
Development of Immersion and Oral Bivalent Nanovaccines for Streptococcosis and Columnaris Disease Prevention in Fry and Fingerling Asian Seabass (Lates calcarifer) Nursery Farms


 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Experimental Designs
2.2. Bacterial Culture and Formalin-Killed Vaccine Preparations
2.3. Formulation of Bivalent Nanovaccines
2.4. Immersion Vaccination
2.5. Experimental Feed Preparation and Oral Vaccination
2.6. Effects of Bivalent Nanovaccine on IgM and Immune Responses of Larvae to Fingerling Stages of Asian Seabass
2.6.1. Fish Sampling
2.6.2. Extraction of Whole-Body Protein
2.6.3. Total RNA Extraction and Preparation of First Strand cDNA
2.6.4. Histology Analysis of Mucosa-Associated Lymphoid Tissues (MALTs)
2.6.5. Total Serum IgM
2.6.6. Assessment of Serum IgM Specific to S. iniae and F. covae Antigens
2.7. Humoral Innate Immune Response Assays
2.7.1. Lysozyme Activity
2.7.2. Bactericidal Activity
2.8. Expression of Immune-Related Genes of Larvae in Fingerling Stages of Asian Seabass Using Quantitative Real-Time PCR (qRT-PCR)
2.9. Growth Performance
2.10. Challenge with S. iniae and F. covae
2.11. Statistical Analysis
3. Results
3.1. Production and Characterization of S. iniae and F. covae Nanovaccines
3.2. Effects of Bivalent Nanovaccine via Immersion and Oral Vaccination on IgM and Immune Responses of Larvae to Fingerling Stages of Asian Seabass
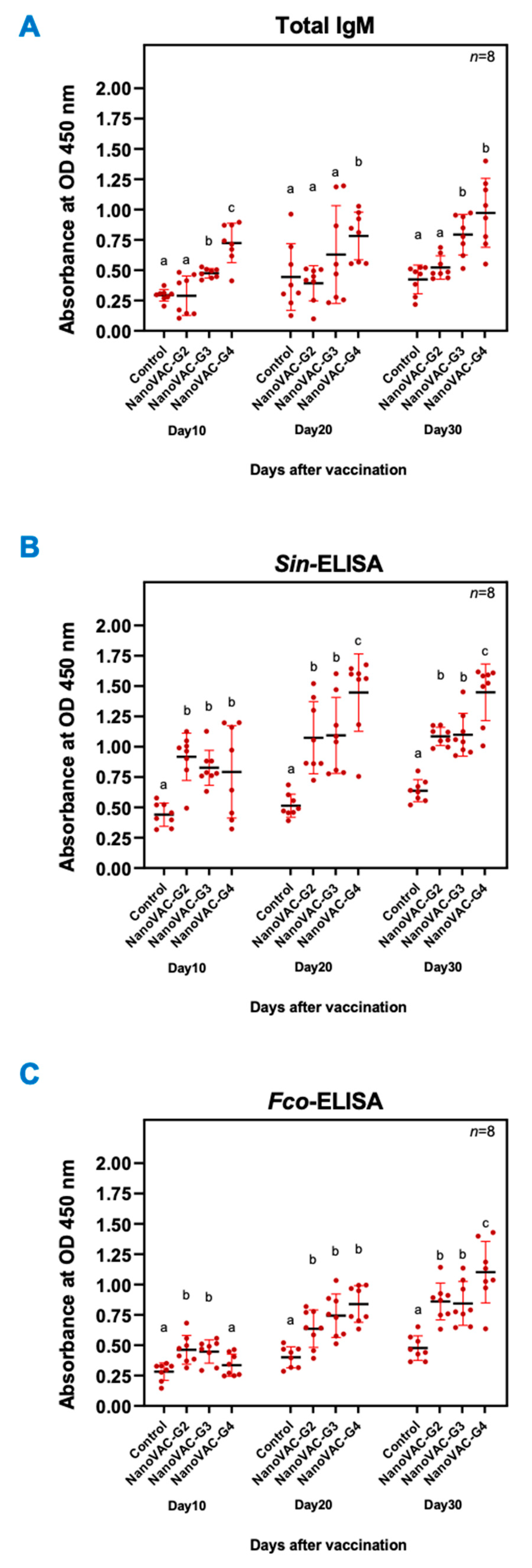
3.2.1. Total Serum IgM
3.2.2. Serum IgM specific for S. iniae and F. covae
3.2.3. Nonspecific Humoral Immune Responses
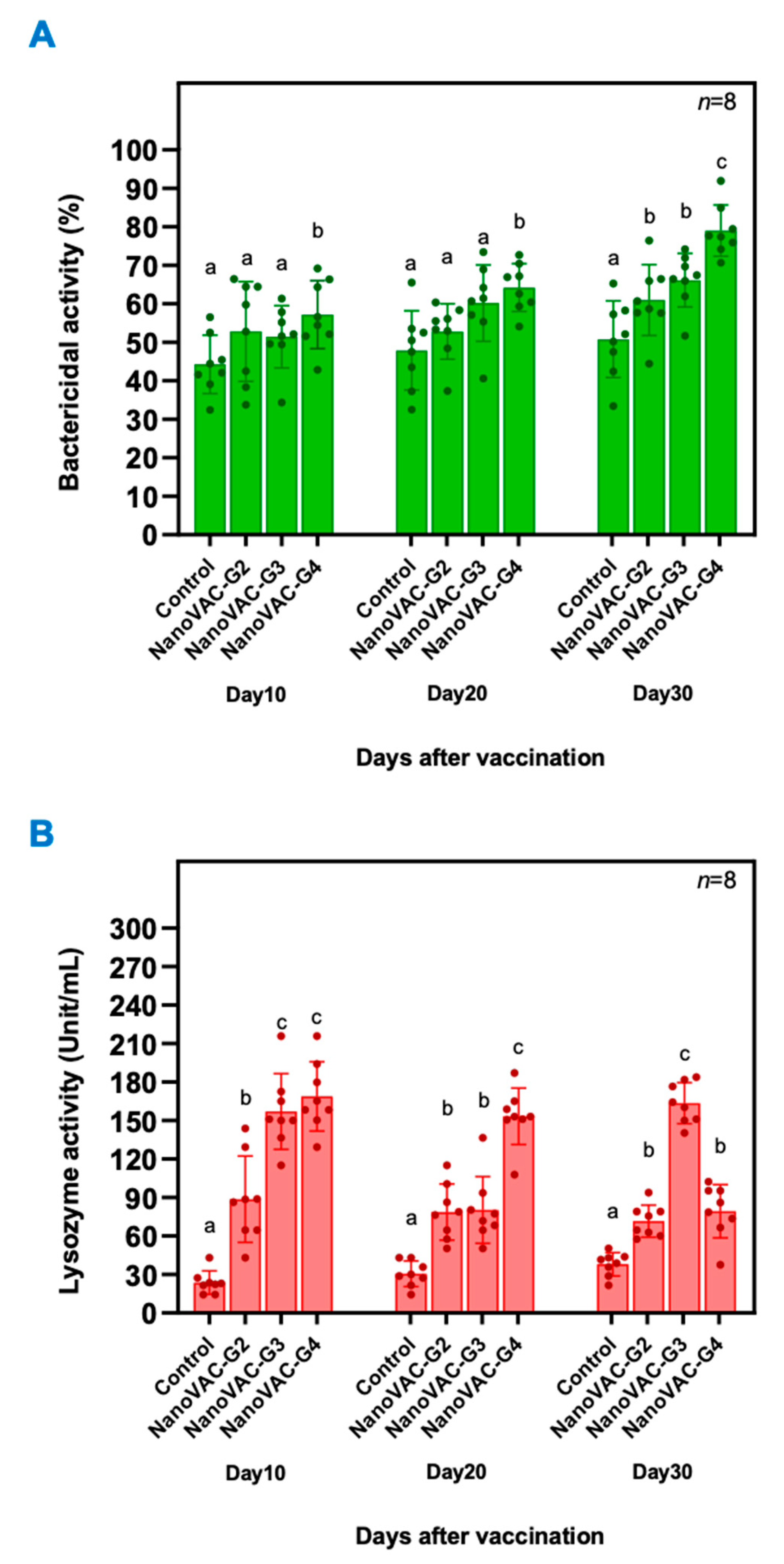
Bactericidal Activity (BA)
Lysozyme Activity
3.3. Effects of Bivalent Nanovaccines on the Expression of Immune-Related Genes
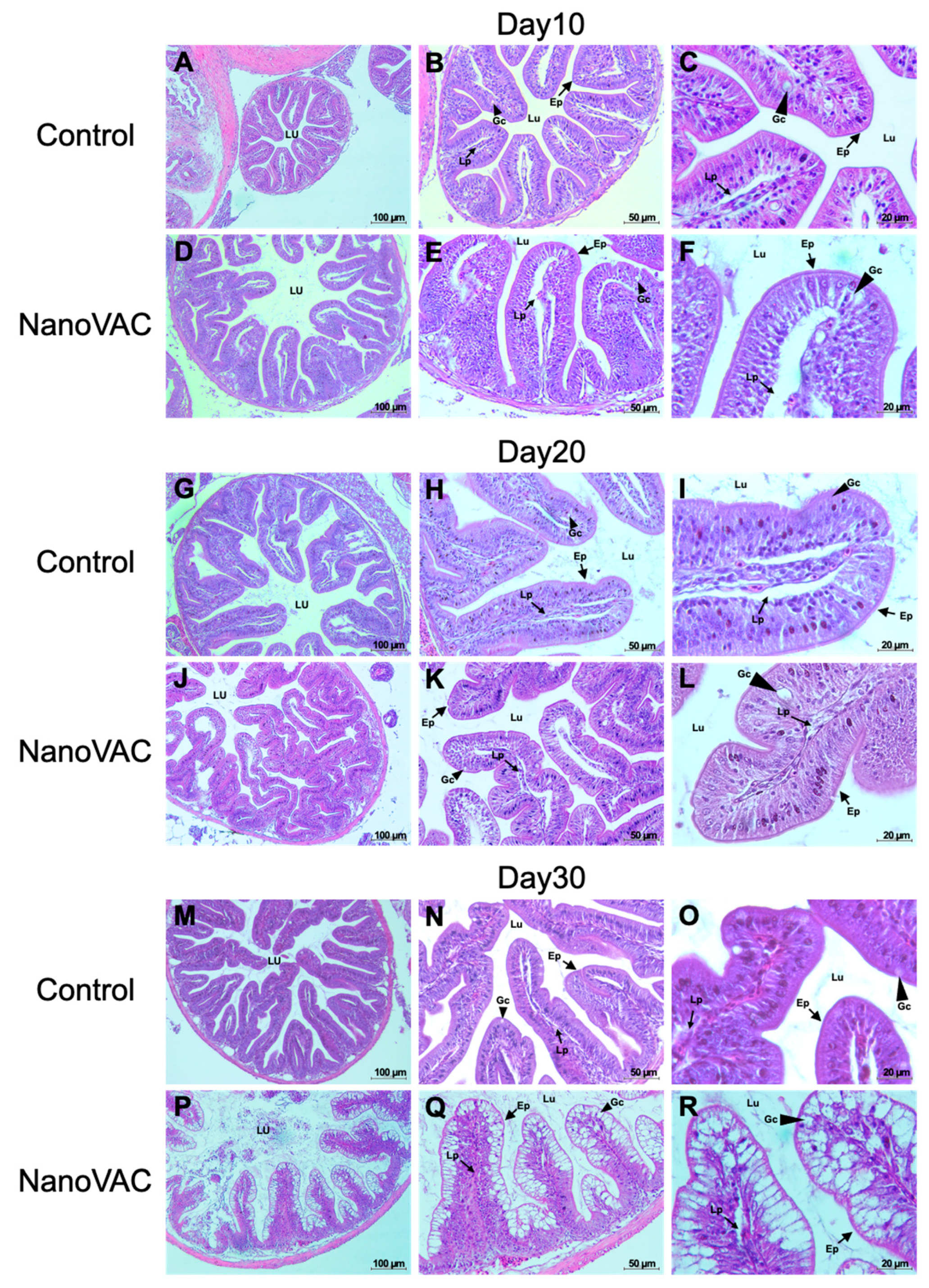
3.4. Effects of Bivalent Nanovaccine on Histopathological Changes in MALTs
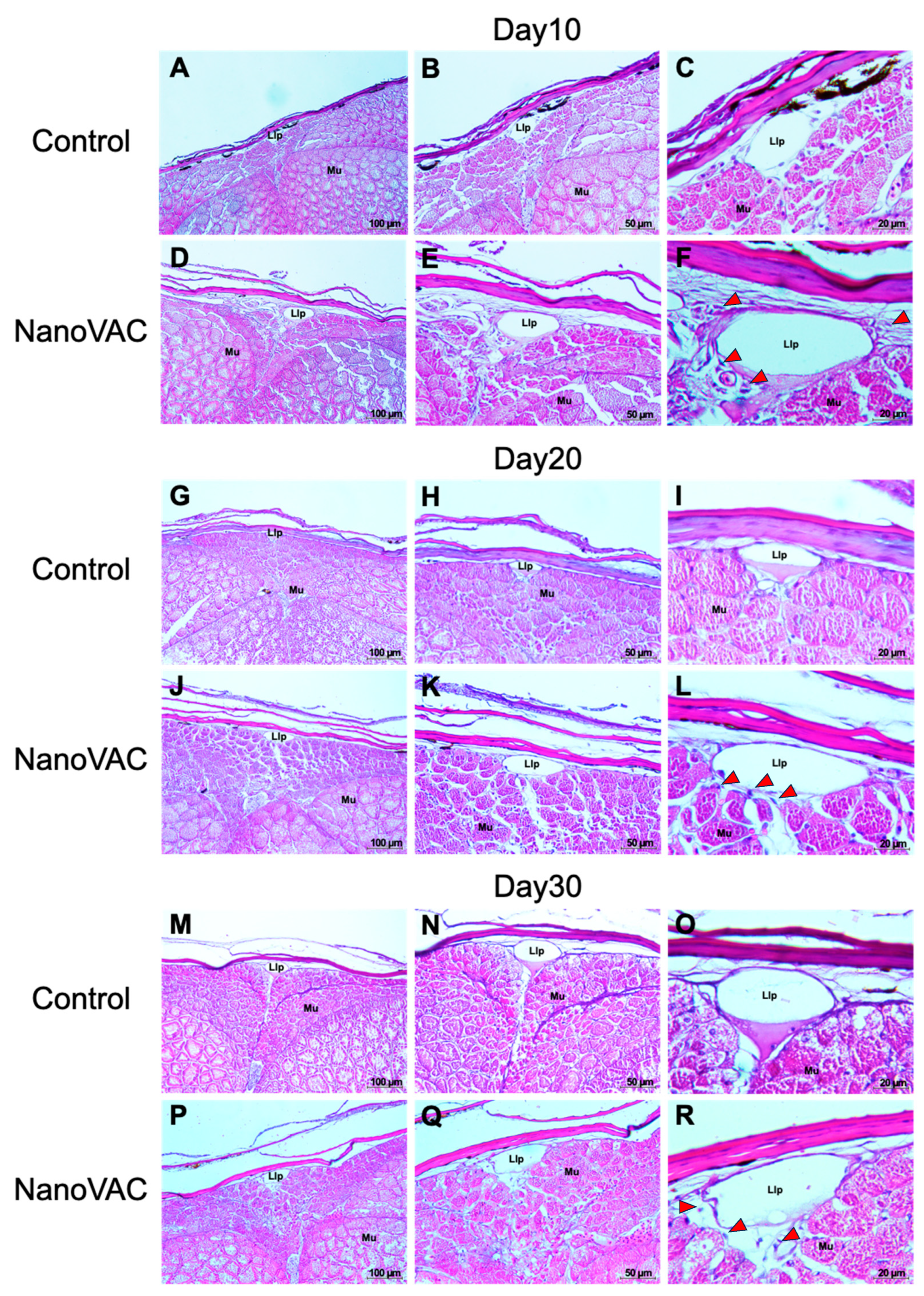
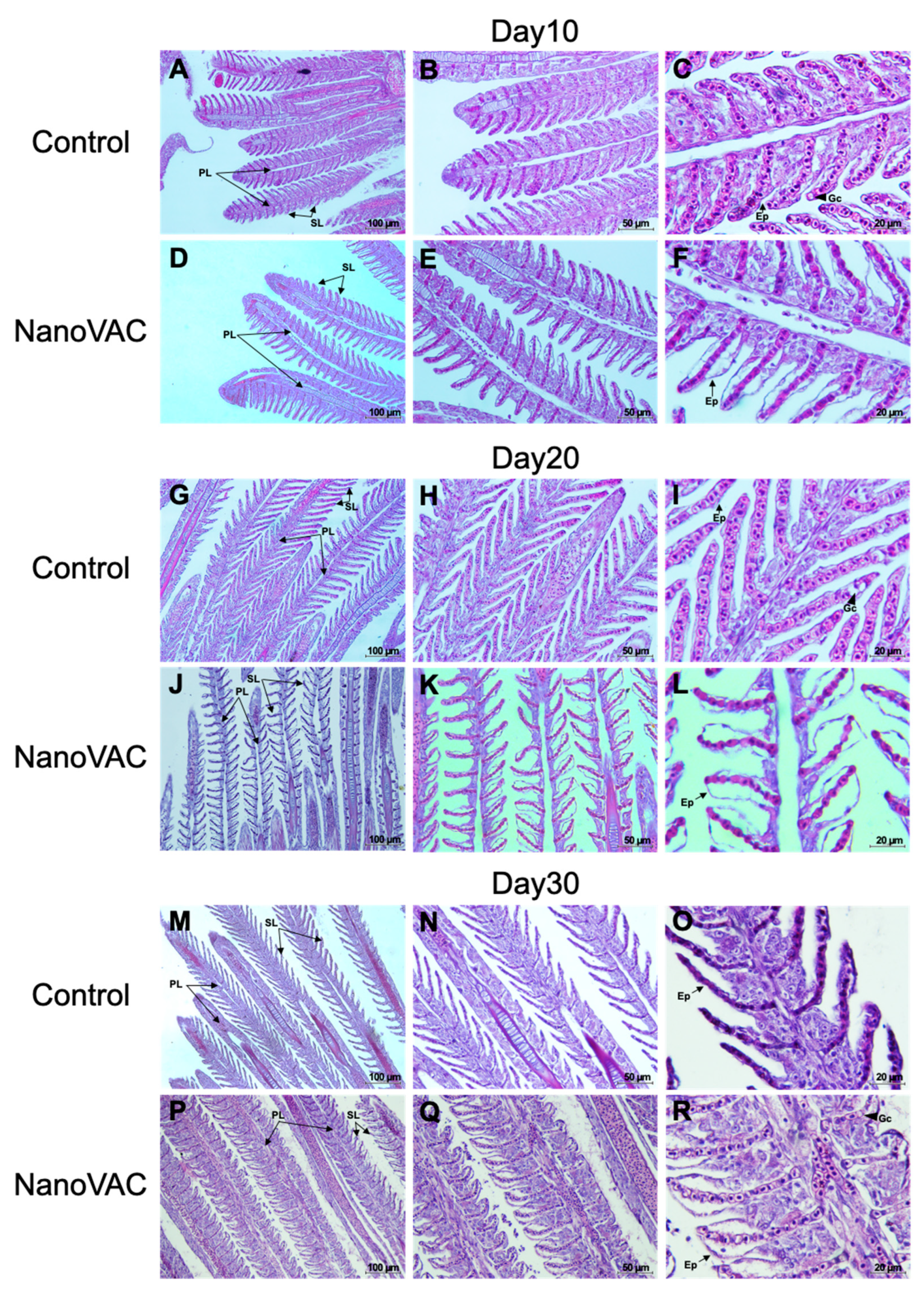
3.4.1. SALT
3.4.2. GIALT
3.4.3. GALT
3.5. Growth Performance
3.6. Challenge Tests with S. iniae and F. covae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lawley, D. Repositioning Australian Farmed Barramundi: Online Consumer Survey Findings; University of Sunshine Coast: Sippy Downs, QLD, Australia, 2010. [Google Scholar]
- Robinson, N.A.; Schipp, G.; Bosmans, J.; Jerry, D.R. Modelling selective breeding in protandrous, batch-reared Asian sea bass (Lates calcarifer, Bloch) using walkback selection. Aquac. Res. 2010, 41, e643–e655. [Google Scholar] [CrossRef]
- Bromage, E.S.; Thomas, A.; Owens, L. Streptococcus iniae, a bacterial infection in barramundi, Lates calcarifer. Dis. Aquat. Org. 1999, 36, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Declercq, A.M.; Haesebrouck, F.; Van den Broeck, W.; Bossier, P.; Decostere, A. Columnaris disease in fish: A review with emphasis on bacterium-host interactions. Vet. Res. 2013, 44, 27. [Google Scholar] [CrossRef] [PubMed]
- Suanyuk, N.; Sukkasame, N.; Tanmark, N.; Yoshida, T.; Itami, T.; Thune, R.; Tantikitti, C.; Supamattaya, K. Streptococcus iniae infection in cultured Asian sea bass (Lates calcarifer) and red tilapia (Oreochromis sp.) in southern Thailand. Songklanakarin J. Sci. Technol. 2010, 32, 341–348. [Google Scholar]
- Tran Vi, H.; Dang Ha, Q.V.; Nguyen Huu, D.; Wergeland, H.I. Experimental Streptococcus iniae infection in barramundi (Lates calcarifer) cultured in Vietnam. Int. J. Aquat. Sci. 2013, 4, 3–12. [Google Scholar]
- Quesada, S.P.; Paschoal, J.A.R.; Reyes, F.G.R. Considerations on the aquaculture development and on the use of veterinary drugs: Special issue for fluoroquinolones da review. J. Food Sci. 2013, 78, R1321–R1333. [Google Scholar] [CrossRef]
- Vinitnantharat, S.; Gravningen, K.; Greger, E. Fish vaccines. Adv. Vet. Med. 1999, 41, 539–550. [Google Scholar]
- Dadar, M.; Dhama, K.; Vakharia, V.N.; Hoseinifar, S.H.; Karthik, K.; Tiwari, R.; Khandia, R.; Munjal, A.; Salgado-Miranda, C.; Joshi, S.K. Advances in aquaculture vaccines against fish pathogens: Global status and current trends. Rev. Fish. Sci. Aquac. 2017, 25, 184–217. [Google Scholar] [CrossRef]
- Plant, K.P.; LaPatra, S.E. Advances in fish vaccine delivery. Dev. Comp. Immunol. 2011, 35, 1256–1262. [Google Scholar] [CrossRef]
- Mondal, H.; Thomas, J. A review on the recent advances and application of vaccines against fish pathogens in aquaculture. Aquac. Int. 2022, 30, 1971–2000. [Google Scholar] [CrossRef]
- Bunnoy, A.; Thompson, K.D.; Thangsunan, P.; Chokmangmeepisarn, P.; Yata, T.; Pirarat, N.; Kitiyodom, S.; Thangsunan, P.; Sukkarun, P.; Prukbenjakul, P.; et al. Development of a bivalent mucoadhesive nanovaccine to prevent francisellosis and columnaris diseases in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2023, 138, 108813. [Google Scholar] [CrossRef] [PubMed]
- Kitiyodom, S.; Yata, T.; Thompson, K.D.; Costa, J.; Elumalai, P.; Katagiri, T.; Temisak, S.; Namdee, K.; Rodkhum, C.; Pirarat, N. Immersion vaccination by a biomimetic-mucoadhesive nanovaccine induces humoral immune response of red tilapia (Oreochromis sp.) against Flavobacterium columnare challenge. Vaccines 2021, 9, 1253. [Google Scholar] [CrossRef] [PubMed]
- Thangsunan, P.; Kitiyodom, S.; Srisapoome, P.; Pirarat, N.; Yata, T.; Thangsunan, P.; Boonrungsiman, S.; Bunnoy, A.; Rodkhum, C. Novel development of cationic surfactant-based mucoadhesive nanovaccine for direct immersion vaccination against Francisella noatunensis subsp. orientalis in red tilapia (Oreochromis sp.). Fish Shellfish Immunol. 2022, 127, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Angulo, C.; Tello-Olea, M.; Reyes-Becerril, M.; Monreal-Escalante, E.; Hernández-Adame, L.; Angulo, M.; Mazon-Suastegui, J.M. Developing oral nanovaccines for fish: A modern trend to fight infectious diseases. Rev. Aquac. 2021, 13, 1172–1192. [Google Scholar] [CrossRef]
- Kent, M.L.; Feist, S.W.; Harper, C.; Hoogstraten-Miller, S.; Mac Law, J.; Sanchez-Morgado, J.M.; Tanguay, R.L.; Sanders, G.E.; Spitsbergen, J.M.; Whipps, C.M. Recommendations for control of pathogens and infectious diseases in fish research facilities. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 240–248. [Google Scholar] [CrossRef]
- Tumree, P.; Bunnoy, A.; Tang, X.; Srisapoome, P. Efficacy of the whole-cell based monovalent and bivalent vaccines of Streptococcus iniae and Flavobacterium covae in fingering Asian seabass (Lates calcarifer). Fish Shellfish Immunol. 2023, 144, 109269. [Google Scholar] [CrossRef]
- Kitiyodom, S.; Yata, T.; Yostawornkul, J.; Kaewmalun, S.; Nittayasut, N.; Suktham, K.; Surassmo, S.; Namdee, K.; Rodkhum, C.; Pirarat, N. Enhanced efficacy of immersion vaccination in tilapia against columnaris disease by chitosan-coated “pathogen-like” mucoadhesive nanovaccines. Fish Shellfish Immunol. 2019, 95, 213–219. [Google Scholar] [CrossRef]
- Fischer, A.H.; Jacobson, K.A.; Rose, J.; Zeller, R. Hematoxylin and eosin staining of tissue and cell sections. Cold Spring Harb. Protoc. 2008, 49, 86. [Google Scholar] [CrossRef]
- Ito, Y.; Yamada, H.; Imoto, T. Colorimetric assay for lysozyme using Micrococcus luteus labeled with a blue dye, remazol brilliant blue R, as a substrate. Chem. Pharm. Bull. 1992, 40, 1523–1526. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bunnoy, A.; Na-Nakorn, U.; Srisapoome, P. Probiotic effects of a novel strain, Acinetobacter KU011TH, on the growth performance, immune responses, and resistance against Aeromonas hydrophila of bighead catfish (Clarias macrocephalus Günther, 1864). Microorganisms 2019, 7, 613. [Google Scholar] [CrossRef] [PubMed]
- Kayansamruaj, P.; Areechon, N.; Unajak, S. Development of fish vaccine in Southeast Asia: A challenge for the sustainability of SE Asia aquaculture. Fish Shellfish Immunol. 2020, 103, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Chokmangmeepisarn, P.; Thangsunan, P.; Kayansamruaj, P.; Rodkhum, C. Resistome characterization of Flavobacterium columnare isolated from freshwater cultured Asian sea bass (Lates calcarifer) revealed diversity of quinolone resistance-associated genes. Aquaculture 2021, 544, 737149. [Google Scholar] [CrossRef]
- Gudding, R.; Van Muiswinkel, W.B. A history of fish vaccination: Science-based disease prevention in aquaculture. Fish Shellfish Immunol. 2013, 35, 1683–1688. [Google Scholar] [CrossRef] [PubMed]
- Leal, C.A.G.; Carvalho-Castro, G.A.; Sacchetin, P.S.C.; Lopes, C.O.; Moraes, A.M.; Figueiredo, H.C.P. Oral and parenteral vaccines against Flavobacterium columnare: Evaluation of humoral immune response by ELISA and in vivo efficiency in Nile tilapia (Oreochromis niloticus). Aquac. Int. 2010, 18, 657–666. [Google Scholar] [CrossRef]
- Priya, T.J.; Kappalli, S. Modern biotechnological strategies for vaccine development in aquaculture-prospects and challenges. Vaccine 2022, 40, 5873–5881. [Google Scholar] [CrossRef] [PubMed]
- Bunnoy, A.; Thangsunan, P.; Chokmangmeepisarn, P.; Yata, T.; Klongklaew, N.; Pirarat, N.; Kitiyodom, S.; Srisapoome, P.; Rodkhum, C. Mucoadhesive cationic lipid-based Flavobacterium oreochromis nanoencapsulation enhanced the efficacy of mucoadhesive immersion vaccination against columnaris disease and strengthened immunity in Asian sea bass (Lates calcarifer). Fish Shellfish Immunol. 2022, 127, 633–646. [Google Scholar] [CrossRef]
- Mody, N.; Sharma, R.; Agrawal, U.; Vyas, S.P. Nanocarriers: A versatile approach for mucosal vaccine delivery. Ther. Deliv. 2015, 6, 231–245. [Google Scholar] [CrossRef]
- Sukkarun, P.; Kitiyodom, S.; Yostawornkul, J.; Chaiin, P.; Yata, T.; Rodkhum, C.; Boonrungsiman, S.; Pirarat, N. Chitosan-polymer based nanovaccine as promising immersion vaccine against Aeromonas veronii challenge in red tilapia (Oreochromis sp.). Fish Shellfish Immunol. 2022, 129, 30–35. [Google Scholar] [CrossRef]
- Pumchan, A.; Sae-Ueng, U.; Prasittichai, C.; Sirisuay, S.; Areechon, N.; Unajak, S. A novel efficient piscine oral nano-vaccine delivery system: Modified halloysite nanotubes (HNTs) preventing streptococcosis disease in tilapia (Oreochromis sp.). Vaccines 2022, 10, 1180. [Google Scholar] [CrossRef]
- Zhang, L.; Zeng, Z.; Hu, C.; Bellis, S.L.; Yang, W.; Su, Y.; Zhang, X.; Wu, Y. Controlled and targeted release of antigens by the intelligent shell for improving the applicability of oral vaccines. Biomaterials 2016, 77, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.-M.; Wei, X.-F.; Zhou, G.-Q.; Liu, M.-Z.; Li, P.-F.; Zhu, B. Mannose functionalized biomimetic nanovaccine enhances immune responses against tilapia lake virus. Aquaculture 2022, 560, 738535. [Google Scholar] [CrossRef]
- Ramos, E.A.; Relucio, J.L.V.; Torres-Villanueva, C.A.T. Gene expression in tilapia following oral delivery of chitosan-encapsulated plasmid DNA incorporated into fish feeds. Mar. Biotechnol. 2005, 7, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.-F.; Gong, Y.-M.; Xia, J.-Y.; Liu, M.-Z.; Li, P.-F.; Wang, G.-X.; Zhu, B. Biomimetic nanovaccine based on erythrocyte membrane enhances immune response and protection against tilapia lake virus. Virology 2023, 580, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.D.; Rodkhum, C.; Bunnoy, A.; Thangsunan, P.; Kitiyodom, S.; Sukkarun, P.; Yostawornkul, J.; Yata, T. Addressing nanovaccine strategies for tilapia. Vaccines 2023, 11, 1356. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Gupta, M.; Gupta, V.; Gogoi, H.; Bhatnagar, R. Novel application of trimethyl chitosan as an adjuvant in vaccine delivery. Int. J. Nanomed. 2018, 13, 7959–7970. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Gao, Y.; Shu, J.; Zhang, C.; Zhao, K. Chitosan-based nanomaterial as immune adjuvant and delivery carrier for vaccines. Vaccines 2022, 10, 1906. [Google Scholar] [CrossRef] [PubMed]
- Plumb, J.A.; Vinitnantharat, S. Vaccination of channel catfish, Ictalurus punctatus (Rafinesque), by immersion and oral booster against Edwardsiella ictalurid. J. Fish Dis. 1993, 16, 65–71. [Google Scholar] [CrossRef]
- Thinh, N.; Kuo, T.; Hung, L.; Loc, T.; Chen, S.; Evensen, Ø.; Schuurman, H.J. Combined immersion and oral vaccination of Vietnamese catfish (Pangasianodon hypophthalmus) confers protection against mortality caused by Edwardsiella ictalurid. Fish Shellfish Immunol. 2009, 27, 773–776. [Google Scholar] [CrossRef]
- Dan, X.-M.; Zhang, T.-W.; Li, Y.-W.; Li, A.-X. Immune responses and immune-related gene expression profile in orange-spotted grouper after immunization with Cryptocaryon irritans vaccine. Fish Shellfish Immunol. 2013, 34, 885–891. [Google Scholar] [CrossRef]
- Gao, Y.; Tang, X.; Sheng, X.; Xing, J.; Zhan, W. Antigen uptake and expression of antigen presentation-related immune genes in flounder (Paralichthys olivaceus) after vaccination with an inactivated Edwardsiella tarda immersion vaccine, following hyperosmotic treatment. Fish Shellfish Immunol. 2016, 55, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Liu, Q.; Ni, C.; Li, S.; Wu, H.; Wang, Q.; Xiao, J.; Zhang, Y. Gene expression profiling in live attenuated Edwardsiella tarda vaccine immunized and challenged zebrafish: Insights into the basic mechanisms of protection seen in immunized fish. Dev. Comp. Immunol. 2013, 40, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Soto, E.; Griffin, M.J.; Tobar, J.A. Mucosal vaccines. In Mucosal Health in Aquaculture; Beck, B.H., Peatman, E., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 297–323. [Google Scholar]
- Aas, I.B.; Austbø, L.; König, M.; Syed, M.; Falk, K.; Hordvik, I.; Koppang, E.O. Transcriptional characterization of the T cell population within the salmonid interbranchial lymphoid tissue. J. Immunol. 2014, 193, 3463–3469. [Google Scholar] [CrossRef] [PubMed]
- Haaparanta, A.; Valtonen, E.; Hoffmann, R.W. Gill anomalies of perch and roach from four lakes differing in water quality. J. Fish Biol. 1997, 50, 575–591. [Google Scholar] [CrossRef]
- Mallatt, J. Fish gill structural changes induced by toxicants and other irritants: A statistical review. Can. J. Fish. Aquat. Sci. 1985, 42, 630–648. [Google Scholar] [CrossRef]
- Santos, R.M.B.; Monteiro, S.; Cortes, R.; Pacheco, F.; Fernandes, L.S. Seasonal effect of land use management on gill histopathology of Barbel and Douro Nase in a Portuguese watershed. Sci. Total Environ. 2021, 764, 142869. [Google Scholar] [CrossRef]
- Adelmann, M.; Köllner, B.; Bergmann, S.M.; Fischer, U.; Lange, B.; Weitschies, W.; Enzmann, P.-J.; Fichtner, D. Development of an oral vaccine for immunisation of rainbow trout (Oncorhynchus mykiss) against viral haemorrhagic septicaemia. Vaccine 2008, 26, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Korbut, R.; Mehrdana, F.; Kania, P.W.; Larsen, M.H.; Frees, D.; Dalsgaard, I.; Jørgensen, L.V.G. Antigen uptake during different life stages of zebrafish (Danio rerio) using a GFP-tagged Yersinia ruckeri. PLoS ONE 2016, 11, e0158968. [Google Scholar] [CrossRef]
- Ohtani, M.; Villumsen, K.R.; Koppang, E.O.; Raida, M.K. Global 3D imaging of Yersinia ruckeri bacterin uptake in rainbow trout fry. PLoS ONE 2015, 10, e0117263. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Names | Genes | Nucleotide Sequences (5′ → 3′) | Annealing Temperature (°C) | Product Size (bp) | Accession Number |
|---|---|---|---|---|---|
| Lc_β-actin | β-actin | F-5′-TACCCCATTGAGCACGGTATTG-3′ R-5′-TCTGGGTCATCTTCTCCCTGTT-3′ | 60 | 160 | XM_018667666.1 |
| Lc_IgM | Immunoglobulin M (IgM) (secreted form) | F-5′-TGTCAAGGTAAACGAGGGAGC-3′ R-5′-TCCCCTGGATCCATTCGTCA-3′ | 60 | 152 | ASM164080v1 |
| Lc_IgT | Immunoglobulin T (IgT) (secreted form) | F-5′-GAGGCAACTTACAGAGGAACCATA-3′ R-5′-CTGGTCACTTCTCCATCAATTTCC-3′ | 60 | 194 | ASM164080v1 |
| Lc_IgD | Immunoglobulin D (IgD) (membrane-bound form) | F-5′-GAGTGTGAATGTTGCTGGGC-3′ R-5′-TTGGCCTGAAAGGTGACGTA-3′ | 60 | 150 | ASM164080v1 |
| Lc_CD4 | CD4 receptor (CD4) | F-5′-AGTGCAATGGATTGGGGTAGATAA-3′ R-5′-GTTGCAGGCTCTGTAACTTTGATT-3′ | 60 | 156 | XM_018672258 |
| Lc_TCRα | T-cell receptor alpha (TCRα) | F-5′-GGCCGTTCGGATAGAAGGAG-3′ R-5′-AGAGCCATTGTGTTCACCGT-3′ | 60 | 153 | ASM164080v1 |
| Lc_MHCIIα | Major histocompatibility complex class IIα (MHCIIα) | F-5′-TTCCTACCTCCCTGATCTACCC-3′ R-5′-CTGAAGTCGCTGTTGGAGTAGT-3′ | 60 | 178 | ASM164080v1 |
| Bacterial Culture | Formulation | Average Diameter (nm) | Zeta Potential (mV) |
|---|---|---|---|
| Streptococcus iniae | Sonicated antigen (bacterial cells) | 203 ± 10 | −36.87 ± 0.93 |
| Polymeric nanovaccine (solution form) | 246 ± 16 | 45.39 ± 1.31 | |
| Polymeric nanovaccine (dry form) | 304 ± 25 | 47.60 ± 0.96 | |
| Flavobacterium covae | Sonicated antigen (bacterial cells) | 324 ± 6 | −21.86 ± 0.89 |
| Polymeric nanovaccine (solution form) | 394 ± 14 | 38.25 ± 1.06 | |
| Polymeric nanovaccine (dry form) | 426 ± 18 | 36.48 ± 1.02 |
| Growth Parameters | Treatments | |||
|---|---|---|---|---|
| Control | NanoVAC-G2 | NanoVAC-G3 | NanoVAC-G4 | |
| Weight gain (WG, g) | 0.542 ± 0.16 | 0.545 ± 0.17 | 0.552 ± 0.12 | 0.544 ± 0.15 |
| Specific growth rate (SGR, %/day) | 7.18 ± 0.75 | 7.18 ± 0.72 | 7.24 ± 0.79 | 7.19 ± 0.74 |
| Average daily gain (ADG, g/fish/day) | 0.0181 ± 0.0016 | 0.0182 ± 0.0018 | 0.0184 ± 0.0014 | 0.0181 ± 0.0011 |
| Feed conversion ratio (FCR) | 1.85 ± 0.44 | 1.83 ± 0.37 | 1.81 ± 0.39 | 1.84 ± 0.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meachasompop, P.; Bunnoy, A.; Keaswejjareansuk, W.; Dechbumroong, P.; Namdee, K.; Srisapoome, P. Development of Immersion and Oral Bivalent Nanovaccines for Streptococcosis and Columnaris Disease Prevention in Fry and Fingerling Asian Seabass (Lates calcarifer) Nursery Farms. Vaccines 2024, 12, 17. https://doi.org/10.3390/vaccines12010017
Meachasompop P, Bunnoy A, Keaswejjareansuk W, Dechbumroong P, Namdee K, Srisapoome P. Development of Immersion and Oral Bivalent Nanovaccines for Streptococcosis and Columnaris Disease Prevention in Fry and Fingerling Asian Seabass (Lates calcarifer) Nursery Farms. Vaccines. 2024; 12(1):17. https://doi.org/10.3390/vaccines12010017
Chicago/Turabian StyleMeachasompop, Pakapon, Anurak Bunnoy, Wisawat Keaswejjareansuk, Piroonrat Dechbumroong, Katawut Namdee, and Prapansak Srisapoome. 2024. "Development of Immersion and Oral Bivalent Nanovaccines for Streptococcosis and Columnaris Disease Prevention in Fry and Fingerling Asian Seabass (Lates calcarifer) Nursery Farms" Vaccines 12, no. 1: 17. https://doi.org/10.3390/vaccines12010017
APA StyleMeachasompop, P., Bunnoy, A., Keaswejjareansuk, W., Dechbumroong, P., Namdee, K., & Srisapoome, P. (2024). Development of Immersion and Oral Bivalent Nanovaccines for Streptococcosis and Columnaris Disease Prevention in Fry and Fingerling Asian Seabass (Lates calcarifer) Nursery Farms. Vaccines, 12(1), 17. https://doi.org/10.3390/vaccines12010017