
ISG15 as a Potent Immune Adjuvant in MVA-Based Vaccines Against Zika Virus and SARS-CoV-2

 , , , , , , , , ,
, , , , , , , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Development of Plasmids Expressing Secretion-Defective ISG15 Mutants
2.3. Ethics Statement
2.4. Analysis of the Impact of ISG15 Variant Expression on Immune Cell Recruitment in Mice by Flow Cytometry
2.5. Co-Infection Assays
2.6. Protein and RNA Extraction and Quantification
2.7. SDS-PAGE and Western Bloting Analysis
2.8. Reverse Transcription and Quantitative PCR (RT-qPCR)
2.9. Immunization Protocols and Evaluation of Vaccine-Induced Immune Responses in Mice
2.9.1. ZIKV Vaccine Immunization and Immune Response Assessment
2.9.2. SARS-CoV-2 Vaccine Immunization and Immune Response Assessment
2.10. Data Analysis and Statistics
3. Results
3.1. Platform-Dependent Modulation of Early Innate Immunity by ISG15 Variants
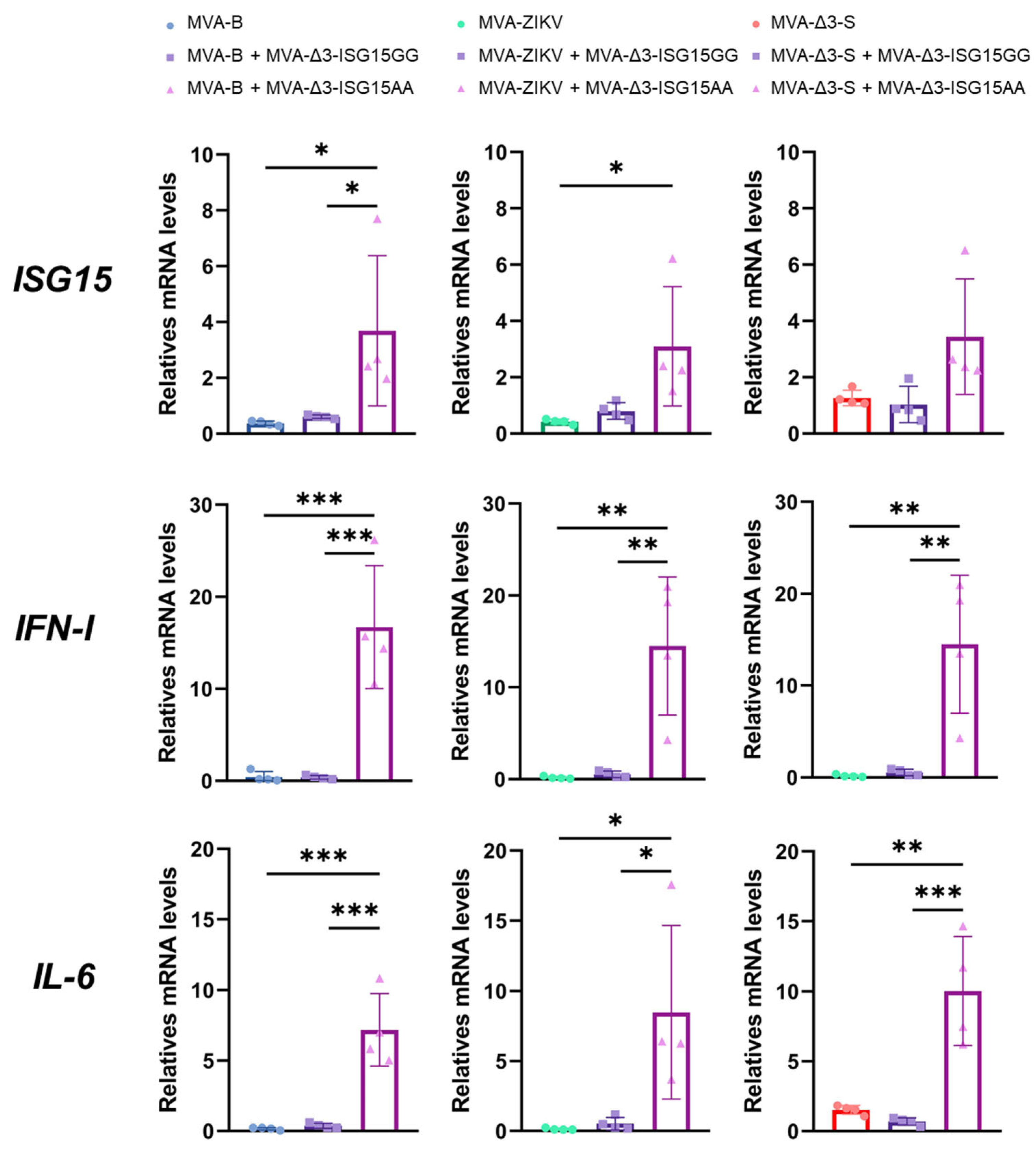
3.2. MVA-Δ3-ISG15AA Enhances Inflammatory Cytokine Responses During Co-Infection with MVA-Based Vaccine Vectors
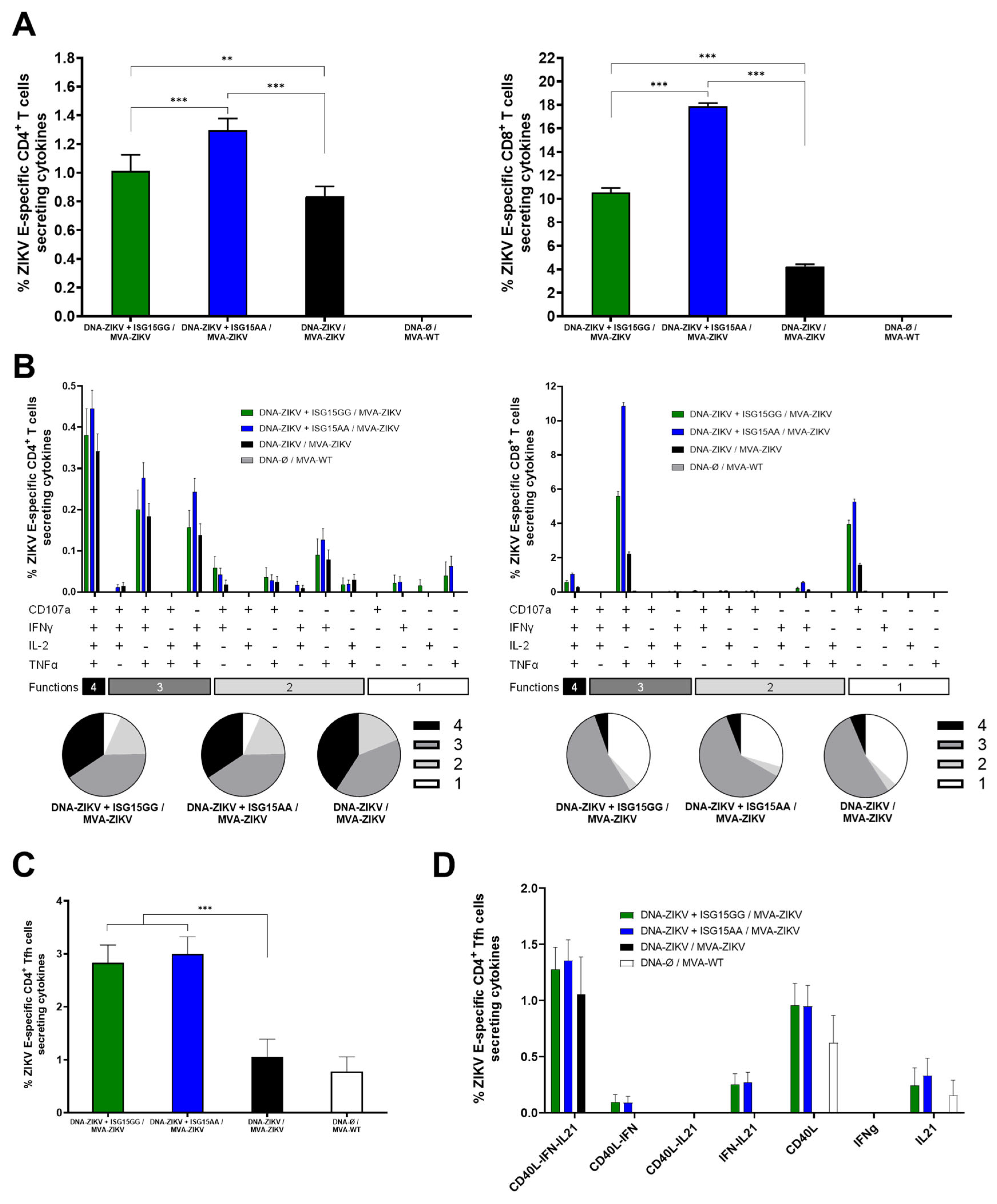
3.3. DNA-ISG15AA Co-Administration Enhances ZIKV-Specific CD4 and CD8 T-Cell Responses in Immunized Mice
3.4. Co-Administration of MVA-Δ3-ISG15 Enhances SARS-CoV-2-Specific T-Cell Responses Induced by MVA-Δ3-S Vaccination in Immunized Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Facciolà, A.; Visalli, G.; Laganà, A.; Di Pietro, A. An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives. Vaccines 2022, 10, 819. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Fox, C.B. New Generation Adjuvants—From Empiricism to Rational Design. Vaccine 2015, 33, B14–B20. [Google Scholar] [CrossRef] [PubMed]
- Berzofsky, J.A.; Ahlers, J.D.; Belyakov, I.M. Strategies for Designing and Optimizing New Generation Vaccines. Nat. Rev. Immunol. 2001, 1, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.R.; Wloch, M.K.; Ye, M.; Reyes, L.R.; Boutsaboualoy, S.; Dunne, C.E.; Chaplin, J.A.; Rusalov, D.; Rolland, A.P.; Fisher, C.L.; et al. Phase 1 Clinical Trials of the Safety and Immunogenicity of Adjuvanted Plasmid DNA Vaccines Encoding Influenza A Virus H5 Hemagglutinin. Vaccine 2010, 28, 2565–2572. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, T.; Wang, Z.; Xu, X. Augmented Immunogenicity of the HPV16 DNA Vaccine via Dual Adjuvant Approach: Integration of CpG ODN into Plasmid Backbone and Co-Administration with IL-28B Gene Adjuvant. Virol. J. 2025, 22, 3. [Google Scholar] [CrossRef]
- Perng, Y.-C.; Lenschow, D.J. ISG15 in Antiviral Immunity and Beyond. Nat. Rev. Microbiol. 2018, 16, 423–439. [Google Scholar] [CrossRef]
- Dos Santos, P.F.; Mansur, D.S. Beyond ISGlylation: Functions of Free Intracellular and Extracellular ISG15. J. Interferon Cytokine Res. 2017, 37, 246–253. [Google Scholar] [CrossRef]
- Swaim, C.D.; Scott, A.F.; Canadeo, L.A.; Huibregtse, J.M. Extracellular ISG15 Signals Cytokine Secretion Through the LFA-1 Integrin Receptor. Mol. Cell 2017, 68, 581–590.e5. [Google Scholar] [CrossRef]
- Owhashi, M.; Taoka, Y.; Ishii, K.; Nakazawa, S.; Uemura, H.; Kambara, H. Identification of a Ubiquitin Family Protein as a Novel Neutrophil Chemotactic Factor. Biochem. Biophys. Res. Commun. 2003, 309, 533–539. [Google Scholar] [CrossRef]
- Napolitano, A.; van der Veen, A.G.; Bunyan, M.; Borg, A.; Frith, D.; Howell, S.; Kjaer, S.; Beling, A.; Snijders, A.P.; Knobeloch, K.-P.; et al. Cysteine-Reactive Free ISG15 Generates IL-1β–Producing CD8α+ Dendritic Cells at the Site of Infection. J. Immunol. 2018, 201, 604–614. [Google Scholar] [CrossRef]
- Álvarez, E.; Falqui, M.; Sin, L.; McGrail, J.P.; Perdiguero, B.; Coloma, R.; Marcos-Villar, L.; Tárrega, C.; Esteban, M.; Gómez, C.E.; et al. Unveiling the Multifaceted Roles of ISG15: From Immunomodulation to Therapeutic Frontiers. Vaccines 2024, 12, 153. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, D.O.; Wise, M.C.; Siefert, R.J.; Yan, J.; Wood, L.M.; Weiner, D.B. Ubiquitin-like Molecule ISG15 Acts as an Immune Adjuvant to Enhance Antigen-Specific CD8 T-Cell Tumor Immunity. Mol. Ther. 2015, 23, 1653–1662. [Google Scholar] [CrossRef]
- Volz, A.; Sutter, G. Modified Vaccinia Virus Ankara: History, Value in Basic Research, and Current Perspectives for Vaccine Development. Adv. Virus Res. 2017, 97, 187–243. [Google Scholar] [CrossRef]
- Nave, L.; Margalit, I.; Tau, N.; Cohen, I.; Yelin, D.; Lienert, F.; Yahav, D. Immunogenicity and Safety of Modified Vaccinia Ankara (MVA) Vaccine—A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Vaccines 2023, 11, 1410. [Google Scholar] [CrossRef] [PubMed]
- Verheust, C.; Goossens, M.; Pauwels, K.; Breyer, D. Biosafety Aspects of Modified Vaccinia Virus Ankara (MVA)-Based Vectors Used for Gene Therapy or Vaccination. Vaccine 2012, 30, 2623–2632. [Google Scholar] [CrossRef] [PubMed]
- Orlova, O.V.; Glazkova, D.V.; Bogoslovskaya, E.V.; Shipulin, G.A.; Yudin, S.M. Development of Modified Vaccinia Virus Ankara-Based Vaccines: Advantages and Applications. Vaccines 2022, 10, 1516. [Google Scholar] [CrossRef]
- Lynch, H.E.; Ray, C.A.; Oie, K.L.; Pollara, J.J.; Petty, I.T.D.; Sadler, A.J.; Williams, B.R.G.; Pickup, D.J. Modified Vaccinia Virus Ankara Can Activate NF-κB Transcription Factors through a Double-Stranded RNA-Activated Protein Kinase (PKR)-Dependent Pathway during the Early Phase of Virus Replication. Virology 2009, 391, 177–186. [Google Scholar] [CrossRef]
- Capone, S.; Brown, A.; Hartnell, F.; Sorbo, M.D.; Traboni, C.; Vassilev, V.; Colloca, S.; Nicosia, A.; Cortese, R.; Folgori, A.; et al. Optimising T Cell (Re)Boosting Strategies for Adenoviral and Modified Vaccinia Ankara Vaccine Regimens in Humans. npj Vaccines 2020, 5, 94. [Google Scholar] [CrossRef]
- Kardani, K.; Bolhassani, A.; Shahbazi, S. Prime-Boost Vaccine Strategy against Viral Infections: Mechanisms and Benefits. Vaccine 2016, 34, 413–423. [Google Scholar] [CrossRef]
- Falivene, J.; Zajac, M.P.D.M.; Pascutti, M.F.; Rodríguez, A.M.; Maeto, C.; Perdiguero, B.; Gómez, C.E.; Esteban, M.; Calamante, G.; Gherardi, M.M. Improving the MVA Vaccine Potential by Deleting the Viral Gene Coding for the IL-18 Binding Protein. PLoS ONE 2012, 7, e32220. [Google Scholar] [CrossRef]
- Alharbi, N.K.; Spencer, A.J.; Hill, A.V.S.; Gilbert, S.C. Deletion of Fifteen Open Reading Frames from Modified Vaccinia Virus Ankara Fails to Improve Immunogenicity. PLoS ONE 2015, 10, e0128626. [Google Scholar] [CrossRef] [PubMed]
- Laghlali, G.; Wiest, M.J.; Karadag, D.; Warang, P.; O’Konek, J.J.; Chang, L.A.; Park, S.; Farazuddin, M.; Landers, J.J.; Janczak, K.W.; et al. Enhanced Mucosal B- and T-Cell Responses against SARS-CoV-2 after Heterologous Intramuscular mRNA Prime/Intranasal Protein Boost Vaccination with a Combination Adjuvant. bioRxiv 2024. bioRxiv:2024.03.28.587260. [Google Scholar] [CrossRef]
- Kolibab, K.; Yang, A.; Derrick, S.C.; Waldmann, T.A.; Perera, L.P.; Morris, S.L. Highly Persistent and Effective Prime/Boost Regimens against Tuberculosis That Use a Multivalent Modified Vaccine Virus Ankara-Based Tuberculosis Vaccine with Interleukin-15 as a Molecular Adjuvant. Clin. Vaccine Immunol. 2010, 17, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Falqui, M.; Perdiguero, B.; Coloma, R.; Albert, M.; Marcos-Villar, L.; McGrail, J.P.; Sorzano, C.Ó.S.; Esteban, M.; Gómez, C.E.; Guerra, S. An MVA-Based Vector Expressing Cell-Free ISG15 Increases IFN-I Production and Improves HIV-1-Specific CD8 T Cell Immune Responses. Front. Cell Infect. Microbiol. 2023, 13, 1187193. [Google Scholar] [CrossRef]
- Perdiguero, B.; Álvarez, E.; Marcos-Villar, L.; Sin, L.; López-Bravo, M.; Valverde, J.R.; Sorzano, C.Ó.S.; Falqui, M.; Coloma, R.; Esteban, M.; et al. B and T Cell Bi-Cistronic Multiepitopic Vaccine Induces Broad Immunogenicity and Provides Protection Against SARS-CoV-2. Vaccines 2024, 12, 1213. [Google Scholar] [CrossRef]
- De Nardo, D.; Kalvakolanu, D.V.; Latz, E. Immortalization of Murine Bone Marrow-Derived Macrophages. Methods Mol. Biol. 2018, 1784, 35–49. [Google Scholar] [CrossRef]
- Gómez, C.E.; Nájera, J.L.; Jiménez, E.P.; Jiménez, V.; Wagner, R.; Graf, M.; Frachette, M.-J.; Liljeström, P.; Pantaleo, G.; Esteban, M. Head-to-Head Comparison on the Immunogenicity of Two HIV/AIDS Vaccine Candidates Based on the Attenuated Poxvirus Strains MVA and NYVAC Co-Expressing in a Single Locus the HIV-1BX08 Gp120 and HIV-1(IIIB) Gag-Pol-Nef Proteins of Clade B. Vaccine 2007, 25, 2863–2885. [Google Scholar] [CrossRef]
- Pérez, P.; Marín, M.Q.; Lázaro-Frías, A.; Jiménez de Oya, N.; Blázquez, A.-B.; Escribano-Romero, E.; Sorzano, C.Ó.S.; Ortego, J.; Saiz, J.-C.; Esteban, M.; et al. A Vaccine Based on a Modified Vaccinia Virus Ankara Vector Expressing Zika Virus Structural Proteins Controls Zika Virus Replication in Mice. Sci. Rep. 2018, 8, 17385. [Google Scholar] [CrossRef]
- García-Arriaza, J.; Garaigorta, U.; Pérez, P.; Lázaro-Frías, A.; Zamora, C.; Gastaminza, P.; Del Fresno, C.; Casasnovas, J.M.; Sorzano, C.Ó.S.; Sancho, D.; et al. COVID-19 Vaccine Candidates Based on Modified Vaccinia Virus Ankara Expressing the SARS-CoV-2 Spike Induce Robust T- and B-Cell Immune Responses and Full Efficacy in Mice. J. Virol. 2021, 95, JVI.02260-20. [Google Scholar] [CrossRef]
- Southern, J.A.; Young, D.F.; Heaney, F.; Baumgärtner, W.K.; Randall, R.E. Identification of an Epitope on the P and V Proteins of Simian Virus 5 That Distinguishes between Two Isolates with Different Biological Characteristics. J. Gen. Virol. 1991, 72, 1551–1557. [Google Scholar] [CrossRef]
- Sun, H.; Chen, X.; Wang, Y. Strategies for the Modification of Vaccinia Virus towards a Better Vaccine Vector. Zoonoses 2025, 5, 994. [Google Scholar] [CrossRef]
- Swaim, C.D.; Canadeo, L.A.; Monte, K.J.; Khanna, S.; Lenschow, D.J.; Huibregtse, J.M. Modulation of Extracellular ISG15 Signaling by Pathogens and Viral Effector Proteins. Cell Rep. 2020, 31, 107772. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Saeed, A.F.U.H.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G.G.; Rao, L.; Duo, Y. Macrophages in Immunoregulation and Therapeutics. Signal Transduct. Target. Ther. 2023, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- Munnur, D.; Teo, Q.; Eggermont, D.; Lee, H.H.Y.; Thery, F.; Ho, J.; van Leur, S.W.; Ng, W.W.S.; Siu, L.Y.L.; Beling, A.; et al. Altered ISGylation Drives Aberrant Macrophage-Dependent Immune Responses during SARS-CoV-2 Infection. Nat. Immunol. 2021, 22, 1416–1427. [Google Scholar] [CrossRef]
- Gómez, C.E.; Perdiguero, B.; Falqui, M.; Marín, M.Q.; Bécares, M.; Sorzano, C.Ó.S.; García-Arriaza, J.; Esteban, M.; Guerra, S. Enhancement of HIV-1 Env-Specific CD8 T Cell Responses Using Interferon-Stimulated Gene 15 as an Immune Adjuvant. J. Virol. 2020, 95, e01155-20. [Google Scholar] [CrossRef]
- Ryan, S.J.; Carlson, C.J.; Tesla, B.; Bonds, M.H.; Ngonghala, C.N.; Mordecai, E.A.; Johnson, L.R.; Murdock, C.C. Warming Temperatures Could Expose More than 1.3 Billion New People to Zika Virus Risk by 2050. Glob. Change Biol. 2021, 27, 84–93. [Google Scholar] [CrossRef]
- Pollard, A.J.; Bijker, E.M. A Guide to Vaccinology: From Basic Principles to New Developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Bogunovic, D.; Byun, M.; Durfee, L.A.; Abhyankar, A.; Sanal, O.; Mansouri, D.; Salem, S.; Radovanovic, I.; Grant, A.V.; Adimi, P.; et al. Mycobacterial Disease and Impaired IFN-γ Immunity in Humans with Inherited ISG15 Deficiency. Science 2012, 337, 1684–1688. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Arriaza, J.; Falqui, M.; Pérez, P.; Coloma, R.; Perdiguero, B.; Álvarez, E.; Marcos-Villar, L.; Astorgano, D.; Campaña-Gómez, I.; Sorzano, C.Ó.S.; et al. ISG15 as a Potent Immune Adjuvant in MVA-Based Vaccines Against Zika Virus and SARS-CoV-2. Vaccines 2025, 13, 696. https://doi.org/10.3390/vaccines13070696
García-Arriaza J, Falqui M, Pérez P, Coloma R, Perdiguero B, Álvarez E, Marcos-Villar L, Astorgano D, Campaña-Gómez I, Sorzano CÓS, et al. ISG15 as a Potent Immune Adjuvant in MVA-Based Vaccines Against Zika Virus and SARS-CoV-2. Vaccines. 2025; 13(7):696. https://doi.org/10.3390/vaccines13070696
Chicago/Turabian StyleGarcía-Arriaza, Juan, Michela Falqui, Patricia Pérez, Rocío Coloma, Beatriz Perdiguero, Enrique Álvarez, Laura Marcos-Villar, David Astorgano, Irene Campaña-Gómez, Carlos Óscar S. Sorzano, and et al. 2025. "ISG15 as a Potent Immune Adjuvant in MVA-Based Vaccines Against Zika Virus and SARS-CoV-2" Vaccines 13, no. 7: 696. https://doi.org/10.3390/vaccines13070696
APA StyleGarcía-Arriaza, J., Falqui, M., Pérez, P., Coloma, R., Perdiguero, B., Álvarez, E., Marcos-Villar, L., Astorgano, D., Campaña-Gómez, I., Sorzano, C. Ó. S., Esteban, M., Gómez, C. E., & Guerra, S. (2025). ISG15 as a Potent Immune Adjuvant in MVA-Based Vaccines Against Zika Virus and SARS-CoV-2. Vaccines, 13(7), 696. https://doi.org/10.3390/vaccines13070696