Abstract
Malaria vaccine research has been ongoing since the 1980s with limited success. However, recent improvements in our understanding of the immune responses required to combat each stage of infection will allow for intelligent design of both antigens and their associated delivery vaccine vehicles/vectors. Synthetic carriers (also known as vectors) are usually particulate and have multiple properties, which can be varied to control how an associated vaccine interacts with the host, and consequently how the immune response develops. This review comprehensively analyzes both historical and recent studies in which synthetic carriers are used to deliver malaria vaccines. Furthermore, the requirements for a synthetic carrier, such as size, charge, and surface chemistry are reviewed in order to understand the design of effective particle-based vaccines against malaria, as well as providing general insights. Synthetic carriers have the ability to alter and direct the immune response, and a better control of particle properties will facilitate improved vaccine design in the near future.
1. Introduction
The World Health Organization estimates that in 2013 there were 198 million cases of malaria; 584,000 of which were fatal [1]. The disease is prevalent in many tropical and subtropical regions of the world, most notably Sub-Saharan Africa, Asia, Latin America and the Middle East [2]. Africa carries the majority of the burden of lethal infections, which are most often mediated by the Plasmodium falciparum parasite strains. Vaccination preventing infection and subsequent transmission is the most likely strategy to reduce the spread and resurgence of the disease. Research into malaria vaccine candidates has been ongoing for many years, with limited progress. This is largely due to the complexity of the parasites’ life cycle, the lack of complete understanding of the body’s natural protective immunity to infection, and the genetic polymorphism of potential antigenic targets [3,4]. The strongest candidate, the recombinant circumsporozoite protein (CSP) based pre-erythrocytic stage RTS,S vaccine (MosquirixTM), has undergone phase III clinical trials and is likely to be licensed in 2015. However, efficacy results have varied from 30% to 50% in reduction rate of clinical episodes of malaria, with short lived and reduced protection in infants [5]. Therefore, further work is needed to develop a long lasting, efficacious vaccine against malaria. New generation attenuated whole parasite vaccines, recombinant technology, and nanotechnology are being actively explored to deliver such a vaccine [6,7]. Strongly immunogenic antigens with limited polymorphism are being discovered using immunomic techniques that allow for the analysis of many proteins identified in genomic data [8]. It is expected that future vaccine formulations will incorporate antigens against multiple stages of the malaria life cycle with improved adjuvants that are able to induce the desired types of protective immune responses.
Adjuvants can be defined as “any substance which when incorporated into a vaccine formulation acts generally to accelerate, prolong or enhance the quality of specific immune responses to vaccine antigens” [9]. Often the adjuvant effect relies on “danger signals” promoting the induction of inflammatory pathways [10]. Conversely, vaccine carriers or vectors (which can also have intrinsic adjuvant properties) are non-specific delivery systems to which recombinant protein, peptide or DNA antigens can be associated to promote their delivery to the antigen presenting cells (APCs) that then initiate an immune response [7]. Numerous carrier/vector characteristics such as size, shape, and surface charge, can be varied to alter the way the associated antigen is seen and processed by APCs and generally interacts with the immune system. This can in turn alter the strength and type of immune response. Furthermore, these properties allow for directing the immune response in a cellular or humoral direction, a practice considered especially important in malaria vaccine design [11].
Vectors historically used in clinical trials are based on attenuated organisms such as genetically altered viruses [5]. Diversification in recent years has seen the development of a wide range of synthetic particles including organic carriers such as liposomes and polymeric microspheres, and inorganic particles based on materials including iron oxide and calcium phosphate [12,13,14,15]. This review will focus on the requirements for a synthetic vaccine vector, both in general and for malaria specifically. It will then examine the properties that make micro- and nanoparticles attractive as vectors, and summarize the literature in which malaria vaccines are delivered with non-conventional vectors with respect to the aforementioned requirements.
2. Vaccine Carrier/Vector Requirements
Historically, successful vaccines have been developed using attenuated and killed organisms. For example, measles and mumps are still prevented using attenuated living viruses while polio vaccines based on both attenuated and inactivated poliovirus have been successful [16,17,18]. Whilst these vaccines have led to a vast reduction in the prevalence of disease, there are those for which progress in vaccine development has been limited, most notably HIV and malaria. In recent years, vaccine research has broadened with subunit and DNA vaccines seeing expanded development at the expense of attenuated and inactivated organism based approaches. Subunit vaccines contain antigens (peptide or protein) from the organism that are the target of specific immune responses [19]. DNA vaccines use genetic material that encodes for the antigens of interest. This material is internalized by cells in the body, which will then process and express the encoded protein. The displayed protein induces dendritic cell (DC) activation and subsequent adaptive immune responses [20]. The major advantages of subunit and DNA vaccines are their inherent safety, the ability to tailor their immune responses, simpler storage and transport requirements and easier development/production [21]. The main drawback of subunit vaccines is their weak immunogenicity, often necessitating advanced delivery systems and/or adjuvant use [22]. Similarly, naked DNA vaccines are quickly removed by the body and hence require delivery vectors. In addition, the immunogenicity evident in mice is often not maintained in humans when the DNA vaccine is tested clinically [23]. This has led to a focus on better vectors for improved cell transfection. Looking towards the future, it seems highly likely that subunit and DNA vaccines will continue to dominate research efforts. Thus, it is important that vectors are developed concurrently to improve their efficacy in a safe, affordable manner.
A successful vector will need to meet a number of requirements for clinical use. It should be designed to enhance and complement the natural response to the vaccine. In addition, vaccine formulations should be stable for extended periods of time and amenable to transport and storage in ambient conditions which are vital in underdeveloped regions [24]. For the same reason, it would be preferable if vaccines could be delivered via a single injection, without the need for secondary and tertiary administration. Production of vaccine vectors needs to be affordable, robust, and scalable. Furthermore, the need to manufacture multiple components (vaccine and vector) will increase costs compared with existing systems [25]. Lastly, and most importantly, a vector needs to be safe for human use, in terms of toxicity, acute and chronic side effects, biocompatibility, and biodegradability, especially for nanoparticle use in vaccines [26]. For particles in particular, size and surface reactivity can allow for diffusion to potentially susceptible areas such as the lungs and brain. Toxicity studies should be undertaken at the preclinical stage and in vivo distribution needs to be mapped.
3. Malaria Vaccine Requirements
The malaria parasite interacts with and affects its host in a distinct manner at each stage of its life cycle (Figure 1). Thus, in order to target each stage of infection (i.e., prevention of cell invasion versus elimination of infected cells), different strategies are required. The problem is exacerbated in that each stage expresses different surface proteins requiring specific responses from the vaccine. The goal of any pre-erythrocytic vaccine is to prevent parasites exiting the liver; for example anti-sporozoite vaccines stop sporozoites infecting the liver [27]. A vaccine targeting sporozoites needs to induce high antibody titers with strong avidity to surface antigens to either prevent the parasite leaving the injection site, or inhibit liver infection [28]. In addition, long lived plasma cells and memory cells may be required to maintain immunity over a long period of time [29]. The molecular mechanisms of hepatocyte invasion are unknown, though a number of proteins thought to be important to the process have been investigated. The CSP is the most abundant surface protein and has been implicated in motility and cell invasion [30,31]. Other identified targets for pre-erythrocytic candidate vaccines include thrombospondin-related anonymous protein (TRAP) which plays roles in motility and invasion, apical membrane antigen 1 (AMA1) involved in hepatocyte invasion and cell traversal protein (CelTOS), and sporozoite microneme protein essential for cell traversal (SPECT), part of the cell traversal mechanics [32,33,34,35,36]. As well as blocking hepatocyte invasion, pre-erythrocytic stage vaccines can also be designed to prevent merozoites exiting into the blood stream by inhibiting parasite maturation inside the liver cells [28]. Activation of sporozoite-specific CD8+ T cells promotes their expansion, cytotoxicity, and cytokine secretion (importantly IFN-γ and TNF-α) [37,38]. These cells then kill infected cells exhibiting the antigen their ancestors initially responded against. Sporozoite antigens such as CSP and TRAP are still exhibited after hepatocyte invasion, along with liver-stage antigens, which can be targeted by vaccines. These include liver-stage antigen 1 (LSA1), involved in liver-stage differentiation, liver-stage antigen 3 (LSA3), and exported protein 1 (Exp1), a membrane protein [39,40]. The use of antigens expressed only in the liver stage is attractive as they are often highly conserved [40,41]. Extremely high cytotoxic T cell responses are needed to combat liver-stage infection. Therefore, a vaccine system needs to target DCs capable of cross-priming into MHC class I pathways and elicit high levels of CD8+ T cells, as well as a supporting CD4+ helper T cell type 1 (Th1) response, which generates the high levels of IL-2 production necessary to sustain CD8+ T cell expansion [42,43].
Blood stage vaccines aim to inhibit erythrocyte invasion and/or parasite reproduction, as well as opsonize infected cells for macrophage removal [28]. A vaccine that meets these requirements will induce high antibody titers with a high affinity for their substrate, cytokine production for phagocyte activation and generation of long-lived plasma and B and T memory cells. Evidence suggests that high antibody titers may be required for protection, and that in mice, natural killer (NK) cells as well as B and T cells play important roles [44,45,46]. Specifically NK and T cell derived IFN-γ promote efficient splenic macrophage activation, and hence parasite clearance. Most individuals living in malaria endemic regions show antibody responses to a large number of Plasmodium blood stage proteins [47]. While this provides potential antigenic targets for vaccines, the pressure placed on the parasite has led to a high degree of allelic variation [48,49]. In many cases this reduces vaccine efficacy throughout a population and promotes further polymorphic immunity to the vaccine. The antigens commonly investigated include merozoite surface protein 1 (MSP1) which is likely to be involved in erythrocyte attachment, merozoite surface protein 2 (MSP2) and merozoite surface protein 3 (MSP3) whose functions are unknown but thought to be essential, and AMA1 and erythrocyte binding antigen 175 (EBA175) which are involved in blood cell invasion [50,51,52,53]. Recent work has focused on Plasmodium falciparum reticulocyte binding-like homologous protein 5 (PfRH5) due to its highly conserved nature [54]. This is exciting as the antigen is thought to play an essential role in erythrocyte invasion [55].
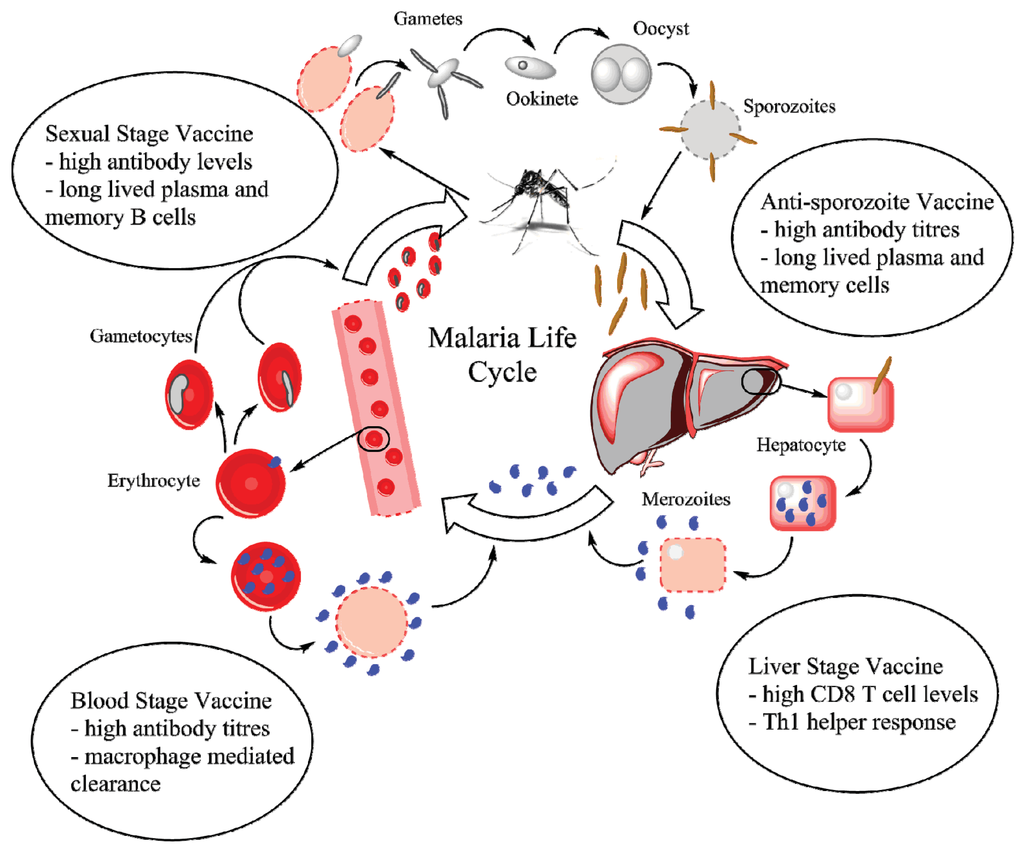
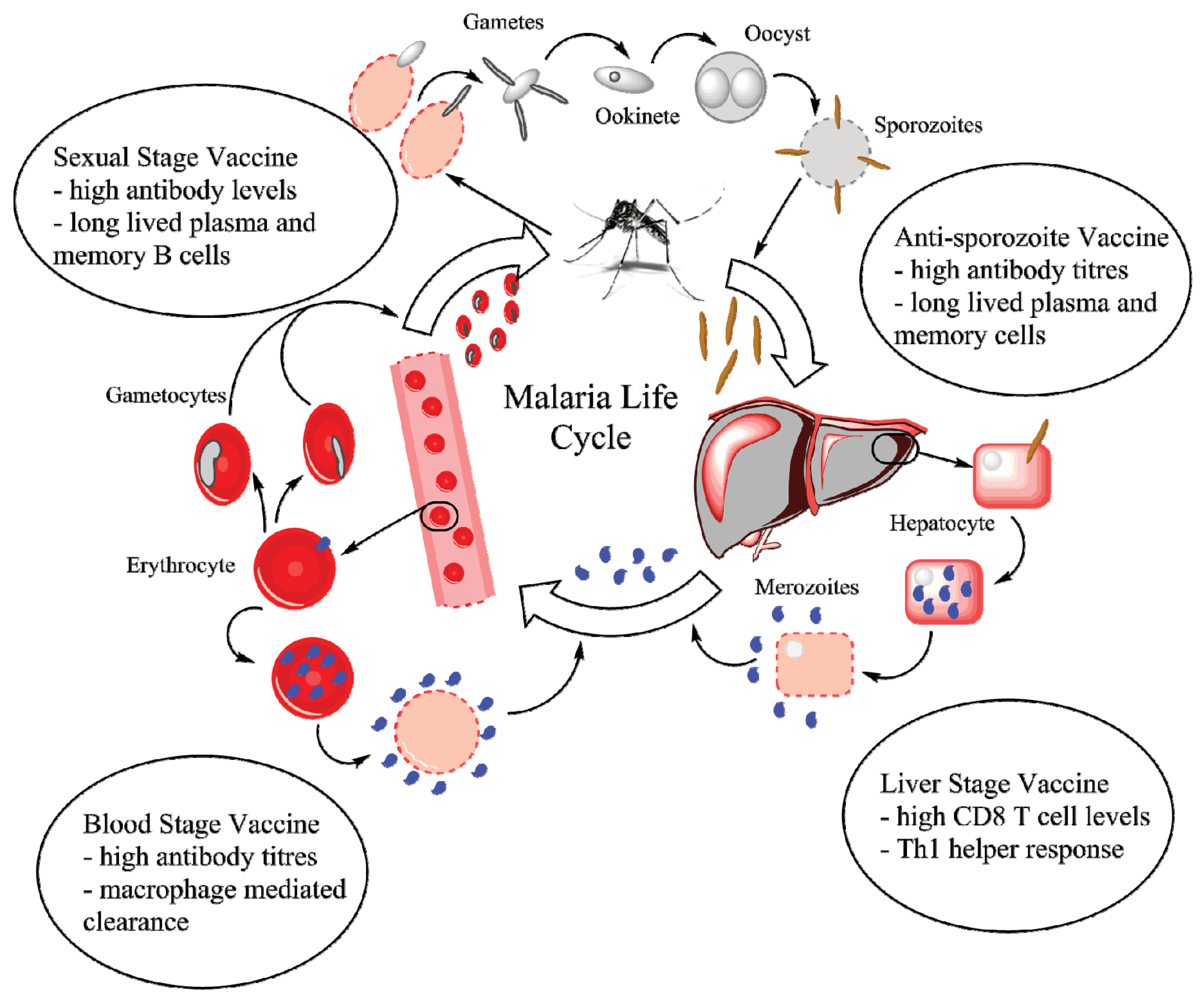
Figure 1.
The life cycle of the malaria parasite. A mosquito delivers sporozoites into the blood stream, which quickly travel to and infect hepatocytes. Inside liver cells, the sporozoites differentiate and proliferate to form merozoites, which upon release infect erythrocytes. Proliferation and release continues inside infected erythrocytes with a small fraction of merozoites differentiating into gametocytes. The gametocyte-infected erythrocytes are taken up by mosquitoes inside which gametes interact to form a zygote (ookinete) and eventually an oocyst, which produces sporozoites. The bubbles contain the primary immune response required against each stage. Figure developed using ChemBioDraw Ultra 14 (PerkinElmer Informatics, Waltham, MA, USA, 2014).
Figure 1.
The life cycle of the malaria parasite. A mosquito delivers sporozoites into the blood stream, which quickly travel to and infect hepatocytes. Inside liver cells, the sporozoites differentiate and proliferate to form merozoites, which upon release infect erythrocytes. Proliferation and release continues inside infected erythrocytes with a small fraction of merozoites differentiating into gametocytes. The gametocyte-infected erythrocytes are taken up by mosquitoes inside which gametes interact to form a zygote (ookinete) and eventually an oocyst, which produces sporozoites. The bubbles contain the primary immune response required against each stage. Figure developed using ChemBioDraw Ultra 14 (PerkinElmer Informatics, Waltham, MA, USA, 2014).
The goal of sexual stage vaccines (transmission blocking vaccines) is to prevent gametocyte uptake by the mosquito and/or to inhibit parasite development within the vector [28]. This would be solely antibody based, with gametocyte, gamete or ookinete antigens targeted to inhibit fertilization or migration. In addition, antibodies can mediate gamete lysis through the human complement system [56]. For antigen which is only expressed on the parasite surface during mosquito inhabitation, long-lived plasma cells and memory B cells would be especially important, as the lack of re-exposure would prevent natural boosting of the response upon infection [57]. A sexual stage vaccine would present no benefit to the vaccinated person and would instead reduce the spread of malaria over the whole population. The most commonly investigated antigens are Plasmodium falciparum surface protein 25 (Pfs25) and Plasmodium vivax surface protein 25 (Pvs25). These proteins are expressed on the surface of the parasites zygote and ookinete stages [58]. Other antigens being investigated include Plasmodium vivax surface protein 230 (Pvs230), Plasmodium falciparum surface protein 230 (Pfs230), and Plasmodium falciparum surface proteins P48/45 (Pfs48/45). These are expressed by the gametocyte and gamete stages and can therefore rely on natural immune boosting [59,60,61]. Vectors for this stage have been suggested to need to promote strong T follicular helper (Tfh) and Th2 responses.
4. Particle Use in Malaria Vaccine Development
This section will highlight a number of historical and recent malaria studies in which a wide range of different materials have been used as particle-based vectors for recombinant protein, peptide, or DNA encoding malarial antigens (summarized in Table 1). Although we have attempted to be comprehensive, we apologize to the authors of papers that may have been inadvertently omitted. There is great variability in these studies in terms of antigenic targets, particle types (highlighted in Figure 2), particle properties, and study goals. However, together they provide useful specific insights into the state of this active and exciting field of research, which will be discussed by categorizing the studies into broad classes based on the type of immune response elicited, the malarial stage they target, and the particle type, to identify and highlight the most informative results.
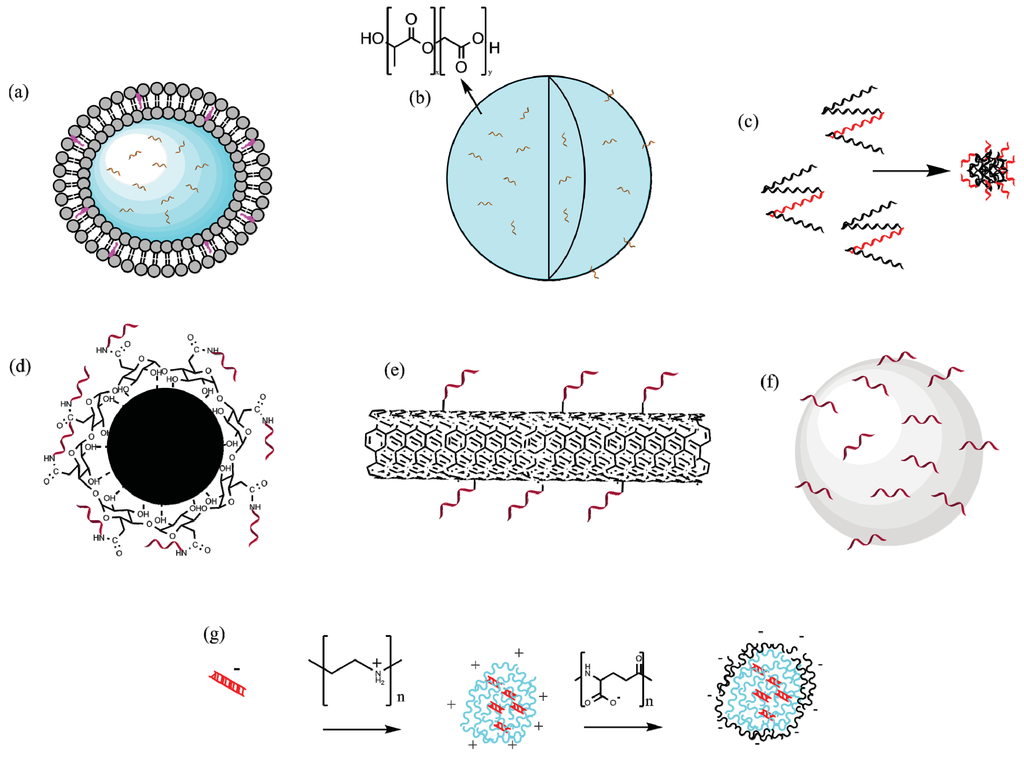
Figure 2.
A selection of different particle types that have been used as vectors for malaria vaccines. Note the different vaccine types and methods of incorporation. (a) PC liposomes, the antigen is encapsulated inside an aqueous compartment; (b) PLGA particles, antigen encapsulated both in and on the surface of particle; (c) Self assembled protein nanoparticles, peptide incorporated into protein to enable repetitive display; (d) Iron oxide nanoparticle, antigen covalently conjugated to polymeric coating; (e) Carbon Nanotube; (f) Hydroxyapatite particle, antigen adsorbed to the surface; (g) PEI/PGA nanoparticle construct, DNA vaccine electrostatically bound to PEI. Figure developed using ChemBioDraw Ultra 14.
Figure 2.
A selection of different particle types that have been used as vectors for malaria vaccines. Note the different vaccine types and methods of incorporation. (a) PC liposomes, the antigen is encapsulated inside an aqueous compartment; (b) PLGA particles, antigen encapsulated both in and on the surface of particle; (c) Self assembled protein nanoparticles, peptide incorporated into protein to enable repetitive display; (d) Iron oxide nanoparticle, antigen covalently conjugated to polymeric coating; (e) Carbon Nanotube; (f) Hydroxyapatite particle, antigen adsorbed to the surface; (g) PEI/PGA nanoparticle construct, DNA vaccine electrostatically bound to PEI. Figure developed using ChemBioDraw Ultra 14.
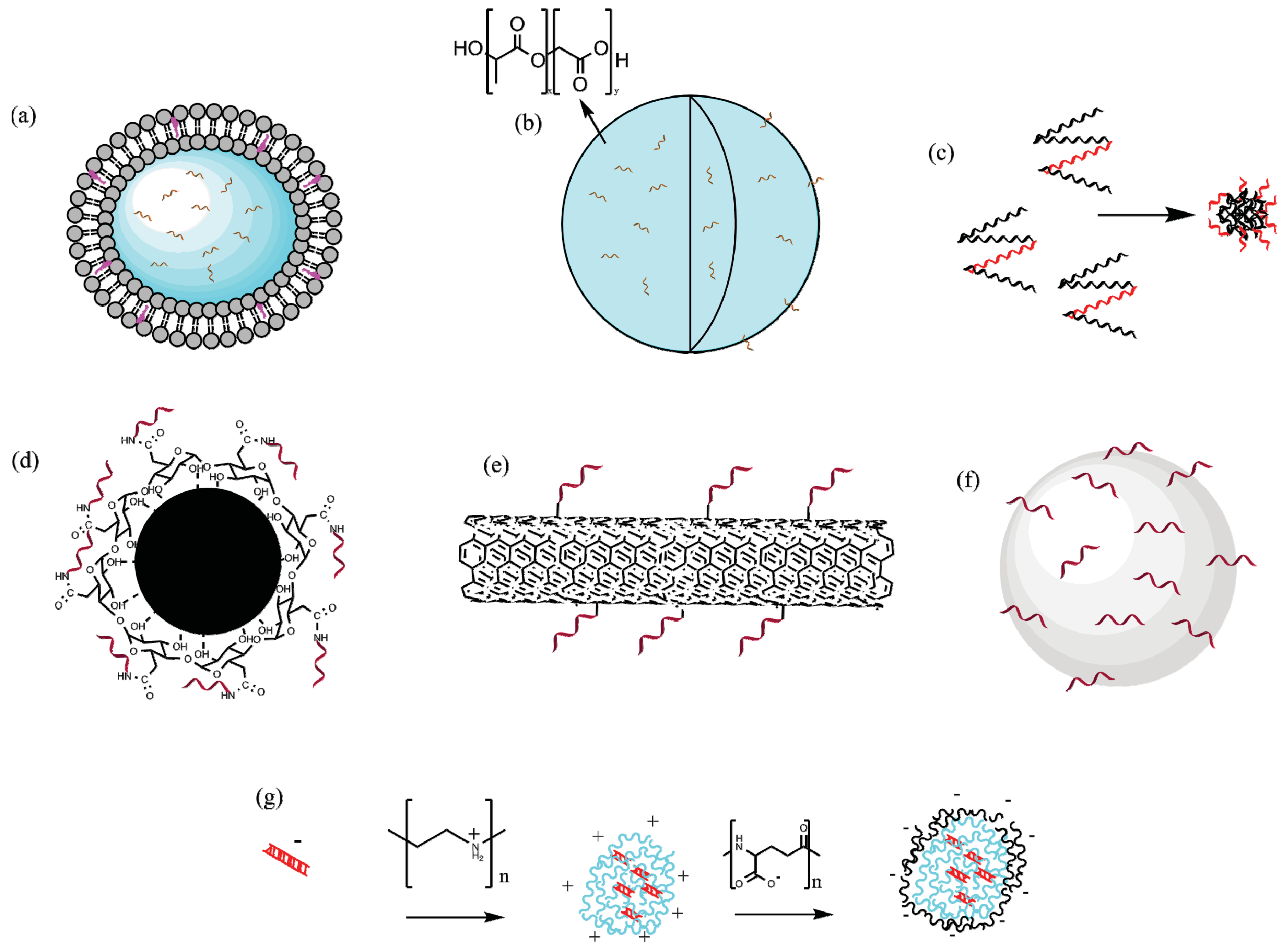

Table 1.
Summary of clinical and preclinical studies incorporating novel synthetic vector strategies.
| Vector | Size | Vaccine | Antibodies | Cytotoxic Response | Ref. |
|---|---|---|---|---|---|
| Pre-Erythrocytic Candidates | |||||
| Lipid Based Particles | |||||
| PC 1 liposomes | Not reported | PyCSP 2 (tetrapeptide B cell, other B, T cell epitopes, RTS,S) | Yes | Some antigens | [12,62,63,64,65,66,67,68,69] |
| Lipid core peptides | Not reported | PyCSP (CD4 and CD8 epitopes) | Minor IgE | ND 3 | [70] |
| ICMV 4 | 180 nm (DLS 5) | PvCSP 6 (VMP001) | Yes | ND | [71] |
| Polymeric Particles | |||||
| PCL 7/PLA 8 | 23–45 µm | PfCSP 9 (tetrapeptide, universal CD4) | Yes | ND | [72] |
| PLA/PLGA 10 | 1–100 µm | PfCSP (tetrapeptide, universal CD4), Pb911 | Yes | Against Pb9 | [13,73,74] |
| PLGA | 0.45–32.1 µm | Pb9 | ND | Yes | [75] |
| Lipid enveloped PLGA | 290 nm (DLS) | PvCSP (VMP001) | Yes | ND | [76] |
| Polystyrene Nanoparticles | 48 nm (DLS) | Pb9 | ND | Yes | [77] |
| Other Particles | |||||
| SAPN 12 | 40 nm (TEM 13, DLS) | PfCSP, PvCSP, CD8 and B epitopes, universal CD4 | Yes | Some antigens | [78,79,80] |
| Polymer coated calcium carbonate | Not reported | PfCSP (B and T cell epitopes) | Yes | Yes | [81] |
| Blood Stage Candidates | |||||
| Lipid Based Particles | |||||
| PC Liposome | Not reported | PfMSP1–19 14 | Yes | ND | [82] |
| PC Liposome | Not reported | PyIMP-66 15 | Yes | ND | [83] |
| E. coli 16 Liposome | Not reported | Py soluble | Yes | Yes | [84] |
| S. cer. 17 Liposome | Not reported | Py soluble | Yes | Yes | [85] |
| pH sensitive Liposome | 325–390 nm | PfMSP1–19 | Minor | ND | [70] |
| Other Organic Particles | |||||
| PEI 18, γ-PGA 19 | 68 nm (DLS) | PyMSP1-C-terminus, PyTAM 20 (DNA) | Yes | With PyTAM | [86,87,88] |
| ISCOM 21 | Not Reported | PfRESA 22 | Yes | ND | [89,90] |
| ISCOM | Not Reported | PfRESA peptides | Yes | ND | [91] |
| Inorganic Particles | |||||
| SPIONs 23 | 20 nm (unknown) | PfMSP1–42 | Yes | ND | [14] |
| PEI coated SPIONs | 147 nm (DLS) | PyMSP1–19 (DNA) | Yes | ND | [92] |
| Gold nanoparticle | 17 nm (TEM) | Pf/PvMSP1–19 | Yes | ND | [93] |
| Quantum Dots | 15 nm (unknown) | PfMSP1–42 | Yes | ND | [94] |
| Carbon Nanotubes | 20–30/500–2000 nm | PvAMA1 4peptides | Yes | ND | [95] |
| Hydroxyapatite | 784 nm (DLS) | MSP1–19 | Yes | ND | [15] |
| Mixed Stage Candidates | |||||
| PLGA | 0.5–2 µm | SPf66, PfMSP2 25 peptides, PfS3 | Yes | ND | [96,97,98,99,100,101,102] |
| PLGA-alginate-RGD | 0.8–1 µm | SPf66, PfS3 | Yes | ND | [103] |
| PLGA | 1–2 µm | PvCSP, MSP1, AMA1, Pvs24 (all with B and T epitopes) | Yes | ND | [104,105] |
| Sexual Stage Candidate | |||||
| Gel core liposomes | 1–1.2 µm | Pfs25 | Yes | ND | [106] |
1: Phosphatidylcholine; 2: Plasmodium yoelii circumsporozoite protein; 3: Not Done; 4: Interbilayer-crosslinked multilamellar vesicles; 5: Dynamic light scattering; 6: Plasmodium vivax; 7: Polycaprolactone; 8: Poly(lactic acid); 9: Plasmodium falciparum; 10: Poly(lactic-co-glycolic acid); 11: Plasmodium berghei; 12: Self-Assembling protein nanoparticle; 13: Transmission electron microscopy; 14: Merozoite Surface Protein 1; 15: Integral Membrane Protein; 16: Escherichia coli; 17: Saccharomyces cerevisiae; 18: Polyethyleneimine; 19: γ-polyglutamic acid; 20: PyGPI8p-transamidase-related protein; 21: Immune stimulating complex; 22: Ring-infected erythrocyte surface antigen; 23: Superparamagnetic iron oxide nanoparticles; 24: Apical Membrane Antigen 1; 25: Merozoite Surface Protein 2.
4.1. Pre-Erythrocytic Vaccines
Anti-sporozoite vaccines attempt to produce antibodies, which agglutinate or otherwise inactivate sporozoites such that they cannot infect hepatocytes. On the other hand, liver-stage vaccines require strong cytotoxic T cell responses to destroy infected cells [28]. Therefore, synthetic particle vector studies focused on meeting either one or both of these requirements are discussed below. A summary of the advantages and disadvantages of the different particle types used is given in Table 2.
Table 2.
Comparison of advantages and disadvantages of different nanoparticle types.
| Particle Type | Advantages | Disadvantages | Ref. |
|---|---|---|---|
| Lipid-Based | Wide size range Antigen encapsulated or on surface Hydrophobic or hydrophilic antigen FDA approved/Non-toxic Biodegradable | Expensive materials Reproducibility issues Oxidative Degradation | [107,108,109,110] |
| PLGA | Antigen encapsulated or on surface Biodegradable FDA approved/Non-toxic Prolonged release of antigen | Antigen degradation Scale-up Antigen burst releases | [110,111,112,113,114] |
| Polystyrene | Biocompatible Non-toxic Wide size range Readily available | Non-biodegradable | [115] |
| SAPN | Repetitive presentation Biodegradable | Complex design Limited Data | [116] |
| PEI/γ-PGA | Good for DNA vaccine Small size | Limited Data | [117] |
| ISCOM | Natural adjuvant Readily available Biodegradable Scalable Well-tolerated | Encapsulation limited Single size | [118,119,120] |
| SPION | Biodegradable Magnetic FDA approved Size control | Coating required Stability issues | [121,122] |
| Quantum Dot | Fluorescent Stable | Toxic materials Non-biodegradable | [123,124,125] |
| Calcium Based | Low cytotoxicity Surface modification | Limited degradability Limited study | [126,127] |
| Gold | Size control Low cytotoxicity | Non-biodegradable Coating required | [128,129] |
4.1.1. Lipid-Based Particles
Lipids are naturally occurring biological molecules whose properties can allow them to form structures in a suitable environment. Liposomes, the most of common of these, are vesicles formed when one or more lipid bilayers enclose an aqueous compartment [130]. Their use as drug delivery agents stretches back to the 1960s with research incorporating malaria antigens beginning in the 1980s [62,131]. Sizes range between 30 nm and several microns, and can be varied alongside lipid composition and surface charge to optimize their carrier potential [132]. For instance, incorporation of PEG into the lipid surface enhances the distribution time in blood [133]. Antigen is commonly encapsulated in the aqueous center of the liposome, with surface conjugation used to a lesser degree [134]. Advantages include the ability to incorporate both hydrophilic and hydrophobic antigens, and the systems’ biodegradability [107]. The safety of liposomes is evidenced by the number of clinically approved drugs in which they are incorporated. Challenges include the use of expensive materials, reproducibility issues, and the potential for oxidative degradation during storage and resultant instability [108,109,110]. In the late 1980s and early 90s, research into the use of liposomal vectors for the delivery of CS antigens culminated in multiple clinical trials. Early results with mice, rabbits, and monkeys demonstrated that antibodies specific to the CS protein (tetrapeptide B cell epitopes) were induced when it was encapsulated in dimyristoyl phosphatidylcholine/cholesterol (PC/Chol) liposomes with responses increasing upon incorporation of the adjuvant lipid A [62,63,64]. Though protection was not analyzed in these reports, a clinical trial using the liposome/antigen/lipid A system adjuvanted with alum was initiated based on the antibody results [12]. The IgG antibody response was greater than those obtained with alum in previous trials. At the highest doses sera inhibited hepatoma invasion by an average of 92%, potentially insufficient to prevent clinical symptoms developing. An anti-sporozoite vaccine needs to completely prevent hepatocyte infection to halt disease progression as any breakthrough infections can lead to blood stage infection. This demonstrates the benefits of having a vaccine that can target both the sporozoite and liver stages of malaria and the reasoning behind multi-stage vaccine development.
Non-liposomal lipid based particles have engendered little interest as malaria vaccine carriers. Moon et al. reported the use of interbilayer-crosslinked multilamellar vesicles (ICMVs) with a Plasmodium vivax CSP antigen, VMP001 [71]. ICMVs are described as nano-sized multilayered lipid vesicles with covalent cross-links between layers [135]. 180 nm particles with antigen both encapsulated and anchored to particle membranes were synthesized and induced a 9-fold greater immune response than particles lacking surface bound antigen [71]. This emphasizes the importance of controlling particle properties, as minor changes can have unexpected effects. When adjuvanted with monophosphoryl lipid A (MPLA), the antibodies targeted multiple epitopes with higher avidity than those produced with soluble protein and produced a balanced T helper response. The particles collected in draining lymph nodes, and the authors concluded that the enhanced, long lived humoral response was due to increased germinal center formation in close proximity to these particle depots and increased CD4+ T cell expansion and differentiation to antigen specific Tfh cells. The authors obtained similar results against the same antigen with 290 nm lipid enveloped poly (lactic-co-glycolic acid) (PLGA) particles [76]. These studies incorporate multiple features, which mark particles as attractive vectors. The ability to control the type of response, to target multiple epitopes, and to induce a long-lived response are hallmarks of successful vaccine design against malaria. Furthermore, enhanced germinal center formation in multiple studies suggests that either the properties of the particles or the lipids composing them are responsible. This provides an avenue for the induction of immunological memory and plasma B cells upon injection, important in the development of a long-lived response.
4.1.2. Polymeric Particles
The descriptor “polymeric particles” encompasses a wide range of structures and conformations. These include natural and synthetic spheres (solid and hollow), nanogels, polymeric micelles, and dendrimers [7,136]. Among these candidates, only particles derived from polyesters and polyamino acids have seen in-depth exploration as malaria vectors, with the focus mainly on the polyesters, polylactic acid, and PLGA. Both solid and hollow particles ranging in size from about 100 nm to several microns have been used in drug and vaccine delivery [96,137]. Antigen/DNA can be encapsulated within the hollow core of a capsule, in the body of a sphere, or attached to the surface of either. A combination of the two is usually present [138]. PLGA is commonly used because both its monomers; lactic acid and glycolic acid, are biodegradable and easily dealt with by the body [111]. Its limited toxicity is further evidenced by incorporation in products approved by the FDA [138]. Targeting ligands can be attached to the surface to enable improved cellular binding and internalization [139]. For example, mannan has been covalently linked to PLGA particles to target C-type lectin receptors (CLRs) on DCs [140]. Other advantages include the ability to prolong the release of antigen and the co-encapsulation of adjuvants [112,113]. A potential problem is the loss of antigen immunogenicity either during synthesis or in the acidic environment formed upon degradation [110,114]. There have also been issues with scale-up and burst releases; large losses of antigen resulting from weak surface adsorption [141]. Thomasin et al. used a B cell CS epitope with universal T helper epitopes from tetanus toxins to synthesize a synthetic antigen, P30B2 [13]. This was loaded in PLGA microparticles with varying molecular weights and lactic: glycolic acid ratios. These properties allowed for control of the rate of antibody production via degradation, with overall titers longer lasting and of similar strength to incomplete Freund’s adjuvant (IFA) for all formulations. The same authors later found that combining microspheres of three different compositions and sizes induced a stronger antibody response to synthetic peptides than any singular particle, with said response similar in intensity and subclass distribution to peptides adjuvanted with IFA [73].
Extremely strong cytotoxic T cell responses are required to combat sporozoite infected hepatocytes and as such a number of studies have incorporated mixtures of B cell and CD8+ T cell epitopes in hopes of eliciting greater overall protection. Men et al. (1997) designed a system using six repeats of the NANP P. falciparum B cell epitope linked to universal tetanus toxin T helper epitopes and the cytotoxic epitope Pb9 in hopes of inducing cytotoxic T cell responses [74]. Chromate assays were used to demonstrate that the CD8+ T cell response was equivalent regardless of whether the peptides were encapsulated or injected with free particles. This suggested that the presence of the polymer was enough to improve MHC class I presentation. Using the same P. berghei epitope conjugated to 50 nm non-inflammatory polystyrene nanoparticles, Wilson et al. induced CD8+ T cell responses comparable to those obtained using montanide [77]. These responses were not developed when the antigen was not directly conjugated to the particle prompting the author to suggest that in vivo delivery of the peptide is vital in immune response development.
4.1.3. Self-Assembling Protein Nanoparticles (SAPNs)
The relatively novel Self-Assembling Protein Nanoparticles (SAPNs) are composed of peptide chains deliberately designed to self-assemble in the right environment to form particles of a certain size and shape [116]. Similar to virus-like particles, they have the ability to repetitively present antigen in an ordered array [142]. This method of display has been linked to increased antibody affinity and suggests potential for strong immunogenicity [143]. Large scale production is possible but no data could be found on the potential toxicity of the constructs [144]. Kaba et al. used an immunodominant B cell epitope from CS to create 25 nm particles [78]. These were able to protect 60% of mice for six months upon initial challenge or 15 months upon secondary challenge when administered in both saline and montanide incomplete Seppic adjuvant 720 (ISA-720). This protection was associated with strong antibody titers, and using serum transfer was shown to be antibody dependent. It was further demonstrated using strains of mice deficient in CD4+ T cells that these cells were necessary for protection. The authors claimed that a sequence in the peptide was aiding DC maturation and enabling T cell activation, rendering adjuvants unnecessary. Later work by this group involved incorporation of PfCSP and PvCSP B and T cell epitopes into their SAPN [79]. A total of 90%–100% of mice were protected with just B cell epitopes incorporated into the SAPN constructs. After a year, 50% of mice were still protected. Using knockout mice and serum/splenocyte transfer the authors demonstrated that partial protection was obtained via both antibody mediated and CD8+ T cell mediated responses. This proved that both components of the system acted in concert to protect the animals. The authors later worked to define the mechanisms by which these responses developed [80]. They demonstrated inhibition of both sporozoite motility and hepatocyte invasion and growth, and linked this to sporozoite lysis via the classical pathway of complement activation. It was further shown that SAPNs are localized with TAP for prolonged periods of time in early endosomes, which is indicative of antigen cross presentation.
4.1.4. Inorganic Particles
Inorganic particles have been used in a limited number of studies. This is generally a result of their lack of degradability and potentially toxic material components. Calcium carbonate is one such material. It has limited biodegradability and was incorporated with the polymers poly-l-lysine and poly-l-glutamic acid to form microparticle constructs loaded with a CS peptide containing a B and two T cell epitopes [81]. They demonstrated that a mixed antibody and cellular response developed, and that cellular immunity alone could not provide protection. The latter is a relatively common theme in the reviewed projects and is evidence that the immune responses against malaria vaccines are rarely simple with the concurrent development of humoral and cellular responses preferable. The differences between these studies limit the conclusions that can be drawn, but it is clear that particles do alter the immune response. The mechanisms by which each does so may differ depending on the particles properties but further investigation is needed to discover how.
4.2. Blood Stage Vaccines
It is thought that a good blood stage vaccine requires strong antibody production along with high levels of IFN-γ production to promote macrophage removal of infected cells [28]. A range of different particles have been used to meet these requirements and the most interesting are examined here.
4.2.1. Lipid Based Particles
Sharma et al. entrapped soluble blood stage antigens inside both PC liposomes and those composed of lipids extracted from E. coli [84]. In contrast to the conventional liposomes, those developed from E. coli elicited CD8+ T cell production with 45% specific lysis of target cells compared to 2% with the PC liposomes. This was credited to the ability of the E. coli lipids to fuse with APCs and deliver antigens directly into the cytosol. These liposomes also induced greater CD4+ T cell responses with strong IL-2 and IFN-γ up-regulation and enhanced antibody titers. The importance of these responses was further evident in the superior survival rates (70% vs. 30%) of the mice following challenge. The authors obtained similar results when they compared liposomes synthesized using lipids from edible yeast Saccharomyces cerevisiae and PC liposomes, encapsulating the same antigens used previously [85]. This indicated development of both Th1 and Th2 helper cells which led the authors to conclude that both are involved in the control of the blood stage infection. The enhanced protection evident in these studies almost certainly results from increased antibody and Th1 development, though this may be aided by CD8+ T cell proliferation and associated cytokine production.
4.2.2. Immune Stimulating Complexes (ISCOMs)
Immune Stimulating Complexes (ISCOMs) are so named for their ability to stimulate both humoral and cellular immune responses. They are spherical cage-like structures, around 40 nm in diameter, which form spontaneously from mixtures of saponin, cholesterol, and phospholipid [118]. Encapsulation has generally been limited to antigens which display amphipathic behavior, though this requirement has been relaxed somewhat with the development of ISCOMATRIX™ [145,146]. ISCOMATRIX™ differs from ISCOMs in that antigen is not incorporated during the synthesis of the structure but added afterwards to create the vaccine formulation [120]. Its successful use has also demonstrated the reproducibility and scalability of the technology. Human trials of saponin-based vaccines have seen them well tolerated [119]. Chopra et al. encapsulated two different ring-infected erythrocyte surface antigen (RESA) peptides by conjugating them to palmitic acid, creating an amphipathic molecule [91]. Both peptides elicited significantly (p < 0.001) higher IgG titers than alum adjuvanted antigen in the four strains of mice tested. Th1 biased responses were favored based on strong IgG2a and IgG2b levels with the authors suggesting that the bias towards these kinds of responses could be controlled by peptide choice.
4.2.3. Inorganic Particles
The only magnetic nanoparticles investigated as vaccine vectors are superparamagnetic iron oxide nanoparticles (SPIONs). These particles range from 1–30 nm and, as with many inorganic nanoparticles, exhibit unusual properties which make them attractive for different applications [147]. SPIONs have a single magnetic domain and can therefore be exposed to a magnetic field without remanence [148]. This has led to their use in magnetic targeting. For example, they have been magnetically concentrated in tumors and used as MRI contrast agents, situations in which organic particles are unsuitable [149,150]. Contrast agents based on SPIONs have been approved by the FDA, evidence of both their large scale capabilities and acceptable safety profile [121]. In contrast to most inorganic particles, they are biodegradable and readily excreted from the body [122]. Aggregation and stability problems are common in solution, though surface functionalization has mitigated this to some degree. This can take the form of polymeric or inorganic coating and/or ligand attachment (also allowing for in vivo targeting) [151]. Pusic et al. conjugated a recombinant MSP1-42 fragment to carboxyl groups on the outside of SPIONs with an amphiphilic polymer coating [14]. The size quoted was less than 20 nm, though the method of measurement was not clear and consequently is difficult to correlate with other studies. Intraperitoneal and intramuscular administration elicited responses from a greater percentage of mice than subcutaneous injection and sera from immunized mice inhibited parasite growth to a greater degree. Inhibition also exceeded that induced via protein administration with Freund’s adjuvant (FA) or ISA-51, potentially due to preferential induction of antibodies specific for inhibitory epitopes. Efficient uptake of the particles (without antigen) by APCs was demonstrated in vitro with expression of pro-inflammatory cytokines (TNF-α, IL-6) enhanced in DCs but not macrophages. In a similar study with quantum dots, the authors found that antibodies elicited against the conjugated peptide were significantly greater than those with FA or ISA-51 [94]. A more balanced Th1/Th2 response was obtained with the quantum dots with the response against SPIONs Th2 biased. Once again this highlights the importance of particle properties, in this case the material of the carrier, in determining the response against conjugated peptide. It should be noted that quantum dots contain heavy metals such as cadmium and zinc which are toxic to the human body [123,124]. This combined with their lack of biodegradability means that they can be only be considered as a model for nanoparticles in vaccine use.
4.2.4. DNA Vaccine Carriers
Few studies have used particles as vectors for malarial DNA vaccines. This is often due to difficulties encountered with encapsulation and degradation [152]. Al-Deen et al. electrostatically conjugated a plasmid vaccine based on the PyMSP119 DNA fragment to the surface of polyetheleneimine (PEI) coated SPIONs (150 nm) via electrostatic interaction [92]. Intraperitoneal injection induced significantly (p < 0.001) greater antibody levels than intramuscular, intradermal and subcutaneous injection. This was attributed to improved access to APCs in and around the peritoneum and the containment of particles at the injection site through use of an external magnet. A strong IgG2a response was generated which correlates with a Th1 biased response. In other work with DNA vaccines Shuaibu et al. designed a layered PEI/γ-polyglutamic acid (γ-PGA) nanoparticle complex [117]. Positively charged PEI complexes with DNA and was then coated with negatively charged γ-PGA. The nanoparticles were less than 100 nm in size, with a zeta potential of −28 mV. Following intravenous administration, between 33% and 60% of mice survived challenge with infected red blood cells, with no survival in groups immunized with naked DNA [86]. IgG1 and IgG2b dominated the antibody response and this along with increased IL-4 production indicated a Th2 biased response. Following this they investigated the use of intraperitoneal injection which completely protected mice upon challenge [87]. Significantly greater IL-12p40, IFN-γ, IL-4 and IL-10 levels suggested a more balanced response than was obtained in the previous study, though this was also the case for intravenously administered particles (which also demonstrated improved survival). The authors suggest the improved survival rates may be a result of an improved DNA expression vector. Using an alternative target, the PyG18p transimidase related protein, similar results were obtained [88]. The authors point to previous work to conclude that effective T cell immunity was the most important correlate for protection against this antigen. In addition they demonstrated that particles without antigen were able to stimulate cytokine production, emphasizing the importance of understanding particle interactions with the body.
As expected the majority of the studies outlined here induced immune responses comparable to traditional adjuvants, though few directly studied how these antibody levels correlated with protection. Different particles and vaccine formulations were able to induce very different helper T cell biases. While no clear conclusion can be drawn on the importance of Th1 and Th2 responses, it is likely that vectors inducing a balanced response are most favorable for use with blood stage vaccine candidates. Once again this highlights one of the most attractive features of particle based vectors, their ability to control the type of immune response developed.
4.3. Multi-Stage Vaccines
A number of vaccine candidates incorporating epitopes from proteins displayed at different stages of the malarial life cycle have been developed in an effort to account for the extreme variability present in some alleles and provide more well-rounded protection. The synthetic Plasmodium falciparum polypeptide vaccine, SPf66 is one such candidate which has undergone clinical testing [153]. It is a polypeptide composed of three blood stage antigens linked via CS protein sequences. Rosas et al. encapsulated this peptide in PLGA in an attempt to improve its immunogenicity [96,98]. Particles of 1.2 µm were developed with peptide both encapsulated and associated with the surface. The IgG titers following subcutaneous injection were higher than those obtained with alum and comparable to FA, but the protection elicited was minimal. In later work by this group, administration method was analyzed in depth [100]. Intradermal injection of PLGA microparticles encapsulating SPf66 induced significantly (p < 0.05) greater antibody titers against SPf66 than subcutaneous injection. High levels of IFN-γ secretion indicated that intradermal injection resulted in a greater Th1 bias than subcutaneous or alum adjuvanted administration especially in the lymph nodes. This was credited to more efficient immune organ targeting as evidenced by significantly higher splenocyte proliferation. These results were reinforced using novel MSP2 peptides and the S3 short synthetic peptide containing epitopes from both sporozoite and blood stages, where intradermal administration induced greater antibody titers than subcutaneous injection and adjuvantation with montanide ISA-720 [101,102]. It was speculated that the intradermal response was higher due to preferential DC vs. macrophage uptake [154]. Most recently, alginate and alginate modified with the amino acid sequence arginine-glycine-aspartate (RGD, involved in cell adhesion) have been incorporated into PLGA particles [103]. These 1 µm sized particles incorporated the antigens SPf66 and S3 more efficiently with reduced surface adsorption than PLGA. In both cases greater antibody titers were seen. This was stated to be a result of alginate enhancing inflammatory cell migration and activation at the injection site. Alginate inclusion led to at least twofold increases in IFN-γ secretion and lymphocyte proliferation with RGD further increasing both 1.4 fold. This along with increasing IgG2a levels implied that the additives were promoting a more balanced CD4+ T cell response. The authors claimed that alginate was known to increase IFN-γ levels and that RGD likely enhanced CD4+ T cell uptake.
4.4. Sexual Stage Vaccines
Very few studies have combined particulate vectors with malaria antigens from the sexual stage. This may in part be due to the difficulty inherent in testing their efficacy. While induction of antibodies is crucial for this stage of protection, it can be difficult to correlate antibody levels with the vaccines capability for inhibiting the spread of malaria. For instance even the most commonly used method of analyzing transmission blocking capability, the standard membrane feeding assay, only measures the oocyst burden. An understanding of how this relates to sporozoite numbers and overall disease transmission is still not clear, limiting its effectiveness [155]. Bhat et al. encapsulated a Pvs24 peptide in PLGA microparticles and delivered them intranasally and intramuscularly [105]. Intranasal delivery induced stronger antibody responses, especially when co-delivered with CpG oligodeoxynucleotide adjuvants, which trigger B-cell proliferation. Use of the sera in a mosquito membrane-feeding assay found a significant decrease in the number of oocysts present. The use of gel-core liposomes with Pfs25 antigen also produced antibody titers slightly better than those obtained with alum [106].
5. Connecting Nanotechnology and Malaria Vaccines
The design of nanoparticles for use as malaria vaccine vectors should be based on an understanding of nanoparticle properties, the desired immune response and how these interact. The importance of carrier properties has only recently become clear. This was evident in the analysis undertaken in the previous section, with few studies giving more than a brief overview of particle characteristics (e.g., some studies summarized in Table 1). It is recommended that future studies utilizing carriers describe particle characteristics in depth, as these can be vital in understanding the reported results. Properties that can be controlled include size, shape, surface charge, hydrophobicity, material, hardness, method of antigen incorporation, crystallinity and surface roughness, and functionality [156,157,158]. A number of these are highlighted in Figure 3. There is no clear consensus on how each property affects the immune response, though a number of them have been studied. The lack of consistency within and the number of uncontrolled variables between these studies often leads to conflicting results. Nevertheless, an understanding of the particle and its properties is crucial to the intelligent design of a malaria vaccine system. This section summarizes and correlates some of the studies on particle characteristics with respect to vaccines in general, and notes how they may be beneficial in future particle-based malaria vaccine research.
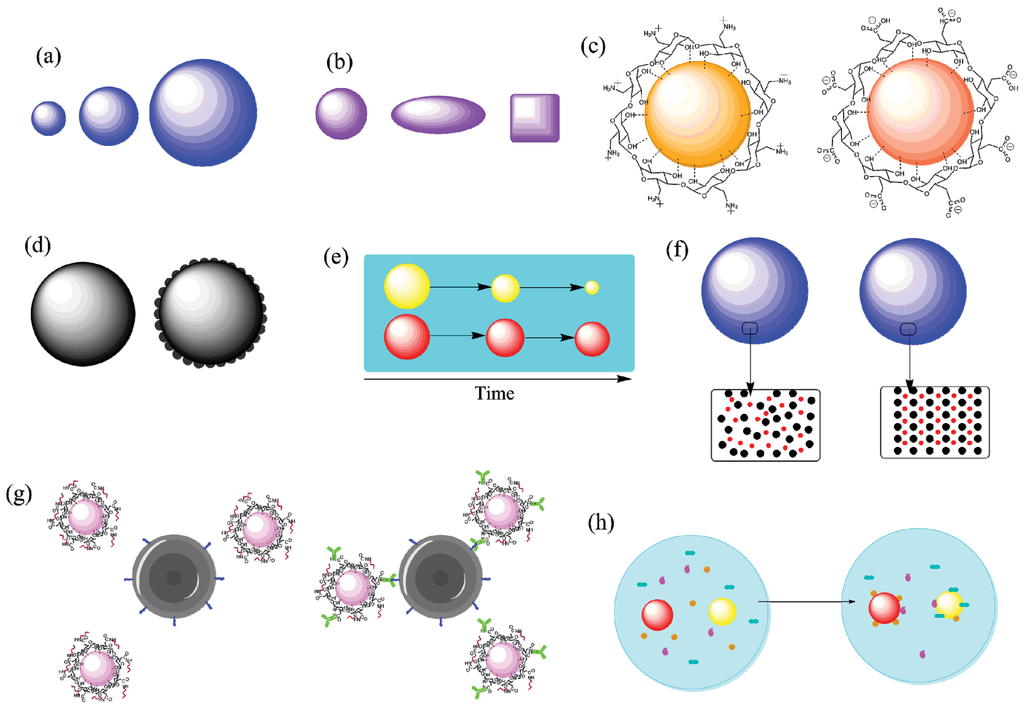
Figure 3.
Properties of nanoparticles that can be controlled via synthetic or post-synthetic modification. (a) Size; (b) Shape; (c) Surface charge; (d) Surface roughness; (e) Biodegradation rate; (f) Crystallinity; (g) Active targeting; (h) Protein corona formation. Figure developed using ChemBioDraw Ultra 14.
Figure 3.
Properties of nanoparticles that can be controlled via synthetic or post-synthetic modification. (a) Size; (b) Shape; (c) Surface charge; (d) Surface roughness; (e) Biodegradation rate; (f) Crystallinity; (g) Active targeting; (h) Protein corona formation. Figure developed using ChemBioDraw Ultra 14.
5.1. Size
In the previous section, the size is given in a number of cases but it is often unclear which size is referred to. The most important size is that seen by the body: the hydrodynamic size. The hydrodynamic size refers to the size of a hard sphere, which diffuses in the same manner as the particle being measured. Unlike sizes derived from dried samples, it accounts for particle solvation and the development of hydration layers. The hydrodynamic size of a particulate system has a strong effect on the particles passage within the body and its uptake by APCs [156]. The majority of nanoparticles can be synthesized in a size-controlled manner, though there are limits on the available range. For example, polymeric, iron oxide, gold, and silica nanoparticles with diameters less than 50 nm have been synthesized [159,160,161,162]. The wide range of monodisperse particles available for purchase currently is evidence of the ease of size control. Use of different reagents in different ratios and layered growth on seed particles are two of the techniques commonly used to limit size during synthesis [161,163]. Little consensus exists in the literature on the optimum size for a vaccine vector. Unlike drugs, where a small size is preferred as it allows improved permeation and better distribution in the body, particle size affects vaccine efficacy in multiple ways. Most importantly, size can determine where in the body the vector travels and consequently which APC populations it interacts with, how it is taken up and processed by cells, and which cell populations uptake favors [164,165,166,167]. These factors then affect the magnitude and type of immune response. It is difficult to disentangle how the delivery and cellular interaction factors mentioned above influence the immune response, and therefore most comparisons are solely based on the responses generated. This has lead different groups to conclude that both small and large particles are preferable [168]. The malarial pre-erythrocytic immune response initially develops in lymph nodes and later on in the liver while the blood stage response is generated in the spleen [37,169]. Therefore, the ability to deliver a vaccine directly to these sites might be beneficial as was first demonstrated by Fifis et al. using the model protein OVA [167]. In analyzing the immune response particles with sizes varying from 20–2000 nm were studied and it was concluded that conjugation of antigen to 40 nm particles was preferable as this conjugate induced high IFN-γ and antibody responses correlated with increased uptake by DCs inclusive of a subset which promotes cross presentation [167]. The same authors later showed a bias towards CD8+ T cell activation with 40–49 nm particles and a CD4+ response for 93–123 nm particles [170]. Particles that were 67 nm produced only weak CD4+ and CD8+ responses. One of the most common issues with malaria vaccines and vaccines in general is a lack of CD8+ T cell induction. This is visible in the RTS,S vaccine and in a number of other candidates undergoing clinical trials with conventional adjuvants [5]. Therefore the ability to focus the response against a malarial pre-erythrocytic antigen could be very valuable. In similar work looking at particle distribution, Reddy et al. used polypropylene sulphide nanoparticles of 20 nm, 45 nm, and 100 nm without antigen to demonstrate that 20 nm particles drained to local lymph nodes to the greatest degree, while 100 nm particles stayed at the injection site [171]. Similarly 74% of 40 nm liposomes injected subcutaneously were taken up by the lymphatics while larger particles remained almost solely at the injection site [172]. There are limited data comparing different sized particles but other studies using small nanoparticles have also seen localization in lymph nodes [173,174,175,176]. This work suggests that the cut-off is ~100 nm, with larger particles staying at the injection site. Lymph node targeting is advantageous as it exposes the vaccine conjugate system to different subsets of APCs at much higher concentrations than are present in the periphery. This can potentially lead to stronger immune responses.
Analyzing the correlations between size and immune response, differences in the literature can be seen, for example in the work of Li et al. and Gutierro et al. [177,178]. The latter suggests that 1000 nm PLGA particles encapsulating bovine serum albumin induce greater immune responses than 500 nm and 200 nm particles while the former measured higher humoral and cellular responses with 230 nm particles compared to 708 nm particles conjugated to OVA. This highlights the importance of factors such as the method of antigen loading, particle size dispersity, particle type, and the route of administration amongst others; all of which may affect the immune response developed [168]. In any case, the ability to travel directly to lymph nodes makes smaller particles (<100 nm) attractive, but very few studies have directly compared the immune response of these particles with larger particles in vivo. In vitro evidence suggested that the optimal size for cellular uptake is 50 nm, although these studies were done with fibroblast, ovarian cancer and brain tumor cell lines, not APCs [179,180]. It has been proposed that this size range represents the thermodynamic optimum for receptor-mediated endocytosis [181,182]. Given that 20–200 nm particles are thought to be ingested via the same mechanism in DCs, it is plausible that a similar trend would be seen [164]. It is clear that hydrodynamic size is an important variable in vector design and should be reported in all cases, however, this was lacking in the studies analyzed in the previous section.
5.2. Surface Charge
One of the most attractive features of particle-based vectors is their amenability to surface modification. This includes both coating and encapsulation of the particle, and reaction of surface functional groups to produce a coating with alternate properties [156]. Coating is mainly used for one of two reasons: to improve the stability of the particle in solution or to alter the properties of the surface and enhance/alter the immune response. Stability can be improved by increasing the repulsive force between particles, thereby reducing their tendency to aggregate. Both steric and electrostatic forces are involved in aggregation and affect stability [183]. Various mechanisms for improving the immune response have been investigated, including altering the surface charge, changing the hydrophobicity, and the attachment of targeting ligands [158]. To take advantage of the negative charge of cell membranes, positively charged particles have been designed towards improved uptake and subsequent immune presentation [184,185]. A common issue with studies of this kind is ensuring that the modification of surface charge does not alter the other properties of the system. For instance, Thomas et al. found that cationic PLGA induced stronger humoral and mucosal immune responses against entrapped hepatitis B surface antigen than unmodified particles but acknowledged that other variables such as size and entrapment efficiency were likely to have influenced the results [186]. Henriksen-Lacey et al. observed that cationic liposomes with a tuberculosis antigen induced greater Th1 and Th17 cytokine responses than those with a neutral charge, potentially due to enhanced antigen absorption and the ability of the cationic material to act as an adjuvant [187]. There are fewer studies which look at nanoparticles, with Villanueva et al. and Cho et al. both reporting greater uptake of cationic particles in cancer cells in vitro [188,189]. In contrast, Foged et al. found no difference in DC uptake between 100 nm cationic and anionic particles without antigen [190]. Larger 1000 nm cationic particles were taken up to a greater degree than equivalent anionic particles. Comparable DC uptake suggests that anionic particles may be better in the nano-size range as they will exhibit less non-specific cellular uptake. It should be noted that none of these studies analyzed the immune response in vivo. Lastly there is evidence that internalization pathways and the type of immune response differ based on particle charge, though further investigation is required [191,192]. This suggests that while there may be potential benefits to altering particle charge in a malaria vaccine formulation, the limited data in this area constrain recommendations. If the antigen in question is charged or DNA based, attachment to an oppositely charged particle could produce a more stable formulation, as was demonstrated with SPIONs, via prolonged presentation [92].
5.3. Targeting Ligands
Any vector to which antigen can be adsorbed or conjugated can similarly be altered with targeting ligands. These ligands enable direct interactions with APCs and consequently they can improve cellular uptake and activation. Pattern recognition receptors (PRRs) on the surface of DCs allow identification of pathogen associated molecular patterns (PAMPs). This receptor subset includes toll-like receptors (TLRs) involved in recognition of and response to pathogen invasion, CLRs which aid endocytosis and nucleotide oligomerization domain (NOD)-like receptors that regulate the immune response. Agonists for these receptors can be conjugated to the surface of vectors to induce maturation, control the nature of the immune response (through TLRs) and/or increase particle uptake and subsequent antigen presentation (through CLRs) [193]. In the majority of TLR work, the agonist has either been encapsulated in a particle or co-administered as an adjuvant [194]. This was seen in a number of the liposomal studies outlined in the previous section, with the incorporation of the TLR-4 agonist lipid A [62,68]. Contrastingly, a number of studies have looked at surface conjugation of CLR ligands [195]. Kwon et al. conjugated monoclonal DEC-205 antibodies to polymeric particles encapsulating OVA, inducing twice as many CD8+ T cells than particles with a control antibody conjugated [196]. Similar enhancements of both CD4+ and CD8+ mediated immunity have also been reported [197,198]. Other receptors which have been targeted include mannose and DC-SIGN. Thomann et al demonstrated equivalent vaccine efficacy with decreased quantities of a cytotoxic ErbB2 antigen and a TLR2 adjuvant using a mannose agonist, while Cruz et al. found that DC-SIGN targeting increased the uptake of PLGA nanoparticles containing a tetanus toxin peptide but not microparticles [199,200]. Using the same particles, the authors demonstrated greater T cell proliferation [201]. Targeting to DCs is mostly required in the periphery, as DC concentrations in lymph nodes are far greater. Drainage via size control is likely preferable as it removes the need for ligand attachment and prevents early antigen presentation which can induce self-tolerance [193]. On the other hand, engaging targeting as a means for enhancing antigen uptake by APC subsets is attractive. Targeted uptake of particles by cross-presenting DCs promoting enhanced CD8+ responses would be useful, for instance in the development of a vaccine against a liver-stage malaria antigen.
5.4. Protein Corona and Material Characteristics
In the previous section, it was evident that a wide range of materials have been used in particle vectors for malaria vaccines. It was also clear that the choice of said material affected the immune response, even when the other characteristics of the particles were similar. For instance, Pusic et al. used quantum dots and SPIONs of a similar size in the same manner, but were only able to induce a strong subcutaneous response with quantum dots [14,94]. The elemental constituents, surface roughness and configuration, crystallinity, and the method of antigen incorporation all potentially alter the particles interaction with the body. Upon injection, particles interact with tissues, fluids, and a wide range of cells. The formation of a protein corona is the most important of these interactions. Serum and plasma contain an array of different proteins, each of which has the potential to adsorb to the surface of a particle [202]. The affinity of this interaction will depend on the particles composition as well as a number of other characteristics (size, charge, hydrophobicity) [203]. The adsorption of fibrinogen and immunoglobulins is dominant on ultra-small SPIONs while apolipoprotein bound more strongly to solid lipid nanoparticles [204,205]. Results of this kind have been demonstrated with a range of particles composed of different materials [206]. Adsorption of a protein layer increases the hydrodynamic size, can potentially influence the structure of the protein, and control the particles’ interaction with cells [207,208]. Wang et al. revealed that caveolae-based endocytosis of polymeric nanoparticles was promoted by the presence of an albumin coating [209]. Furthermore, it has been suggested that the presence of known opsonins, such as complement and immunoglobulins in the protein corona could be beneficial in the design of vaccines [203]. While this is just speculation, an understanding of protein coronae development and structure is as important as that of size in the design of malaria vaccine vectors. The other factor that needs to be considered in the choice of material is biocompatibility. This includes both biodegradability and toxicity; requirements which tend to limit the applicability of inorganic materials as was evident in the previous section [210].
5.5. Method of Antigen Incorporation
The method by which the subunit or DNA vaccine is combined with the particle system affects the subsequent immune response. Figure 2 highlights how the different systems investigated incorporate antigen. Particles can be mixed with the vaccine as are traditional adjuvants, or the antigen can be adsorbed or covalently bound to the surface [211]. Adsorption relies upon attractive forces such as electrostatic and van der Waal interactions and is weaker than covalent bonding [212]. Antigenic epitopes can also be incorporated into the particle framework, as in the previously mentioned SAPNs [142]. Hollow and porous nanoparticles allow for encapsulation of antigen, which is typically added to the initial reagent mixture [62]. The release rate depends on the degradation of the particle in vivo. It is evident that the encapsulation method used will lead to differences in the degree of antigen protection, the swiftness and length of the immune response, and the intracellular processing of the antigen. It was previously mentioned that encapsulation combined with surface display induced nine-fold greater antibody titers against a P. vivax CSP antigen than encapsulation alone in ICMVs [71]. While many of the variables alluded to in this section have only seen limited study, their existence demonstrates the possibilities nanoparticles provide as vectors, especially when compared with traditional adjuvants.
6. Conclusions
It is readily apparent that particle-based vaccine vectors can induce immunity relevant to the development of protection against malaria. A direct comparison between different synthetic vectors is difficult, as each study incorporates distinct antigens and injection regimes. Immune responses against specific antigens can be directed through particle selection, although this is not a guarantee that the same bias will result when other antigens are used. Effective improvement of vectors has been demonstrated through enhancement of both humoral and cellular responses, and the control of the type of cellular response, though this would benefit from a stronger mechanistic understanding. Few in-depth reports of in vivo particle interaction mechanisms are provided in the studies analyzed. These types of studies are generally extremely complex but can offer in-depth insight into not just the particle being tested, but those with similar properties without conjecture. Vital characteristics including protein corona structure, and nanoparticle size and charge are often neglected. As was evident in the previous section, the importance of these factors cannot be underestimated.
Exploration into the manipulation and control of immune responses via a wide range of particle properties is producing promising results, which should influence future studies. SAPN development is producing systems which induce high antibody titers with high avidity for their epitopes, a requirement of both anti-sporozoite and anti-blood stage vaccine constructs. ICMVs and polymeric particles have been able to induce long-lived memory cells while PEI-γPGA systems are promising transfection agents. Size-dependent cytotoxic T cell induction may enable effective targeting of infected hepatocytes, overcoming an issue common to malaria vaccine candidates. Future vectors can be designed to complement different antigens and potentially create a multi-stage vaccine. Furthermore, systems incorporating multiple epitopes or multilayered particles capable of multiple antigen releases are possibilities. The positive results induced using biodegradable materials with proven toxicity profiles suggest that further research into particulate vectors will be beneficial.
Acknowledgments
Magdalena Plebanski is an NHMRC Senior Research Fellow and Cordelia Selomulya an ARC Future Research Fellow. Magdalena Plebanski and Cordelia Selomulya are in receipt of ARC DP150101058 Grant. Liam Powles is supported by an Australian Postgraduate Award.
Author Contributions
Liam Powles reviewed the literature and developed the manuscript and the figures with supervision from Magdalena Plebanski, Cordelia Selomulya, and Sue D. Xiang. All authors revised, edited and proofread the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- WHO. World Malaria Report 2014; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Murray, C.J.L.; Rosenfeld, L.C.; Lim, S.S.; Andrews, K.G.; Foreman, K.J.; Haring, D.; Fullman, N.; Naghavi, M.; Lozano, R.; Lopez, A.D. Global malaria mortality between 1980 and 2010: A systematic analysis. Lancet 2012, 379, 413–431. [Google Scholar] [CrossRef]
- Playfair, J.H.L.; Taverne, J.; Bate, C.A.W.; Brian de Souza, J. The malaria vaccine: Anti-parasite or anti-disease? Immunol. Today 1990, 11, 25–27. [Google Scholar] [CrossRef]
- Conway, D.J. Natural selection on polymorphic malaria antigens and the search for a vaccine. Parasitol. Today 1997, 13, 26–29. [Google Scholar] [CrossRef]
- Schwartz, L.; Brown, G.V.; Genton, B.; Moorthy, V.S. A review of malaria vaccine clinical projects based on the WHO rainbow table. Malar. J. 2012. [Google Scholar] [CrossRef] [PubMed]
- Seder, R.A.; Chang, L.-J.; Enama, M.E.; Zephir, K.L.; Sarwar, U.N.; Gordon, I.J.; Holman, L.A.; James, E.R.; Billingsley, P.F.; Gunasekera, A.; et al. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 2013, 341, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.E.; Williamson, D.; Titball, R. Vaccine delivery using nanoparticles. Front. Cell. Infect. Microbiol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Doolan, D.L. Plasmodium immunomics. Int. J. Parasitol. 2011, 41, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Vogel, F.R. Immunologic adjuvants for modern vaccine formulations. Ann. N.Y. Acad. Sci. 1995, 754, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Schijns, V.E.J.C. Immunological concepts of vaccine adjuvant activity: Commentary. Curr. Opin. Immunol. 2000, 12, 456–463. [Google Scholar] [CrossRef]
- Beeson, J.G.; Osier, F.H.A.; Engwerda, C.R. Recent insights into humoral and cellular immune responses against malaria. Trends Parasitol. 2008, 24, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Fries, L.F.; Gordon, D.M.; Richards, R.L.; Egan, J.E.; Hollingdale, M.R.; Gross, M.; Silverman, C.; Alving, C.R. Liposomal malaria vaccine in humans: A safe and potent adjuvant strategy. Proc. Natl. Acad. Sci. USA 1992, 89, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Thomasin, C.; Corradin, G.; Men, Y.; Merkle, H.P.; Gander, B. Tetanus toxoid and synthetic malaria antigen containing poly (lactide)/poly (lactide-co-glycolide) microspheres: Importance of polymer degradation and antigen release for immune response. J. Controll. Release 1996, 41, 131–145. [Google Scholar] [CrossRef]
- Pusic, K.; Aguilar, Z.; McLoughlin, J.; Kobuch, S.; Xu, H.; Tsang, M.; Wang, A.; Hui, G. Iron oxide nanoparticles as a clinically acceptable delivery platform for a recombinant blood-stage human malaria vaccine. FASEB J. 2013, 27, 1153–1166. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Goyal, A.K.; Khatri, K.; Mishra, N.; Mehta, A.; Vaidya, B.; Tiwari, S.; Paliwal, R.; Paliwal, S. Development of self-assembled nanoceramic carrier construct(s) for vaccine delivery. J. Biomater. Appl. 2009, 24, 65–84. [Google Scholar]
- Ruigrok, R.W.H.; Gerlier, D. Structure of the measles virus H glycoprotein sheds light on an efficient vaccine. Proc. Natl. Acad. Sci. USA 2007, 104, 20639–20640. [Google Scholar] [CrossRef] [PubMed]
- Hilleman, M.R.; Buynak, E.B.; Weibel, R.E.; Stokes, J. Live, attenuated mumps-virus vaccine. New Engl. J. Med. 1968, 278, 227–232. [Google Scholar] [CrossRef] [PubMed]
- American Academy of Pediatrics Committee on Infectious Diseases. Poliomyelitis prevention: Recommendations for use of inactivated poliovirus vaccine and live oral poliovirus vaccine. Pediatrics 1997, 99, 300–305. [Google Scholar]
- Schild, G.; Corbel, M.; Corran, P.; Minor, P. Vaccines: Past, Present and Future. In Vaccines; Perlmann, P., Wigzell, H., Eds.; Springer: Berlin, Germany; Heidelberg, Germany, 1999; Volume 133, pp. 1–19. [Google Scholar]
- Ulmer, J.B.; Donnelly, J.J.; Liu, M.A. DNA Vaccines: Immunogenicity and Preclinical Efficacy. In Vaccines; Perlmann, P., Wigzell, H., Eds.; Springer: Berlin, Germany; heidelberg, Germany, 1999; Volume 133, pp. 43–55. [Google Scholar]
- Robinson, H.L.; Pertmer, T.M. DNA vaccines for viral infections: Basic studies and applications. In Advances in Virus Research; Academic Press: Waltham, MA, USA, 2000; Volume 55, pp. 1–74. [Google Scholar]
- Male, D.; Brostoff, J.; Roth, D.; Roitt, I. Immunology, 8 ed.; Saunders Elsevier: Philadelphia, PA, USA, 2012. [Google Scholar]
- Ferraro, B.; Morrow, M.P.; Hutnick, N.A.; Shin, T.H.; Lucke, C.E.; Weiner, D.B. Clinical applications of DNA vaccines: Current progress. Clin. Infect. Dis. 2011, 53, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Friede, M.; Aguado, M.T. Need for new vaccine formulations and potential of particulate antigen and DNA delivery systems. Adv. Drug Deliv. Rev. 2005, 57, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, J.B.; Valley, U.; Rappuoli, R. Vaccine manufacturing: Challenges and solutions. Nat. Biotech. 2006, 24, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic Potential of Materials at the Nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Crompton, P.D.; Pierce, S.K.; Miller, L.H. Advances and challenges in malaria vaccine development. J. Clin. Investig. 2010, 120, 4168–4178. [Google Scholar] [CrossRef] [PubMed]
- Riley, E.M.; Stewart, V.A. Immune mechanisms in malaria: New insights in vaccine development. Nat. Med. 2013, 19, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Finney, O.C.; Keitany, G.J.; Smithers, H.; Kaushansky, A.; Kappe, S.; Wang, R. Immunization with genetically attenuated P. falciparum parasites induces long-lived antibodies that efficiently block hepatocyte invasion by sporozoites. Vaccine 2014, 32, 2135–2138. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Coppi, A.; Natarajan, R.; Pradel, G.; Bennett, B.L.; James, E.R.; Roggero, M.A.; Corradin, G.; Persson, C.; Tewari, R.; Sinnis, P. The malaria circumsporozoite protein has two functional domains, each with distinct roles as sporozoites journey from mosquito to mammalian host. J. Exp. Med. 2011, 208, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.; Spaccapelo, R.; Bistoni, F.; Holder, A.A.; Crisanti, A. Function of region I and II adhesive motifs of Plasmodium falciparum circumsporozoite protein in sporozoite motility and infectivity. J. Biol. Chem. 2002, 277, 47613–47618. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.A.; Thathy, V.; Frevert, U.; Robson, K.J.H.; Crisanti, A.; Nussenzweig, V.; Nussenzweig, R.S.; Ménard, R. TRAP Is necessary for gliding motility and infectivity of Plasmodium sporozoites. Cell 1997, 90, 511–522. [Google Scholar] [CrossRef]
- Baldacci, P.; Ménard, R. The elusive malaria sporozoite in the mammalian host. Mol. Microbiol. 2004, 54, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Silvie, O.; Franetich, J.-F.; Charrin, S.; Mueller, M.S.; Siau, A.; Bodescot, M.; Rubinstein, E.; Hannoun, L.; Charoenvit, Y.; Kocken, C.H.; et al. A role for apical membrane antigen 1 during invasion of hepatocytes by Plasmodium falciparum sporozoites. J. Biol. Chem. 2004, 279, 9490–9496. [Google Scholar] [CrossRef] [PubMed]
- Kariu, T.; Ishino, T.; Yano, K.; Chinzei, Y.; Yuda, M. CelTOS, a novel malarial protein that mediates transmission to mosquito and vertebrate hosts. Mol. Microbiol. 2006, 59, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Ishino, T.; Yano, K.; Chinzei, Y.; Yuda, M. Cell-passage activity is required for the malarial parasite to cross the liver sinusoidal cell layer. PLoS Biol. 2004, 2, e4. [Google Scholar] [CrossRef] [PubMed]
- Cockburn, I.A.; Tse, S.-W.; Radtke, A.J.; Srinivasan, P.; Chen, Y.-C.; Sinnis, P.; Zavala, F. Dendritic cells and hepatocytes use distinct pathways to process protective antigen from plasmodium in vivo. PLoS Pathog. 2011, 7, e1001318. [Google Scholar] [CrossRef] [PubMed]
- Butler, N.S.; Schmidt, N.W.; Harty, J.T. Differential effector pathways regulate memory CD8 T cell immunity against Plasmodium berghei versus P. yoelii sporozoites. J. Immunol. 2010, 184, 2528–2538. [Google Scholar] [CrossRef] [PubMed]
- Günther, K.; Tümmler, M.; Arnold, H.-H.; Ridley, R.; Goman, M.; Scaife, J.G.; Lingelbach, K. An exported protein of Plasmodium falciparum is synthesized as an integral membrane protein. Mol. Biochem. Parasitol. 1991, 46, 149–157. [Google Scholar] [CrossRef]
- Mikolajczak, S.A.; Sacci, J.J.B.; de la Vega, P.; Camargo, N.; VanBuskirk, K.; Krzych, U.; Cao, J.; Jacobs-Lorena, M.; Cowman, A.F.; Kappe, S.H.I. Disruption of the Plasmodium falciparum liver-stage antigen-1 locus causes a differentiation defect in late liver-stage parasites. Cell. Microbiol. 2011, 13, 1250–1260. [Google Scholar] [CrossRef] [PubMed]
- Prieur, E.; Druilhe, P. The malaria candidate vaccine liver stage antigen-3 is highly conserved in Plasmodium falciparum isolates from diverse geographical areas. Malar. J. 2009. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.W.; Butler, N.S.; Badovinac, V.P.; Harty, J.T. Extreme CD8 T cell requirements for anti-malarial liver-stage immunity following immunization with radiation attenuated sporozoites. PLoS Pathog. 2010, 6, e1000998. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.W.; Podyminogin, R.L.; Butler, N.S.; Badovinac, V.P.; Tucker, B.J.; Bahjat, K.S.; Lauer, P.; Reyes-Sandoval, A.; Hutchings, C.L.; Moore, A.C.; et al. Memory CD8 T cell responses exceeding a large but definable threshold provide long-term immunity to malaria. Proc. Natl. Acad. Sci. USA 2008, 105, 14017–14022. [Google Scholar] [CrossRef] [PubMed]
- Osier, F.H.A.; Fegan, G.; Polley, S.D.; Murungi, L.; Verra, F.; Tetteh, K.K.A.; Lowe, B.; Mwangi, T.; Bull, P.C.; Thomas, A.W.; et al. Breadth and magnitude of antibody responses to multiple Plasmodium falciparum merozoite antigens are associated with protection from clinical malaria. Infect. Immun. 2008, 76, 2240–2248. [Google Scholar] [CrossRef] [PubMed]
- McCall, M.B.; Roestenberg, M.; Ploemen, I.; Teirlinck, A.; Hopman, J.; de Mast, Q.; Dolo, A.; Doumbo, O.K.; Luty, A.; van der Ven, A.J. Memory-like IFN-γ response by NK cells following malaria infection reveals the crucial role of T cells in NK cell activation by P. falciparum. Eur. J. Immunol. 2010, 40, 3472–3477. [Google Scholar] [CrossRef] [PubMed]
- McCall, M.B.B.; Sauerwein, R.W. Interferon-γ—Central mediator of protective immune responses against the pre-erythrocytic and blood stage of malaria. J. Leukocyte Biol. 2010, 88, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.C.; Corran, P.H.; Mangia, E.; Gaunt, M.W.; Li, Q.; Tetteh, K.K.A.; Polley, S.D.; Conway, D.J.; Holder, A.A.; Bacarese-Hamilton, T.; et al. Profiling the antibody immune response against blood stage malaria vaccine candidates. Clin. Chem. 2007, 53, 1244–1253. [Google Scholar] [CrossRef] [PubMed]
- Volkman, S.K.; Hartl, D.L.; Wirth, D.F.; Nielsen, K.M.; Choi, M.; Batalov, S.; Zhou, Y.; Plouffe, D.; le Roch, K.G.; Abagyan, R.; et al. Excess polymorphisms in genes for membrane proteins in Plasmodium falciparum. Science 2002, 298, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.H.; Roberts, T.; Shahabuddin, M.; McCutchan, T.F. Analysis of sequence diversity in the Plasmodium falciparum merozoite surface protein-1 (MSP-1). Mol. Biochem. Parasitol. 1993, 59, 1–14. [Google Scholar] [CrossRef]
- Goel, V.K.; Li, X.; Chen, H.; Liu, S.-C.; Chishti, A.H.; Oh, S.S. Band 3 is a host receptor binding merozoite surface protein 1 during the Plasmodium falciparum invasion of erythrocytes. Proc. Natl. Acad. Sci. USA 2003, 100, 5164–5169. [Google Scholar] [CrossRef] [PubMed]
- Cowman, A.F.; Crabb, B.S. Invasion of red blood cells by malaria parasites. Cell 2006, 124, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Remarque, E.J.; Faber, B.W.; Kocken, C.H.M.; Thomas, A.W. Apical membrane antigen 1: A malaria vaccine candidate in review. Trends Parasitol. 2008, 24, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Lee Sim, B.K.; Toyoshima, T.; David Haynes, J.; Aikawa, M. Localization of the 175-kilodalton erythrocyte binding antigen in micronemes of Plasmodium falciparum merozoites. Mol. Biochem. Parasitol. 1992, 51, 157–159. [Google Scholar] [CrossRef]
- Douglas, A.D.; Baldeviano, G.C.; Miura, K.; Wright, G.J.; Draper, S.J. PfRH5 vaccine efficacy against heterologous strain blood-stage Plasmodium falciparum. Lancet 2014. [Google Scholar] [CrossRef]
- Bustamante, L.Y.; Bartholdson, S.J.; Crosnier, C.; Campos, M.G.; Wanaguru, M.; Nguon, C.; Kwiatkowski, D.P.; Wright, G.J.; Rayner, J.C. A full-length recombinant Plasmodium falciparum PfRH5 protein induces inhibitory antibodies that are effective across common PfRH5 genetic variants. Vaccine 2013, 31, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Healer, J.; McGuinness, D.; Hopcroft, P.; Haley, S.; Carter, R.; Riley, E. Complement-mediated lysis of Plasmodium falciparum gametes by malaria-immune human sera is associated with antibodies to the gamete surface antigen Pfs230. Infect. Immun. 1997, 65, 3017–3023. [Google Scholar] [PubMed]
- Vaughan, A.M.; Kappe, S.H.I. Malaria vaccine development: Persistent challenges. Curr. Opin. Immunol. 2012, 24, 324–331. [Google Scholar] [PubMed]
- Barr, P.J.; Green, K.M.; Gibson, H.L.; Bathurst, I.C.; Quakyi, I.A.; Kaslow, D.C. Recombinant Pfs25 protein of Plasmodium falciparum elicits malaria transmission-blocking immunity in experimental animals. J. Exp. Med. 1991, 174, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, M.; Sato, C.; Otsuki, H.; Sattabongkot, J.; Kaneko, O.; Torii, M.; Tsuboi, T. Plasmodium vivax gametocyte protein Pvs230 is a transmission-blocking vaccine candidate. Vaccine 2012, 30, 1807–1812. [Google Scholar] [CrossRef] [PubMed]
- Eksi, S.; Czesny, B.; van Gemert, G.-J.; Sauerwein, R.W.; Eling, W.; Williamson, K.C. Malaria transmission-blocking antigen, Pfs230, mediates human red blood cell binding to exflagellating male parasites and oocyst production. Mol. Microbiol. 2006, 61, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, D.R.; Angov, E.; Kariuki, T.; Kumar, N. A potent malaria transmission blocking vaccine based on codon harmonized full length Pfs48/45 expressed in Escherichia coli. PLoS ONE 2009, 4, e6352. [Google Scholar] [PubMed]
- Richards, R.; Hayre, M.; Hockmeyer, W.; Alving, C. Liposomes, lipid A, and aluminum hydroxide enhance the immune response to a synthetic malaria sporozoite antigen. Infect. Immun. 1988, 56, 682–686. [Google Scholar] [PubMed]
- Alving, C.R.; Richards, R.L.; Moss, J.; Alving, L.I.; Clements, J.D.; Shiba, T.; Kotani, S.; Wirtz, R.A.; Hockmeyer, W.T. Effectiveness of liposomes as potential carriers of vaccines: Applications to cholera toxin and human malaria sporozoite antigen. Vaccine 1986, 4, 166–172. [Google Scholar] [CrossRef]
- Richards, R.L.; Swartz, G.M., Jr.; Schultz, C.; Hayre, M.D.; Ward, G.S.; Ballou, W.R.; Chulay, J.D.; Hockmeyer, W.T.; Berman, S.L.; Alving, C.R. Immunogenicity of liposomal malaria sporozoite antigen in monkeys: Adjuvant effects of aluminium hydroxide and non-pyrogenic liposomal lipid A. Vaccine 1989, 7, 506–512. [Google Scholar] [CrossRef]
- White, K.; Krzych, U.; Gordon, D.M.; Porter, T.G.; Richards, R.L.; Alving, C.R.; Deal, C.D.; Hollingdale, M.; Silverman, C.; Sylvester, D.R. Induction of cytolytic and antibody responses using Plasmodium falciparum repeatless circumsporozoite protein encapsulated in liposomes. Vaccine 1993, 11, 1341–1346. [Google Scholar] [CrossRef]
- Verma, J.; Rao, M.; Amselem, S.; Krzych, U.; Alving, C.; Green, S.; Wassef, N. Adjuvant effects of liposomes containing lipid A: Enhancement of liposomal antigen presentation and recruitment of macrophages. Infect. Immun. 1992, 60, 2438–2444. [Google Scholar] [PubMed]
- Wang, R.; Charoenvit, Y.; Corradin, G.; Porrozzi, R.; Hunter, R.L.; Glenn, G.; Alving, C.R.; Church, P.; Hoffman, S.L. Induction of protective polyclonal antibodies by immunization with a Plasmodium yoelii circumsporozoite protein multiple antigen peptide vaccine. J. Immunol. 1995, 154, 2784–2793. [Google Scholar] [PubMed]
- Richards, R.L.; Rao, M.; Wassef, N.M.; Glenn, G.M.; Rothwell, S.W.; Alving, C.R. Liposomes containing lipid A serve as an adjuvant for induction of antibody and cytotoxic T-cell responses against RTS,S malaria antigen. Infect. Immun. 1998, 66, 2859–2865. [Google Scholar] [PubMed]
- Heppner, D.; Gordon, D.; Gross, M.; Wellde, B.; Leitner, W.; Krzych, U.; Schneider, I.; Wirtz, R.; Richards, R.; Trofa, A. Safety, immunogenicity, and efficacy of Plasmodium falciparum repeatless circumsporozoite protein vaccine encapsulated in liposomes. J. Infect. Dis. 1996, 174, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.P.; Jadon, R.S.; Goyal, A.K.; Mishra, N.; Gupta, P.N.; Khatri, K.; Tyagi, R. pH sensitive liposomes enhances immunogenicity of 19 kDa carboxyl-terminal fragment of Plasmodium falciparum. Int. J. Pharm. Sci. Nanotechnol. 2008, 1, 78–86. [Google Scholar]
- Moon, J.J.; Suh, H.; Li, A.V.; Ockenhouse, C.F.; Yadava, A.; Irvine, D.J. Enhancing humoral responses to a malaria antigen with nanoparticle vaccines that expand Tfh cells and promote germinal center induction. Proc. Natl. Acad. Sci. USA 2012, 109, 1080–1085. [Google Scholar] [CrossRef] [PubMed]
- Amselem, S.; Alving, C.R.; Domb, A.J. Polymeric biodegradable lipospheres™ as vaccine delivery systems. Polymers Adv. Technol. 1992, 3, 351–357. [Google Scholar] [CrossRef]
- Men, Y.; Gander, B.; Merkle, H.P.; Corradin, G. Induction of sustained and elevated immune responses to weakly immunogenic synthetic malarial peptides by encapsulation in biodegradable polymer microspheres. Vaccine 1996, 14, 1442–1450. [Google Scholar] [PubMed]
- Men, Y.; Tamber, H.; Audran, R.; Gander, B.; Corradin, G. Induction of a cytotoxic T lymphocyte response by immunization with a malaria specific CTL peptide entrapped in biodegradable polymer microspheres. Vaccine 1997, 15, 1405–1412. [Google Scholar] [CrossRef]
- Nixon, D.F.; Hioe, C.; Chen, P.-D.; Bian, Z.; Kuebler, P.; Li, M.-L.; Qiu, H.; Li, X.-M.; Singh, M.; Richardson, J.; et al. Synthetic peptides entrapped in microparticles can elicit cytotoxic T cell activity. Vaccine 1996, 14, 1523–1530. [Google Scholar] [CrossRef]
- Moon, J.J.; Suh, H.; Polhemus, M.E.; Ockenhouse, C.F.; Yadava, A.; Irvine, D.J. Antigen-displaying lipid-enveloped PLGA nanoparticles as delivery agents for a Plasmodium vivax malaria vaccine. PLoS ONE 2012, 7, e31472. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.L.; Xiang, S.D.; Plebanski, M. Montanide, Poly I: C and nanoparticle based vaccines promote differential suppressor and effector cell expansion: A study of induction of CD8 T cells to a minimal Plasmodium berghei epitope. Front. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Kaba, S.A.; Brando, C.; Guo, Q.; Mittelholzer, C.; Raman, S.; Tropel, D.; Aebi, U.; Burkhard, P.; Lanar, D.E. A nonadjuvanted polypeptide nanoparticle vaccine confers long-lasting protection against rodent malaria. J. Immunol. 2009, 183, 7268–7277. [Google Scholar] [CrossRef] [PubMed]
- Kaba, S.A.; McCoy, M.E.; Doll, T.A.; Brando, C.; Guo, Q.; Dasgupta, D.; Yang, Y.; Mittelholzer, C.; Spaccapelo, R.; Crisanti, A. Protective antibody and CD8+ T-cell responses to the Plasmodium falciparum circumsporozoite protein induced by a nanoparticle vaccine. PLoS ONE 2012, 7, e48304. [Google Scholar] [CrossRef] [PubMed]
- McCoy, M.E.; Golden, H.E.; Doll, T.A.; Yang, Y.; Kaba, S.A.; Burkhard, P.; Lanar, D.E. Mechanisms of protective immune responses induced by the Plasmodium falciparum circumsporozoite protein-based, self-assembling protein nanoparticle vaccine. Malar. J. 2013. [Google Scholar] [CrossRef]
- Powell, T.J.; Tang, J.; DeRome, M.E.; Mitchell, R.A.; Jacobs, A.; Deng, Y.; Palath, N.; Cardenas, E.; Boyd, J.G.; Nardin, E. Plasmodium falciparum synthetic LbL microparticle vaccine elicits protective neutralizing antibody and parasite-specific cellular immune responses. Vaccine 2013, 31, 1898–1904. [Google Scholar] [CrossRef] [PubMed]
- Burghaus, P.A.; Wellde, B.T.; Hall, T.; Richards, R.L.; Egan, A.F.; Riley, E.M.; Ballou, W.R.; Holder, A.A. Immunization of Aotus nancymai with recombinant C terminus of Plasmodium falciparum merozoite surface protein 1 in liposomes and alum adjuvant does not induce protection against a challenge infection. Infect. Immun. 1996, 64, 3614–3619. [Google Scholar] [PubMed]
- Sharma, S.K.; Gupta, C.; Dwivedi, V.; Misra-Bhattacharya, S.; Mohammad, O. Prophylactic potential of liposomized integral membrane protein of Plasmodium yoelii nigeriensis against blood stage infection in BALB/c mice. Vaccine 2007, 25, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Farah, D.; Misra-Bhattacharya, S.; Bajpai, P.; Agarwal, A.; Mohammad, O. Escheriosome entrapped soluble blood stage antigens impart protective immunity against a multi-drug resistant isolate of Plasmodium yoelii nigeriensis in BALB/c mice. Vaccine 2006, 24, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, V.; Vasco, A.; Vedi, S.; Dangi, A.; Arif, K.; Bhattacharya, S.M.; Owais, M. Adjuvanticity and protective immunity of Plasmodium yoelii nigeriensis blood-stage soluble antigens encapsulated in fusogenic liposome. Vaccine 2009, 27, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Shuaibu, M.; Cherif, M.; Kurosaki, T.; Helegbe, G.; Kikuchi, M.; Yanagi, T.; Sasaki, H.; Hirayama, K. Effect of nanoparticle coating on the immunogenicity of plasmid DNA vaccine encoding P. yoelii MSP-1 C-terminal. Vaccine 2011, 29, 3239–3247. [Google Scholar] [CrossRef] [PubMed]
- Cherif, M.S.; Shuaibu, M.N.; Kurosaki, T.; Helegbe, G.K.; Kikuchi, M.; Yanagi, T.; Tsuboi, T.; Sasaki, H.; Hirayama, K. Immunogenicity of novel nanoparticle-coated MSP-1 C-terminus malaria DNA vaccine using different routes of administration. Vaccine 2011, 29, 9038–9050. [Google Scholar] [CrossRef] [PubMed]
- Cherif, M.S.; Shuaibu, M.N.; Kodama, Y.; Kurosaki, T.; Helegbe, G.K.; Kikuchi, M.; Ichinose, A.; Yanagi, T.; Sasaki, H.; Yui, K. Nanoparticle formulation enhanced protective immunity provoked by PYGPI8p-transamidase related protein (PyTAM) DNA vaccine in Plasmodium yoelii malaria model. Vaccine 2014, 32, 1998–2006. [Google Scholar] [CrossRef] [PubMed]
- Sjölander, A.; Lövgren, K.; Ståhl, S.; Åslund, L.; Hansson, M.; Nygren, P.-Å.; Larsson, M.; Hagstedt, M.; Wåhlin, B.; Berzins, K. High antibody responses in rabbits immunized with influenza virus ISCOMs containing a repeated sequence of the Plasmodium falciparum antigen Pf155/RESA. Vaccine 1991, 9, 443–450. [Google Scholar]
- Sjölander, A.; Hansson, M.; Lövgren, K.; Wåhlin, B.; Berzins, K.; Perlmann, P. Immunogenicity in rabbits and monkeys of influenza ISCOMs conjugated with repeated sequences of the Plasmodium falciparum antigen Pf155/RESA. Parasite Immunol. 1993, 15, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Chopra, N.; Biswas, S.; Thomas, B.; Sabhnani, L.; Rao, D. Inducing protective antibodies against ring-infected erythrocyte surface peptide antigen of Plasmodium falciparum using immunostimulating complex (ISCOMs) delivery. Med. Microbiol. Immunol. 2000, 189, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Nawwab Al-Deen, F.; Ma, C.; Xiang, S.D.; Selomulya, C.; Plebanski, M.; Coppel, R.L. On the efficacy of malaria DNA vaccination with magnetic gene vectors. J. Controll. Release 2013, 168, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Parween, S.; Gupta, P.K.; Chauhan, V.S. Induction of humoral immune response against PfMSP-1(19) and PvMSP-1(19) using gold nanoparticles along with alum. Vaccine 2011, 29, 2451–2460. [Google Scholar] [CrossRef] [PubMed]
- Pusic, K.; Xu, H.; Stridiron, A.; Aguilar, Z.; Wang, A.; Hui, G. Blood stage merozoite surface protein conjugated to nanoparticles induce potent parasite inhibitory antibodies. Vaccine 2011, 29, 8898–8908. [Google Scholar] [CrossRef] [PubMed]
- Yandar, N.; Pastorin, G.; Prato, M.; Bianco, A.; Patarroyo, M.E.; Manuel Lozano, J. Immunological profile of a Plasmodium vivax AMA-1 N-terminus peptide-carbon nanotube conjugate in an infected Plasmodium berghei mouse model. Vaccine 2008, 26, 5864–5873. [Google Scholar] [CrossRef] [PubMed]
- Rosas, J.; Hernández, R.; Gascon, A.; Igartua, M.; Guzman, F.; Patarroyo, M.; Pedraz, J. Biodegradable PLGA microspheres as a delivery system for malaria synthetic peptide SPf66. Vaccine 2001, 19, 4445–4451. [Google Scholar] [CrossRef]
- Carcaboso, A.; Hernandez, R.; Igartua, M.; Rosas, J.; Patarroyo, M.; Pedraz, J. Potent, long lasting systemic antibody levels and mixed Th1/Th2 immune response after nasal immunization with malaria antigen loaded PLGA microparticles. Vaccine 2004, 22, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Rosas, J.; Pedraz, J.; Hernandez, R.; Gascon, A.; Igartua, M.; Guzman, F.; Rodrı́guez, R.; Cortes, J.; Patarroyo, M. Remarkably high antibody levels and protection against P. falciparum malaria in Aotus monkeys after a single immunisation of SPf66 encapsulated in PLGA microspheres. Vaccine 2002, 20, 1707–1710. [Google Scholar] [PubMed]
- Carcaboso, A.; Hernández, R.; Igartua, M.; Gascón, A.; Rosas, J.; Patarroyo, M.; Pedraz, J. Immune response after oral administration of the encapsulated malaria synthetic peptide SPf66. Int. J. Pharm. 2003, 260, 273–282. [Google Scholar] [CrossRef]
- Carcaboso, Á.M.; Hernández, R.M.; Igartua, M.; Rosas, J.E.; Patarroyo, M.E.; Pedraz, J.L. Enhancing immunogenicity and reducing dose of microparticulated synthetic vaccines: Single intradermal administration. Pharm. Res. 2004, 21, 121–126. [Google Scholar]
- Mata, E.; Igartua, M.; Hernández, R.M.; Rosas, J.E.; Patarroyo, M.E.; Pedraz, J.L. Comparison of the adjuvanticity of two different delivery systems on the induction of humoral and cellular responses to synthetic peptides. Drug Deliv. 2010, 17, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Mata, E.; Carcaboso, A.; Hernandez, R.; Igartua, M.; Corradin, G.; Pedraz, J. Adjuvant activity of polymer microparticles and montanide ISA 720 on immune responses to Plasmodium falciparum MSP2 long synthetic peptides in mice. Vaccine 2007, 25, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Mata, E.; Igartua, M.; Patarroyo, M.E.; Pedraz, J.L.; Hernández, R.M. Enhancing immunogenicity to PLGA microparticulate systems by incorporation of alginate and RGD-modified alginate. Eur. J. Pharm. Sci. 2011, 44, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Seth, R.K.; Kumar, S.; Ali, R.; Mohan, T.; Biswas, S.; Rao, D. Induction of cell-mediated immune responses to peptide antigens of P. vivax in microparticles using intranasal immunization. Immunol. Investig. 2010, 39, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Seth, R.K.; Babu, J.; Biswas, S.; Rao, D. Induction of mucosal and systemic humoral immune responses in murine system by intranasal immunization with peptide antigens of P. vivax and CpG oligodeoxynucleotide (ODN) in microparticle delivery. Int. Immunopharmacol. 2009, 9, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Goyal, A.K.; Mishra, N.; Khatri, K.; Vaidya, B.; Mehta, A.; Wu, Y.; Vyas, S.P. Development and characterization of novel carrier gel core liposomes based transmission blocking malaria vaccine. J. Controll. Release 2009, 140, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Van Rooijen, N.; van Nieuwmegen, R. Liposomes in immunology: Multilamellar phosphatidylcholine liposomes as a simple, biodegradable and harmless adjuvant without any immunogenic activity of its own. Immunol. Investig. 1980, 9, 243–256. [Google Scholar] [CrossRef]
- Toh, M.-R.; Chiu, G.N.C. Liposomes as sterile preparations and limitations of sterilisation techniques in liposomal manufacturing. Asian J. Pharm. Sci. 2013, 8, 88–95. [Google Scholar] [CrossRef]
- Barenholz, Y. Liposome application: Problems and prospects. Curr. Opin. Colloid Interface Sci. 2001, 6, 66–77. [Google Scholar] [CrossRef]
- Patravale, V.; Prabhu, P. Potential of nanocarriers in antigen delivery: The path to successful vaccine delivery. Nanocarriers 2014. [Google Scholar] [CrossRef]
- Anderson, J.M.; Shive, M.S. Biodegradation and biocompatibility of PLA and PLGA microspheres. Adv. Drug Deliv. Rev. 2012, 64, 72–82. [Google Scholar] [CrossRef]
- Slütter, B.; Bal, S.; Keijzer, C.; Mallants, R.; Hagenaars, N.; Que, I.; Kaijzel, E.; van Eden, W.; Augustijns, P.; Löwik, C.; et al. Nasal vaccination with N-trimethyl chitosan and PLGA based nanoparticles: Nanoparticle characteristics determine quality and strength of the antibody response in mice against the encapsulated antigen. Vaccine 2010, 28, 6282–6291. [Google Scholar] [CrossRef] [PubMed]
- Elamanchili, P.; Lutsiak, C.M.; Hamdy, S.; Diwan, M.; Samuel, J. “Pathogen-mimicking” nanoparticles for vaccine delivery to dendritic cells. J. Immunother. 2007, 30, 378–395. [Google Scholar] [CrossRef] [PubMed]
- Mallapragada, S.K.; Narasimhan, B. Immunomodulatory biomaterials. Int. J. Pharm. 2008, 364, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Loos, C.; Syrovets, T.; Musyanovych, A.; Mailänder, V.; Landfester, K.; Nienhaus, G.U.; Simmet, T. Functionalized polystyrene nanoparticles as a platform for studying bio-nano interactions. Beilstein J. Nanotechnol. 2014, 5, 2403–2412. [Google Scholar] [CrossRef] [PubMed]
- Raman, S.; Machaidze, G.; Lustig, A.; Aebi, U.; Burkhard, P. Structure-based design of peptides that self-assemble into regular polyhedral nanoparticles. Nanomedicine 2006, 2, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Kurosaki, T.; Kitahara, T.; Fumoto, S.; Nishida, K.; Nakamura, J.; Niidome, T.; Kodama, Y.; Nakagawa, H.; To, H.; Sasaki, H. Ternary complexes of pDNA, polyethylenimine, and γ-polyglutamic acid for gene delivery systems. Biomaterials 2009, 30, 2846–2853. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.T.; Brown, L.E.; Deliyannis, G.; Pearse, M.J. ISCOMTM-based vaccines: The second decade. Immunol. Cell Biol. 2005, 83, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.J.; Pedersen, G.; Madhun, A.S.; Svindland, S.; Sævik, M.; Breakwell, L.; Hoschler, K.; Willemsen, M.; Campitelli, L.; Nøstbakken, J.K.; et al. Evaluation of a virosomal H5N1 vaccine formulated with Matrix M™ adjuvant in a phase I clinical trial. Vaccine 2011, 29, 8049–8059. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-X.; Xie, Y.; Ye, Y.-P. ISCOMs and ISCOMATRIX™. Vaccine 2009, 27, 4388–4401. [Google Scholar] [CrossRef] [PubMed]
- Amiji, M.M. Nanotechnology for Cancer Therapy; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2006. [Google Scholar]
- Kunzmann, A.; Andersson, B.; Thurnherr, T.; Krug, H.; Scheynius, A.; Fadeel, B. Toxicology of engineered nanomaterials: Focus on biocompatibility, biodistribution and biodegradation. Biochim. Biophys. Acta 2011, 1810, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Biju, V.; Mundayoor, S.; Omkumar, R.V.; Anas, A.; Ishikawa, M. Bioconjugated quantum dots for cancer research: Present status, prospects and remaining issues. Biotechnol. Adv. 2010, 28, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Ron, H. A toxicologic review of quantum dots: Toxicity depends on physicochemical and environmental factors. Environ. Health Perspect. 2006, 114, 165–172. [Google Scholar]
- Chan, W.C.W.; Nie, S. Quantum dot bioconjugates for ultrasensitive nonisotopic detection. Science 1998, 281, 2016–2018. [Google Scholar] [CrossRef] [PubMed]
- Loo, S.; Moore, T.; Banik, B.; Alexis, F. Biomedical applications of hydroxyapatite nanoparticles. Curr. Pharm. Biotechnol. 2010, 11, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Epple, M.; Ganesan, K.; Heumann, R.; Klesing, J.; Kovtun, A.; Neumann, S.; Sokolova, V. Application of calcium phosphate nanoparticles in biomedicine. J. Mater. Chem. 2010, 20, 18–23. [Google Scholar] [CrossRef]
- Boisselier, E.; Astruc, D. Gold nanoparticles in nanomedicine: Preparations, imaging, diagnostics, therapies and toxicity. Chem. Soc. Rev. 2009, 38, 1759–1782. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ralston, J.; Sedev, R.; Beattie, D.A. Functionalized gold nanoparticles: Synthesis, structure and colloid stability. J. Colloid Interface Sci. 2009, 331, 251–262. [Google Scholar] [CrossRef] [PubMed]
- De Araújo Lopes, S.C.; dos Santos Giuberti, C.; Rocha, T.G.R.; dos Santos Ferreira, D.; Leite, E.A.; Oliveira, M.C. Liposomes as Carriers of Anticancer Drugs. In Cancer Treatment—Conventional and Innovative Approaches; Rangel, L., Ed.; InTech: Rijeka, Croatia, 2013. [Google Scholar]
- Bangham, A.D.; Standish, M.M.; Watkins, J.C. Diffusion of univalent ions across the lamellae of swollen phospholipids. J. Mol. Biol. 1965, 13, 238–252. [Google Scholar] [CrossRef]
- Koo, O.M.; Rubinstein, I.; Onyuksel, H. Role of nanotechnology in targeted drug delivery and imaging: A concise review. Nanomedicine 2005, 1, 193–212. [Google Scholar] [CrossRef] [PubMed]
- Papahadjopoulos, D.; Allen, T.M.; Gabizon, A.; Mayhew, E.; Matthay, K.; Huang, S.K.; Lee, K.D.; Woodle, M.C.; Lasic, D.D.; Redemann, C. Sterically stabilized liposomes: Improvements in pharmacokinetics and antitumor therapeutic efficacy. Proc. Natl. Acad. Sci. USA 1991, 88, 11460–11464. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Nishida, M.; Maekawa, N.; Yamamura, H.; Tanaka, Y.; Kasai, M.; Taneichi, M.; Uchida, T. An increased adjuvanticity of liposomes by the inclusion of phosphatidylserine in immunization with surface-coupled liposomal antigen. Int. Arch. Allergy Immunol. 2005, 136, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.J.; Suh, H.; Bershteyn, A.; Stephan, M.T.; Liu, H.; Huang, B.; Sohail, M.; Luo, S.; Ho Um, S.; Khant, H.; et al. Interbilayer-crosslinked multilamellar vesicles as synthetic vaccines for potent humoral and cellular immune responses. Nat. Mater. 2011, 10, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Kowalczuk, A.; Trzcinska, R.; Trzebicka, B.; Müller, A.H.; Dworak, A.; Tsvetanov, C.B. Loading of polymer nanocarriers: Factors, mechanisms and applications. Prog. Polymer Sci. 2014, 39, 43–86. [Google Scholar] [CrossRef]
- Yilgor, P.; Hasirci, N.; Hasirci, V. Sequential BMP-2/BMP-7 delivery from polyester nanocapsules. J. Biomed. Mater. Res. 2010, 93, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; le Breton, A.; Préat, V. PLGA-based nanoparticles: An overview of biomedical applications. J. Controll. Release 2012, 161, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Betancourt, T.; Byrne, J.D.; Sunaryo, N.; Crowder, S.W.; Kadapakkam, M.; Patel, S.; Casciato, S.; Brannon-Peppas, L. PEGylation strategies for active targeting of PLA/PLGA nanoparticles. J. Biomed. Mater. Res. Part A 2009, 91, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Hamdy, S.; Haddadi, A.; Shayeganpour, A.; Samuel, J.; Lavasanifar, A. Activation of antigen-specific T cell-responses by mannan-decorated PLGA nanoparticles. Pharm. Res. 2011, 28, 2288–2301. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, B.M.; Schwendeman, S.P. Characterization of the initial burst release of a model peptide from poly(d,l-lactide-co-glycolide) microspheres. J. Controll. Release 2002, 82, 289–307. [Google Scholar] [CrossRef]
- Schroeder, U.; Graff, A.; Buchmeier, S.; Rigler, P.; Silvan, U.; Tropel, D.; Jockusch, B.M.; Aebi, U.; Burkhard, P.; Schoenenberger, C.-A. Peptide nanoparticles serve as a powerful platform for the immunogenic display of poorly antigenic actin determinants. J. Mol. Biol. 2009, 386, 1368–1381. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, Y.-H. High epitope density in a single protein molecule significantly enhances antigenicity as well as immunogenicity: A novel strategy for modern vaccine development and a preliminary investigation about B cell discrimination of monomeric proteins. Eur. J. Immunol. 2005, 35, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Dasgupta, D.; Doll, T.A.; Burkhard, P.; Lanar, D.E. Expression, purification and refolding of a self-assembling protein nanoparticle (SAPN) malaria vaccine. Methods 2013, 60, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Scheerlinck, J.-P.Y.; Greenwood, D.L.V. Virus-sized vaccine delivery systems. Drug Discov. Today 2008, 13, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Pearse, M.J.; Drane, D. ISCOMATRIX® adjuvant for antigen delivery. Adv. Drug Deliv. Rev. 2005, 57, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Gupta, M. Synthesis and surface engineering of iron oxide nanoparticles for biomedical applications. Biomaterials 2005, 26, 3995–4021. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.H.; Salabas, E.E.L.; Schüth, F. Magnetic nanoparticles: Synthesis, protection, functionalization, and application. Angew. Chem. Int. Ed. 2007, 46, 1222–1244. [Google Scholar] [CrossRef] [PubMed]
- Alexiou, C.; Arnold, W.; Klein, R.J.; Parak, F.G.; Hulin, P.; Bergemann, C.; Erhardt, W.; Wagenpfeil, S.; Lübbe, A.S. Locoregional cancer treatment with magnetic drug targeting. Cancer Res. 2000, 60, 6641–6648. [Google Scholar] [PubMed]
- Babes, L.; Denizot, B.; Amp, X.; Tanguy, G.; le Jeune, J.J.; Jallet, P. Synthesis of iron oxide nanoparticles used as MRI contrast agents: A parametric study. J. Colloid Interface Sci. 1999, 212, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Forge, D.; Port, M.; Roch, A.; Robic, C.; Vander Elst, L.; Muller, R.N. Magnetic iron oxide nanoparticles: Synthesis, stabilization, vectorization, physicochemical characterizations, and biological applications. Chem. Rev. 2008, 108, 2064–2110. [Google Scholar] [CrossRef] [PubMed]
- Luten, J.; van Nostrum, C.F.; de Smedt, S.C.; Hennink, W.E. Biodegradable polymers as non-viral carriers for plasmid DNA delivery. J. Controll. Release 2008, 126, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Graves Patricia, M.; Gelband, H. Vaccines for preventing malaria (SPf66). In Cochrane Database of Systematic Reviews; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Newman, K.D.; Elamanchili, P.; Kwon, G.S.; Samuel, J. Uptake of poly(d,l-lactic-co-glycolic acid) microspheres by antigen-presenting cells in vivo. J. Biomed. Mater. Res. 2002, 60, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Churcher, T.S.; Blagborough, A.M.; Delves, M.; Ramakrishnan, C.; Kapulu, M.C.; Williams, A.R.; Biswas, S.; Da, D.F.; Cohuet, A.; Sinden, R.E. Measuring the blockade of malaria transmission—An analysis of the standard membrane feeding assay. Int. J. Parasitol. 2012, 42, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Purwada, A.; Roy, K.; Singh, A. Engineering vaccines and niches for immune modulation. Acta Biomater. 2013, 10, 1728–1740. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tan, J.; Thomas, A.; Ou-Yang, D.; Muzykantov, V.R. The shape of things to come: Importance of design in nanotechnology for drug delivery. Ther. Deliv. 2012, 3, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Stellacci, F. Effect of surface properties on nanoparticle-cell interactions. Small 2010, 6, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.P.; Geckeler, K.E. Polymer nanoparticles: Preparation techniques and size-control parameters. Prog. Polymer Sci. 2011, 36, 887–913. [Google Scholar] [CrossRef]
- Byrne, J.; Telling, N.; Coker, V.; Pattrick, R.; van der Laan, G.; Arenholz, E.; Tuna, F.; Lloyd, J. Control of nanoparticle size, reactivity and magnetic properties during the bioproduction of magnetite by Geobacter sulfurreducens. Nanotechnology 2011, 22, 455709. [Google Scholar] [CrossRef] [PubMed]
- Sau, T.K.; Pal, A.; Jana, N.; Wang, Z.; Pal, T. Size controlled synthesis of gold nanoparticles using photochemically prepared seed particles. J. Nanoparticle Res. 2001, 3, 257–261. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Haynes, C.L. Synthesis and characterization of biocompatible and size-tunable multifunctional porous silica nanoparticles. Chem. Mater. 2009, 21, 3979–3986. [Google Scholar] [CrossRef]
- Hou, Y.; Kondoh, H.; Ohta, T.; Gao, S. Size-controlled synthesis of nickel nanoparticles. Appl. Surf. Sci. 2005, 241, 218–222. [Google Scholar] [CrossRef]
- Xiang, S.D.; Scholzen, A.; Minigo, G.; David, C.; Apostolopoulos, V.; Mottram, P.L.; Plebanski, M. Pathogen recognition and development of particulate vaccines: Does size matter? Methods 2006, 40, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Gu, W.; Xu, Z.P. Re-considering how particle size and other properties of antigen-adjuvant complexes impact on the immune responses. J. Colloid Interface Sci. 2013, 395, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fifis, T.; Gamvrellis, A.; Crimeen-Irwin, B.; Pietersz, G.A.; Li, J.; Mottram, P.L.; McKenzie, I.F.C.; Plebanski, M. Size-dependent immunogenicity: Therapeutic and protective properties of nano-vaccines against tumors. J. Immunol. 2004, 173, 3148–3154. [Google Scholar] [CrossRef] [PubMed]
- Oyewumi, M.O.; Kumar, A.; Cui, Z. Nano-microparticles as immune adjuvants: Correlating particle sizes and the resultant immune responses. Expert Rev. Vaccines 2010, 9, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Good, M.F.; Doolan, D.L. Malaria vaccine design: Immunological considerations. Immunity 2010, 33, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Mottram, P.L.; Leong, D.; Crimeen-Irwin, B.; Gloster, S.; Xiang, S.D.; Meanger, J.; Ghildyal, R.; Vardaxis, N.; Plebanski, M. Type 1 and 2 immunity following vaccination is influenced by nanoparticle size: Formulation of a model vaccine for respiratory syncytial virus. Mol. Pharm. 2006, 4, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.T.; Rehor, A.; Schmoekel, H.G.; Hubbell, J.A.; Swartz, M.A. In vivo targeting of dendritic cells in lymph nodes with poly(propylene sulfide) nanoparticles. J. Controll. Release 2006, 112, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Oussoren, C.; Zuidema, J.; Crommelin, D.J.A.; Storm, G. Lymphatic uptake and biodistribution of liposomes after subcutaneous injection: II. Influence of liposomal size, lipid composition and lipid dose. Biochim. Biophys. Acta 1997, 1328, 261–272. [Google Scholar] [CrossRef]
- Kourtis, I.C.; Hirosue, S.; de Titta, A.; Kontos, S.; Stegmann, T.; Hubbell, J.A.; Swartz, M.A. Peripherally administered nanoparticles target monocytic myeloid cells, secondary lymphoid organs and tumors in mice. PLoS ONE 2013, 8, e61646. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.N.; Vokali, E.; Lund, A.W.; Hubbell, J.A.; Swartz, M.A. Targeting the tumor-draining lymph node with adjuvanted nanoparticles reshapes the anti-tumor immune response. Biomaterials 2014, 35, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.T.; van der Vlies, A.J.; Simeoni, E.; Angeli, V.; Randolph, G.J.; O’Neil, C.P.; Lee, L.K.; Swartz, M.A.; Hubbell, J.A. Exploiting lymphatic transport and complement activation in nanoparticle vaccines. Nat. Biotech. 2007, 25, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Manolova, V.; Flace, A.; Bauer, M.; Schwarz, K.; Saudan, P.; Bachmann, M.F. Nanoparticles target distinct dendritic cell populations according to their size. Eur. J. Immunol. 2008, 38, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Gutierro, I.; Hernández, R.M.; Igartua, M.; Gascón, A.R.; Pedraz, J.L. Size dependent immune response after subcutaneous, oral and intranasal administration of BSA loaded nanospheres. Vaccine 2002, 21, 67–77. [Google Scholar] [CrossRef]
- Li, X.; Sloat, B.R.; Yanasarn, N.; Cui, Z. Relationship between the size of nanoparticles and their adjuvant activity: Data from a study with an improved experimental design. Eur. J. Pharm. Biopharm. 2011, 78, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Chithrani, B.D.; Chan, W.C.W. Elucidating the mechanism of cellular uptake and removal of protein-coated gold nanoparticles of different sizes and shapes. Nano Lett. 2007, 7, 1542–1550. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Wu, S.-H.; Hung, Y.; Mou, C.-Y. Size effect on cell uptake in well-suspended, uniform mesoporous silica nanoparticles. Small 2009, 5, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Li, J.; Bao, G.; Zhang, S. Variable nanoparticle-cell adhesion strength regulates cellular uptake. Phys. Rev. Lett. 2010. [Google Scholar] [CrossRef]
- Albanese, A.; Tang, P.S.; Chan, W.C.W. The effect of nanoparticle size, shape, and surface chemistry on biological systems. Annu. Rev. Biomed. Eng. 2012, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, J.; Watanabe, W. Physical and chemical stability of drug nanoparticles. Adv. Drug Deliv. Rev. 2011, 63, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Yeung, T.; Gilbert, G.E.; Shi, J.; Silvius, J.; Kapus, A.; Grinstein, S. Membrane phosphatidylserine regulates surface charge and protein localization. Science 2008, 319, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.D.; Fuchsberger, M.; Karlson, T.D.L.; Hardy, C.L.; Selomulya, C.; Plebanski, M. Nanoparticles, Immunomodulation and Vaccine Delivery. In Handbook of Immunological Properties of Engineered Nanomaterials; Dobrovolskaia, M.A., McNeil, S.E., Eds.; World Scientific Publishing: Hackensack, NJ, USA, 2013; pp. 449–475. [Google Scholar]
- Thomas, C.; Gupta, V.; Ahsan, F. Influence of surface charge of PLGA particles of recombinant hepatitis B surface antigen in enhancing systemic and mucosal immune responses. Int. J. Pharm. 2009, 379, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Henriksen-Lacey, M.; Christensen, D.; Bramwell, V.W.; Lindenstrøm, T.; Agger, E.M.; Andersen, P.; Perrie, Y. Liposomal cationic charge and antigen adsorption are important properties for the efficient deposition of antigen at the injection site and ability of the vaccine to induce a CMI response. J. Controll. Release 2010, 145, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.C.; Xie, J.; Wurm, P.A.; Xia, Y. Understanding the role of surface charges in cellular adsorption versus internalization by selectively removing gold nanoparticles on the cell surface with a I2/KI etchant. Nano Lett. 2009, 9, 1080–1084. [Google Scholar] [CrossRef]
- Villanueva, A.; Canete, M.; Roca, A.G.; Calero, M.; Veintemillas-Verdaguer, S.; Serna, C.J.; del Puerto Morales, M.; Miranda, R. The influence of surface functionalization on the enhanced internalization of magnetic nanoparticles in cancer cells. Nanotechnology 2009. [Google Scholar] [CrossRef] [PubMed]
- Foged, C.; Brodin, B.; Frokjaer, S.; Sundblad, A. Particle size and surface charge affect particle uptake by human dendritic cells in an in vitro model. Int. J. Pharm. 2005, 298, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Badiee, A.; Jaafari, M.R.; Khamesipour, A.; Samiei, A.; Soroush, D.; Kheiri, M.T.; Barkhordari, F.; McMaster, W.R.; Mahboudi, F. The role of liposome charge on immune response generated in BALB/c mice immunized with recombinant major surface glycoprotein of Leishmania (rgp63). Exp. Parasitol. 2009, 121, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Harush-Frenkel, O.; Rozentur, E.; Benita, S.; Altschuler, Y. Surface charge of nanoparticles Determines their endocytic and transcytotic pathway in Polarized MDCK cells. Biomacromolecules 2008, 9, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.T.; Swartz, M.A.; Hubbell, J.A. Targeting dendritic cells with biomaterials: Developing the next generation of vaccines. Trends Immunol. 2006, 27, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Duthie, M.S.; Windish, H.P.; Fox, C.B.; Reed, S.G. Use of defined TLR ligands as adjuvants within human vaccines. Immunol. Rev. 2011, 239, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Demento, S.L.; Siefert, A.L.; Bandyopadhyay, A.; Sharp, F.A.; Fahmy, T.M. Pathogen-associated molecular patterns on biomaterials: A paradigm for engineering new vaccines. Trends Biotechnol. 2011, 29, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.J.; James, E.; Shastri, N.; Fréchet, J.M.J. In vivo targeting of dendritic cells for activation of cellular immunity using vaccine carriers based on pH-responsive microparticles. Proc. Natl. Acad. Sci. USA 2005, 102, 18264–18268. [Google Scholar] [CrossRef] [PubMed]
- Trumpfheller, C.; Finke, J.S.; López, C.B.; Moran, T.M.; Moltedo, B.; Soares, H.; Huang, Y.; Schlesinger, S.J.; Park, C.G.; Nussenzweig, M.C.; et al. Intensified and protective CD4+ T cell immunity in mice with anti-dendritic cell HIV gag fusion antibody vaccine. J. Exp. Med. 2006, 203, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Bonifaz, L.C.; Bonnyay, D.P.; Charalambous, A.; Darguste, D.I.; Fujii, S.-I.; Soares, H.; Brimnes, M.K.; Moltedo, B.; Moran, T.M.; Steinman, R.M. In vivo targeting of antigens to maturing dendritic cells via the DEC-205 receptor improves T cell vaccination. J. Exp. Med. 2004, 199, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; Tacken, P.J.; Fokkink, R.; Joosten, B.; Stuart, M.C.; Albericio, F.; Torensma, R.; Figdor, C.G. Targeted PLGA nano- but not microparticles specifically deliver antigen to human dendritic cells via DC-SIGN in vitro. J. Controll. Release 2010, 144, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Thomann, J.-S.; Heurtault, B.; Weidner, S.; Brayé, M.; Beyrath, J.; Fournel, S.; Schuber, F.; Frisch, B. Antitumor activity of liposomal ErbB2/HER2 epitope peptide-based vaccine constructs incorporating TLR agonists and mannose receptor targeting. Biomaterials 2011, 32, 4574–4583. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; Tacken, P.J.; Fokkink, R.; Figdor, C.G. The influence of PEG chain length and targeting moiety on antibody-mediated delivery of nanoparticle vaccines to human dendritic cells. Biomaterials 2011, 32, 6791–6803. [Google Scholar] [CrossRef] [PubMed]
- Cedervall, T.; Lynch, I.; Lindman, S.; Berggård, T.; Thulin, E.; Nilsson, H.; Dawson, K.A.; Linse, S. Understanding the nanoparticle-protein corona using methods to quantify exchange rates and affinities of proteins for nanoparticles. Proc. Natl. Acad. Sci. USA 2007, 104, 2050–2055. [Google Scholar] [CrossRef] [PubMed]
- Nel, A.E.; Mädler, L.; Velegol, D.; Xia, T.; Hoek, E.M.; Somasundaran, P.; Klaessig, F.; Castranova, V.; Thompson, M. Understanding biophysicochemical interactions at the nano-bio interface. Nature materials 2009, 8, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Jansch, M.; Stumpf, P.; Graf, C.; Rühl, E.; Müller, R.H. Adsorption kinetics of plasma proteins on ultrasmall superparamagnetic iron oxide (USPIO) nanoparticles. Int. J. Pharm. 2012, 428, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Göppert, T.M.; Müller, R.H. Adsorption kinetics of plasma proteins on solid lipid nanoparticles for drug targeting. Int. J. Pharm. 2005, 302, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Saptarshi, S.R.; Duschl, A.; Lopata, A.L. Interaction of nanoparticles with proteins: Relation to bio-reactivity of the nanoparticle. J. Nanobiotechnol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Monopoli, M.P.; Walczyk, D.; Campbell, A.; Elia, G.; Lynch, I.; Baldelli Bombelli, F.; Dawson, K.A. Physical-chemical aspects of protein corona: Relevance to in vitro and in vivo biological impacts of nanoparticles. J. Am. Chem. Soc. 2011, 133, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Klein, J. Probing the interactions of proteins and nanoparticles. Proc. Natl. Acad. Sci. USA 2007, 104, 2029–2030. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tiruppathi, C.; Minshall, R.D.; Malik, A.B. Size and dynamics of caveolae studied using nanoparticles in living endothelial cells. ACS Nano 2009, 3, 4110–4116. [Google Scholar] [CrossRef] [PubMed]
- Fadeel, B.; Garcia-Bennett, A.E. Better safe than sorry: Understanding the toxicological properties of inorganic nanoparticles manufactured for biomedical applications. Adv. Drug Deliv. Rev. 2010, 62, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Seth, A.; Wibowo, N.; Zhao, C.-X.; Mitter, N.; Yu, C.; Middelberg, A.P.J. Nanoparticle vaccines. Vaccine 2014, 32, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Chesko, J.; Kazzaz, J.; Ugozzoli, M.; O’Hagan, D.T.; Singh, M. An investigation of the factors controlling the adsorption of protein antigens to anionic PLG microparticles. J. Pharm. Sci. 2005, 94, 2510–2519. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).