
High-Throughput Screening for the Prevalence of Neutralizing Antibodies against Human Adenovirus Serotype 5
 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
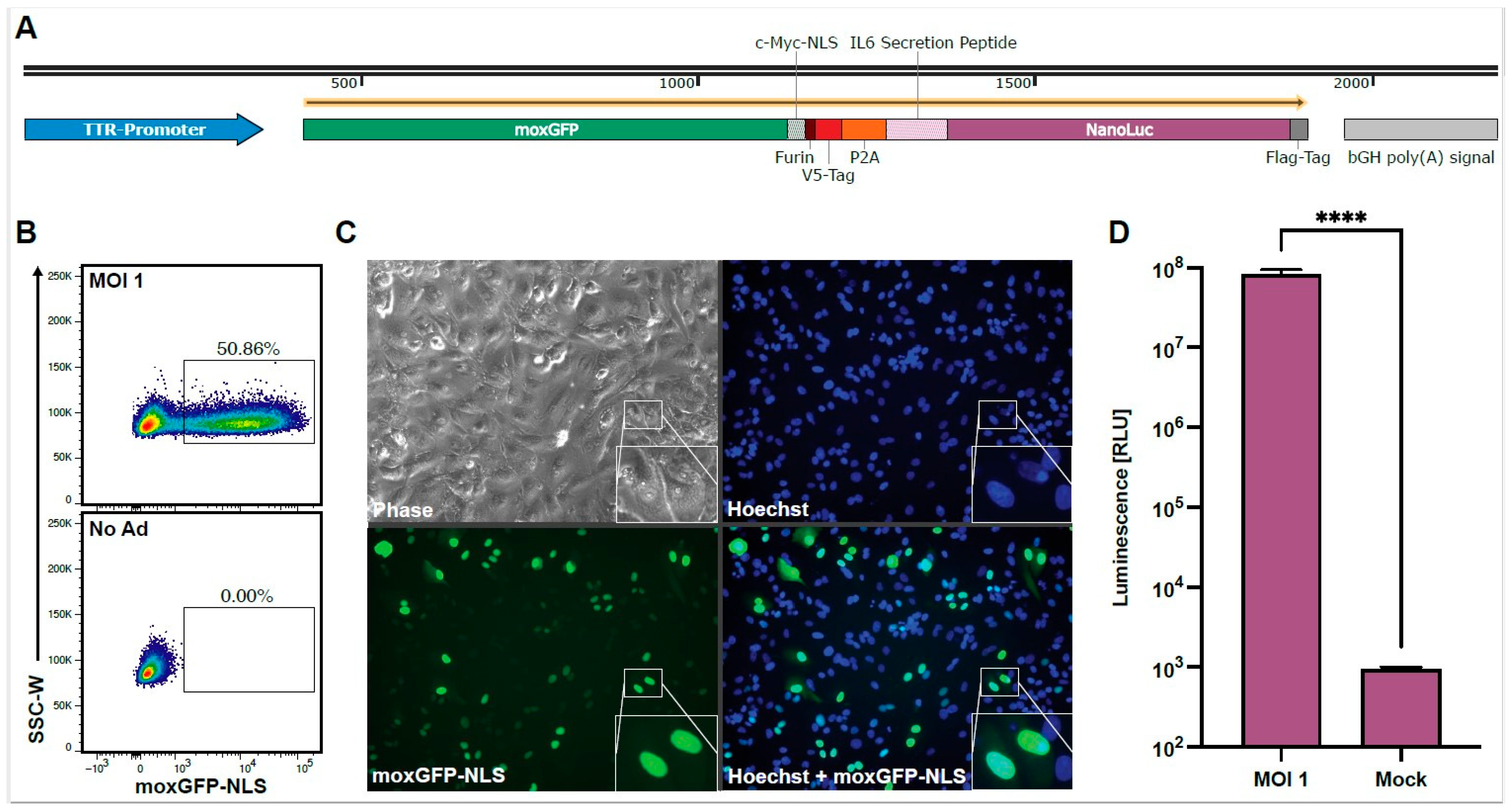
2.1. Generation of the HAdV-C5-Reporter Virus
2.2. Titration of the HAdV-C5-Reporter Virus Stock
2.3. Serum Samples
2.4. HAdV-C5-Neutralization Assay
2.5. Fluorescence Microscopy
2.6. Flow Cytometry
2.7. NanoLuc Luminescence Analysis
2.8. Cell-Titer Blue Cell Viability Assay
2.9. Anti-HAdV-C5 IgG ELISA
3. Results
3.1. Optimization of Assay Parameters
3.2. Validation of HAdV-C5-Neutralization Measurements
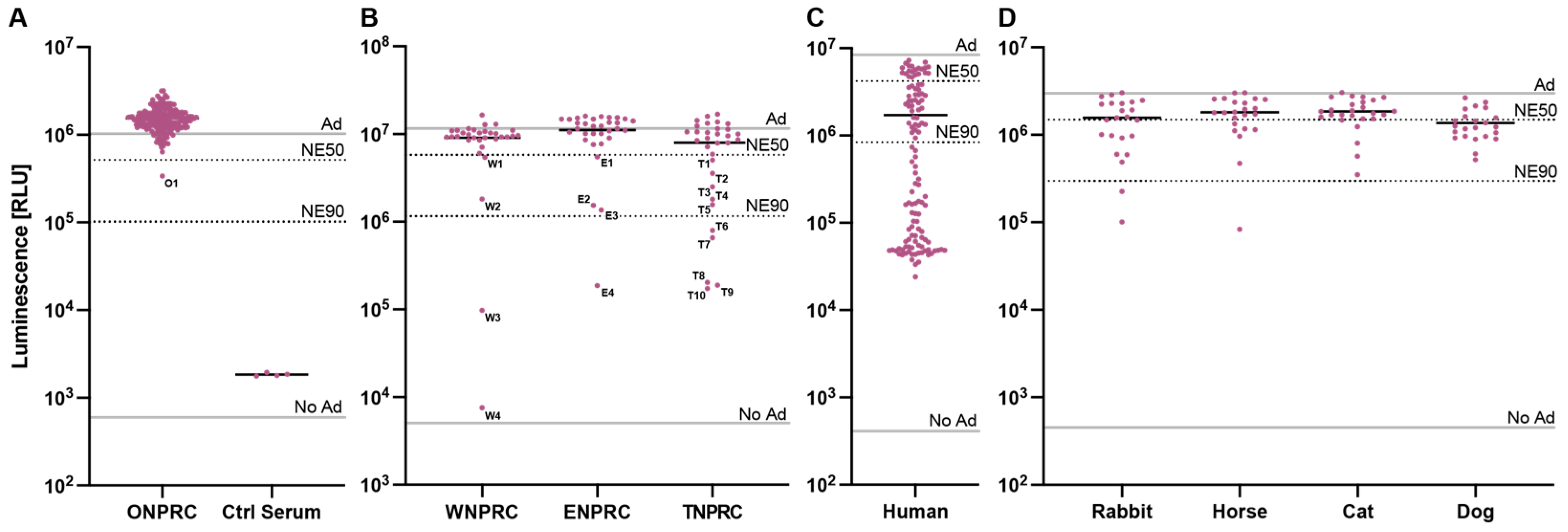
3.3. Neutralizing Antibodies against HAdV-C5 in Non-Human Primates
3.4. Neutralizing Antibodies against HAdV-C5 in Other Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, Z.; Anselmo, A.C.; Mitragotri, S. Viral vector-based gene therapies in the clinic. Bioeng. Transl. Med. 2022, 7, e10258. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Samulski, R.J. Engineering adeno-associated virus vectors for gene therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.G.; Jin, H.-T.; West, E.E.; Penaloza-MacMaster, P.; Wieland, A.; Zilliox, M.J.; McElrath, M.J.; Barouch, D.H.; Ahmed, R. Comparative analysis of simian immunodeficiency virus gag-specific effector and memory CD8+T cells induced by different adenovirus vectors. J. Virol. 2013, 87, 1359–1372. [Google Scholar] [CrossRef]
- Farhad, T.; Neves, K.; Arbuthnot, P.; Maepa, M.B. Adenoviral vectors: Potential as anti-HBV vaccines and therapeutics. Genes 2022, 13, 1941. [Google Scholar] [CrossRef] [PubMed]
- Chuang, I.; Sedegah, M.; Cicatelli, S.; Spring, M.; Polhemus, M.; Tamminga, C.; Patterson, N.; Guerrero, M.; Bennett, J.W.; McGrath, S.; et al. DNA prime/adenovirus boost malaria vaccine encoding P. falciparum CSP and AMA1 Induces sterile protection associated with cell-mediated immunity. PLoS ONE 2013, 8, e55571. [Google Scholar] [CrossRef] [PubMed]
- Jeyanathan, M.; Fritz, D.K.; Afkhami, S.; Aguirre, E.; Howie, K.J.; Zganiacz, A.; Dvorkin-Gheva, A.; Thompson, M.R.; Silver, R.F.; Cusack, R.P.; et al. Aerosol delivery, but not intramuscular injection, of adenovirus-vectored tuberculosis vaccine induces respiratory-mucosal immunity in humans. J. Clin. Investig. 2022, 7, 155655. [Google Scholar] [CrossRef] [PubMed]
- Smaill, F.; Jeyanathan, M.; Smieja, M.; Medina, M.F.; Thanthrige-Don, N.; Zganiacz, A.; Yin, C.; Heriazon, A.; Damjanovic, D.; Puri, L.; et al. A Human type 5 adenovirus–based tuberculosis vaccine induces robust T cell responses in humans despite preexisting anti-adenovirus immunity. Sci. Transl. Med. 2013, 5, 205ra134. [Google Scholar] [CrossRef]
- Kerstetter, L.J.; Buckley, S.; Bliss, C.M.; Coughlan, L. Adenoviral Vectors as Vaccines for Emerging Avian Influenza Viruses. Front. Immunol. 2020, 11, 607333. [Google Scholar] [CrossRef]
- Davison, A.J.; Benkő, M.; Harrach, B. Genetic content and evolution of adenoviruses. J. Gen. Virol. 2003, 84, 2895–2908. [Google Scholar] [CrossRef]
- Graham, F.L.; Prevec, L. Methods for construction of adenovirus vectors. Mol. Biotechnol. 1995, 3, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Hitt, M.; Bett, J.A.; Addison, C.L.; Prevec, L.; Graham, F.L. Techniques for human adenovirus vector construction and characterization. In Methods in Molecular Genetics; Elsevier: Amsterdam, The Netherlands, 1995; pp. 13–30. [Google Scholar]
- Choi, V.W.; Asokan, A.; Haberman, R.A.; Samulski, R.J. Production of recombinant adeno-associated viral vectors for in vitro and in vivo use. Curr. Protoc. Mol. Biol. 2007, 78, 16.25.1–16.25.24. [Google Scholar] [CrossRef]
- Travieso, T.; Li, J.; Mahesh, S.; Mello, J.D.F.R.E.; Blasi, M. The use of viral vectors in vaccine development. NPJ Vaccines 2022, 7, 1–10. [Google Scholar] [CrossRef]
- Knowles, M.K.; Roberts, D.; Craig, S.; Sheen, M.; Nadin-Davis, S.A.; Wandeler, A.I. In vitro and in vivo genetic stability studies of a human adenovirus type 5 recombinant rabies glycoprotein vaccine (ONRAB). Vaccine 2009, 27, 2662–2668. [Google Scholar] [CrossRef]
- Götting, J.; Baier, C.; Panagiota, V.; Maecker-Kolhoff, B.; Dhingra, A.; Heim, A. High genetic stability of co-circulating human adenovirus type 31 lineages over 59 years. Virus Evol. 2022, 8, veac067. [Google Scholar] [CrossRef]
- Chavda, V.P.; Bezbaruah, R.; Valu, D.; Patel, B.; Kumar, A.; Prasad, S.; Kakoti, B.B.; Kaushik, A.; Jesawadawala, M. Adenoviral Vector-Based Vaccine Platform for COVID-19: Current Status. Vaccines 2023, 11, 432. [Google Scholar] [CrossRef]
- Gao, G.P.; Yang, Y.; Wilson, J.M. Biology of adenovirus vectors with E1 and E4 deletions for liver-directed gene therapy. J. Virol. 1996, 70, 8934–8943. [Google Scholar] [CrossRef]
- Ricobaraza, A.; Gonzalez-Aparicio, M.; Mora-Jimenez, L.; Lumbreras, S.; Hernandez-Alcoceba, R. High-Capacity Adenoviral Vectors: Expanding the Scope of Gene Therapy. Int. J. Mol. Sci. 2020, 21, 3643. [Google Scholar] [CrossRef]
- Alba, R.; Bosch, A.; Chillon, M. Gutless adenovirus: Last-generation adenovirus for gene therapy. Gene Ther. 2005, 12, S18–S27. [Google Scholar] [CrossRef] [PubMed]
- Morsy, M.A.; Gu, M.; Motzel, S.; Zhao, J.; Lin, J.; Su, Q.; Allen, H.; Franlin, L.; Parks, R.J.; Graham, F.L.; et al. An adenoviral vector deleted for all viral coding sequences results in enhanced safety and extended expression of a leptin transgene. Proc. Natl. Acad. Sci. USA 1998, 95, 7866–7871. [Google Scholar] [CrossRef] [PubMed]
- Fiolet, T.; Kherabi, Y.; MacDonald, C.-J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: A narrative review. Clin. Microbiol. Infect. 2022, 28, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.-C.; Li, Y.-H.; Guan, X.-H.; Hou, L.-H.; Wang, W.-J.; Li, J.-X.; Wu, S.-P.; Wang, B.-S.; Wang, Z.; Wang, L.; et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: A dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 2020, 395, 1845–1854. [Google Scholar] [CrossRef]
- Hou, L.; Guo, X.; Jin, P.; Wu, S.; Zhu, J.; Pan, H.; Wang, X.; Song, Z.; Wan, J.; Cui, L.; et al. Heterologous AD5-nCOV plus CoronaVac versus homologous CoronaVac vaccination: A randomized phase 4 trial. Nat. Med. 2022, 28, 401–409. [Google Scholar] [CrossRef]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; del Rio, C.; et al. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): A double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 2008, 372, 1881–1893. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.; Buchbinder, S.; Duerr, A. Overview of STEP and Phambili trial results: Two phase IIb test-of-concept studies investigating the efficacy of MRK adenovirus type 5 gag/pol/nef subtype B HIV vaccine. Curr. Opin. HIV AIDS 2010, 5, 357–361. [Google Scholar] [CrossRef]
- Sprangers, M.C.; Lakhai, W.; Koudstaal, W.; Verhoeven, M.; Koel, B.F.; Vogels, R.; Goudsmit, J.; Havenga, M.J.E.; Kostense, S. Quantifying adenovirus-neutralizing antibodies by luciferase transgene detection: Addressing preexisting immunity to vaccine and gene therapy vectors. J. Clin. Microbiol. 2003, 41, 5046–5052. [Google Scholar] [CrossRef]
- Liu, Q.; Nie, J.; Huang, W.; Meng, S.; Yuan, B.; Gao, D.; Xu, X.; Wang, Y. Comparison of Two high-throughput assays for quantification of adenovirus type 5 neutralizing antibodies in a population of donors in China. PLoS ONE 2012, 7, e37532. [Google Scholar] [CrossRef] [PubMed]
- Aste-Amézaga, M.; Bett, A.J.; Wang, F.; Casimiro, D.R.; Antonello, J.M.; Patel, D.K.; Dell, E.C.; Franlin, L.L.; Dougherty, N.M.; Bennett, P.S.; et al. Quantitative adenovirus neutralization assays based on the secreted alkaline phosphatase reporter gene: Application in epidemiologic studies and in the design of adenovector vaccines. Hum. Gene Ther. 2004, 15, 293–304. [Google Scholar] [CrossRef]
- Bruder, J.T.; Semenova, E.; Chen, P.; Limbach, K.; Patterson, N.B.; Stefaniak, M.E.; Konovalova, S.; Thomas, C.; Hamilton, M.; King, C.R.; et al. Modification of Ad5 hexon hypervariable regions circumvents pre-existing Ad5 neutralizing antibodies and induces protective immune responses. PLoS ONE 2012, 7, e33920. [Google Scholar] [CrossRef]
- Untergasser, Production of Adenoviral Vectors for Application in Test Animals. 2021. Available online: https://www.untergasser.de/lab/protocols/adeno_vectors_production_animal_v1_0.htm (accessed on 20 December 2023).
- Luo, J.; Deng, Z.-L.; Luo, X.; Tang, N.; Song, W.-X.; Chen, J.; Sharff, K.A.; Luu, H.H.; Haydon, R.C.; Kinzler, K.W.; et al. A protocol for rapid generation of recombinant adenoviruses using the AdEasy system. Nat. Protoc. 2007, 2, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.A.; Russo, R.C.; Thurston, R.V. Trimmed Spearman-Karber method for estimating median lethal concentrations in toxicity bioassays. Environ. Sci. Technol. 1977, 11, 714–719. [Google Scholar] [CrossRef]
- Wettengel, J.M.; Hansen-Palmus, L.; Yusova, S.; Rust, L.; Biswas, S.; Carson, J.; Ryu, J.; Bimber, B.N.; Hennebold, J.D.; Burwitz, B.J. A Multifunctional and Highly Adaptable Reporter System for CRISPR/Cas Editing. Int. J. Mol. Sci. 2023, 24, 8271. [Google Scholar] [CrossRef]
- Burwitz, B.J.; Wettengel, J.M.; Mück-Häusl, M.A.; Ringelhan, M.; Ko, C.; Festag, M.M.; Hammond, K.B.; Northrup, M.; Bimber, B.N.; Jacob, T.; et al. Hepatocytic expression of human sodium-taurocholate cotransporting polypeptide enables hepatitis B virus infection of macaques. Nat. Commun. 2017, 8, 2146. [Google Scholar] [CrossRef]
- Costantini, L.M.; Baloban, M.; Markwardt, M.L.; Rizzo, M.A.; Guo, F.; Verkhusha, V.V.; Snapp, E.L. A palette of fluorescent proteins optimized for diverse cellular environments. Nat. Commun. 2015, 6, 7670. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V.; Lee, W. Identification of the human c-myc protein nuclear translocation signal. Mol. Cell. Biol. 1988, 8, 4048–4054. [Google Scholar] [PubMed]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Chng, J.; Wang, T.; Nian, R.; Lau, A.; Hoi, K.M.; Ho, S.C.; Gagnon, P.; Bi, X.; Yang, Y. Cleavage efficient 2A peptides for high level monoclonal antibody expression in CHO cells. In mAbs; Taylor & Francis: London, UK, 2015. [Google Scholar] [CrossRef]
- Biswas, S.; Rust, L.N.; Wettengel, J.M.; Yusova, S.; Fischer, M.; Carson, J.N.; Johnson, J.; Wei, L.; Thode, T.; Kaadige, M.R.; et al. Long-term hepatitis B virus infection of rhesus macaques requires suppression of host immunity. Nat. Commun. 2022, 13, 2995. [Google Scholar] [CrossRef] [PubMed]
- Kumin, D.; Hofmann, C.; Rudolph, M.; Both, G.W.; Loser, P. Biology of ovine adenovirus infection of nonpermissive cells. J. Virol. 2002, 76, 10882–10893. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mast, T.C.; Kierstead, L.; Gupta, S.B.; Nikas, A.A.; Kallas, E.G.; Novitsky, V.; Mbewe, B.; Pitisuttithum, P.; Schechter, M.; Vardas, E.; et al. International epidemiology of human pre-existing adenovirus (Ad) type-5, type-6, type-26 and type-36 neutralizing antibodies: Correlates of high Ad5 titers and implications for potential HIV vaccine trials. Vaccine 2010, 28, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Nwanegbo, E.; Vardas, E.; Gao, W.; Whittle, H.; Sun, H.; Rowe, D.; Robbins, P.D.; Gambotto, A. Prevalence of neutralizing antibodies to adenoviral serotypes 5 and 35 in the adult populations of The Gambia, South Africa, and the United States. Clin. Vaccine Immunol. 2004, 11, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Zhou, Y.; Wu, H.; Wang, Z.; Zhan, Y.; Feng, X.; Geng, R.; Wu, Y.; Kong, W.; Yu, X. Seroprevalence of neutralizing antibodies to human adenovirus type 5 in healthy adults in China. J. Med. Virol. 2012, 84, 1408–1414. [Google Scholar] [CrossRef]
- Gentschev, I.; Patil, S.S.; Petrov, I.; Cappello, J.; Adelfinger, M.; Szalay, A.A. Oncolytic virotherapy of canine and feline cancer. Viruses 2014, 6, 2122–2137. [Google Scholar] [CrossRef] [PubMed]
- Frisbie, D.; Ghivizzani, S.; Robbins, P.; Evans, C.; McIlwraith, C. Treatment of experimental equine osteoarthritis by in vivo delivery of the equine interleukin-1 receptor antagonist gene. Gene Ther. 2002, 9, 12–20. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wettengel, J.M.; Naka, H.; Dissen, G.A.; Torgerson, J.; Pounder, M.; Mueller, S.F.; Mueller, E.; Hagen, P.; Brandt, M.; Protzer, U.; et al. High-Throughput Screening for the Prevalence of Neutralizing Antibodies against Human Adenovirus Serotype 5. Vaccines 2024, 12, 155. https://doi.org/10.3390/vaccines12020155
Wettengel JM, Naka H, Dissen GA, Torgerson J, Pounder M, Mueller SF, Mueller E, Hagen P, Brandt M, Protzer U, et al. High-Throughput Screening for the Prevalence of Neutralizing Antibodies against Human Adenovirus Serotype 5. Vaccines. 2024; 12(2):155. https://doi.org/10.3390/vaccines12020155
Chicago/Turabian StyleWettengel, Jochen M., Hiroaki Naka, Gregory A. Dissen, Jeffrey Torgerson, Michelle Pounder, Simon F. Mueller, Elisabeth Mueller, Philipp Hagen, Micah Brandt, Ulrike Protzer, and et al. 2024. "High-Throughput Screening for the Prevalence of Neutralizing Antibodies against Human Adenovirus Serotype 5" Vaccines 12, no. 2: 155. https://doi.org/10.3390/vaccines12020155
APA StyleWettengel, J. M., Naka, H., Dissen, G. A., Torgerson, J., Pounder, M., Mueller, S. F., Mueller, E., Hagen, P., Brandt, M., Protzer, U., & Burwitz, B. J. (2024). High-Throughput Screening for the Prevalence of Neutralizing Antibodies against Human Adenovirus Serotype 5. Vaccines, 12(2), 155. https://doi.org/10.3390/vaccines12020155