
The Association of HIV-1 Neutralization in Aviremic Children and Adults with Time to ART Initiation and CD4+/CD8+ Ratios

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
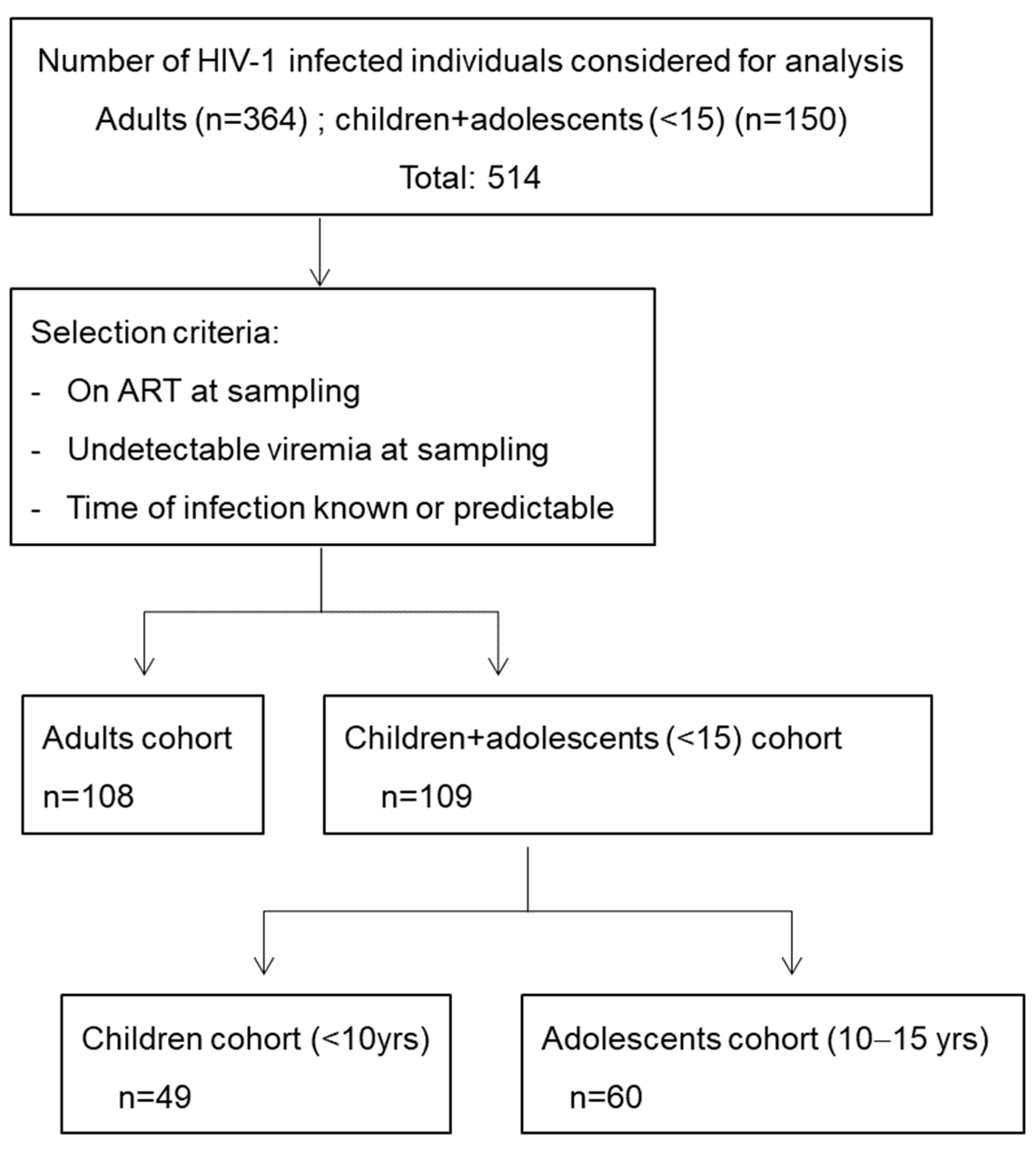
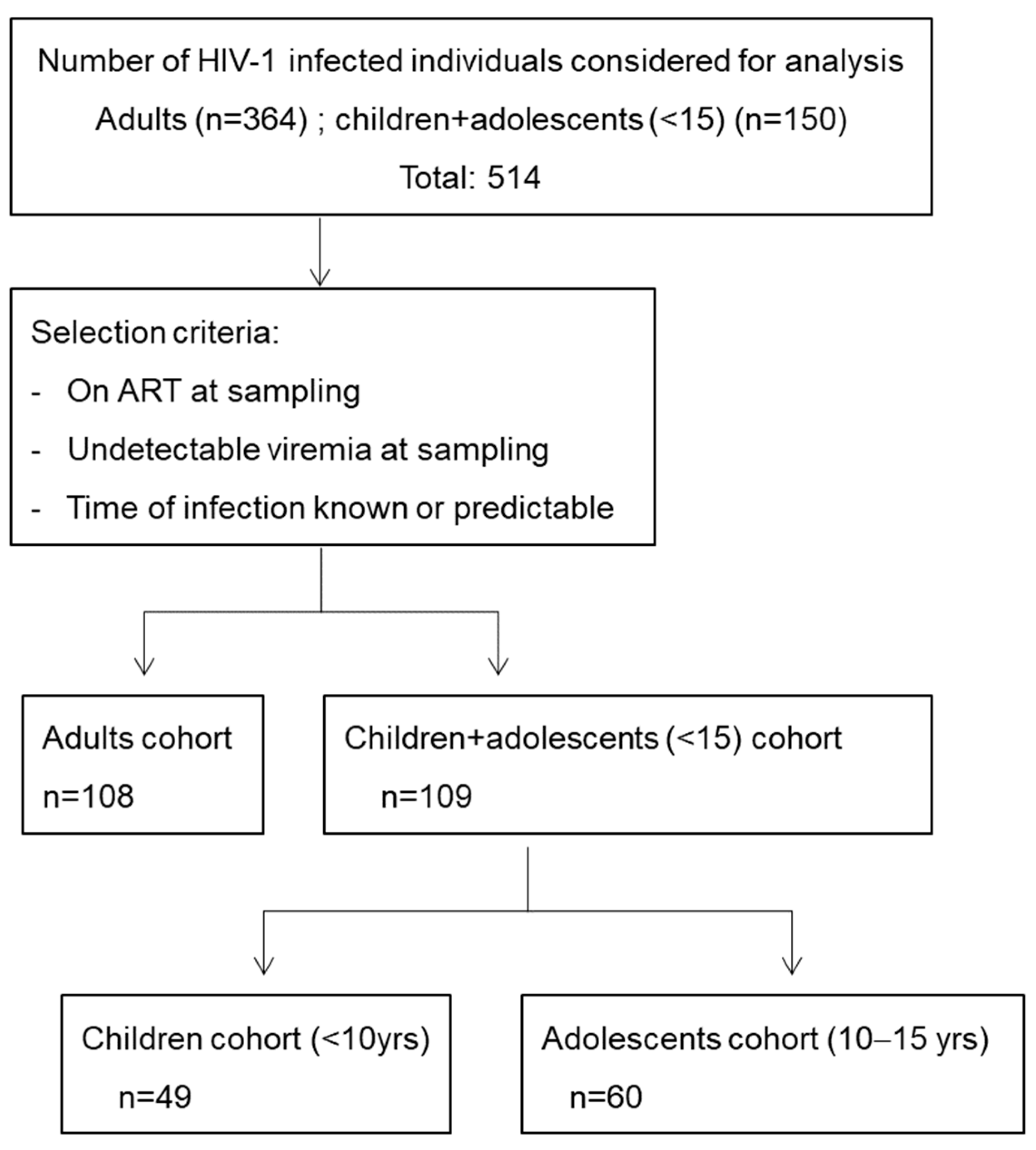
2.1. Study Participants
2.2. Neutralization Assays
2.3. CD4+ and CD8+ T Cell Counts
2.4. Statistical Analysis
3. Results
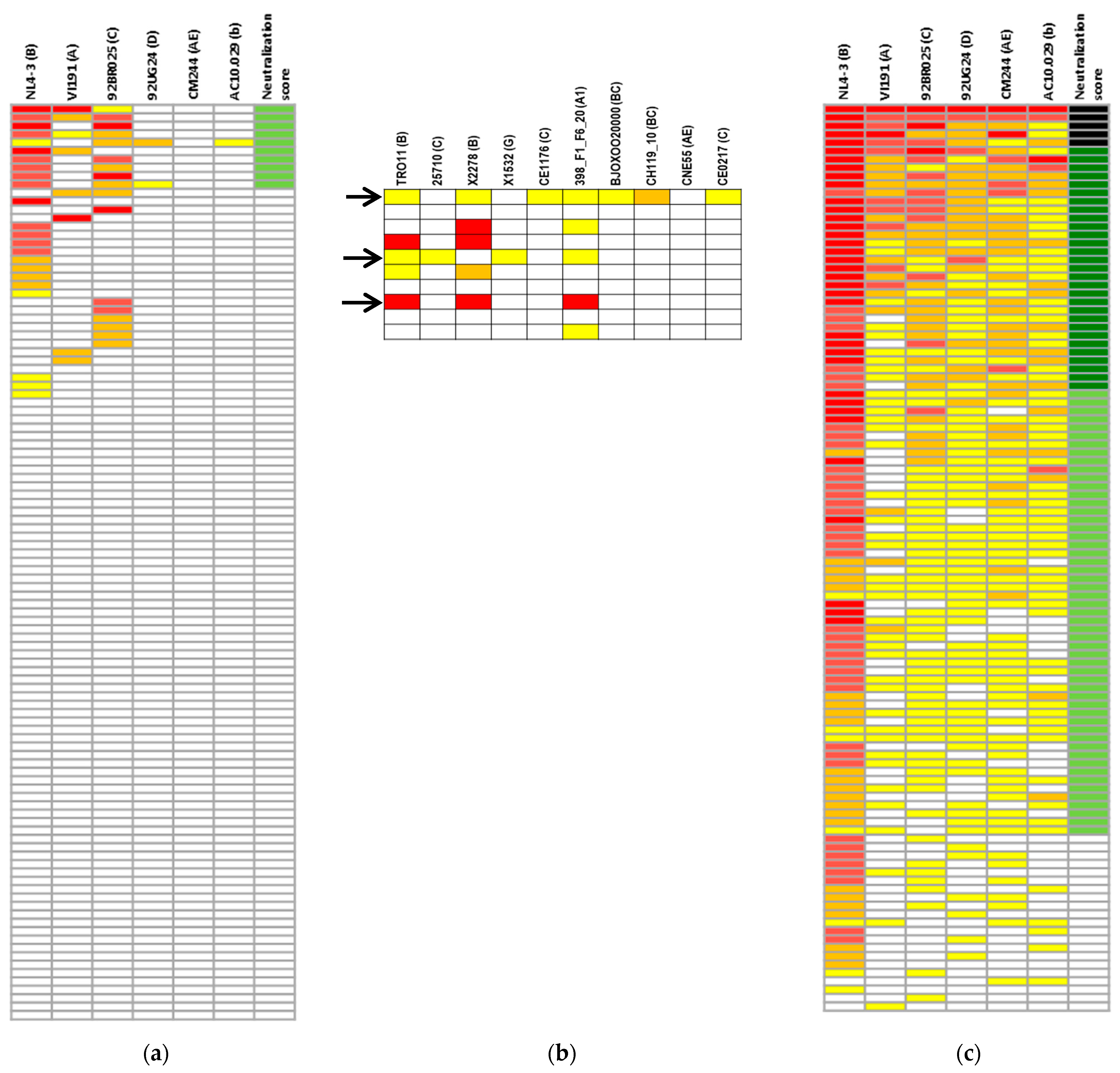
3.1. Screening for Cross-Neutralizing Sera in Children and Adolescents
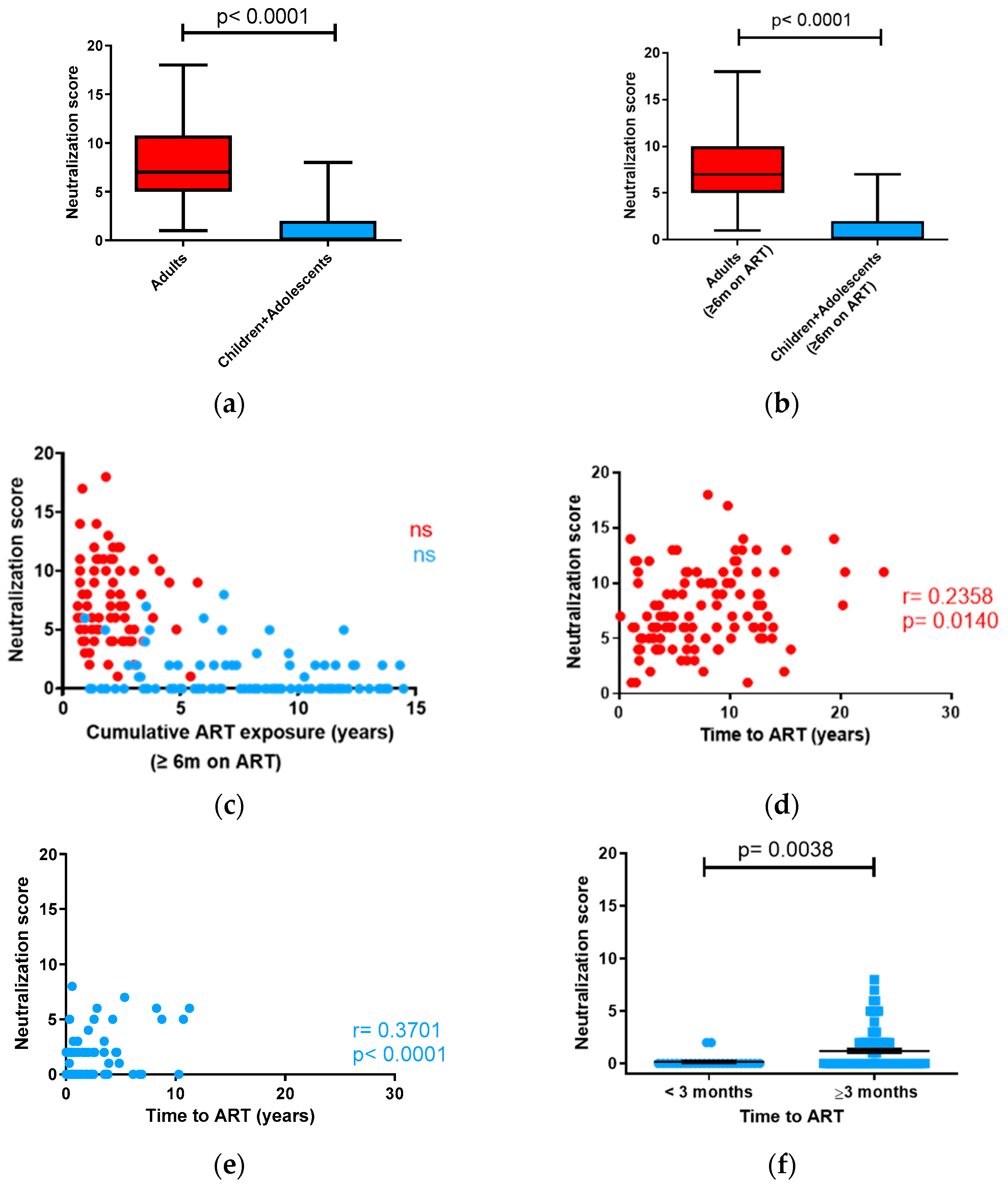
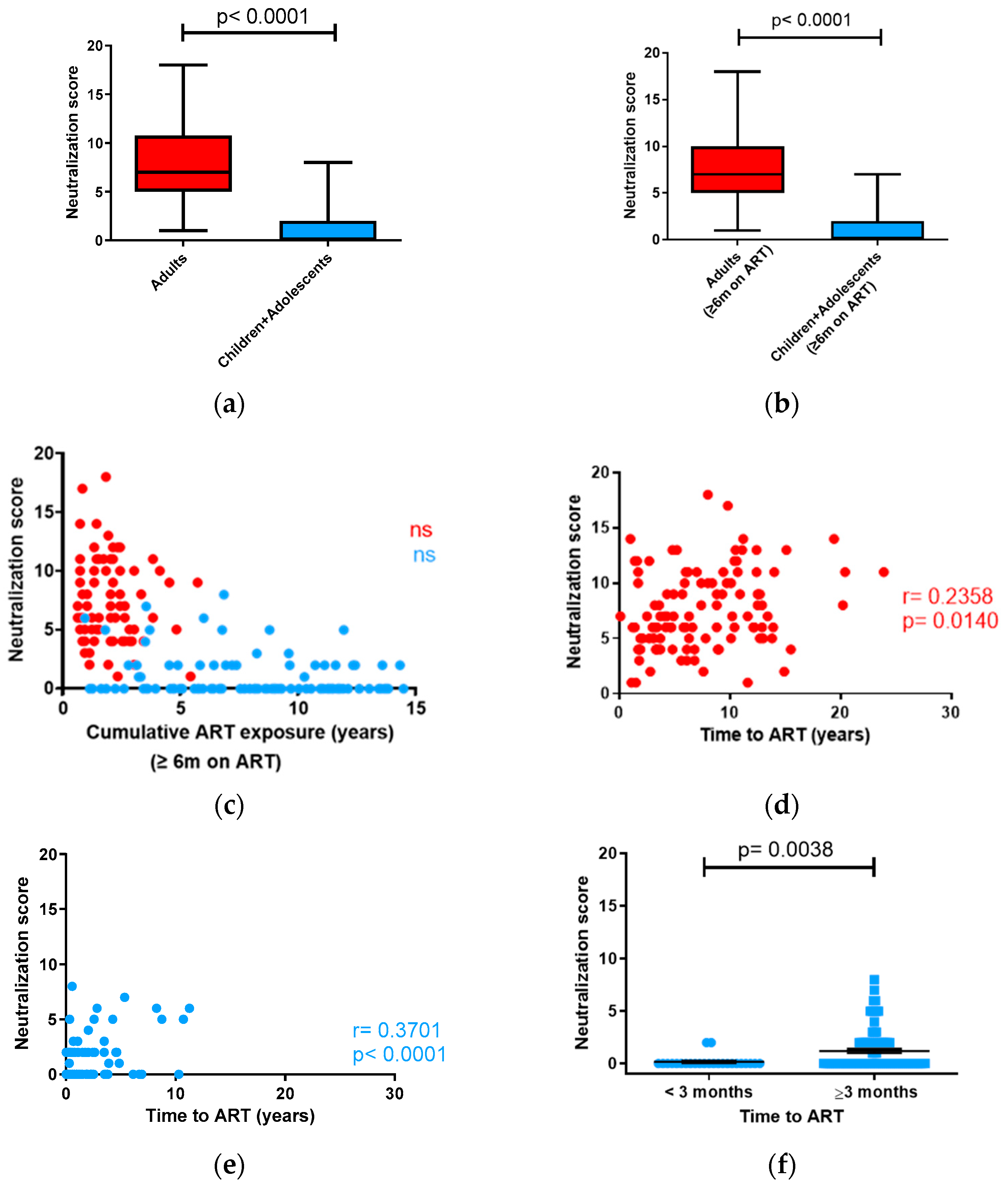
3.2. Neutralizing Antibody Response in Children vs. Adults
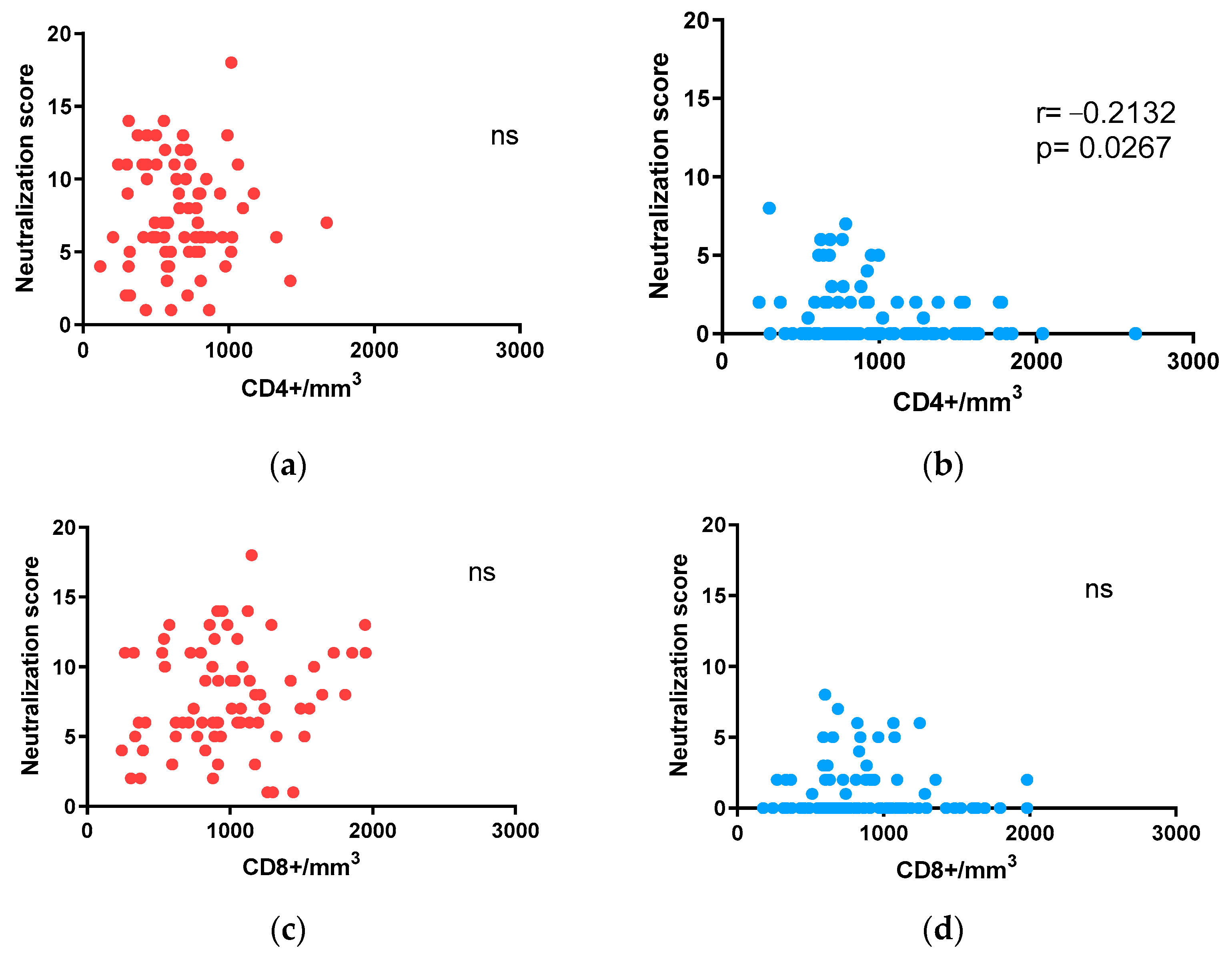
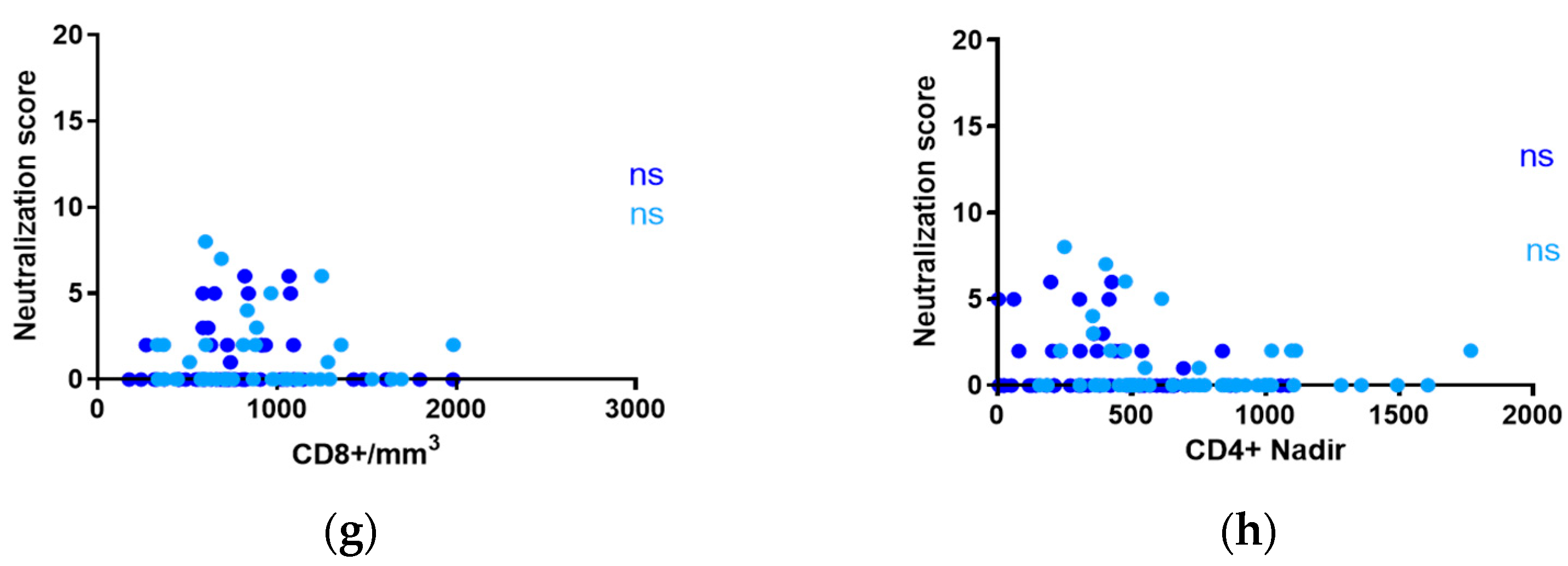
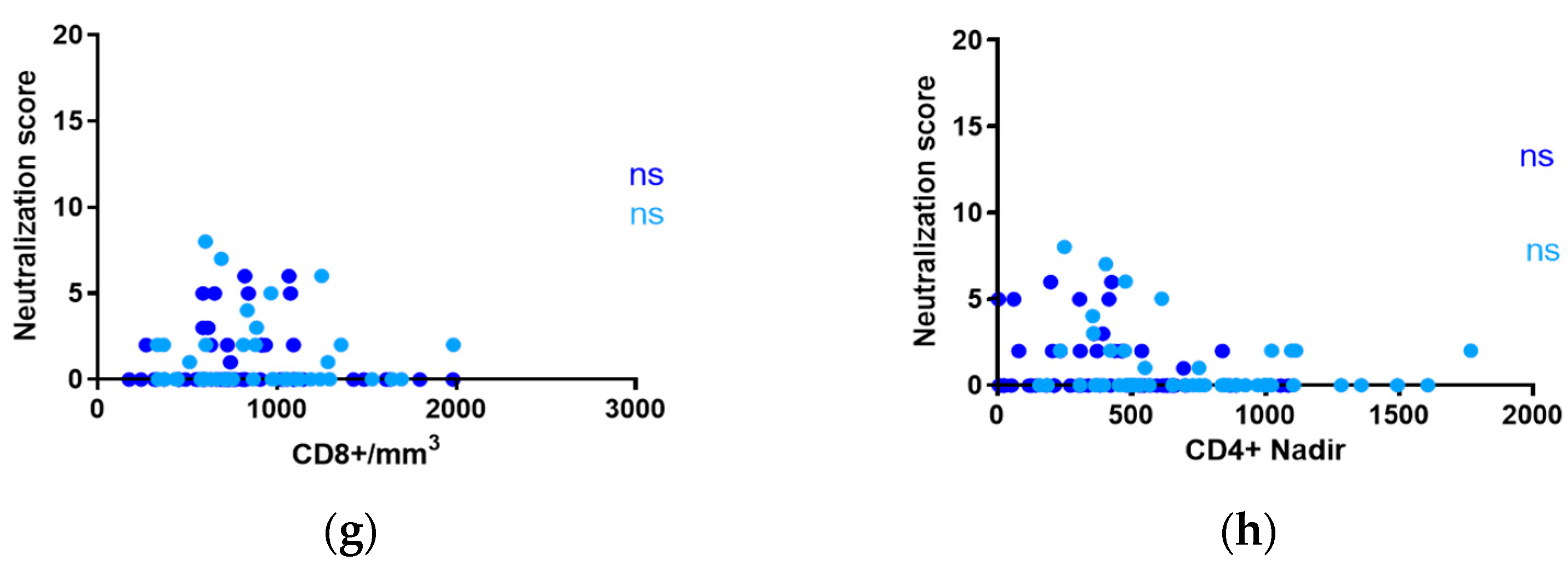
3.3. Influence of CD4+ and CD8+ T-Cell Counts on Neutralization in Adults and Children with ART-Suppressed Viremia
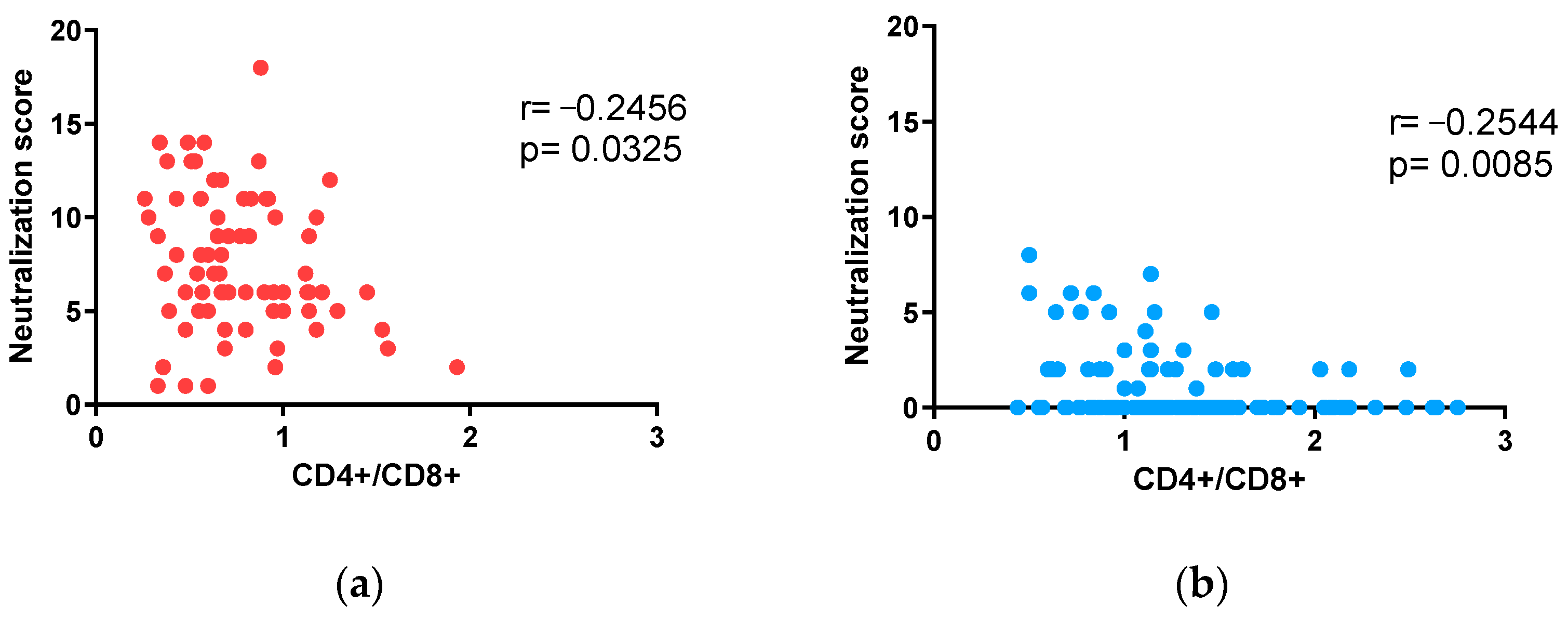
3.4. Impact of CD4+/CD8+ T-Lymphocyte Ratios on Neutralization in Adults and Children under 15 Years of Age with ART-Suppressed Viremia
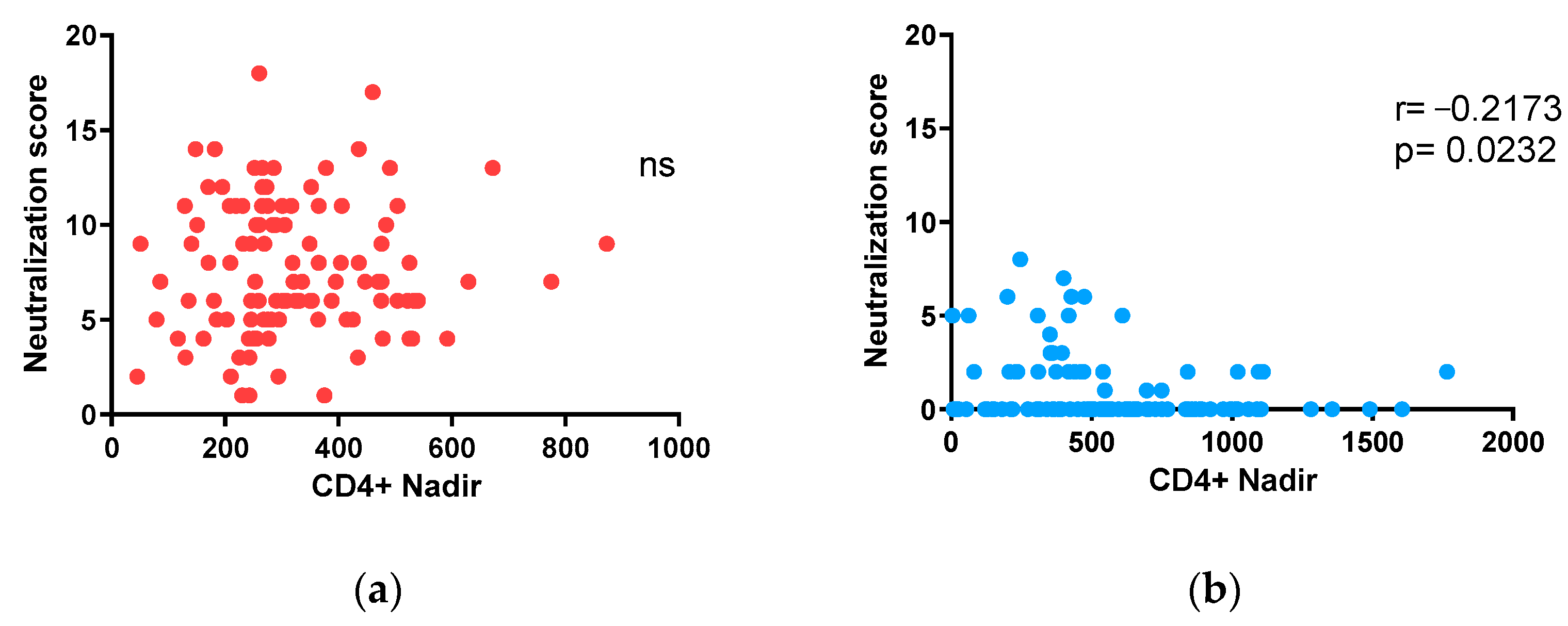
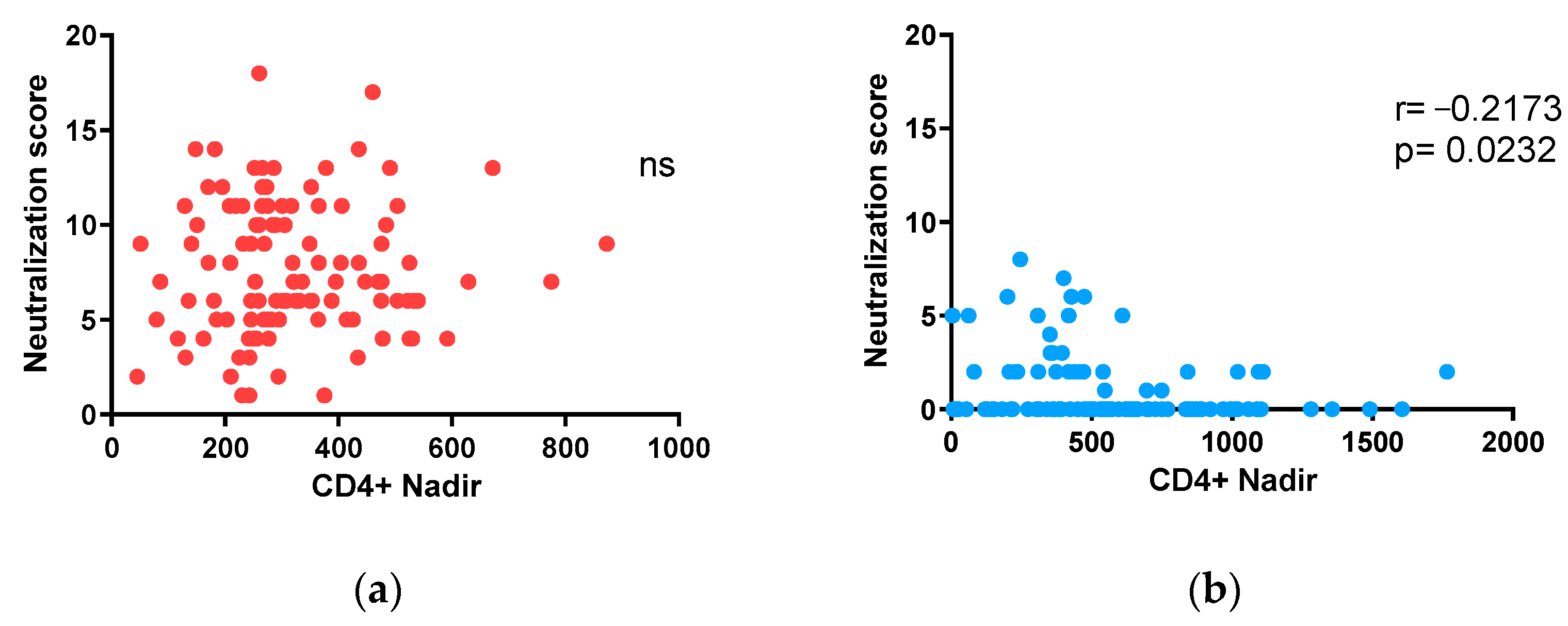
3.5. Impact of nadir CD4+ T-Lymphocyte Counts on Neutralization in Adults and Children under 15 on ART
3.6. Neutralizing Antibody Response in Children under 10 Years of Age and Adolescents (10 to 15 Years of Age) with ART-Suppressed Viremia
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collaboration, H.-C.; Ray, M.; Logan, R.; Sterne, J.A.; Hernandez-Diaz, S.; Robins, J.M.; Sabin, C.; Bansi, L.; van Sighem, A.; de Wolf, F.; et al. The effect of combined antiretroviral therapy on the overall mortality of HIV-infected individuals. AIDS 2010, 24, 123–137. [Google Scholar] [CrossRef]
- SeyedAlinaghi, S.; Afsahi, A.M.; Moradi, A.; Parmoon, Z.; Habibi, P.; Mirzapour, P.; Dashti, M.; Ghasemzadeh, A.; Karimi, E.; Sanaati, F.; et al. Current ART, determinants for virologic failure and implications for HIV drug resistance: An umbrella review. AIDS Res. Ther. 2023, 20, 74. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, S.A. Correlates of protection induced by vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Decker, J.M.; Wang, S.; Hui, H.; Kappes, J.C.; Wu, X.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Kilby, J.M.; Saag, M.S.; et al. Antibody neutralization and escape by HIV-1. Nature 2003, 422, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Bar, K.J.; Tsao, C.Y.; Iyer, S.S.; Decker, J.M.; Yang, Y.; Bonsignori, M.; Chen, X.; Hwang, K.K.; Montefiori, D.C.; Liao, H.X.; et al. Early low-titer neutralizing antibodies impede HIV-1 replication and select for virus escape. PLoS Pathog. 2012, 8, e1002721. [Google Scholar] [CrossRef]
- Haynes, B.F.; Burton, D.R.; Mascola, J.R. Multiple roles for HIV broadly neutralizing antibodies. Sci. Transl. Med. 2019, 11, eaaz2686. [Google Scholar] [CrossRef]
- Caskey, M.; Klein, F.; Nussenzweig, M.C. Broadly neutralizing anti-HIV-1 monoclonal antibodies in the clinic. Nat. Med. 2019, 25, 547–553. [Google Scholar] [CrossRef]
- Haynes, B.F.; Wiehe, K.; Borrow, P.; Saunders, K.O.; Korber, B.; Wagh, K.; McMichael, A.J.; Kelsoe, G.; Hahn, B.H.; Alt, F.; et al. Strategies for HIV-1 vaccines that induce broadly neutralizing antibodies. Nat. Rev. Immunol. 2023, 23, 142–158. [Google Scholar] [CrossRef]
- Locci, M.; Havenar-Daughton, C.; Landais, E.; Wu, J.; Kroenke, M.A.; Arlehamn, C.L.; Su, L.F.; Cubas, R.; Davis, M.M.; Sette, A.; et al. Human circulating PD-1+CXCR3-CXCR5+ memory Tfh cells are highly functional and correlate with broadly neutralizing HIV antibody responses. Immunity 2013, 39, 758–769. [Google Scholar] [CrossRef]
- Borrow, P.; Moody, M.A. Immunologic characteristics of HIV-infected individuals who make broadly neutralizing antibodies. Immunol. Rev. 2017, 275, 62–78. [Google Scholar] [CrossRef]
- Ferreira, C.B.; Merino-Mansilla, A.; Llano, A.; Perez, I.; Crespo, I.; Llinas, L.; Garcia, F.; Gatell, J.M.; Yuste, E.; Sanchez-Merino, V. Evolution of broadly cross-reactive HIV-1-neutralizing activity: Therapy-associated decline, positive association with detectable viremia, and partial restoration of B-cell subpopulations. J. Virol. 2013, 87, 12227–12236. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.S.; Madiga, M.C.; Hermanus, T.; Moore, P.L.; Wibmer, C.K.; Tumba, N.L.; Werner, L.; Mlisana, K.; Sibeko, S.; Williamson, C.; et al. The neutralization breadth of HIV-1 develops incrementally over four years and is associated with CD4+ T cell decline and high viral load during acute infection. J. Virol. 2011, 85, 4828–4840. [Google Scholar] [CrossRef] [PubMed]
- Hraber, P.; Seaman, M.S.; Bailer, R.T.; Mascola, J.R.; Montefiori, D.C.; Korber, B.T. Prevalence of broadly neutralizing antibody responses during chronic HIV-1 infection. AIDS 2014, 28, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Tomaras, G.D.; Binley, J.M.; Gray, E.S.; Crooks, E.T.; Osawa, K.; Moore, P.L.; Tumba, N.; Tong, T.; Shen, X.; Yates, N.L.; et al. Polyclonal B cell responses to conserved neutralization epitopes in a subset of HIV-1-infected individuals. J. Virol. 2011, 85, 11502–11519. [Google Scholar] [CrossRef]
- Muenchhoff, M.; Adland, E.; Karimanzira, O.; Crowther, C.; Pace, M.; Csala, A.; Leitman, E.; Moonsamy, A.; McGregor, C.; Hurst, J.; et al. Nonprogressing HIV-infected children share fundamental immunological features of nonpathogenic SIV infection. Sci. Transl. Med. 2016, 8, 358ra125. [Google Scholar] [CrossRef]
- Ditse, Z.; Muenchhoff, M.; Adland, E.; Jooste, P.; Goulder, P.; Moore, P.L.; Morris, L. HIV-1 Subtype C-Infected Children with Exceptional Neutralization Breadth Exhibit Polyclonal Responses Targeting Known Epitopes. J. Virol. 2018, 92, e00878-18. [Google Scholar] [CrossRef]
- Walker, L.M.; Simek, M.D.; Priddy, F.; Gach, J.S.; Wagner, D.; Zwick, M.B.; Phogat, S.K.; Poignard, P.; Burton, D.R. A limited number of antibody specificities mediate broad and potent serum neutralization in selected HIV-1 infected individuals. PLoS Pathog. 2010, 6, e1001028. [Google Scholar] [CrossRef]
- Mofenson, L.M.; Korelitz, J.; Meyer, W.A., 3rd; Bethel, J.; Rich, K.; Pahwa, S.; Moye, J., Jr.; Nugent, R.; Read, J. The relationship between serum human immunodeficiency virus type 1 (HIV-1) RNA level, CD4 lymphocyte percent, and long-term mortality risk in HIV-1-infected children. National Institute of Child Health and Human Development Intravenous Immunoglobulin Clinical Trial Study Group. J. Infect. Dis. 1997, 175, 1029–1038. [Google Scholar] [CrossRef]
- Schommers, P.; Kim, D.S.; Schlotz, M.; Kreer, C.; Eggeling, R.; Hake, A.; Stecher, M.; Park, J.; Radford, C.E.; Dingens, A.S.; et al. Dynamics and durability of HIV-1 neutralization are determined by viral replication. Nat. Med. 2023, 29, 2763–2774. [Google Scholar] [CrossRef]
- de Jose, M.I.; Jimenez de Ory, S.; Espiau, M.; Fortuny, C.; Navarro, M.L.; Soler-Palacin, P.; Munoz-Fernandez, M.A.; Working Groups of Co, R.; BioBank, H.H. A new tool for the paediatric HIV research: General data from the Cohort of the Spanish Paediatric HIV Network (CoRISpe). BMC Infect. Dis. 2013, 13, 2. [Google Scholar] [CrossRef]
- Medina-Ramirez, M.; Sanchez-Merino, V.; Sanchez-Palomino, S.; Merino-Mansilla, A.; Ferreira, C.B.; Perez, I.; Gonzalez, N.; Alvarez, A.; Alcocer-Gonzalez, J.M.; Garcia, F.; et al. Broadly cross-neutralizing antibodies in HIV-1 patients with undetectable viremia. J. Virol. 2011, 85, 5804–5813. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Merino, V.; Fabra-Garcia, A.; Gonzalez, N.; Nicolas, D.; Merino-Mansilla, A.; Manzardo, C.; Ambrosioni, J.; Schultz, A.; Meyerhans, A.; Mascola, J.R.; et al. Detection of Broadly Neutralizing Activity within the First Months of HIV-1 Infection. J. Virol. 2016, 90, 5231–5245. [Google Scholar] [CrossRef] [PubMed]
- deCamp, A.; Hraber, P.; Bailer, R.T.; Seaman, M.S.; Ochsenbauer, C.; Kappes, J.; Gottardo, R.; Edlefsen, P.; Self, S.; Tang, H.; et al. Global panel of HIV-1 Env reference strains for standardized assessments of vaccine-elicited neutralizing antibodies. J. Virol. 2014, 88, 2489–2507. [Google Scholar] [CrossRef] [PubMed]
- Rusert, P.; Kouyos, R.D.; Kadelka, C.; Ebner, H.; Schanz, M.; Huber, M.; Braun, D.L.; Hoze, N.; Scherrer, A.; Magnus, C.; et al. Determinants of HIV-1 broadly neutralizing antibody induction. Nat. Med. 2016, 22, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Marcelino, R.; Gramacho, F.; Martin, F.; Brogueira, P.; Janeiro, N.; Afonso, C.; Badura, R.; Valadas, E.; Mansinho, K.; Caldeira, L.; et al. Antibody response against selected epitopes in the HIV-1 envelope gp41 ectodomain contributes to reduce viral burden in HIV-1 infected patients. Sci. Rep. 2021, 11, 8993. [Google Scholar] [CrossRef]
- Sarzotti-Kelsoe, M.; Bailer, R.T.; Turk, E.; Lin, C.L.; Bilska, M.; Greene, K.M.; Gao, H.; Todd, C.A.; Ozaki, D.A.; Seaman, M.S.; et al. Optimization and validation of the TZM-bl assay for standardized assessments of neutralizing antibodies against HIV-1. J. Immunol. Methods 2014, 409, 131–146. [Google Scholar] [CrossRef] [PubMed]
- van Gils, M.J.; Euler, Z.; Schweighardt, B.; Wrin, T.; Schuitemaker, H. Prevalence of cross-reactive HIV-1-neutralizing activity in HIV-1-infected patients with rapid or slow disease progression. AIDS 2009, 23, 2405–2414. [Google Scholar] [CrossRef] [PubMed]
- Mellors, J.W.; Munoz, A.; Giorgi, J.V.; Margolick, J.B.; Tassoni, C.J.; Gupta, P.; Kingsley, L.A.; Todd, J.A.; Saah, A.J.; Detels, R.; et al. Plasma viral load and CD4+ lymphocytes as prognostic markers of HIV-1 infection. Ann. Intern. Med. 1997, 126, 946–954. [Google Scholar] [CrossRef]
- Prendergast, A.; Mphatswe, W.; Tudor-Williams, G.; Rakgotho, M.; Pillay, V.; Thobakgale, C.; McCarthy, N.; Morris, L.; Walker, B.D.; Goulder, P. Early virological suppression with three-class antiretroviral therapy in HIV-infected African infants. AIDS 2008, 22, 1333–1343. [Google Scholar] [CrossRef]
- McManus, M.; Henderson, J.; Gautam, A.; Brody, R.; Weiss, E.R.; Persaud, D.; Mick, E.; Luzuriaga, K.; Investigators, P. Quantitative Human Immunodeficiency Virus (HIV)-1 Antibodies Correlate With Plasma HIV-1 RNA and Cell-associated DNA Levels in Children on Antiretroviral Therapy. Clin. Infect. Dis. 2019, 68, 1725–1732. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Children | Adults | ||||
|---|---|---|---|---|---|
| Neutralization Category | Neutralization Score a | n | (%) | n | (%) |
| Elite | 14–18 | 0 | 0 | 5 | 4.6 |
| Broad | 10–13 | 0 | 0 | 29 | 27.0 |
| Cross | 5–9 | 10 | 9.2 | 53 | 49.0 |
| Weak or none | <5 | 99 | 90.8 | 21 | 19.4 |
| Characteristics | Children + Adolescents (<15 years) | Adults | ||
|---|---|---|---|---|
| Children (<10 years) | Adolescents (10–15 years) | Total (<15 years) | ||
| Total number of individuals | 49 | 60 | 109 | 108 |
| Number (%) of individuals by gender | ||||
| Male | 23 (46.9) | 24 (40) | 47 (43.1) | 94 (87.1) |
| Female | 26 (53.1) | 36 (60) | 62 (56.9) | 14 (12.9) |
| Number (%) of individuals by risk group | ||||
| Injecting drug user | 0 (0.0) | 0 (0.0) | 0 (0.0) | 4 (3.7) |
| Injecting drug user + homosexual | 0 (0.0) | 0 (0.0) | 0 (0.0) | 4 (3.7) |
| Bisexual | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (0.9) |
| Heterosexual contact | 0 (0.0) | 0 (0.0) | 0 (0.0) | 24 (22.2) |
| Hemophiliac | 0 (0.0) | 0 (0.0) | 0 (0.0) | 1 (0.9) |
| Homosexual | 0 (0.0) | 0 (0.0) | 0 (0.0) | 69 (63.8) |
| Vertical transmission | 49 (100) | 60(100) | 109 (100) | 0 (0.0) |
| Unknown | 0 (0) | 0 (0) | 0 (0) | 5 (4.6) |
| Breastfeeding | ||||
| Yes | 10 | 8 | 18 | - |
| No | 14 | 23 | 37 | - |
| Unknown | 25 | 29 | 54 | 108 |
| Median (range) age (years) a | 7.3 (1.5–9.8) | 12.6 (10–14.7) | 10.8 (1.5–14.7) | 42.0 (27.0–78.0) |
| Median (range) time post-infection (years) a | 7.3 (1.5–9.8) b | 12.6 (10–14.7) b | 10.8 (1.5–14.7) b | 8.8 (1.1–24.0) |
| Median (range) time to ART (years) a | 0.7 (0.0–6.7) b | 0.8 (0.0–11.3) b | 1.4 (0.0–12.6) b | 6.8 (0.1–23.9) |
| Median (range) time on ART (years) a | 5.0 (0.1–9.5) | 10.45 (0.9–14.7) | 6.9 (0.1–14.7) | 1.3 (0.0–5.7) |
| Median (range) nadir CD4+ cells/mm3 a | 696 (154–1765) | 424 (5–1088) | 488 (5–1765) | 289 (45–873) |
| Median (range) CD4+ cells/mm3 a | 1067 (300–2632) | 814 (236–1846) | 897 (236–2632) | 640 (117–1673) |
| Median (range) CD8+ cells/mm3 a | 780.5 (330–1982) | 749.5 (176–1983) | 754.5 (176–1983) | 916 (243–1950) |
| Median (range) CD4+/CD8+ ratio | 1.25 (0.50–2.50) | 1.10 (0.40–2.80) | 1.20 (0.40–2.80) | 0.68 (0.26–1.93) |
| Viral RNA copies/mL plasma | <50 | <50 | <50 | <50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchez-Merino, V.; Martin-Serrano, M.; Beltran, M.; Lazaro-Martin, B.; Cervantes, E.; Oltra, M.; Sainz, T.; Garcia, F.; Navarro, M.L.; Yuste, E. The Association of HIV-1 Neutralization in Aviremic Children and Adults with Time to ART Initiation and CD4+/CD8+ Ratios. Vaccines 2024, 12, 8. https://doi.org/10.3390/vaccines12010008
Sanchez-Merino V, Martin-Serrano M, Beltran M, Lazaro-Martin B, Cervantes E, Oltra M, Sainz T, Garcia F, Navarro ML, Yuste E. The Association of HIV-1 Neutralization in Aviremic Children and Adults with Time to ART Initiation and CD4+/CD8+ Ratios. Vaccines. 2024; 12(1):8. https://doi.org/10.3390/vaccines12010008
Chicago/Turabian StyleSanchez-Merino, Victor, Miguel Martin-Serrano, Manuela Beltran, Beatriz Lazaro-Martin, Eloisa Cervantes, Manuel Oltra, Talia Sainz, Felipe Garcia, Maria Luisa Navarro, and Eloisa Yuste. 2024. "The Association of HIV-1 Neutralization in Aviremic Children and Adults with Time to ART Initiation and CD4+/CD8+ Ratios" Vaccines 12, no. 1: 8. https://doi.org/10.3390/vaccines12010008
APA StyleSanchez-Merino, V., Martin-Serrano, M., Beltran, M., Lazaro-Martin, B., Cervantes, E., Oltra, M., Sainz, T., Garcia, F., Navarro, M. L., & Yuste, E. (2024). The Association of HIV-1 Neutralization in Aviremic Children and Adults with Time to ART Initiation and CD4+/CD8+ Ratios. Vaccines, 12(1), 8. https://doi.org/10.3390/vaccines12010008