
The aroA and luxS Double-Gene Mutant Strain Has Potential to Be a Live Attenuated Vaccine against Salmonella Typhimurium


 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Strains, Primers, Plasmids and Cell Lines
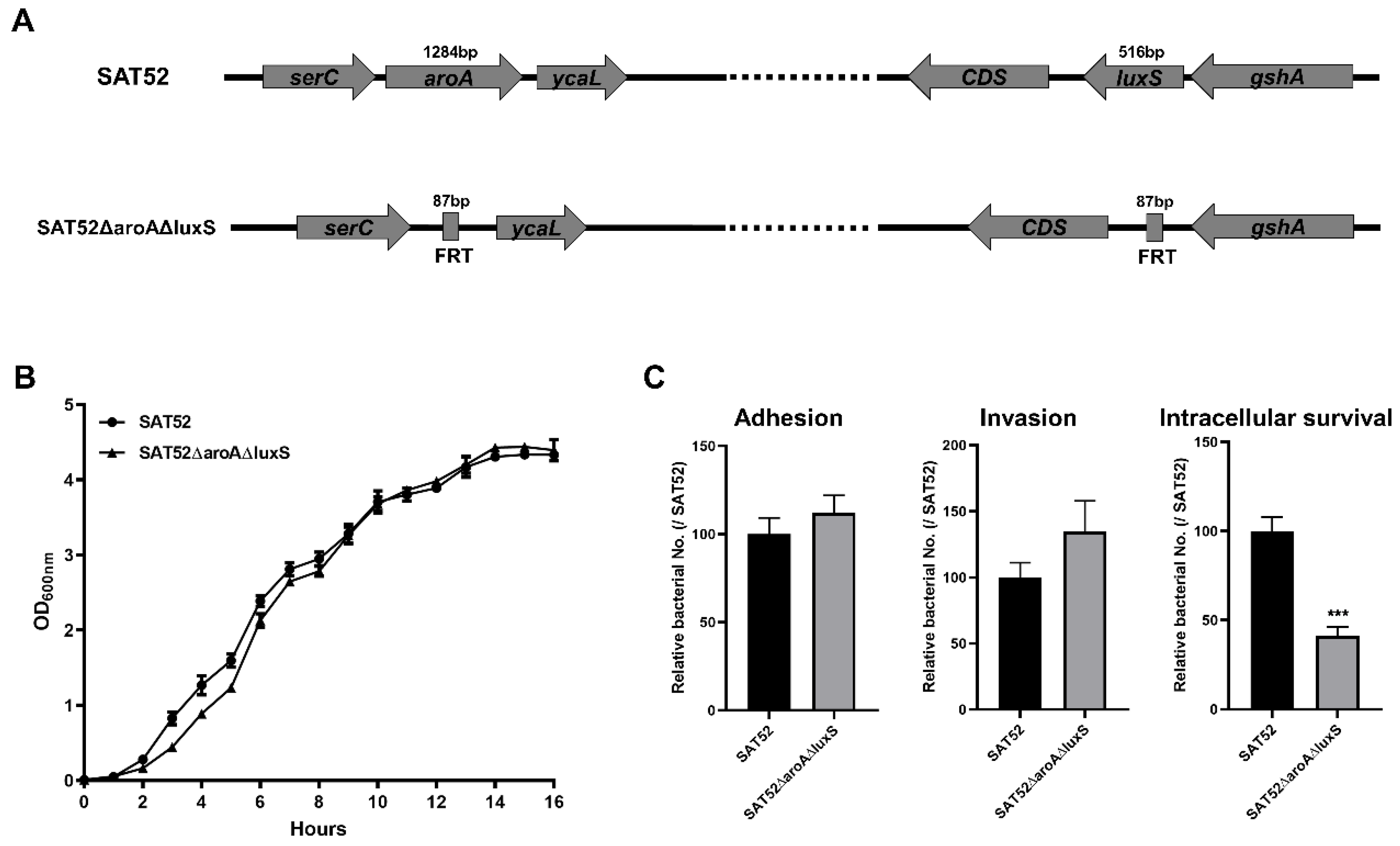
2.3. Construction of Mutant Strain SAT52ΔaroAΔluxS
2.4. Growth Assay
2.5. Bacterial Infection In Vitro
2.6. LD50 Determination
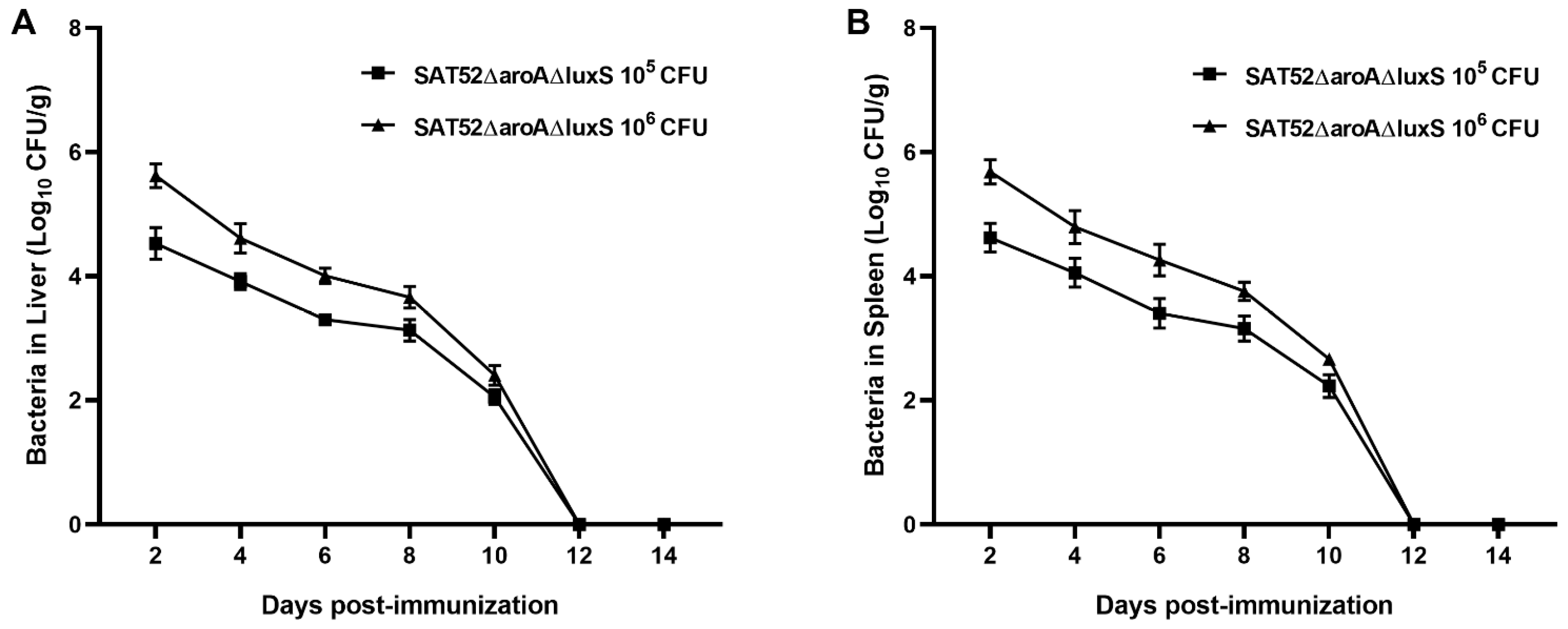
2.7. In Vivo Persistence Assay
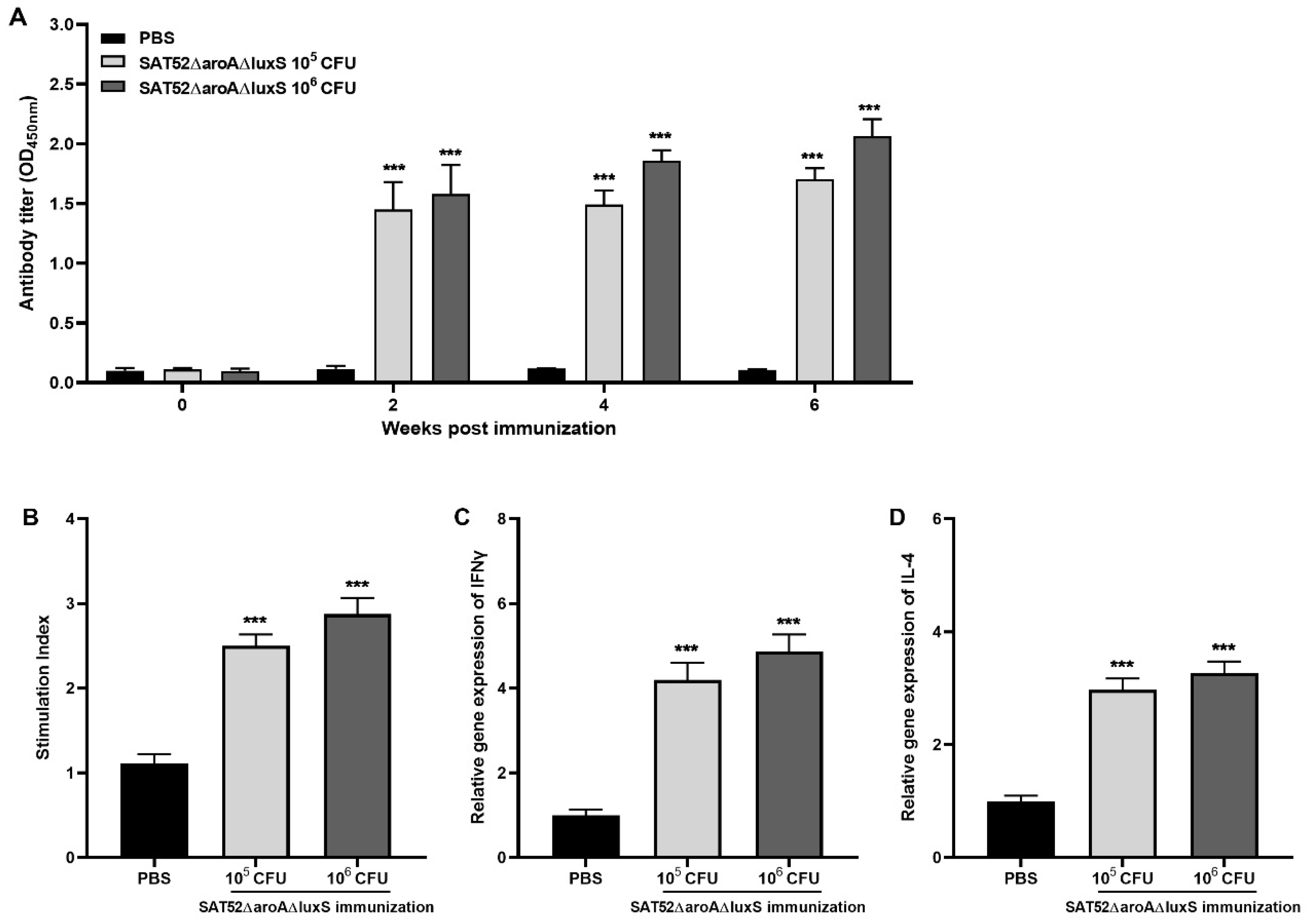
2.8. Humoral Immune Responses
2.9. Cell-Mediated Immune Responses by Splenocyte Proliferation
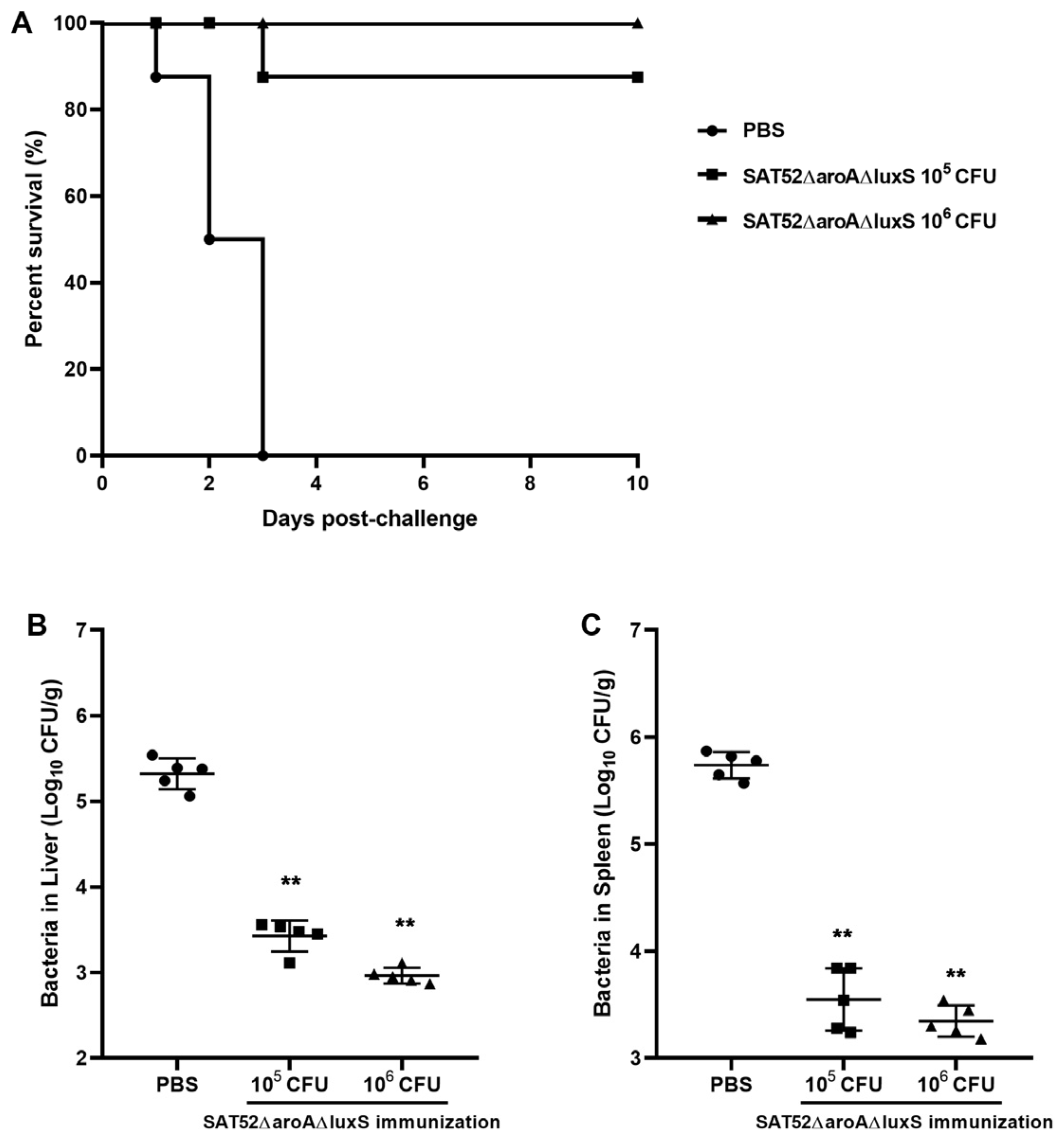
2.10. Immune Protection Assay
2.11. Statistical Analysis
3. Results
3.1. SAT52ΔaroAΔluxS Exhibited Reduced Bacterial Intracellular Survival in Cells
3.2. Double Mutation of aroA and luxS Genes Attenuated SAT52 Virulence In Vivo
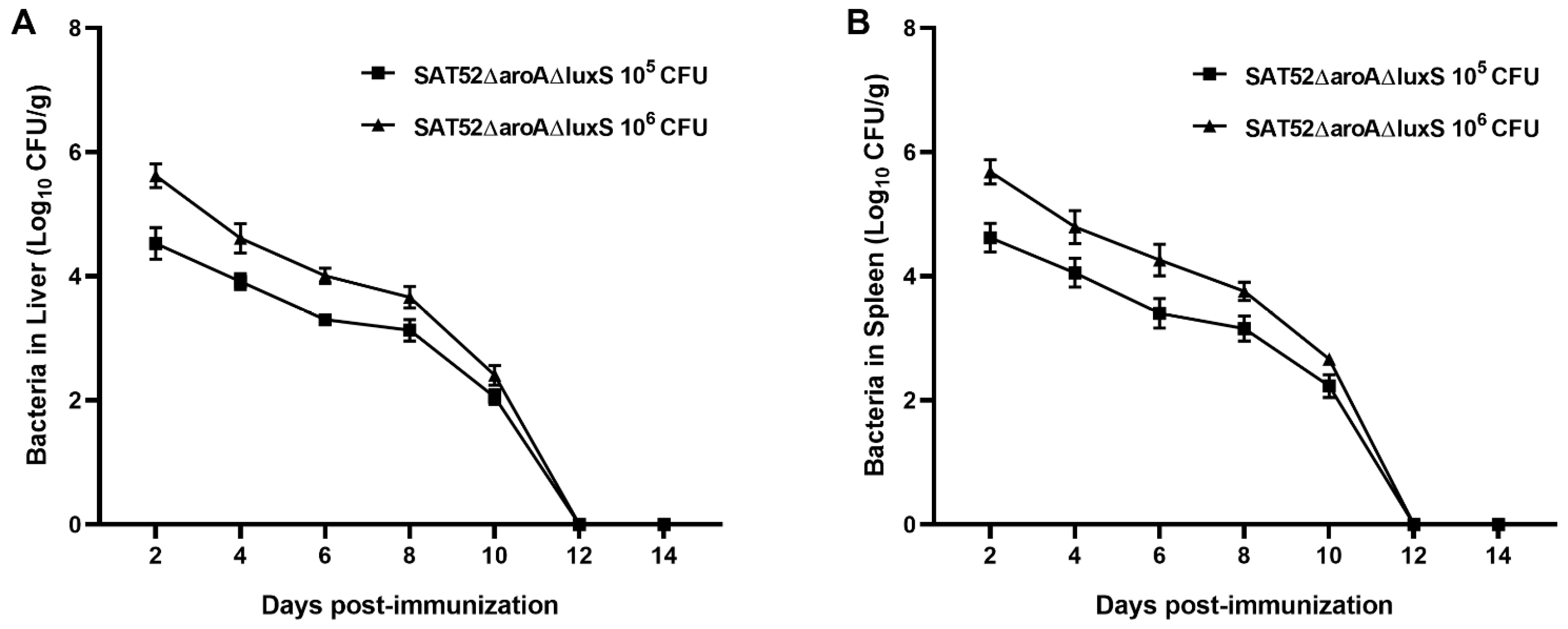
3.3. SAT52ΔaroAΔluxS Could Be Cleaned by Hosts within Two Weeks
3.4. SAT52ΔaroAΔluxS Effectively Induced Immune Responses
3.5. Vaccine Protection Efficacy
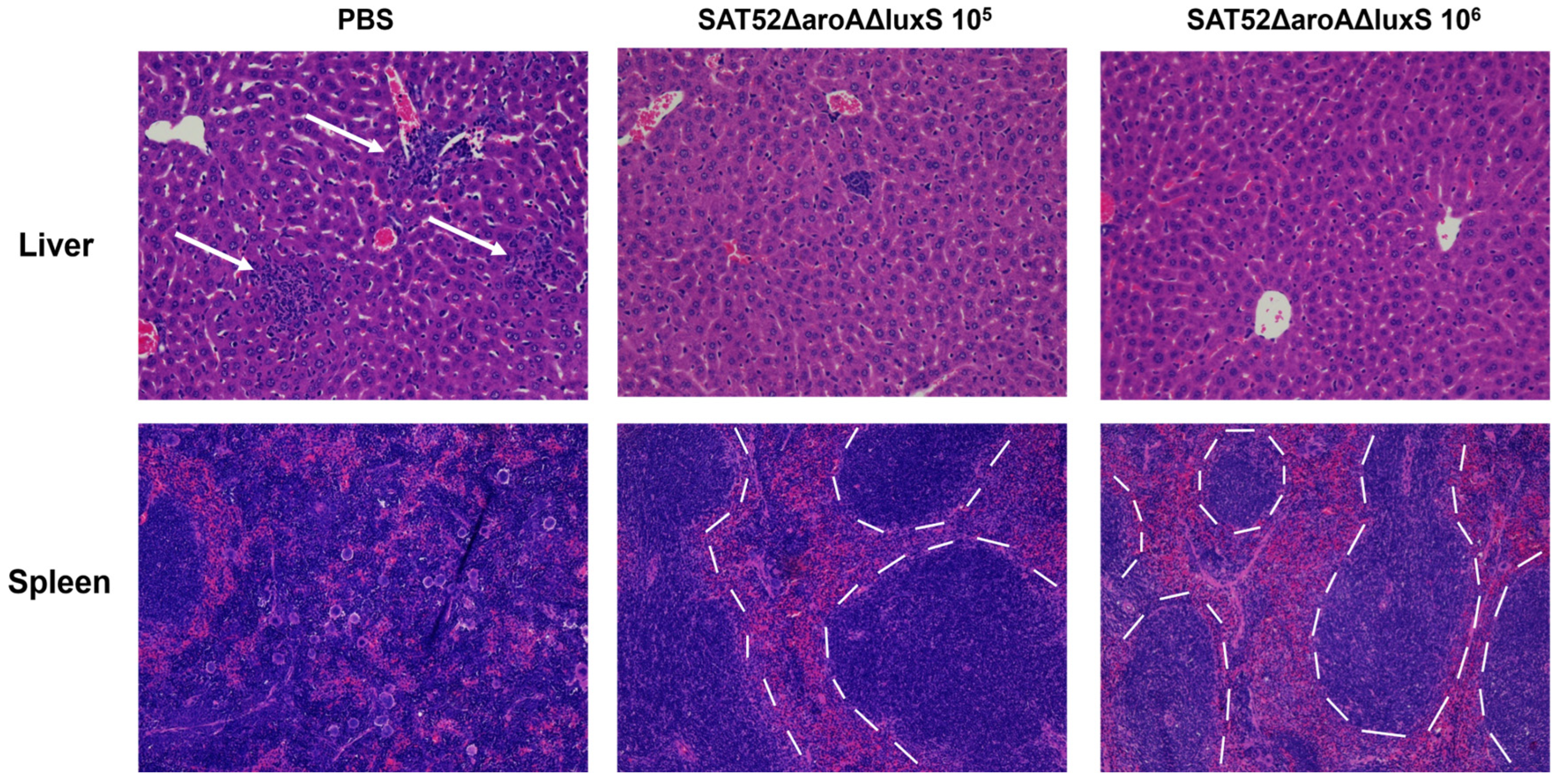
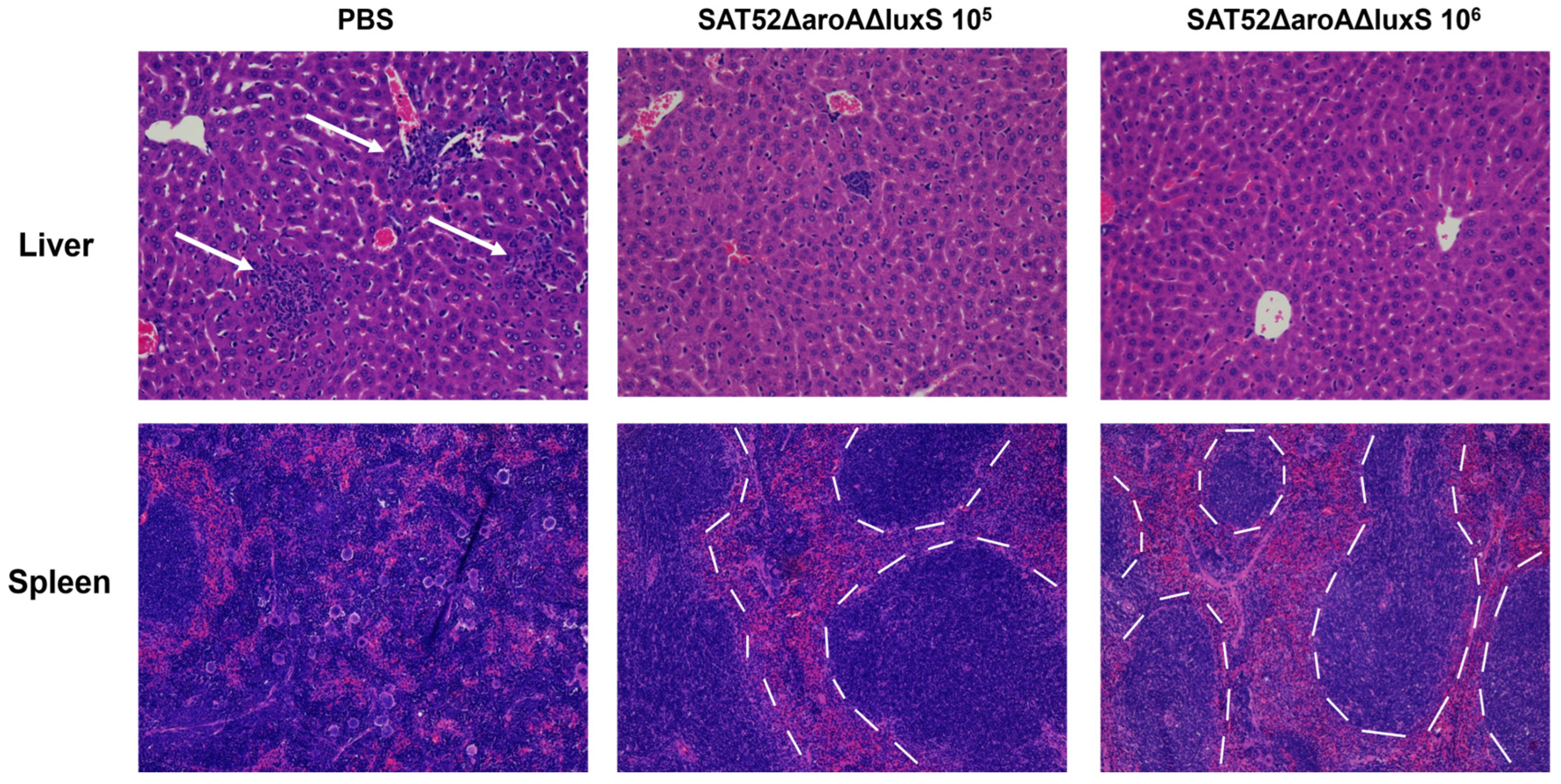
3.6. Histopathological Examination of Challenged Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chousalkar, K.; Gast, R. Review of egg-related salmonellosis and reduction strategies in United States, Australia, United Kingdom and New Zealand. Crit. Rev. Microbiol. 2018, 44, 290–303. [Google Scholar] [CrossRef]
- De la Cruz, M.L.; Conrado, I. Vaccination as a control strategy against Salmonella infection in pigs: A systematic review and meta-analysis of the literature. Res. Vet. Sci. 2017, 114, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Salmonella investigations in ruminants. Vet. Rec. 2017, 181, 366–367. [Google Scholar] [CrossRef] [PubMed]
- Lund, S.; Tahir, M. Outbreak of monophasic Salmonella typhimurium Sequence Type 34 linked to chocolate products. Ann. Med. Surg. 2022, 82, 5. [Google Scholar] [CrossRef] [PubMed]
- Branchu, P.; Bawn, M. Genome Variation and Molecular Epidemiology of Salmonella enterica Serovar Typhimurium Pathovariants. Infect. Immun. 2018, 86, 17. [Google Scholar] [CrossRef] [PubMed]
- Ramatla, T.; Tawana, M. One Health Perspective of Salmonella serovars in South Africa Using Pooled Prevalence: Systematic Review and Meta-Analysis. Int. J. Microbiol. 2022, 2022, 8952669. [Google Scholar] [CrossRef] [PubMed]
- Patra, S.D.; Mohakud, N.K. Prevalence and multidrug resistance in Salmonella enterica Typhimurium: An overview in South East Asia. World J. Microbiol. Biotechnol. 2021, 37, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Z.; Ge, H.W. Research progress on antibiotic resistance of Salmonella. Food Qual. Saf. 2022, 6, 10. [Google Scholar] [CrossRef]
- Tennant, S.M.; MacLennan, C.A. Nontyphoidal salmonella disease: Current status of vaccine research and development. Vaccine 2016, 34, 2907–2910. [Google Scholar] [CrossRef]
- MacLennan, C.A. Antibodies and protection against invasive Salmonella disease. Front. Immunol. 2014, 5, 4. [Google Scholar] [CrossRef]
- Jit, M.; Cooper, B. The role of vaccines in combating antimicrobial resistance. Eur. J. Public Health 2020, 30, 1. [Google Scholar] [CrossRef]
- Ji, H.J.; Jang, A.Y. Development of Live Attenuated Salmonella typhimurium Vaccine Strain Using Radiation Mutation Enhancement Technology (R-MET). Front. Immunol. 2022, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.Y.; McWhorter, A.R. Challenges in Vaccinating Layer Hens against Salmonella Typhimurium. Vaccines 2020, 8, 696. [Google Scholar] [CrossRef] [PubMed]
- Tennant, S.M.; Levine, M.M. Live attenuated vaccines for invasive Salmonella infections. Vaccine 2015, 33, C36–C41. [Google Scholar] [CrossRef] [PubMed]
- Germanier, R.; Fuer, E. Isolation and characterization of Gal E mutant Ty 21a of Salmonella typhi: A candidate strain for a live, oral typhoid vaccine. J. Infect. Dis. 1975, 131, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.M.; Layton, S.L. Scarless and site-directed mutagenesis in Salmonella enteritidis chromosome. BMC Biotechnol. 2007, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.X.; Liu, J. Development of a Method for Simultaneous Generation of Multiple Genetic Modification in Salmonella enterica Serovar Typhimurium. Front. Genet. 2020, 11, 14. [Google Scholar] [CrossRef]
- Hoiseth, S.K.; Stocker, B.A. Aromatic-dependent Salmonella typhimurium are non-virulent and effective as live vaccines. Nature 1981, 291, 238–239. [Google Scholar] [CrossRef]
- Hindle, Z.; Chatfield, S.N. Characterization of Salmonella enterica derivatives harboring defined aroC and Salmonella pathogenicity island 2 type III secretion system (ssaV) mutations by immunization of healthy volunteers. Infect. Immun. 2002, 70, 3457–3467. [Google Scholar] [CrossRef]
- Groisman, E.A.; Duprey, A. How the PhoP/PhoQ System Controls Virulence and Mg2+ Homeostasis: Lessons in Signal Transduction, Pathogenesis, Physiology, and Evolution. Microbiol. Mol. Biol. Rev. 2021, 85, 61. [Google Scholar] [CrossRef]
- Tennant, S.M.; Wang, J.Y. Engineering and Preclinical Evaluation of Attenuated Nontyphoidal Salmonella Strains Serving as Live Oral Vaccines and as Reagent Strains. Infect. Immun. 2011, 79, 4175–4185. [Google Scholar] [CrossRef] [PubMed]
- Tacket, C.O.; Hone, D.M. Comparison of the safety and immunogenicity of delta aroC delta aroD and delta cya delta crp Salmonella typhi strains in adult volunteers. Infect. Immun. 1992, 60, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Mercado-Lubo, R.; Gauger, E.J. A Salmonella enterica serovar Typhimurium succinate dehydrogenase/fumarate reductase double mutant is avirulent and immunogenic in BALB/c mice. Infect. Immun. 2008, 76, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Bai, H. Deletion of luxS further attenuates the virulence of the avian pathogenic Escherichia coli aroA mutant. Microb. Pathog. 2015, 88, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Yang, D.H. The Ferric Uptake Regulator Represses Type VI Secretion System Function by Binding Directly to the clpV Promoter in Salmonella enterica Serovar Typhimurium. Infect. Immun. 2019, 87, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Dai, J. DotU expression is highly induced during in vivo infection and responsible for virulence and Hcp1 secretion in avian pathogenic Escherichia coli. Front. Microbiol. 2014, 5, 588. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Liu, X. Escherichia coli Type III Secretion System 2 ATPase EivC Is Involved in the Motility and Virulence of Avian Pathogenic Escherichia coli. Front. Microbiol. 2016, 7, 14. [Google Scholar] [CrossRef]
- Gilles, H.J. Calculation of the index of acute toxicity by the method of linear regression. Comparison with the method of “Karber and Behrens”. Eur. J. Toxicol. Environ. Hyg. J. Eur. Toxicol. 1974, 7, 77–84. [Google Scholar]
- Dogra, V.; Verma, S. Development of OMP based indirect ELISA to gauge the antibody titers in bovines against Pasteurella multocida. Iran. J. Vet. Res. 2015, 16, 350–356. [Google Scholar]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Hajam, I.A.; Kim, J. Oral immunization with a novel attenuated Salmonella gallinarum encoding infectious bronchitis virus spike protein induces protective immune responses against fowl typhoid and infectious bronchitis in chickens. Vet. Res. 2018, 49, 10. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.W.; Wang, X.L. Development and Evaluation of a Trivalent Riemerella anatipestifer-Inactivated Vaccine. Clin. Vaccine Immunol. 2013, 20, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Haselbeck, A.H.; Panzner, U. Current perspectives on invasive nontyphoidal Salmonella disease. Curr. Opin. Infect. Dis. 2017, 30, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Herve, C.; Laupeze, B. The how’s and what’s of vaccine reactogenicity. Npj Vaccines 2019, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Oo, W.T. Salmonella Typhi Vi polysaccharide conjugate vaccine protects infants and children against typhoid fever. Lancet 2021, 398, 643–644. [Google Scholar] [CrossRef] [PubMed]
- Pham, O.H.; McSorley, S.J. Protective host immune responses to Salmonella infection. Future Microbiol. 2015, 10, 101–110. [Google Scholar] [CrossRef]
- Ali, A.; Waris, A. Recent advancement, immune responses, and mechanism of action of various vaccines against intracellular bacterial infections. Life Sci. 2023, 314, 121332. [Google Scholar] [CrossRef]
- Higginson, E.E.; Simon, R. Animal Models for Salmonellosis: Applications in Vaccine Research. Clin. Vaccine Immunol. 2016, 23, 746–756. [Google Scholar] [CrossRef]
- Thompson, A.; Fulde, M. The metabolic pathways utilized by Salmonella typhimurium during infection of host cells. Environ. Microbiol. Rep. 2018, 10, 140–154. [Google Scholar] [CrossRef]
- Felgner, S.; Frahm, M. aroA-Deficient Salmonella enterica Serovar Typhimurium Is More Than a Metabolically Attenuated Mutant. MBio 2016, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Salehi, T.Z.; Tabatabaei, S. Assessment of immunity against avian colibacillosis induced by an aroA mutant containing increased serum survival gene in broilers. Braz. J. Microbiol. 2012, 43, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.; Favaro, C. Effect of Spray Escherichia coli Vaccine on the Immunity of Poultry. Avian Dis. 2013, 57, 671–676. [Google Scholar]
- Sholpan, A.; Lamas, A. Salmonella spp. quorum sensing: An overview from environmental persistence to host cell invasion. AIMS Microbiol. 2021, 7, 238–256. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Shin, D. Implication of quorum sensing in Salmonella enterica serovar typhimurium virulence: The luxS gene is necessary for expression of genes in pathogenicity island 1. Infect. Immun. 2007, 75, 4885–4890. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.; Jun, S. Engineered Attenuated Salmonella typhimurium Expressing Neoantigen Has Anticancer Effects. ACS Synth. Biol. 2021, 10, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Karavolos, M.H.; Bulmer, D.M. LuxS affects flagellar phase variation independently of quorum sensing in Salmonella enterica serovar typhimurium. J. Bacteriol. 2008, 190, 769–771. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, D.; Cazevieille, C. A homologue of the Agrobacterium tumefaciens VirB and Bordetella pertussis Ptl type IV secretion systems is essential for intracellular survival of Brucella suis. Mol. Microbiol. 1999, 33, 1210–1220. [Google Scholar] [CrossRef]
- Valentine, P.J.; Devore, B.P. Identification of three highly attenuated Salmonella typhimurium mutants that are more immunogenic and protective in mice than a prototypical aroA mutant. Infect. Immun. 1998, 66, 3378–3383. [Google Scholar] [CrossRef]
- Sinha, K.; Mastroeni, P. Salmonella typhimurium aroA, htrA, and aroD htrA mutants cause progressive infections in athymic (nu/nu) BALB/c mice. Infect. Immun. 1997, 65, 1566–1569. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′ to 3′) a | Usage |
|---|---|---|
| aroAMu-F | CACGGCCAGTCTGTGGGGTTTTTATTTCTGTTTTTTGAGAGTTGAGTTTCTGTAGGCTGGAGCTGCTTC | Construction of aroA gene mutation |
| aroAMu-R | TGCGAATACGGACTCGGCGCGCCAGCCCGTCGACTGGCGCAACAGAAGACCATATGAATATCCTCCTTAG | Construction of aroA gene mutation |
| aroAout-F | GAGTTTATCCAGGTCGCTGA | Identification of aroA gene mutation |
| aroAout-R | GCGCATTATACGCGCCAA | Identification of aroA gene mutation |
| aroAin-F | CAATTACACCCTTTCTGCCGA | Identification of aroA gene mutation |
| aroAin-R | GGTCAGGAACTGGCTGGAA | Identification of aroA gene mutation |
| luxSMu-F | AACAAAGAGTTCAGTTTATTTTTAAAAAATTATCGGAGGTGACTAAATGGTGTAGGCTGGAGCTGCTTC | Construction of luxS gene mutation |
| luxSMu-R | CATAAACCGGGGTTAATTTAAATACTGGAACCGCTTACAAATAAGACTACATATGAATATCCTCCTTAG | Construction of luxS gene mutation |
| luxSout-F | AACGTGACGCTTCAGTACGT | Identification of luxS gene mutation |
| luxSout-R | TTAACAGGCCAGGCATTAC | Identification of luxS gene mutation |
| luxSin-F | AGATAGCTTCGCAGTCGATCAT | Identification of luxS gene mutation |
| luxSin-R | TCACTGAGCGAGTGCATCTG | Identification of luxS gene mutation |
| mGAPDHRT-F | GCACAGTCAAGGCCGAGAAT | Real-time PCR for mouse GAPDH |
| mGAPDHRT-R | GCCTTCTCCATGGTGGTGAA | Real-time PCR for mouse GAPDH |
| mIFNγ RT-F | TCAAGTGGCATAGATGTGGAAGAA | Real-time PCR for mouse IFNγ |
| mIFNγ RT-R | TGGCTCTGCAGGATTTTCATG | Real-time PCR for mouse IFNγ |
| mIL4 RT-F | TCGGCATTTTGAACGAGGTC | Real-time PCR for mouse IL-4 |
| mIL4 RT-R | GAAAAGCCCGAAAGAGTCTC | Real-time PCR for mouse IL-4 |
| Bacterial Strains | Dose of Challenge (CFUs) | No. of Deaths/No. of Total Mice | LD50 Value (CFUs) |
|---|---|---|---|
| SAT52 | 1 × 109 | 8/8 | 3.16 × 104 |
| 1 × 108 | 8/8 | ||
| 1 × 107 | 8/8 | ||
| 1 × 106 | 8/8 | ||
| 1 × 105 | 5/8 | ||
| 1 × 104 | 3/8 | ||
| SAT52ΔaroAΔluxS | 1 × 109 | 8/8 | 4.22 × 107 |
| 1 × 108 | 7/8 | ||
| 1 × 107 | 0/8 | ||
| 1 × 106 | 0/8 | ||
| 1 × 105 | 0/8 | ||
| 1 × 104 | 0/8 | ||
| PBS | / | 0/8 | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, W.; Yang, D.; Wu, X.; Zhang, B.; Wang, X.; Hu, J.; Qi, J.; Tian, M.; Bao, Y.; Wang, S. The aroA and luxS Double-Gene Mutant Strain Has Potential to Be a Live Attenuated Vaccine against Salmonella Typhimurium. Vaccines 2024, 12, 162. https://doi.org/10.3390/vaccines12020162
Zuo W, Yang D, Wu X, Zhang B, Wang X, Hu J, Qi J, Tian M, Bao Y, Wang S. The aroA and luxS Double-Gene Mutant Strain Has Potential to Be a Live Attenuated Vaccine against Salmonella Typhimurium. Vaccines. 2024; 12(2):162. https://doi.org/10.3390/vaccines12020162
Chicago/Turabian StyleZuo, Wei, Denghui Yang, Xiaojun Wu, Beibei Zhang, Xinyu Wang, Jiangang Hu, Jingjing Qi, Mingxing Tian, Yanqing Bao, and Shaohui Wang. 2024. "The aroA and luxS Double-Gene Mutant Strain Has Potential to Be a Live Attenuated Vaccine against Salmonella Typhimurium" Vaccines 12, no. 2: 162. https://doi.org/10.3390/vaccines12020162
APA StyleZuo, W., Yang, D., Wu, X., Zhang, B., Wang, X., Hu, J., Qi, J., Tian, M., Bao, Y., & Wang, S. (2024). The aroA and luxS Double-Gene Mutant Strain Has Potential to Be a Live Attenuated Vaccine against Salmonella Typhimurium. Vaccines, 12(2), 162. https://doi.org/10.3390/vaccines12020162