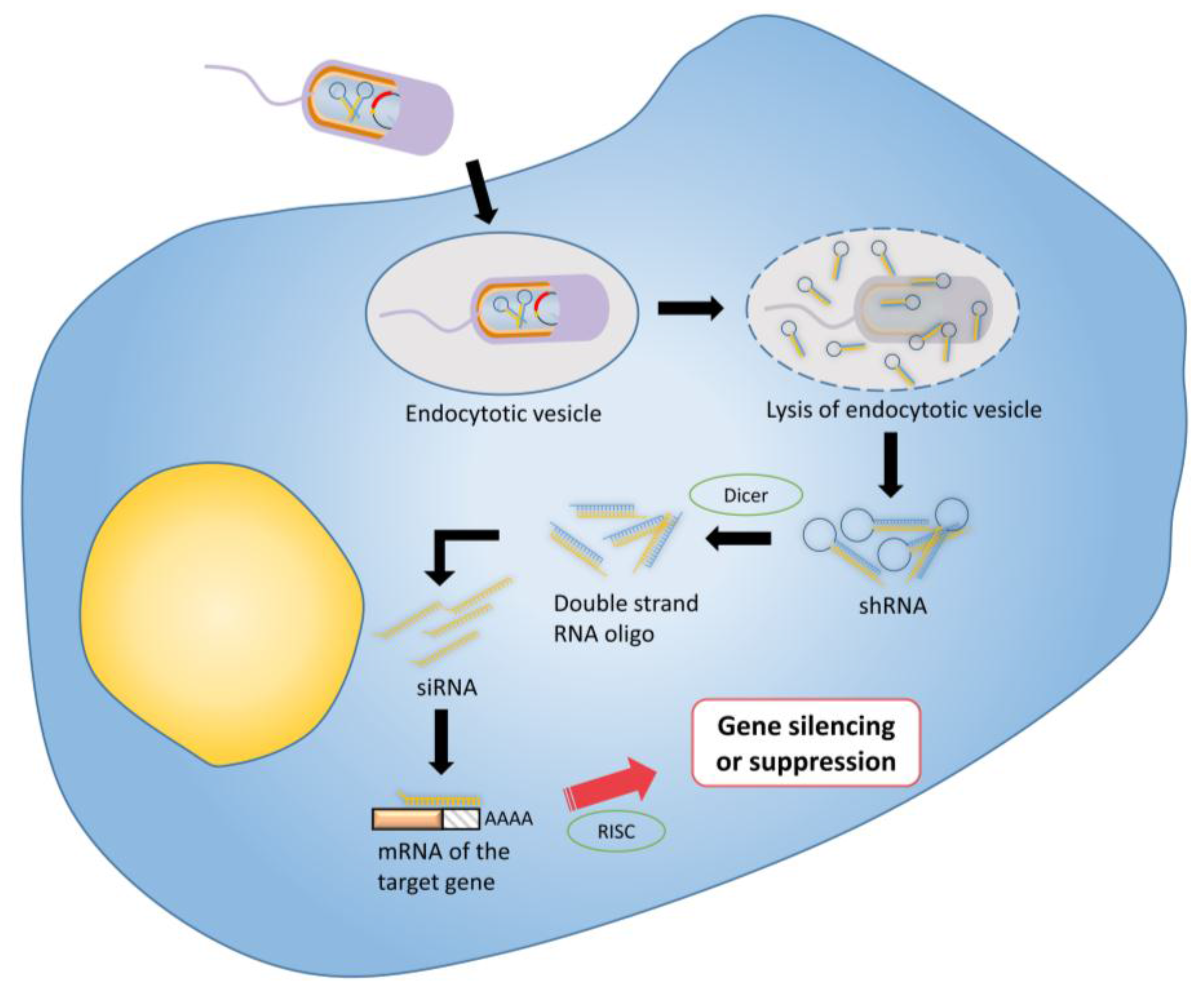
3.4. RNA Interference
3.5. Cell Death Inducer

3.6. Pro-Drug Activating Enzyme + (Drug)
3.7. Immune Stimulatory Molecules
3.8. Tumor Antigen
3.9. In Vivo Tumor Imaging
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Shata, M.T.; Stevceva, L.; Agwale, S.; Lewis, G.K.; Hone, D.M. Recent advances with recombinant bacterial vaccine vectors. Mol. Med. Today 2000, 6, 66–71. [Google Scholar] [CrossRef]
- Carleton, H.A. Pathogenic bacteria as vaccine vectors: Teaching old bugs new tricks. Yale J. Biol. Med. 2010, 83, 217–222. [Google Scholar] [PubMed]
- Van, T.T.H.; Lin, Y.-C.; Van, T.N.N.; Nguyen, T.Q.; Le, T.T.H.; Do, T.H.; Truong, N.H.; Coloe, P.J.; Smooker, P.M. Salmonella as a vaccine vector for influenza virus. Procedia Vaccinol. 2013, 7, 23–27. [Google Scholar] [CrossRef]
- Goya, A.K.; Khatri, K.; Mishra, N.; Vyas, S.P. New patents on mucosal delivery of vaccines. Expert Opin. Ther. Pat. 2008, 18, 1271–1288. [Google Scholar]
- Hone, D.M.; Harris, A.M.; Chatfield, S.; Dougan, G.; Levine, M.M. Construction of genetically defined double aro mutants of Salmonella Typhi. Vaccine 1991, 9, 810–816. [Google Scholar] [CrossRef]
- Strahan, K.; Chatfield, S.N.; Tite, J.; Dougan, G.; Hormaeche, C.E. Impaired resistance to infection does not increase the virulence of Salmonella htrA live vaccines for mice. Microb. Pathog. 1992, 12, 311–317. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Noriega, F.; Losonsky, G.A.; Sztein, M.B.; Wasserman, S.S.; Nataro, J.P.; Levine, M.M. Safety, immunogenicity, and transmissibility in humans of CVD 1203, a live oral Shigella flexneri 2a vaccine candidate attenuated by deletions in aroA and virG. Infect. Immun. 1996, 64, 4542–4548. [Google Scholar] [PubMed]
- van Loon, F.P.; Clemens, J.D.; Chakraborty, J.; Rao, M.R.; Kay, B.A.; Sack, D.A.; Yunus, M.; Ali, M.; Svennerholm, A.M.; Holmgren, J. Field trial of inactivated oral cholera vaccines in Bangladesh: Results from 5 years of follow-up. Vaccine 1996, 14, 162–166. [Google Scholar] [CrossRef]
- Olanratmanee, T.; Levine, M.; Losonsky, G.; Thisyakorn, U.; Cryz, S., Jr. Safety and immunogenicity of Salmonella Typhi Ty21a liquid formulation vaccine in 4 to 6 year old Thai children. J. Infect. Dis. 1992, 166, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Mouricout, M. Interactions between the enteric pathogen and the host. An assortment of bacterial lectins and a set of glycoconjugate receptors. Adv. Exp. Med. Biol. 1997, 412, 109–123. [Google Scholar] [PubMed]
- Leclerc, C. New technologies for vaccine development. Med. Sci. (Paris) 2007, 23, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Gerdts, V.; Mutwiri, G.K.; Tikoo, S.K.; Babiuk, L.A. Mucosal delivery of vaccines in domestic animals. Vet. Res. 2006, 37, 487–510. [Google Scholar] [CrossRef] [PubMed]
- Chinchilla, M.; Pasetti, M.F.; Medina-Moreno, S.; Wang, J.Y.; Gomez-Duarte, O.G.; Stout, R.; Levine, M.M.; Galen, J.E. Enhanced immunity to Plasmodium falciparum circumsporozoite protein (PfCSP) by using Salmonella enterica serovar Typhi expressing PfCSP and a PfCSP-encoding DNA vaccine in a heterologous prime-boost strategy. Infect. Immun. 2007, 75, 3769–3779. [Google Scholar] [CrossRef] [PubMed]
- Saxena, M.; Van, T.T.H.; Baird, F.J.; Coloe, P.J.; Smooker, P.M. Pre-existing immunity against vaccine vectors––Friend or foe? Microbiology 2013, 159, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Germanier, R.; Furer, E. Isolation and characterization of Gal E mutant Ty21a of Salmonella Typhi - candidate strain for a live, oral typhoid vaccine. J. Infect. Dis. 1975, 131, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Oettinger, T.; Jorgensen, M.; Ladefoged, A.; Haslov, K.; Andersen, P. Development of the Mycobacterium bovis BCG vaccine: Review of the historical and biochemical evidence for a genealogical tree. Tuber. Lung Dis. 1999, 79, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Galan, J.E.; Curtiss, R., 3rd. Virulence and vaccine potential of phoP mutants of Salmonella Typhimurium. Microb. Pathog. 1989, 6, 433–443. [Google Scholar] [CrossRef]
- Coloe, P.J.; Alderton, M.R.; Gerraty, N.L.; Christopher, W.; Smith, S.C. Aromatic vitamin-dependent Salmonellae as vaccines in food animals: Efficacy and persistence. Dev. Biol. Stand. 1995, 84, 263–267. [Google Scholar] [PubMed]
- Bachtiar, E.W.; Sheng, K.C.; Fifis, T.; Gamvrellis, A.; Plebanski, M.; Coloe, P.J.; Smooker, P.M. Delivery of a heterologous antigen by a registered Salmonella vaccine (STM1). FEMS Microbiol. Lett. 2003, 227, 211–217. [Google Scholar] [CrossRef]
- Cheminay, C.; Hensel, M. Rational design of Salmonella recombinant vaccines. Int. J. Med. Microbiol. 2008, 298, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.J.; Clare, S.; Pickard, D.; Clark, S.O.; Kelly, D.L.F.; Ghany, M.A.E.; Hale, C.; Dietrich, J.; Andersen, P.; Marsh, P.D.; et al. Characterisation of a live Salmonella vaccine stably expressing the Mycobacterium tuberculosis Ag85B–ESAT6 fusion protein. Vaccine 2009, 27, 6894–6904. [Google Scholar] [CrossRef] [PubMed]
- Darji, A.; Guzman, C.A.; Gerstel, B.; Wachholz, P.; Timmis, K.N.; Wehland, J.; Chakraborty, T.; Weiss, S. Oral somatic transgene vaccination using attenuated S. Typhimurium. Cell 1997, 91, 765–775. [Google Scholar] [CrossRef]
- Curtiss, R., 3rd; Kelly, S.M. Salmonella Typhimurium deletion mutants lacking adenylate cyclase and cyclic AMP receptor protein are avirulent and immunogenic. Infect. Immun. 1987, 55, 3035–3043. [Google Scholar] [PubMed]
- Hall, H.K.; Foster, J.W. The role of fur in the acid tolerance response of Salmonella Typhimurium is physiologically and genetically separable from its role in iron acquisition. J. Bacteriol. 1996, 178, 5683–5691. [Google Scholar] [PubMed]
- Zhang, X.; Wanda, S.-Y.; Brenneman, K.; Kong, W.; Zhang, X.; Roland, K.; Curtiss, R. Improving Salmonella vector with rec mutation to stabilize the DNA cargoes. BMC Microbiol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.Y.; Han, S.J.; Cha, H.R.; Seo, S.U.; Song, J.H.; Chung, S.H.; Kweon, M.N. Eye mucosa: An efficient vaccine delivery route for inducing protective immunity. J. Immunol. 2010, 185, 3610–3619. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, L.; Clements, J.D. Oral immunization using live attenuated Salmonella spp. as carriers of foreign antigens. Clin. Microbiol. Rev. 1992, 5, 328–342. [Google Scholar] [PubMed]
- Kotton, C.N.; Hohmann, E.L. Enteric pathogens as vaccine vectors for foreign antigen delivery. Infect. Immun. 2004, 72, 5535–5547. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.M.; Tacket, C.O.; Sztein, M.B. Host-Salmonella interaction: Human trials. Microb. Infect. 2001, 3, 1271–1279. [Google Scholar] [CrossRef]
- McGhie, E.J.; Brawn, L.C.; Hume, P.J.; Humphreys, D.; Koronakis, V. Salmonella takes control: Effector-driven manipulation of the host. Curr. Opin. Microbiol. 2009, 12, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.L.; Tsolis, R.M.; Baumler, A.J.; Adams, L.G. Pathogenesis of Salmonella-induced enteritis. Braz. J. Med. Biol. Res. 2003, 36, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.C.; Galan, J.E. Manipulation of the host actin cytoskeleton by Salmonella—All in the name of entry. Curr. Opin. Microbiol. 2005, 8, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, E.L.; Oletta, C.A.; Miller, S.I. Evaluation of a phoP/phoQ-deleted, aroA-deleted live oral Salmonella Typhi vaccine strain in human volunteers. Vaccine 1996, 14, 19–24. [Google Scholar] [CrossRef]
- Nardelli-Haefliger, D.; Kraehenbuhl, J.P.; Curtiss, R., 3rd; Schodel, F.; Potts, A.; Kelly, S.; de Grandi, P. Oral and rectal immunization of adult female volunteers with a recombinant attenuated Salmonella Typhi vaccine strain. Infect. Immun. 1996, 64, 5219–5224. [Google Scholar] [PubMed]
- Shi, H.; Santander, J.; Brenneman, K.E.; Wanda, S.Y.; Wang, S.; Senechal, P.; Sun, W.; Roland, K.L.; Curtiss, R. Live recombinant Salmonella Typhi vaccines constructed to investigate the role of rpoS in eliciting immunity to a heterologous antigen. PLoS ONE 2010, 5, e11142. [Google Scholar] [CrossRef] [PubMed]
- Husseiny, M.I.; Hensel, M. Evaluation of Salmonella live vaccines with chromosomal expression cassettes for translocated fusion proteins. Vaccine 2009, 27, 3780–3787. [Google Scholar] [CrossRef] [PubMed]
- Matic, J.N.; Terry, T.D.; van Bockel, D.; Maddocks, T.; Tinworth, D.; Jennings, M.P.; Djordjevic, S.P.; Walker, M.J. Development of non-antibiotic-resistant, chromosomally based, constitutive and inducible expression systems for aroA-attenuated Salmonella enterica serovar Typhimurium. Infect. Immun. 2009, 77, 1817–1826. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, S.J.; Simmons, C.P.; Strugnell, R.A. In vitro and in vivo stability of recombinant plasmids in a vaccine strain of Salmonella enterica var. Typhimurium. FEMS Immunol. Med. Microbiol. 2003, 37, 111–119. [Google Scholar] [CrossRef]
- Million-Weaver, S.; Camps, M. Mechanisms of plasmid segregation: Have multicopy plasmids been overlooked? Plasmid 2014, 75, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Galen, J.E.; Pasetti, M.F.; Tennant, S.; Ruiz-Olvera, P.; Sztein, M.B.; Levine, M.M. Salmonella enterica serovar Typhi live vector vaccines finally come of age. Immunol. Cell Biol. 2009, 87, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Garcillán-Barcia, M.P.; Alvarado, A.; de la Cruz, F. Identification of bacterial plasmids based on mobility and plasmid population biology. FEMS Microbiol. Rev. 2011, 35, 936–956. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M. Paradigms of plasmid organization. Mol. Microbiol. 2000, 37, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Galan, J.E.; Nakayama, K.; Curtiss, R., 3rd. Cloning and characterization of the asd gene of Salmonella Typhimurium: Use in stable maintenance of recombinant plasmids in Salmonella vaccine strains. Gene 1990, 94, 29–35. [Google Scholar] [CrossRef]
- Schodel, F.; Peterson, D.; Hughes, J.; Wirtz, R.; Milich, D. Hybrid hepatitis B virus core antigen as a vaccine carrier moiety: I. presentation of foreign epitopes. J. Biotechnol. 1996, 44, 91–96. [Google Scholar] [CrossRef]
- Yang, X.; Hinnebusch, B.J.; Trunkle, T.; Bosio, C.M.; Suo, Z.; Tighe, M.; Harmsen, A.; Becker, T.; Crist, K.; Walters, N.; et al. Oral vaccination with Salmonella simultaneously expressing Yersinia pestis F1 and V antigens protects against bubonic and pneumonic plague. J. Immunol. 2007, 178, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, E.L.; Oletta, C.A.; Loomis, W.P.; Miller, S.I. Macrophage-inducible expression of a model antigen in Salmonella Typhimurium enhances immunogenicity. Proc. Natl. Acad. Sci. USA 1995, 92, 2904–2908. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, Y.; Shi, H.; Sun, W.; Roland, K.L.; Curtiss, R., 3rd. Comparison of a regulated delayed antigen synthesis system with in vivo-inducible promoters for antigen delivery by live attenuated Salmonella vaccines. Infect. Immun. 2011, 79, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Christman, M.F.; Morgan, R.W.; Jacobson, F.S.; Ames, B.N. Positive control of a regulon for defenses against oxidative stress and some heat-shock proteins in Salmonella Typhimurium. Cell 1985, 41, 753–762. [Google Scholar] [CrossRef]
- Guan, L.; Mu, W.; Champeimont, J.; Wang, Q.; Wu, H.; Xiao, J.; Lubitz, W.; Zhang, Y.; Liu, Q. Iron-regulated lysis of recombinant Escherichia coli in host releases protective antigen and confers biological containment. Infect. Immun. 2011, 79, 2608–2618. [Google Scholar] [CrossRef] [PubMed]
- Garcia Vescovi, E.; Soncini, F.C.; Groisman, E.A. Mg2+ as an extracellular signal: Environmental regulation of Salmonella virulence. Cell 1996, 84, 165–174. [Google Scholar] [CrossRef]
- Chu, T.; Ni, C.; Zhang, L.; Wang, Q.; Xiao, J.; Zhang, Y.; Liu, Q. A quorum sensing-based in vivo expression system and its application in multivalent bacterial vaccine. Microb. Cell Fact. 2015. [Google Scholar] [CrossRef] [PubMed]
- Galen, J.E.; Levine, M.M. Can a “flawless” live vector vaccine strain be engineered? Trends. Microbiol. 2001, 9, 372–376. [Google Scholar] [CrossRef]
- Haddad, D.; Liljeqvist, S.; Kumar, S.; Hansson, M.; Stahl, S.; Perlmann, H.; Perlmann, P.; Berzins, K. Surface display compared to periplasmic expression of a malarial antigen in Salmonella Typhimurium and its implications for immunogenicity. FEMS Immunol. Med. Microbiol. 1995, 12, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.Y.; Curtiss, R., 3rd. Immune responses dependent on antigen location in recombinant attenuated Salmonella Typhimurium vaccines following oral immunization. FEMS Immunol. Med. Microbiol. 2003, 37, 99–104. [Google Scholar] [CrossRef]
- Hobom, G.; Arnold, N.; Ruppert, A. OmpA fusion proteins for presentation of foreign antigens on the bacterial outer membrane. Dev. Biol. Stand. 1995, 84, 255–262. [Google Scholar] [PubMed]
- Charbit, A. Maltodextrin transport through lamB. Front. Biosci. 2003, 8, s265–s274. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, A.; Igimi, S. Innate and acquired immune responses induced by recombinant Lactobacillus casei displaying flagellin-fusion antigen on the cell-surface. Vaccine 2010, 28, 3409–3415. [Google Scholar] [CrossRef] [PubMed]
- Shahabi, V.; Maciag, P.C.; Rivera, S.; Wallecha, A. Live, attenuated strains of Listeria and Salmonella as vaccine vectors in cancer treatment. Bioeng. Bugs 2010, 1, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.K. Live-attenuated Salmonella as a prototype vaccine vector for passenger immunogens in humans: Are we there yet? Expert Rev. Vaccines 2007, 6, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Curtiss, R., III. Bacterial infectious disease control by vaccine development. J. Clin. Invest. 2002, 110, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Byrd, C.M.; Bolken, T.C.; Jones, K.F.; Warren, T.K.; Vella, A.T.; McDonald, J.; King, D.; Blackwood, Z.; Hruby, D.E. Biological consequences of antigen and cytokine co-expression by recombinant Streptococcus gordonii vaccine vectors. Vaccine 2002, 20, 2197–2205. [Google Scholar] [CrossRef]
- Chabalgoity, J.A.; Baz, A.; Rial, A.; Grille, S. The relevance of cytokines for development of protective immunity and rational design of vaccines. Cytokine Growth Factor Rev. 2007, 18, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Braat, H.; Rottiers, P.; Hommes, D.W.; Huyghebaert, N.; Remaut, E.; Remon, J.P.; van Deventer, S.J.; Neirynck, S.; Peppelenbosch, M.P.; Steidler, L. A phase I trial with transgenic bacteria expressing interleukin-10 in Crohn’s disease. Clin. Gastroenterol. Hepatol. 2006, 4, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Bruhn, K.W.; Craft, N.; Miller, J.F. Listeria as a vaccine vector. Microb. Infect. 2007, 9, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Sztein, M.B.; Tanner, M.K.; Polotsky, Y.; Orenstein, J.M.; Levine, M.M. Cytotoxic T lymphocytes after oral immunization with attenuated vaccine strains of Salmonella Typhi in humans. J. Immunol. 1995, 155, 3987–3993. [Google Scholar] [PubMed]
- Sztein, M.B. Cell-mediated immunity and antibody responses elicited by attenuated Salmonella enterica serovar Typhi strains used as live oral vaccines in humans. Clin. Infect. Dis. 2007, 45, S15–S19. [Google Scholar] [CrossRef] [PubMed]
- Salerno-Goncalves, R.; Fernandez-Vina, M.; Lewinsohn, D.M.; Sztein, M.B. Identification of a human HLA-E-restricted CD8+ T cell subset in volunteers immunized with Salmonella enterica serovar Typhi strain Ty21a typhoid vaccine. J. Immunol. 2004, 173, 5852–5862. [Google Scholar] [CrossRef] [PubMed]
- Viret, J.F.; Favre, D.; Wegmuller, B.; Herzog, C.; Que, J.U.; Cryz, S.J., Jr.; Lang, A.B. Mucosal and systemic immune responses in humans after primary and booster immunizations with orally administered invasive and noninvasive live attenuated bacteria. Infect. Immun. 1999, 67, 3680–3685. [Google Scholar] [PubMed]
- Seegers, J.F.M.L. Lactobacilli as live vaccine delivery vectors: Progress and prospects. Trends Biotechnol. 2002, 20, 508–515. [Google Scholar] [CrossRef]
- Pontes, D.S.; de Azevedo, M.S.P.; Chatel, J.-M.; Langella, P.; Azevedo, V.; Miyoshi, A. Lactococcus lactis as a live vector: Heterologous protein production and DNA delivery systems. Protein Expr. Purif. 2011, 79, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Marelli, B.; Perez, A.R.; Banchio, C.; de Mendoza, D.; Magni, C. Oral immunization with live Lactococcus lactis expressing rotavirus VP8* subunit induces specific immune response in mice. J. Virol. Methods 2011, 175, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Mielcarek, N.; Alonso, S.; Locht, C. Nasal vaccination using live bacterial vectors. Adv. Drug Del. Rev. 2001, 51, 55–69. [Google Scholar] [CrossRef]
- Wilson, R.L.; Hruby, D.E. Commensal bacteria as a novel delivery system for subunit vaccines directed against agents of bioterrorism. Adv. Drug Del. Rev. 2005, 57, 1392–1402. [Google Scholar] [CrossRef] [PubMed]
- Oggioni, M.R.; Manganelli, R.; Contorni, M.; Tommasino, M.; Pozzi, G. Immunization of mice by oral colonization with live recombinant commensal Streptococci. Vaccine 1995, 13, 775–779. [Google Scholar] [PubMed]
- Johnson, P.V.; Blair, B.M.; Zeller, S.; Kotton, C.N.; Hohmann, E.L. Attenuated Listeria monocytogenes vaccine vectors expressing influenza A nucleoprotein: Preclinical evaluation and oral inoculation of volunteers. Microbiol. Immunol. 2011, 55, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Eypper, E.H.; Johnson, P.V.; Purro, E.I.; Hohmann, E.L. Transcutaneous immunization of healthy volunteers with an attenuated Listeria monocytogenes vaccine strain and cholera toxin adjuvant. Vaccine 2013, 31, 3257–3261. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.Y.; Chua, S.Q.; Foo, D.G.W.; Locht, C.; Chow, V.T.; Poh, C.L.; Alonso, S. Highly attenuated Bordetella pertussis strain BPZE1 as a potential live vehicle for delivery of heterologous vaccine candidates. Infect. Immun. 2008, 76, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Maciag, P.C.; Radulovic, S.; Rothman, J. The first clinical use of a live-attenuated Listeria monocytogenes vaccine: A Phase I safety study of Lm-LLO-E7 in patients with advanced carcinoma of the cervix. Vaccine 2009, 27, 3975–3983. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chatfield, S.; Stratford, R.; Bedwell, J.; Bentley, M.; Sulsh, S.; Giemza, R.; Smith, S.; Bongard, E.; Cosgrove, C.A.; et al. Ability of SPI2 mutant of S. Typhi to effectively induce antibody responses to the mucosal antigen enterotoxigenic E. coli heat labile toxin B subunit after oral delivery to humans. Vaccine 2007, 25, 4175–4182. [Google Scholar] [CrossRef] [PubMed]
- Bumann, D.; Behre, C.; Behre, K.; Herz, S.; Gewecke, B.; Gessner, J.E.; von Specht, B.U.; Baumann, U. Systemic, nasal and oral live vaccines against Pseudomonas aeruginosa: A clinical trial of immunogenicity in lower airways of human volunteers. Vaccine 2010, 28, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Aebischer, T.; Bumann, D.; Epple, H.J.; Metzger, W.; Schneider, T.; Cherepnev, G.; Walduck, A.K.; Kunkel, D.; Moos, V.; Loddenkemper, C.; et al. Correlation of T cell response and bacterial clearance in human volunteers challenged with Helicobacter pylori revealed by randomised controlled vaccination with Ty21a-based Salmonella vaccines. Gut 2008, 57, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Xu de, Q.; Cisar, J.O.; Osorio, M.; Wai, T.T.; Kopecko, D.J. Core-linked LPS expression of Shigella dysenteriae serotype 1 O-antigen in live Salmonella Typhi vaccine vector Ty21a: Preclinical evidence of immunogenicity and protection. Vaccine 2007, 25, 6167–6175. [Google Scholar] [PubMed]
- Roland, K.L.; Cloninger, C.; Kochi, S.K.; Thomas, L.J.; Tinge, S.A.; Rouskey, C.; Killeen, K.P. Construction and preclinical evaluation of recombinant Peru-15 expressing high levels of the cholera toxin B subunit as a vaccine against enterotoxigenic Escherichia coli. Vaccine 2007, 25, 8574–8584. [Google Scholar] [CrossRef] [PubMed]
- Van, T.T.H.; Lin, Y.C.; Coloe, P.J.; Smooker, P.M. From animals to humans: Can vaccines make the transition? In Vaccinations: Procedures, Types and Controversy; Bezio, A.I., Campbell, B.E., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2012; pp. 1–63. [Google Scholar]
- Kasinskas, R.W.; Forbes, N.S. Salmonella Typhimurium lacking ribose chemoreceptors localize in tumor quiescence and induce apoptosis. Cancer Res. 2007, 67, 3201–3209. [Google Scholar] [CrossRef] [PubMed]
- Baban, C.K.; Cronin, M.; O’Hanlon, D.; O’Sullivan, G.C.; Tangney, M. Bacteria as vectors for gene therapy of cancer. Bioeng. Bugs 2010, 1, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.A.; Shabahang, S.; Timiryasova, T.M.; Zhang, Q.; Beltz, R.; Gentschev, I.; Goebel, W.; Szalay, A.A. Visualization of tumors and metastases in live animals with bacteria and vaccinia virus encoding light-emitting proteins. Nat. Biotechnol. 2004, 22, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Kerkar, S.P.; Restifo, N.P. Cellular constituents of immune escape within the tumor microenvironment. Cancer Res. 2012, 72, 3125–3130. [Google Scholar] [CrossRef] [PubMed]
- Sitkovsky, M.V.; Lukashev, D.; Apasov, S.; Kojima, H.; Koshiba, M.; Caldwell, C.; Ohta, A.; Thiel, M. Physiological control of immune response and inflammatory tissue damage by hypoxia-inducible factors and adenosine A2A receptors. Annu. Rev. Immunol. 2004, 22, 657–682. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, H.A.; Theurich, S.; Shimabukuro-Vornhagen, A.; Holtick, U.; Stippel, D.L.; von Bergwelt-Baildon, M. Overcoming tumor-mediated immunosuppression. Immunotherapy 2014, 6, 973–988. [Google Scholar] [CrossRef] [PubMed]
- Leschner, S.; Weiss, S. Salmonella-allies in the fight against cancer. J. Mol. Med. (Berl.) 2010, 88, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Möse, J.R.; Möse, G. Oncolysis by Clostridia. I. activity of Clostridium Butyricum (M-55) and other nonpathogenic clostridia against the ehrlich carcinoma. Cancer Res. 1964, 24, 212–216. [Google Scholar]
- Luo, X.; Li, Z.; Lin, S.; Le, T.; Ittensohn, M.; Bermudes, D.; Runyab, J.D.; Shen, S.Y.; Chen, J.; King, I.C.; et al. Antitumor effect of VNP20009, an attenuated Salmonella, in murine tumor models. Oncol. Res. 2001, 12, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yang, M.; Li, X.-M.; Jiang, P.; Baranov, E.; Li, S.; Xu, M.; Penman, S.; Hoffman, R.M. Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing Salmonella Typhimurium. Proc. Natl. Acad. Sci. USA 2005, 102, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Mengesha, A.; Wei, J.Z.; Zhou, S.F.; Wei, M.Q. Clostridial spores to treat solid tumours-potential for a new therapeutic modality. Curr. Gene Ther. 2010, 10, 15–26. [Google Scholar] [PubMed]
- Umer, B.; Good, D.; Ann, J.; Duan, W.; Wei, M.Q. Clostridial spores for cancer therapy: Targeting solid tumour microenvironment. J. Toxicol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.G.; Chen, Y.; Wu, Z.P.; Liu, X.; Liu, C.; Zhou, Y.C.; Wu, C.L.; Jin, C.G.; Gu, Y.L.; Li, J.; et al. Orally administered DNA vaccine delivery by attenuated Salmonella Typhimurium targeting fetal liver kinase 1 inhibits murine Lewis lung carcinoma growth and metastasis. Biol. Pharm. Bull. 2010, 33, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Seavey, M.M.; Maciag, P.C.; Al-Rawi, N.; Sewell, D.; Paterson, Y. An anti-vascular endothelial growth factor receptor 2/fetal liver kinase-1 Listeria monocytogenes anti-angiogenesis cancer vaccine for the treatment of primary and metastatic her-2/neu+ breast tumors in a mouse model. J. Immunol. 2009, 182, 5537–5546. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Chen, X.; Kou, L.; Hu, B.; Zhu, L.P.; Fan, Y.R.; Wu, Z.W.; Wang, J.J.; Xu, G.X. Selenium-Bifidobacterium longum as a delivery system of endostatin for inhibition of pathogenic bacteria and selective regression of solid tumor. Exp. Ther. Med. 2010, 1, 129–135. [Google Scholar] [PubMed]
- Li, X.; Fu, G.F.; Fan, Y.R.; Liu, W.H.; Liu, X.J.; Wang, J.J.; Xu, G.X. Bifidobacterium adolescentis as a delivery system of endostatin for cancer gene therapy: Selective inhibitor of angiogenesis and hypoxic tumor growth. Cancer Gene Ther. 2003, 10, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.A.N.; Kou, L.E.I.; Wang, J.-J.; Xu, G.-X. Therapeutic efficacy of Bifidobacterium longum-mediated human interleukin-2 with endostatin or TRAIL in transplanted tumors in mice. Exp. Ther. Med. 2012, 3, 481–486. [Google Scholar] [PubMed]
- Xu, Y.F.; Zhu, L.P.; Hu, B.; Fu, G.F.; Zhang, H.Y.; Wang, J.J.; Xu, G.X. A new expression plasmid in Bifidobacterium longum as a delivery system of endostatin for cancer gene therapy. Cancer Gene Ther. 2007, 14, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.L.; Shiau, A.L. Endostatin gene therapy delivered by Salmonella choleraesuis in murine tumor models. J. Gene Med. 2004, 6, 1382–1393. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Zhu, X.; Chen, L.; Li, S.; Ren, D. Oral administration of attenuated S. Typhimurium carrying shRNA-expressing vectors as a cancer therapeutic. Cancer Biol. Ther. 2008, 7, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhao, P.; Zhou, Z.; Liu, J.; Qin, L.; Wang, H. Using attenuated Salmonella Typhi as tumor targeting vector for MDR1 siRNA delivery. Cancer Biol. Ther. 2007, 6, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Inhibition of tumor growth using Salmonella expressing Fas Ligand. J. Natl. Cancer Inst. 2008, 100, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.M.; Green, J.; Williams, P.J.; Tazzyman, S.; Hunt, S.; Harmey, J.H.; Kehoe, S.C.; Lewis, C.E. Bacterial delivery of a novel cytolysin to hypoxic areas of solid tumors. Gene Ther. 2009, 16, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Ganai, S.; Arenas, R.B.; Forbes, N.S. Tumour-targeted delivery of TRAIL using Salmonella Typhimurium enhances breast cancer survival in mice. Br. J. Cancer 2009, 101, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, B.; Cheng, X.; Qiao, Y.; Tang, B.; Chen, G.; Wei, J.; Liu, X.; Cheng, W.; Du, P.; et al. Salmonella-mediated tumor-targeting TRAIL gene therapy significantly suppresses melanoma growth in mouse model. Cancer Sci. 2012, 103, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Guan, G.F.; Zhao, M.; Liu, L.M.; Jin, C.S.; Sun, K.; Zhang, D.J.; Yu, D.J.; Cao, H.W.; Lu, Y.Q.; Wen, L.J. Salmonella Typhimurium mediated delivery of Apoptin in human laryngeal cancer. Int. J. Med. Sci. 2013, 10, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Zhang, J.; Zhang, J.; Liu, Q.; Wang, S.; Wu, S.; Peng, X.; Shao, J.; Huang, W. Suppression of murine melanoma growth by a vaccine of attenuated Salmonella carrying heat shock protein 70 and Herpes simplex virus-thymidine kinase genes. Oncol. Rep. 2012, 27, 798–806. [Google Scholar] [PubMed]
- Pawelek, J.M.; Low, K.B.; Bermudes, D. Tumor-targeted Salmonella as a novel anticancer vector. Cancer Res. 1997, 57, 4537–4544. [Google Scholar]
- Tang, W.; He, Y.; Zhou, S.; Ma, Y.; Liu, G. A novel Bifidobacterium infantis-mediated TK/GCV suicide gene therapy system exhibits antitumor activity in a rat model of bladder cancer. J. Exp. Clin. Cancer Res. 2009, 28, 155. [Google Scholar] [CrossRef]
- Palffy, R.; Gardlik, R.; Hodosy, J.; Behuliak, M.; Resko, P.; Radvansky, J.; Celec, P. Bacteria in gene therapy: Bactofection versus alternative gene therapy. Gene Ther. 2005, 13, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Wahid, A.; Faubert, G. Mucosal delivery of a transmission-blocking DNA vaccine encoding Giardia lamblia CWP2 by Salmonella Typhimurium bactofection vehicle. Vaccine 2007, 25, 8372–8383. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Lan, H.; Li, S.; Han, X.; Gao, T.; Ren, D. Synergistic antitumor efficacy of suicide/ePNP gene and 6-methylpurine 2’-deoxyriboside via Salmonella against murine tumors. Cancer Gene Ther. 2008, 15, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.L.; Shiau, A.L. Systemic administration of attenuated Salmonella choleraesuis carrying thrombospondin-1 gene leads to tumor-specific transgene expression, delayed tumor growth and prolonged survival in the murine melanoma model. Cancer Gene Ther. 2005, 12, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Sizemore, D.R.; Branstrom, A.A.; Sadoff, J.C. Attenuated Shigella as a DNA delivery vehicle for DNA-mediated immunization. Science 1995, 270, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, G.; Bubert, A.; Gentschev, I.; Sokolovic, Z.; Simm, A.; Catic, A.; Kaufmann, S.H.; Hess, J.; Szalay, A.A.; Goebel, W. Delivery of antigen-encoding plasmid DNA into the cytosol of macrophages by attenuated suicide Listeria monocytogenes. Nat. Biotechnol. 1998, 16, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Buttaro, C.; Fruehauf, J.H. Engineered E. coli as vehicles for targeted therapeutics. Curr. Gene Ther. 2010, 10, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Friedlos, F.; Lehouritis, P.; Ogilvie, L.; Hedley, D.; Davies, L.; Bermudes, D.; King, I.; Martin, J.; Marais, R.; Springer, C.J. Attenuated Salmonella targets prodrug activating enzyme carboxypeptidase G2 to mouse melanoma and human breast and colon carcinomas for effective suicide gene therapy. Clin. Cancer Res. 2008, 14, 4259–4266. [Google Scholar] [CrossRef] [PubMed]
- Loessner, H.; Weiss, S. Bacteria-mediated DNA transfer in gene therapy and vaccination. Expert Opin. Biol. Ther. 2004, 4, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Fujimori, M.; Hamaji, Y.; Hama, Y.; Ito, K.; Amano, J.; Taniguchi, S. Genetically engineered Bifidobacterium longum for tumor-targeting enzyme-prodrug therapy of autochthonous mammary tumors in rats. Cancer Sci. 2006, 97, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Yazawa, K.; Fujimori, M.; Nakamura, T.; Sasaki, T.; Amano, J.; Kano, Y.; Taniguchi, S. Bifidobacterium longum as a delivery system for gene therapy of chemically induced rat mammary tumors. Breast Cancer Res. Treat. 2001, 66, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.P.; Yin, Y.; Xing, J.; Li, C.; Kou, L.; Hu, B.; Wu, Z.W.; Wang, J.J.; Xu, G.X. Therapeutic efficacy of Bifidobacterium longum-mediated human granulocyte colony-stimulating factor and/or endostatin combined with cyclophosphamide in mouse-transplanted tumors. Cancer Sci. 2009, 100, 1986–1990. [Google Scholar] [CrossRef] [PubMed]
- Howarth, J.L.; Lee, Y.B.; Uney, J.B. Using viral vectors as gene transfer tools (Cell Biology and Toxicology Special Issue: ETCS-UK 1 day meeting on genetic manipulation of cells). Cell Biol. Toxicol. 2010, 26, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Al-Dosari, M.S.; Gao, X. Nonviral gene delivery: Principle, limitations, and recent progress. AAPS J. 2009, 11, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Schoen, C.; Kolb-Maurer, A.; Geginat, G.; Loffler, D.; Bergmann, B.; Stritzker, J.; Szalay, A.A.; Pilgrim, S.; Goebel, W. Bacterial delivery of functional messenger RNA to mammalian cells. Cell Microbiol. 2005, 7, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Southwick, F.S.; Purich, D.L. Listeria and Shigella actin-based motility in host cells. Trans. Am. Clin. Climatol. Assoc. 1998, 109, 160–173. [Google Scholar] [PubMed]
- Tilney, L.G.; Portnoy, D.A. Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite, Listeria monocytogenes. J. Cell Biol. 1989, 109, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Darji, A.; zur Lage, S.; Garbe, A.I.; Chakraborty, T.; Weiss, S. Oral delivery of DNA vaccines using attenuated Salmonella Typhimurium as carrier. FEMS Immunol. Med. Microbiol. 2000, 27, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Paterson, Y.; Guirnalda, P.D.; Wood, L.M. Listeria and Salmonella bacterial vectors of tumor-associated antigens for cancer immunotherapy. Semin. Immunol. 2010, 22, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Wallecha, A.; Carroll, K.D.; Maciag, P.C.; Rivera, S.; Shahabi, V.; Paterson, Y. Multiple effector mechanisms induced by recombinant Listeria monocytogenes anticancer immunotherapeutics. Adv. Appl. Microbiol. 2009, 66, 1–27. [Google Scholar] [PubMed]
- Stark, F.C.; Sad, S.; Krishnan, L. Intracellular bacterial vectors that induce CD8+ T cells with similar cytolytic abilities but disparate memory phenotypes provide contrasting tumor protection. Cancer Res. 2009, 69, 4327–4334. [Google Scholar] [CrossRef] [PubMed]
- Gardlik, R.; Behuliak, M.; Palffy, R.; Celec, P.; Li, C.J. Gene therapy for cancer: Bacteria-mediated anti-angiogenesis therapy. Gene Ther. 2011, 18, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.J.; Xu, H.M.; Ma, D.Y.; Hu, Q.G.; Huang, X.F.; Jiang, W.H.; Li, S.F.; Jia, K.Z.; Huang, Q.L.; Hua, Z.C. Enhanced therapeutic effect by combination of tumor-targeting Salmonella and endostatin in murine melanoma model. Cancer Biol. Ther. 2005, 4, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.F.; Li, X.; Hou, Y.Y.; Fan, Y.R.; Liu, W.H.; Xu, G.X. Bifidobacterium longum as an oral delivery system of endostatin for gene therapy on solid liver cancer. Cancer Gene Ther. 2005, 12, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Kou, L.; Li, C.; Zhu, L.P.; Fan, Y.R.; Wu, Z.W.; Wang, J.J.; Xu, G.X. Bifidobacterium longum as a delivery system of TRAIL and endostatin cooperates with chemotherapeutic drugs to inhibit hypoxic tumor growth. Cancer Gene Ther. 2009, 16, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, Z.; Wientjes, M.G.; Au, J.L.S. Delivery of siRNA therapeutics: Barriers and carriers. AAPS J. 2010, 12, 492–503. [Google Scholar] [PubMed]
- Khatri, N.I.; Rathi, M.N.; Baradia, D.P.; Trehan, S.; Misra, A. in vivo delivery aspects of miRNA, shRNA and siRNA. Crit. Rev. Ther. Drug Carrier Syst. 2012, 29, 487–527. [Google Scholar] [CrossRef] [PubMed]
- Lage, H.; Kruhn, A. Bacterial delivery of RNAi effectors: Transkingdom RNAi. J. Vis. Exp. 2010. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.Y.; Xie, F.; Woodle, M.C. In vivo application of RNA interference: From functional genomics to therapeutics. Adv. Genet. 2005, 54, 117–142. [Google Scholar] [PubMed]
- Xiang, S.; Keates, A.C.; Fruehauf, J.; Yang, Y.; Guo, H.; Nguyen, T.; Li, C.J. In vitro and in vivo gene silencing by TransKingdom RNAi (tkRNAi). Methods Mol. Biol. 2009, 487, 147–160. [Google Scholar] [PubMed]
- Zhao, H.F.; L’Abbe, D.; Jolicoeur, N.; Wu, M.; Li, Z.; Yu, Z.; Shen, S.H. High-throughput screening of effective siRNAs from RNAi libraries delivered via bacterial invasion. Nat. Methods 2005, 2, 967–973. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Fruehauf, J.; Li, C.J. Short hairpin RNA-expressing bacteria elicit RNA interference in mammals. Nat. Biotechnol. 2006, 24, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Morin, P.J. beta-Catenin signaling and cancer. Bioessays 1999, 21, 1021–1030. [Google Scholar] [CrossRef]
- Zhang, L.; Gao, L.; Zhao, L.; Guo, B.; Ji, K.; Tian, Y.; Wang, J.; Yu, H.; Hu, J.; Kalvakolanu, D. Intratumoral delivery and suppression of prostate tumor growth by attenuated Salmonella enterica serovar Typhimurium carrying plasmid-based small interfering RNAs. Cancer Res. 2007, 67, 5859–5864. [Google Scholar] [CrossRef] [PubMed]
- Blache, C.A.; Manuel, E.R.; Kaltcheva, T.I.; Wong, A.N.; Ellenhorn, J.D.; Blazar, B.R.; Diamond, D.J. Systemic delivery of Salmonella Typhimurium transformed with IDO shRNA enhances intratumoral vector colonization and suppresses tumor growth. Cancer Res. 2012, 72, 6447–6456. [Google Scholar] [CrossRef] [PubMed]
- Thorburn, A. Death receptor-induced cell killing. Cell. Signal. 2004, 16, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Backendorf, C.; Noteborn, M.H. Apoptin towards safe and efficient anticancer therapies. Adv. Exp. Med. Biol. 2014, 818, 39–59. [Google Scholar] [PubMed]
- Weyel, D.; Sedlacek, H.H.; Muller, R.; Brusselbach, S. Secreted human beta-glucuronidase: A novel tool for gene-directed enzyme prodrug therapy. Gene Ther. 2000, 7, 224–231. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Springer, C.J.; Niculescu-Duvaz, I. Prodrug-activating systems in suicide gene therapy. J. Clin. Invest. 2000, 105, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Moolten, F.L. Tumor chemosensitivity conferred by inserted herpes thymidine kinase genes: Paradigm for a prospective cancer control strategy. Cancer Res. 1986, 46, 5276–5281. [Google Scholar] [PubMed]
- Fox, M.E.; Lemmon, M.J.; Mauchline, M.L.; Davis, T.O.; Giaccia, A.J.; Minton, N.P.; Brown, J.M. Anaerobic bacteria as a delivery system for cancer gene therapy: In vitro activation of 5-fluorocytosine by genetically engineered Clostridia. Gene Ther. 1996, 3, 173–178. [Google Scholar] [PubMed]
- Nuyts, S.; Theys, J.; Landuyt, W.; van Mellaert, L.; Lambin, P.; Anne, J. Increasing specificity of anti-tumor therapy: Cytotoxic protein delivery by non-pathogenic clostridia under regulation of radio-induced promoters. Anticancer Res. 2001, 21, 857–861. [Google Scholar] [PubMed]
- Nakamura, T.; Sasaki, T.; Fujimori, M.; Yazawa, K.; Kano, Y.; Amano, J.; Taniguchi, S. Cloned cytosine deaminase gene expression of Bifidobacterium longum and application to enzyme/pro-drug therapy of hypoxic solid tumors. Biosci. Biotechnol. Biochem. 2002, 66, 2362–2366. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Theys, J.; Landuyt, W.; Anne, J.; Lambin, P. Optimization of tumor-targeted gene delivery by engineered attenuated Salmonella Typhimurium. Anticancer Res. 2002, 22, 3261–3266. [Google Scholar] [PubMed]
- Theys, J.; Pennington, O.; Dubois, L.; Anlezark, G.; Vaughan, T.; Mengesha, A.; Landuyt, W.; Anné, J.; Burke, P.J.; Dûrre, P.; et al. Repeated cycles of Clostridium-directed enzyme prodrug therapy result in sustained antitumour effects in vivo. Br. J. Cancer 2006, 95, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.J.; van Zijl, P.; Fox, M.E.; Mauchline, M.L.; Giaccia, A.J.; Minton, N.P.; Brown, J.M. Anaerobic bacteria as a gene delivery system that is controlled by the tumor microenvironment. Gene Ther. 1997, 4, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Lan, H.; Liang, S.; Gao, T.; Ren, D. Suicide gene/prodrug therapy using Salmonella-mediated delivery of Escherichia coli purine nucleoside phosphorylase gene and 6-methoxypurine 2’-deoxyriboside in murine mammary carcinoma 4T1 model. Cancer Sci. 2008, 99, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Nemunaitis, J.; Cunningham, C.; Senzer, N.; Kuhn, J.; Cramm, J.; Litz, C.; Cavagnolo, R.; Cahill, A.; Clairmont, C.; Sznol, M. Pilot trial of genetically modified, attenuated Salmonella expressing the E. coli cytosine deaminase gene in refractory cancer patients. Cancer Gene Ther. 2003, 10, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Borrello, I.; Pardoll, D. GM-CSF-based cellular vaccines: A review of the clinical experience. Cytokine Growth Factor Rev. 2002, 13, 185–193. [Google Scholar] [CrossRef]
- O’Connell, L.; O’Sullivan, G.C.; Collins, J.K.; Shanahan, F. The Fas counterattack: Fas-mediated T cell killing by colon cancer cells expressing Fas ligand. J. Exp. Med. 1996, 184, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Shevach, E.M. CD4+CD25+ suppressor T cells: More questions than answers. Nat. Rev. Immunol. 2002, 2, 389–400. [Google Scholar] [PubMed]
- Bui, J.D.; Schreiber, R.D. Cancer immunosurveillance, immunoediting and inflammation: Independent or interdependent processes? Curr. Opin. Immunol. 2007, 19, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Salmonella Typhimurium engineered to produce CCL21 inhibit tumor growth. Cancer Immunol. Immunother. 2009, 58, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.S.; Chae, Y.S.; Hong, J.; Park, Y.K. Antitumor therapeutic effects of a genetically engineered Salmonella Typhimurium harboring TNF-alpha in mice. Appl. Microbiol. Biotechnol. 2011, 89, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. IL-18-producing Salmonella inhibit tumor growth. Cancer Gene Ther. 2008, 15, 787–794. [Google Scholar] [CrossRef] [PubMed]
- al-Ramadi, B.K.; Fernandez-Cabezudo, M.J.; El-Hasasna, H.; Al-Salam, S.; Bashir, G.; Chouaib, S. Potent anti-tumor activity of systemically-administered IL2-expressing Salmonella correlates with decreased angiogenesis and enhanced tumor apoptosis. Clin. Immunol. 2009, 130, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, M.; Le’Negrate, G.; Krajewska, M.; Reed, J.C. Attenuated Salmonella engineered to produce human cytokine LIGHT inhibit tumor growth. Proc. Natl. Acad. Sci. USA 2007, 104, 12879–12883. [Google Scholar] [CrossRef] [PubMed]
- Agorio, C.; Schreiber, F.; Sheppard, M.; Mastroeni, P.; Fernandez, M.; Martinez, M.A.; Chabalgoity, J.A. Live attenuated Salmonella as a vector for oral cytokine gene therapy in melanoma. J. Gene Med. 2007, 9, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Finn, O.J. T-cell death and cancer immune tolerance. Cell Death Differ. 2007, 15, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D. Does the immune system see tumors as foreign or self? Annu. Rev. Immunol. 2003, 21, 807–839. [Google Scholar] [CrossRef] [PubMed]
- Hobohm, U.; Stanford, J.L.; Grange, J.M. Pathogen-associated molecular pattern in cancer immunotherapy. Crit. Rev. Immunol. 2008, 28, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Wallecha, A. Cancer immunotherapy using recombinant Listeria monocytogenes: Transition from bench to clinic. Hum. Vaccines 2011, 7, 497–505. [Google Scholar] [CrossRef]
- Hernandez-Luna, M.A.; Luria-Perez, R.; Huerta-Yepez, S. Therapeutic intervention alternatives in cancer, using attenuated live bacterial vectors: Salmonella enterica as a carrier of heterologous molecules. Rev. Invest. Clin. 2013, 65, 65–73. [Google Scholar] [PubMed]
- Wood, L.M.; Guirnalda, P.D.; Seavey, M.M.; Paterson, Y. Cancer immunotherapy using Listeria monocytogenes and listerial virulence factors. Immunol. Res. 2008, 42, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Seavey, M.M.; Pan, Z.K.; Maciag, P.C.; Wallecha, A.; Rivera, S.; Paterson, Y.; Shahabi, V. A novel human Her-2/neu chimeric molecule expressed by Listeria monocytogenes can elicit potent HLA-A2 restricted CD8-positive T cell responses and impact the growth and spread of Her-2/neu-positive breast tumors. Clin. Cancer Res. 2009, 15, 924–932. [Google Scholar] [CrossRef]
- Shahabi, V.; Seavey, M.M.; Maciag, P.C.; Rivera, S.; Wallecha, A. Development of a live and highly attenuated Listeria monocytogenes-based vaccine for the treatment of Her2/neu-overexpressing cancers in human. Cancer Gene Ther. 2011, 18, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Gunn, G.R.; Zubair, A.; Peters, C.; Pan, Z.K.; Wu, T.C.; Paterson, Y. Two Listeria monocytogenes vaccine vectors that express different molecular forms of human papilloma virus-16 (HPV-16) E7 induce qualitatively different T cell immunity that correlates with their ability to induce regression of established tumors immortalized by HPV-16. J. Immunol. 2001, 167, 6471–6479. [Google Scholar] [PubMed]
- Sewell, D.A.; Pan, Z.K.; Paterson, Y. Listeria-based HPV-16 E7 vaccines limit autochthonous tumor growth in a transgenic mouse model for HPV-16 transformed tumors. Vaccine 2008, 26, 5315–5320. [Google Scholar] [CrossRef] [PubMed]
- Sewell, D.A.; Shahabi, V.; Gunn, G.R., 3rd; Pan, Z.K.; Dominiecki, M.E.; Paterson, Y. Recombinant Listeria vaccines containing PEST sequences are potent immune adjuvants for the tumor-associated antigen human papillomavirus-16 E7. Cancer Res. 2004, 64, 8821–8825. [Google Scholar] [CrossRef] [PubMed]
- Maciag, P.C.; Seavey, M.M.; Pan, Z.K.; Ferrone, S.; Paterson, Y. Cancer immunotherapy targeting the high molecular weight melanoma-associated antigen protein results in a broad antitumor response and reduction of pericytes in the tumor vasculature. Cancer Res. 2008, 68, 8066–8075. [Google Scholar] [CrossRef] [PubMed]
- Wallecha, A.; Maciag, P.C.; Rivera, S.; Paterson, Y.; Shahabi, V. Construction and characterization of an attenuated Listeria monocytogenes strain for clinical use in cancer immunotherapy. Clin. Vaccine Immunol. 2009, 16, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hou, J.; Lin, Z.; Zhuo, H.; Chen, D.; Zhang, X.; Chen, Y.; Sun, B. Attenuated Listeria monocytogenes as a cancer vaccine vector for the delivery of CD24, a biomarker for hepatic cancer stem cells. Cell. Mol. Immunol. 2014, 11, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Overdevest, J.B.; Knubel, K.H.; Duex, J.E.; Thomas, S.; Nitz, M.D.; Harding, M.A.; Smith, S.C.; Frierson, H.F.; Conaway, M.; Theodorescu, D. CD24 expression is important in male urothelial tumorigenesis and metastasis in mice and is androgen regulated. Proc. Natl. Acad. Sci. USA 2012, 109, E3588–E3596. [Google Scholar] [CrossRef] [PubMed]
- Senner, V.; Sturm, A.; Baur, I.; Schrell, U.H.; Distel, L.; Paulus, W. CD24 promotes invasion of glioma cells in vivo. J. Neuropathol. Exp. Neurol. 1999, 58, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Baumann, P.; Cremers, N.; Kroese, F.; Orend, G.; Chiquet-Ehrismann, R.; Uede, T.; Yagita, H.; Sleeman, J.P. CD24 expression causes the acquisition of multiple cellular properties associated with tumor growth and metastasis. Cancer Res. 2005, 65, 10783–10793. [Google Scholar] [CrossRef] [PubMed]
- Cummins, J.; Tangney, M. Bacteria and tumours: Causative agents or opportunistic inhabitants? Infect. Agent. Cancer 2013. [Google Scholar] [CrossRef] [PubMed]
- Panteli, J.T.; Forkus, B.A.; Van Dessel, N.; Forbes, N.S. Genetically modified bacteria as a tool to detect microscopic solid tumor masses with triggered release of a recombinant biomarker. Integr. Biol. (Camb.) 2015, 7, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Li, Y.; Li, Z.; Xie, X.; Lu, L. In vivo monitoring the process of tumor growth, metastasis and bacterial infection expressing GFP via real-time optical imaging. J. Biomed. Nanotechnol. 2013, 9, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Cronin, M.; Akin, A.R.; Collins, S.A.; Meganck, J.; Kim, J.-B.; Baban, C.K.; Joyce, S.A.; van Dam, G.M.; Zhang, N.; van Sinderen, D.; et al. High resolution in vivo bioluminescent imaging for the study of bacterial tumour targeting. PLoS ONE 2012, 7, e30940. [Google Scholar] [CrossRef] [PubMed]
- Riedel, C.U.; Monk, I.R.; Casey, P.G.; Morrissey, D.; O’Sullivan, G.C.; Tangney, M.; Hill, C.; Gahan, C.G.M. Improved Luciferase tagging system for Listeria monocytogenes allows real-time monitoring in vivo and in vitro. Appl. Environ. Microbiol. 2007, 73, 3091–3094. [Google Scholar] [CrossRef] [PubMed]
- Tjuvajev, J.G.; Avril, N.; Oku, T.; Sasajima, T.; Miyagawa, T.; Joshi, R.; Safer, M.; Beattie, B.; DiResta, G.; Daghighian, F.; et al. Imaging herpes virus thymidine kinase gene transfer and expression by positron emission tomography. Cancer Res. 1998, 58, 4333–4341. [Google Scholar] [PubMed]
- Brader, P.; Stritzker, J.; Riedl, C.C.; Zanzonico, P.; Cai, S.; Burnazi, E.M.; Ghani, E.R.; Hricak, H.; Szalay, A.A.; Fong, Y.; et al. Escherichia coli Nissle 1917 facilitates tumor detection by positron emission tomography and optical imaging. Clin. Cancer Res. 2008, 14, 2295–2302. [Google Scholar] [CrossRef] [PubMed]
- Soghomonyan, S.A.; Doubrovin, M.; Pike, J.; Luo, X.; Ittensohn, M.; Runyan, J.D.; Balatoni, J.; Finn, R.; Tjuvajev, J.G.; Blasberg, R.; et al. Positron emission tomography (PET) imaging of tumor-localized Salmonella expressing HSV1-TK. Cancer Gene Ther. 2005, 12, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Tjuvajev, J.; Blasberg, R.; Luo, X.; Zheng, L.M.; King, I.; Bermudes, D. Salmonella-based tumor-targeted cancer therapy: Tumor amplified protein expression therapy (TAPET) for diagnostic imaging. J. Control. Release 2001, 74, 313–315. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, I.Y.C.; Van, T.T.H.; Smooker, P.M. Live-Attenuated Bacterial Vectors: Tools for Vaccine and Therapeutic Agent Delivery. Vaccines 2015, 3, 940-972. https://doi.org/10.3390/vaccines3040940
Lin IYC, Van TTH, Smooker PM. Live-Attenuated Bacterial Vectors: Tools for Vaccine and Therapeutic Agent Delivery. Vaccines. 2015; 3(4):940-972. https://doi.org/10.3390/vaccines3040940
Chicago/Turabian StyleLin, Ivan Y. C., Thi Thu Hao Van, and Peter M. Smooker. 2015. "Live-Attenuated Bacterial Vectors: Tools for Vaccine and Therapeutic Agent Delivery" Vaccines 3, no. 4: 940-972. https://doi.org/10.3390/vaccines3040940
APA StyleLin, I. Y. C., Van, T. T. H., & Smooker, P. M. (2015). Live-Attenuated Bacterial Vectors: Tools for Vaccine and Therapeutic Agent Delivery. Vaccines, 3(4), 940-972. https://doi.org/10.3390/vaccines3040940