
Degradation of Poliovirus Sabin 2 Genome After Electron Beam Irradiation

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Poliomyelitis Virus Propagation
2.2. Inactivation by Electron Beam Irradiation or Chemicals
2.3. Determination of Residual Infectious Activity
2.4. Determination of D-Antigen
2.5. Quantitative Polymerase Chain Reaction (qPCR)
2.6. Electrochemical Study
2.7. Statistics
3. Results
3.1. Residual Infectious Activity and D-Antigen Content of Irradiated Samples
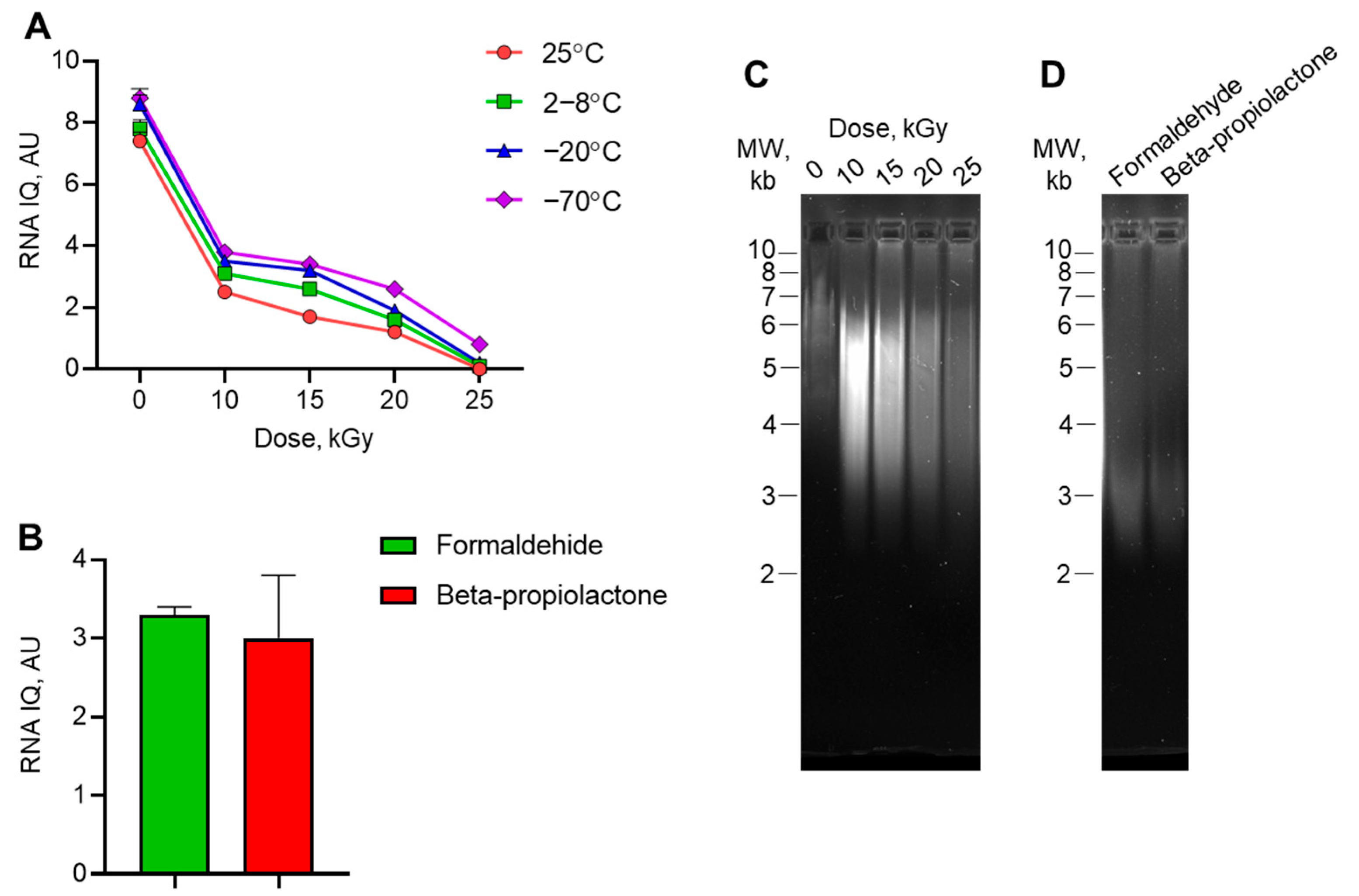
3.2. Quality of RNA Isolated from Irradiated Virus
3.3. 3D Left Segment of Poliovirus Most Sensitive to Accelerated Electrons
3.4. Electroanalysis of Poliovirus RNA Degradation on SPE/SWCNT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Piniaeva, A.; Ignatyev, G.; Kozlovskaya, L.; Ivin, Y.; Kovpak, A.; Ivanov, A.; Shishova, A.; Antonova, L.; Khapchaev, Y.; Feldblium, I.; et al. Immunogenicity and Safety of Inactivated Sabin-Strain Polio Vaccine “PoliovacSin”: Clinical Trials Phase I and II. Vaccines 2021, 9, 565. [Google Scholar] [CrossRef]
- Dowdle, W.R.; De Gourville, E.; Kew, O.M.; Pallansch, M.A.; Wood, D.J. Polio eradication: The OPV paradox. Rev. Med. Virol. 2003, 13, 277–291. [Google Scholar] [CrossRef] [PubMed]
- Polio Eradication Strategy 2022–2026: Executive Summary; World Health Organization: Geneva, Switzerland, 2021.
- Bakker, W.A.M.; Thomassen, Y.E.; van’t Oever, A.G.; Westdijk, J.; van Oijen, M.G.C.T.; Sundermann, L.C.; van’t Veld, P.; Sleeman, E.; van Nimwegen, F.W.; Hamidi, A.; et al. Inactivated polio vaccine development for technology transfer using attenuated Sabin poliovirus strains to shift from Salk-IPV to Sabin-IPV. Vaccine 2011, 29, 7188–7196. [Google Scholar] [CrossRef]
- Sanders, B.; Koldijk, M.; Schuitemaker, H. Inactivated Viral Vaccines. In Vaccine Analysis: Strategies, Principles, and Control; Springer: Berlin/Heidelberg, Germany, 2014; pp. 45–80. [Google Scholar]
- Walter, K.S.; Altamirano, J.; Huang, C.; Carrington, Y.J.; Zhou, F.; Andrews, J.R.; Maldonado, Y. Rapid emergence and transmission of virulence-associated mutations in the oral poliovirus vaccine following vaccination campaigns. NPJ Vaccines 2023, 8, 137. [Google Scholar] [CrossRef]
- Rojas-Bonilla, M.; Coulliette-Salmond, A.; Belgasmi, H.; Wong, K.; Sayyad, L.; Vega, E.; Grimoldi, F.; Oberste, M.S.; Rüttimann, R. Environmental Surveillance for Risk Assessment in the Context of a Phase 2 Clinical Trial of Type 2 Novel Oral Polio Vaccine in Panama. Viruses 2021, 13, 1355. [Google Scholar] [CrossRef]
- Bislava, S.B.; Daja, A.; Oderinde, B.S.; Uzairu, S.M. Prevalence of vaccine-derived poliovirus in sewage waters in Maiduguri, Borno State, Nigeria. Le Infez. Med. 2024, 32, 90–98. [Google Scholar] [CrossRef]
- Wilton, T.; Dunn, G.; Eastwood, D.; Minor, P.D.; Martin, J. Effect of Formaldehyde Inactivation on Poliovirus. J. Virol. 2014, 88, 11955–11964. [Google Scholar] [CrossRef] [PubMed]
- Zhdanov, D.D.; Ivin, Y.Y.; Shishparenok, A.N.; Kraevskiy, S.V.; Kanashenko, S.L.; Agafonova, L.E.; Shumyantseva, V.V.; Gnedenko, O.V.; Pinyaeva, A.N.; Kovpak, A.A.; et al. Perspectives for the creation of a new type of vaccine preparations based on pseudovirus particles using polio vaccine as an example. Biomed. Khim. 2023, 69, 253–280. [Google Scholar] [CrossRef]
- Sabbaghi, A.; Miri, S.M.; Keshavarz, M.; Zargar, M.; Ghaemi, A. Inactivation methods for whole influenza vaccine production. Rev. Med. Virol. 2019, 29, e2074. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.S. Application of radiation technology in vaccines development. Clin. Exp. Vaccine Res. 2015, 4, 145–158. [Google Scholar] [CrossRef]
- Quarleri, J. Poliomyelitis is a current challenge: Long-term sequelae and circulating vaccine-derived poliovirus. GeroScience 2023, 45, 707–717. [Google Scholar] [CrossRef]
- Bahar, M.W.; Porta, C.; Fox, H.; Macadam, A.J.; Fry, E.E.; Stuart, D.I. Mammalian expression of virus-like particles as a proof of principle for next generation polio vaccines. NPJ Vaccines 2021, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Lentz, K.N.; Smith, A.D.; Geisler, S.C.; Cox, S.; Buontempo, P.; Skelton, A.; DeMartino, J.; Rozhon, E.; Schwartz, J.; Girijavallabhan, V.; et al. Structure of poliovirus type 2 Lansing complexed with antiviral agent SCH48973: Comparison of the structural and biological properties of the three poliovirus serotypes. Structure 1997, 5, 961–978. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Kwee, E.J.; Cleveland, M.H.; Cole, K.D.; Lin-Gibson, S.; He, H.-J. Quantitation and integrity evaluation of RNA genome in lentiviral vectors by direct reverse transcription-droplet digital PCR (direct RT-ddPCR). Sci. Rep. 2023, 13, 14470. [Google Scholar] [CrossRef] [PubMed]
- Wurtzer, S.; Duvivier, M.; Accrombessi, H.; Levert, M.; Richard, E.; Moulin, L. Assessing RNA integrity by digital RT-PCR: Influence of extraction, storage, and matrices. Biol. Methods Protoc. 2024, 9, bpae053. [Google Scholar] [CrossRef]
- Burton, J.; Love, H.; Richards, K.; Burton, C.; Summers, S.; Pitman, J.; Easterbrook, L.; Davies, K.; Spencer, P.; Killip, M.; et al. The effect of heat-treatment on SARS-CoV-2 viability and detection. J. Virol. Methods 2021, 290, 114087. [Google Scholar] [CrossRef]
- Sabat, J.; Subhadra, S.; Rath, S.; Ho, L.M.; Kanungo, S.; Panda, S.; Mandal, M.C.; Dash, S.; Pati, S.; Turuk, J. Yielding quality viral RNA by using two different chemistries: A comparative performance study. Biotechniques 2021, 71, 510–515. [Google Scholar] [CrossRef]
- Van Holm, W.; Ghesquière, J.; Boon, N.; Verspecht, T.; Bernaerts, K.; Zayed, N.; Chatzigiannidou, I.; Teughels, W. A Viability Quantitative PCR Dilemma: Are Longer Amplicons Better? Appl. Environ. Microbiol. 2021, 87, e0265320. [Google Scholar] [CrossRef]
- Ogata, H.; Claverie, J.-M. Unique genes in giant viruses: Regular substitution pattern and anomalously short size. Genome Res. 2007, 17, 1353–1361. [Google Scholar] [CrossRef]
- Agafonova, L.E.; Zhdanov, D.D.; Gladilina; Yu, A.; Shisparenok, A.N.; Shumyantseva, V.V. Electrochemical approach for the analysis of DNA degradation in native DNA and apoptotic cells. Heliyon 2023, 10, e25602. [Google Scholar] [CrossRef]
- Sher, M.; Faheem, A.; Asghar, W.; Cinti, S. Nano-engineered screen-printed electrodes: A dynamic tool for detection of viruses. TrAC Trends Anal. Chem. 2021, 143, 116374. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Xin, X.; Su, J.; Song, S. Research Progress of Electrochemical Biosensors for Diseases Detection in China: A Review. Biosensors 2025, 15, 231. [Google Scholar] [CrossRef] [PubMed]
- Brazaca, L.C.; dos Santos, P.L.; de Oliveira, P.R.; Rocha, D.P.; Stefano, J.S.; Kalinke, C.; Abarza Muñoz, R.A.; Bonacin, J.A.; Janegitz, B.C.; Carrilho, E. Biosensing strategies for the electrochemical detection of viruses and viral diseases—A review. Anal. Chim. Acta 2021, 1159, 338384. [Google Scholar] [CrossRef] [PubMed]
- Tamborelli, A.; Mujica, M.L.; Gallay, P.; Vaschetti, V.; Reartes, D.; Delpino, R.; Bravo, L.; Bollo, S.; Rodríguez, M.; Rubianes, M.D.; et al. Biosensing strategies for the detection of SARS-CoV-2 nucleic acids. J. Pharm. Biomed. Anal. 2023, 232, 115370. [Google Scholar] [CrossRef]
- Paleček, E.; Bartošík, M. Electrochemistry of nucleic acids. Chem. Rev. 2012, 112, 3427–3481. [Google Scholar] [CrossRef]
- Ferapontova, E.E. DNA Electrochemistry and Electrochemical Sensors for Nucleic Acids. Annu. Rev. Anal. Chem. Palo Alto. Calif 2018, 11, 197–218. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Pronina, V.V.; Bulko, T.V.; Agafonova, L.E. Electroanalysis in Pharmacogenomic Studies: Mechanisms of Drug Interaction with DNA. Biochemistry 2024, 89, S224–S233. [Google Scholar] [CrossRef]
- Agafonova, L.; Tikhonova, E.; Sanzhakov, M.; Kostryukova, L.; Shumyantseva, V. Electrochemical Studies of the Interaction of Phospholipid Nanoparticles with dsDNA. Processes 2022, 10, 2324. [Google Scholar] [CrossRef]
- Shumyantseva, V.V.; Berezhnova, A.V.; Agafonova, L.E.; Bulko, T.V.; Veselovsky, A. V Electrochemical Analysis of the Interaction between DNA and Abiraterone D4A Metabolite. J. Anal. Chem. 2024, 79, 873–882. [Google Scholar] [CrossRef]
- Thomassen, Y.E.; Welle, J.; van Eikenhorst, G.; van der Pol, L.A.; Bakker, W.A.M. Transfer of an adherent Vero cell culture method between two different rocking motion type bioreactors with respect to cell growth and metabolic rates. Process Biochem. 2012, 47, 288–296. [Google Scholar] [CrossRef]
- Jiang, S.D.; Pye, D.; Cox, J.C. Inactivation of poliovirus with beta-propiolactone. J. Biol. Stand. 1986, 14, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Ozer, T.; Geiss, B.J.; Henry, C.S. Review-Chemical and Biological Sensors for Viral Detection. J. Electrochem. Soc. 2020, 167, 37523. [Google Scholar] [CrossRef] [PubMed]
- Stempkowska, I.; Ligaj, M.; Jasnowska, J.; Langer, J.; Filipiak, M. Electrochemical response of oligonucleotides on carbon paste electrode. Bioelectrochemistry 2007, 70, 488–494. [Google Scholar] [CrossRef]
- Brabec, V.; Koudelka, J. Oxidation of deoxyribonucleic acid at carbon electrodes. The effect of the quality of the deoxyribonucleic acid sample. J. Electroanal. Chem. Interfacial Electrochem. 1980, 116, 793–805. [Google Scholar] [CrossRef]
- Morozova, O.V.; Bakhvalova, V.N.; Potapova, O.F.; Grishechkin, A.E.; Isaeva, E.I.; Aldarov, K.V.; Klinov, D.V.; Vorovich, M.F. Evaluation of immune response and protective effect of four vaccines against the tick-borne encephalitis virus. Vaccine 2014, 32, 3101–3106. [Google Scholar] [CrossRef]
- Werzberger, A.; Mensch, B.; Kuter, B.; Brown, L.; Lewis, J.; Sitrin, R.; Miller, W.; Shouval, D.; Wiens, B.; Calandra, G. A controlled trial of a formalin-inactivated hepatitis A vaccine in healthy children. N. Engl. J. Med. 1992, 327, 453–457. [Google Scholar] [CrossRef]
- Khoshnood, S.; Arshadi, M.; Akrami, S.; Koupaei, M.; Ghahramanpour, H.; Shariati, A.; Sadeghifard, N.; Heidary, M. An overview on inactivated and live-attenuated SARS-CoV-2 vaccines. J. Clin. Lab. Anal. 2022, 36, e24418. [Google Scholar] [CrossRef]
- Bhatia, S.S.; Pillai, S.D. Ionizing Radiation Technologies for Vaccine Development—A Mini Review. Front. Immunol. 2022, 13, 845514. [Google Scholar] [CrossRef]
- Lawrence, R.M.; Zook, J.D.; Hogue, B.G. Full inactivation of alphaviruses in single particle and crystallized forms. J. Virol. Methods 2016, 236, 237–244. [Google Scholar] [CrossRef]
- Elveborg, S.; Monteil, V.; Mirazimi, A. Methods of Inactivation of Highly Pathogenic Viruses for Molecular, Serology or Vaccine Development Purposes. Pathogens 2022, 11, 271. [Google Scholar] [CrossRef] [PubMed]
- Howley, P.M.; Knipe, D.M.; Whelan, S.P.J. Fields Virology: Emerging Viruses, 7th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2021; ISBN 978-1-975112-54-7. [Google Scholar]
- Comas-Garcia, M. Packaging of Genomic RNA in Positive-Sense Single-Stranded RNA Viruses: A Complex Story. Viruses 2019, 11, 253. [Google Scholar] [CrossRef] [PubMed]
- Grieb, T.; Forng, R.-Y.; Brown, R.; Owolabi, T.; Maddox, E.; Mcbain, A.; Drohan, W.N.; Mann, D.M.; Burgess, W.H. Effective use of Gamma Irradiation for Pathogen Inactivation of Monoclonal Antibody Preparations. Biologicals 2002, 30, 207–216. [Google Scholar] [CrossRef]
- Ranawat, P.; Rawat, S. Radiation resistance in thermophiles: Mechanisms and applications. World J. Microbiol. Biotechnol. 2017, 33, 112. [Google Scholar] [CrossRef]
- Song, Y.; Gorbatsevych, O.; Liu, Y.; Mugavero, J.; Shen, S.H.; Ward, C.B.; Asare, E.; Jiang, P.; Paul, A.V.; Mueller, S.; et al. Limits of variation, specific infectivity, and genome packaging of massively recoded poliovirus genomes. Proc. Natl. Acad. Sci. USA 2017, 114, E8731–E8740. [Google Scholar] [CrossRef]
- Venkatesh, A.G.; Brickner, H.; Looney, D.; Hall, D.A.; Aronoff-Spencer, E. Clinical detection of Hepatitis C viral infection by yeast-secreted HCV-core: Gold-binding-peptide. Biosens. Bioelectron. 2018, 119, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Sha, Z.; Xu, J.; Wang, Y.; Ma, L.; Li, X.; Chen, Y.; Gu, H.-Y. Direct electrochemistry & enzyme characterization of fresh tobacco RNA. J. Electroanal. Chem. 2023, 931, 117156. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Region Name | Sense Primer (5′-3′) | Antisense Primer (5′-3′) | Product Size, bp | Start Position |
|---|---|---|---|---|---|
| 1 | CloSpa | aacagctctggggttgtacc | tggtttcgtgcttctaagttgc | 118 | 5 |
| 2 | IRES | tccccggtgacattgcatag | caaagtagtcggttccgcca | 373 | 176 |
| 3 | VP4 | gcgcccaagtttcatcacag | tttagcatgggagcggtctta | 202 | 752 |
| 4 | VP2 left | tccccaaacattgaggcgtg | tctggaactgcaaacacccc | 386 | 955 |
| 5 | VP2 right | agaatgcgaatccaggcgaa | tggcacagtgatgttgcgta | 371 | 1388 |
| 6 | VP3 left | gtaaccagtacctgaccgca | gagagacacaagatcggcgt | 229 | 1796 |
| 7 | VP3 right | ttttgcggctcaatgatggc | tatctcgcagtaagcgcaca | 325 | 2125 |
| 8 | VP1 left | tggtgacatgattgaggggg | gactctgatcgcgttcgtct | 219 | 2486 |
| 9 | VP1 middle | cggacatgcattgaaccaagt | attcccacgtagggcactga | 162 | 2922 |
| 10 | VP1 right | gtaccactagcgggtcaagc | cccttttctggtagtggggt | 261 | 3142 |
| 11 | 2A | gcaaaatgccgtgagtgtca | ctgctccatagcttcctcctc | 367 | 3465 |
| 12 | 2B | tgagtcacttggtgctgcat | ttctttagccactgccacgg | 231 | 3849 |
| 13 | 2C left | ctgcaaagggactggagtgg | agtggtgcaaacctcttgga | 237 | 4163 |
| 14 | 2C middle | gagcaagcaccgtattgagc | tgcctttctcttctagcgagg | 287 | 4467 |
| 15 | 2C right | accaactccagtcggatcac | aactgaattgccttgccaca | 212 | 4786 |
| 16 | 3A | ccctccggagtgtatcaacg | gcccagcgaacagcttgta | 209 | 5157 |
| 17 | 3C | aggccctgggtttgattacg | ggtttgtcgtccaccgagat | 397 | 5436 |
| 18 | 3D left | aaccaaacttgaacccagcg | tccgcatttccttggtgtct | 362 | 6051 |
| 19 | 3D middle | gtggagcagggaaaatccag | ggcatgccgccttttacg | 359 | 6487 |
| 20 | 3D right | tacccccatgaggttgatgc | acggcggtacaatgtagagt | 369 | 6985 |
| Dose, kGy | Temperature, °C | |||
|---|---|---|---|---|
| 25 | 2–8 | −20 | −70 | |
| 0 | 100% | 100% | 100% | 100% |
| 10 | 0 | ND | ND | ND |
| 15 | 0 | ND | ND | ND |
| 20 | 0 | ND | ND | ND |
| 25 | 0 | 27.5 ± 3.0 | 46.3 ± 5.2% | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhdanov, D.D.; Shishparenok, A.N.; Ivin, Y.Y.; Kovpak, A.A.; Piniaeva, A.N.; Levin, I.V.; Budnik, S.V.; Shilov, O.A.; Churyukin, R.S.; Agafonova, L.E.; et al. Degradation of Poliovirus Sabin 2 Genome After Electron Beam Irradiation. Vaccines 2025, 13, 824. https://doi.org/10.3390/vaccines13080824
Zhdanov DD, Shishparenok AN, Ivin YY, Kovpak AA, Piniaeva AN, Levin IV, Budnik SV, Shilov OA, Churyukin RS, Agafonova LE, et al. Degradation of Poliovirus Sabin 2 Genome After Electron Beam Irradiation. Vaccines. 2025; 13(8):824. https://doi.org/10.3390/vaccines13080824
Chicago/Turabian StyleZhdanov, Dmitry D., Anastasia N. Shishparenok, Yury Y. Ivin, Anastasia A. Kovpak, Anastasia N. Piniaeva, Igor V. Levin, Sergei V. Budnik, Oleg A. Shilov, Roman S. Churyukin, Lubov E. Agafonova, and et al. 2025. "Degradation of Poliovirus Sabin 2 Genome After Electron Beam Irradiation" Vaccines 13, no. 8: 824. https://doi.org/10.3390/vaccines13080824
APA StyleZhdanov, D. D., Shishparenok, A. N., Ivin, Y. Y., Kovpak, A. A., Piniaeva, A. N., Levin, I. V., Budnik, S. V., Shilov, O. A., Churyukin, R. S., Agafonova, L. E., Berezhnova, A. V., Shumyantseva, V. V., & Ishmukhametov, A. A. (2025). Degradation of Poliovirus Sabin 2 Genome After Electron Beam Irradiation. Vaccines, 13(8), 824. https://doi.org/10.3390/vaccines13080824