

Unconjugated Multi-Epitope Peptides Adjuvanted with ALFQ Induce Durable and Broadly Reactive Antibodies to Human and Avian Influenza Viruses

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Influenza Viruses
2.3. Influenza Peptides
2.4. ALFQ Formulation with Composite Influenza Peptides
2.5. Murine Immunizations using ALFQ Formulated Composite Peptides (LHNVD-105)
2.6. Antisera ELISA: Detection of Antibodies That Bind to Influenza Peptides and Viruses
2.7. Microneutralization Assay (MNA) for Determination of Serum Neutralizing Antibodies
2.8. Phylogenetic Reconstructions of Influenza Strains
2.9. Statistical Analysis
3. Results
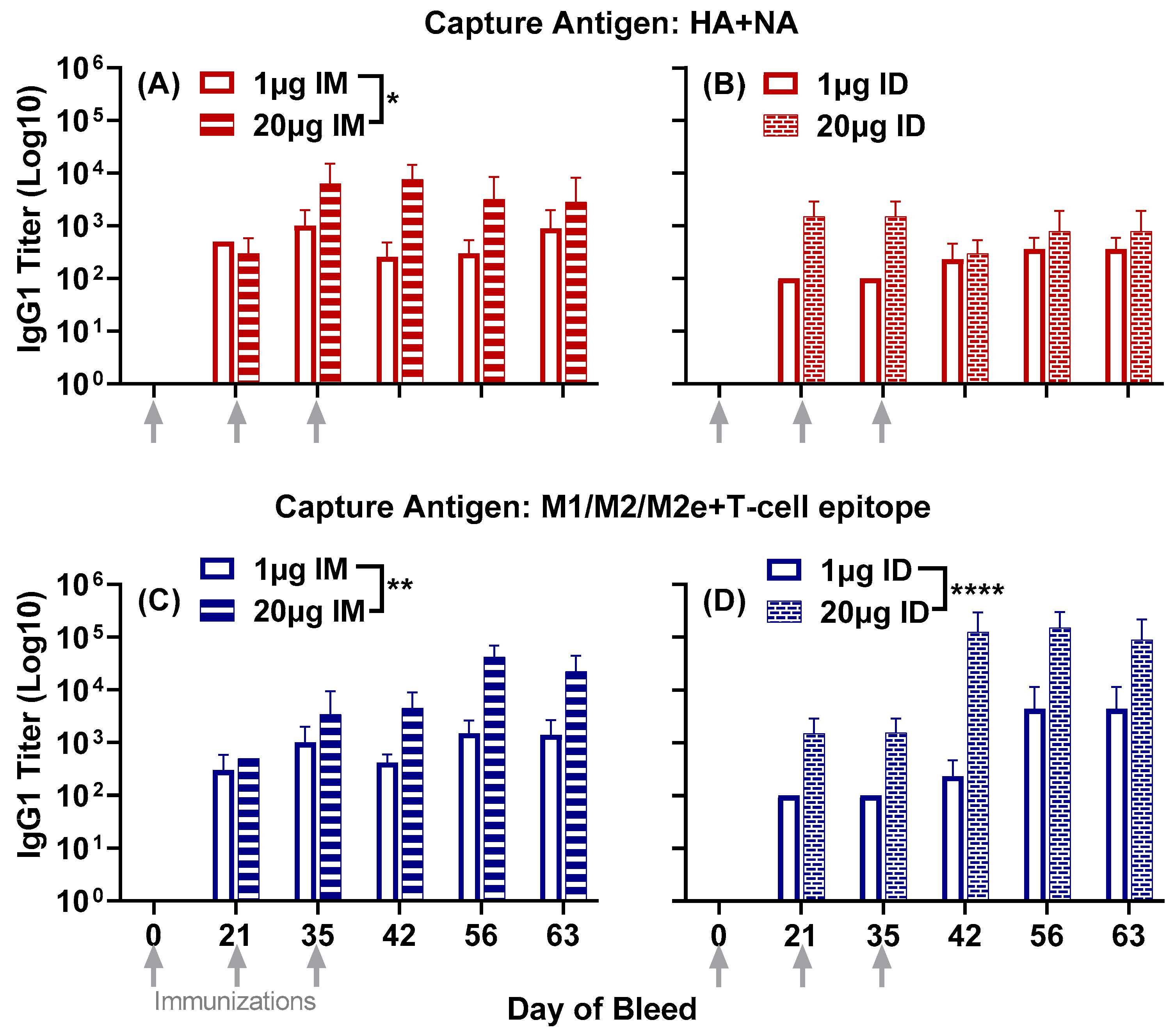
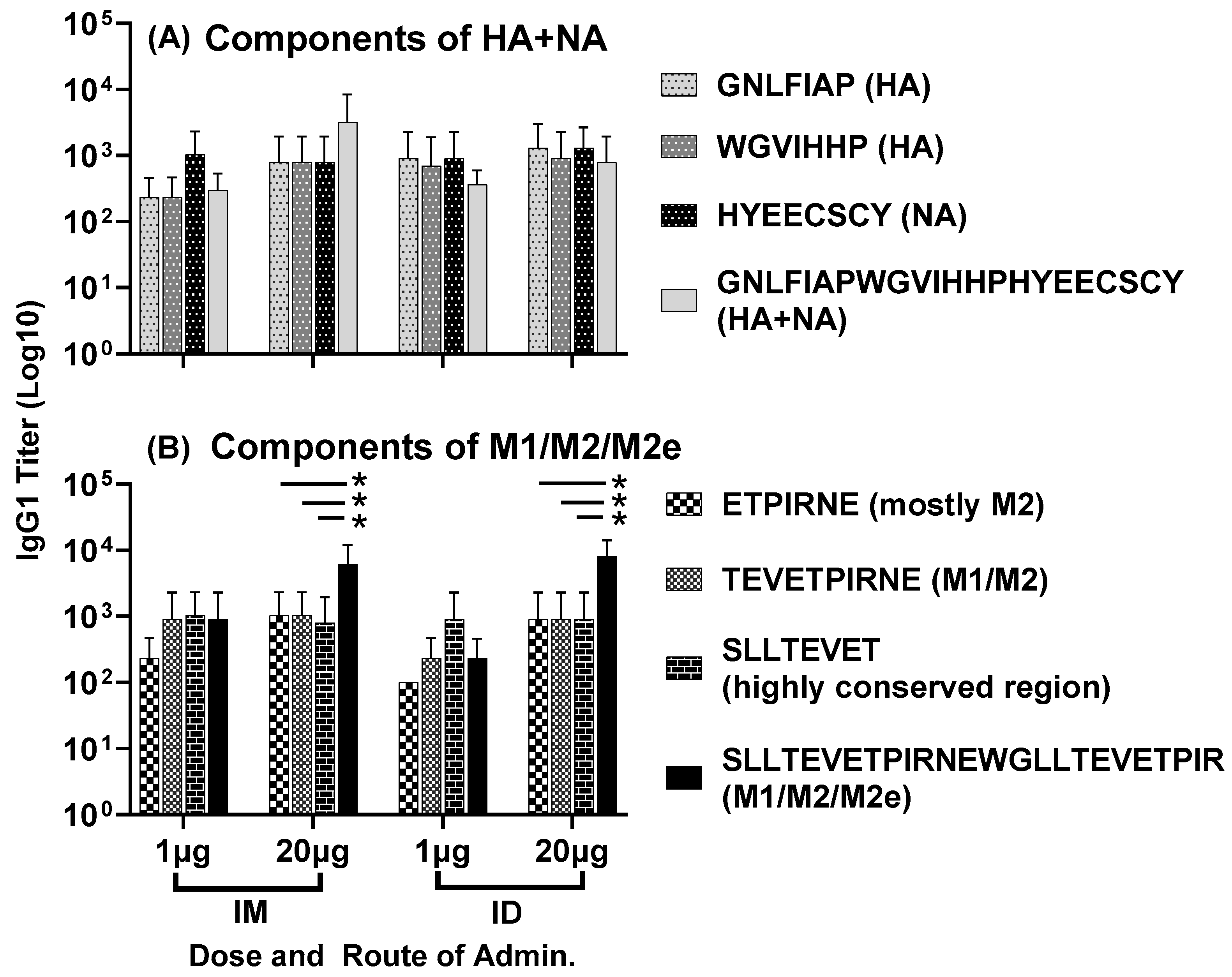
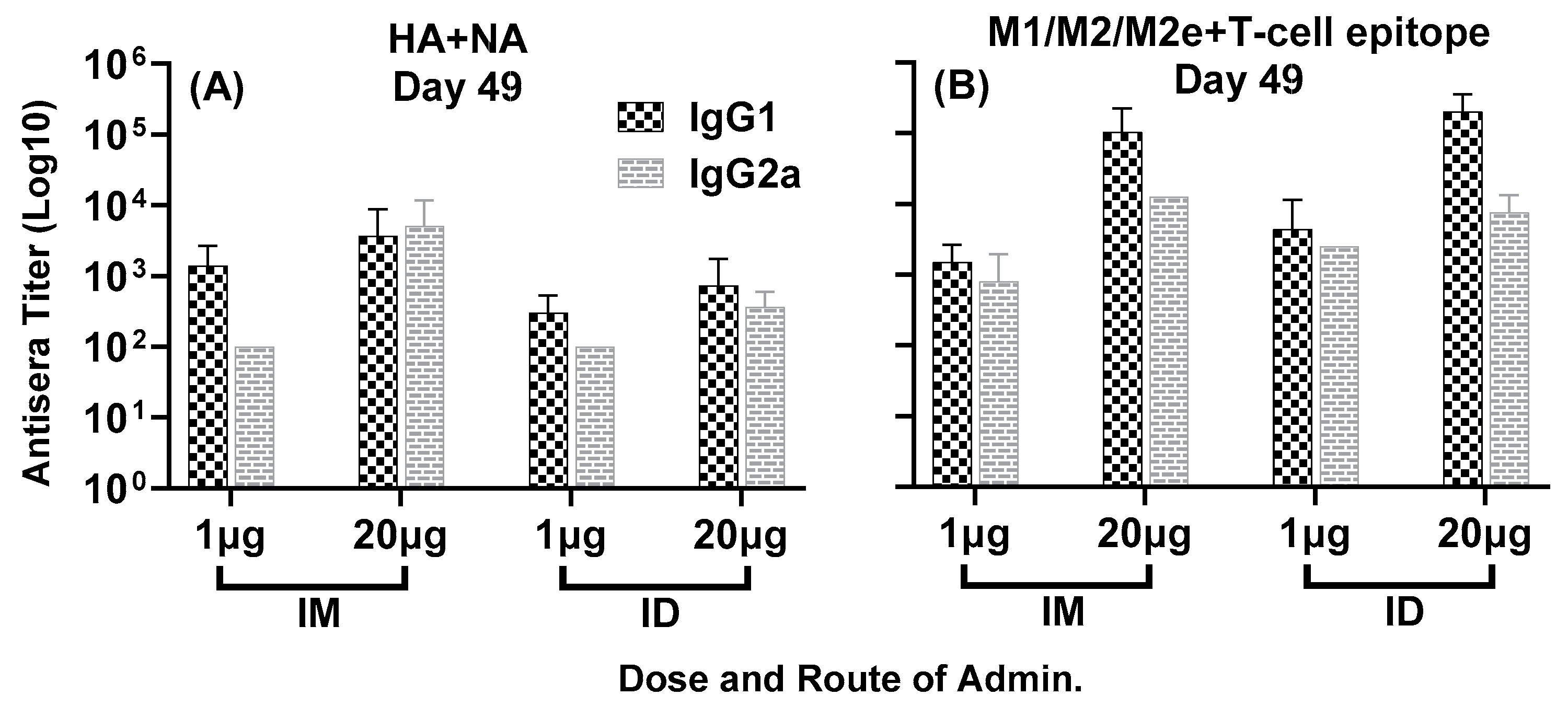
3.1. Antisera Responses to Composite Peptide Immunogens and Individual Peptide Components
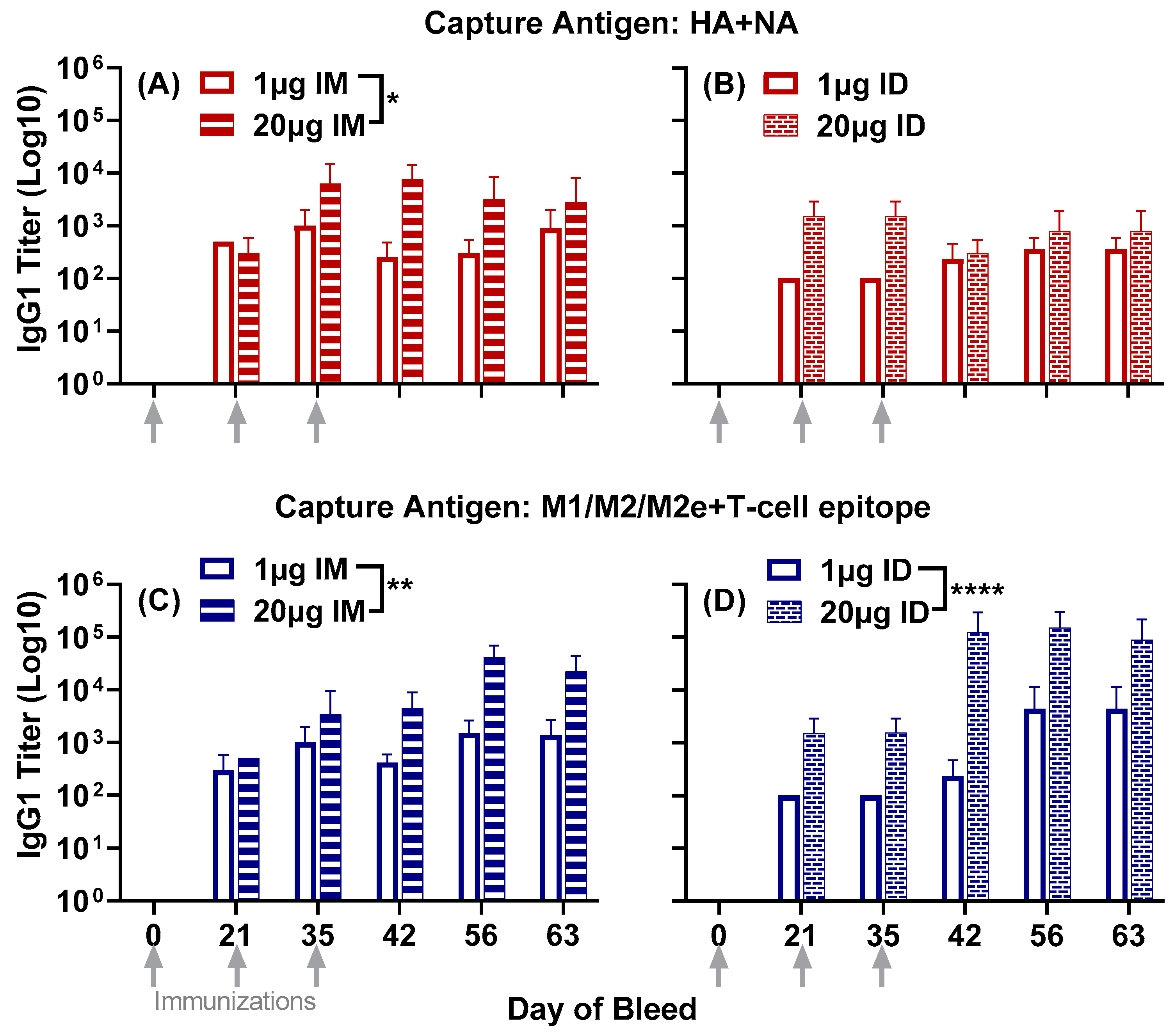

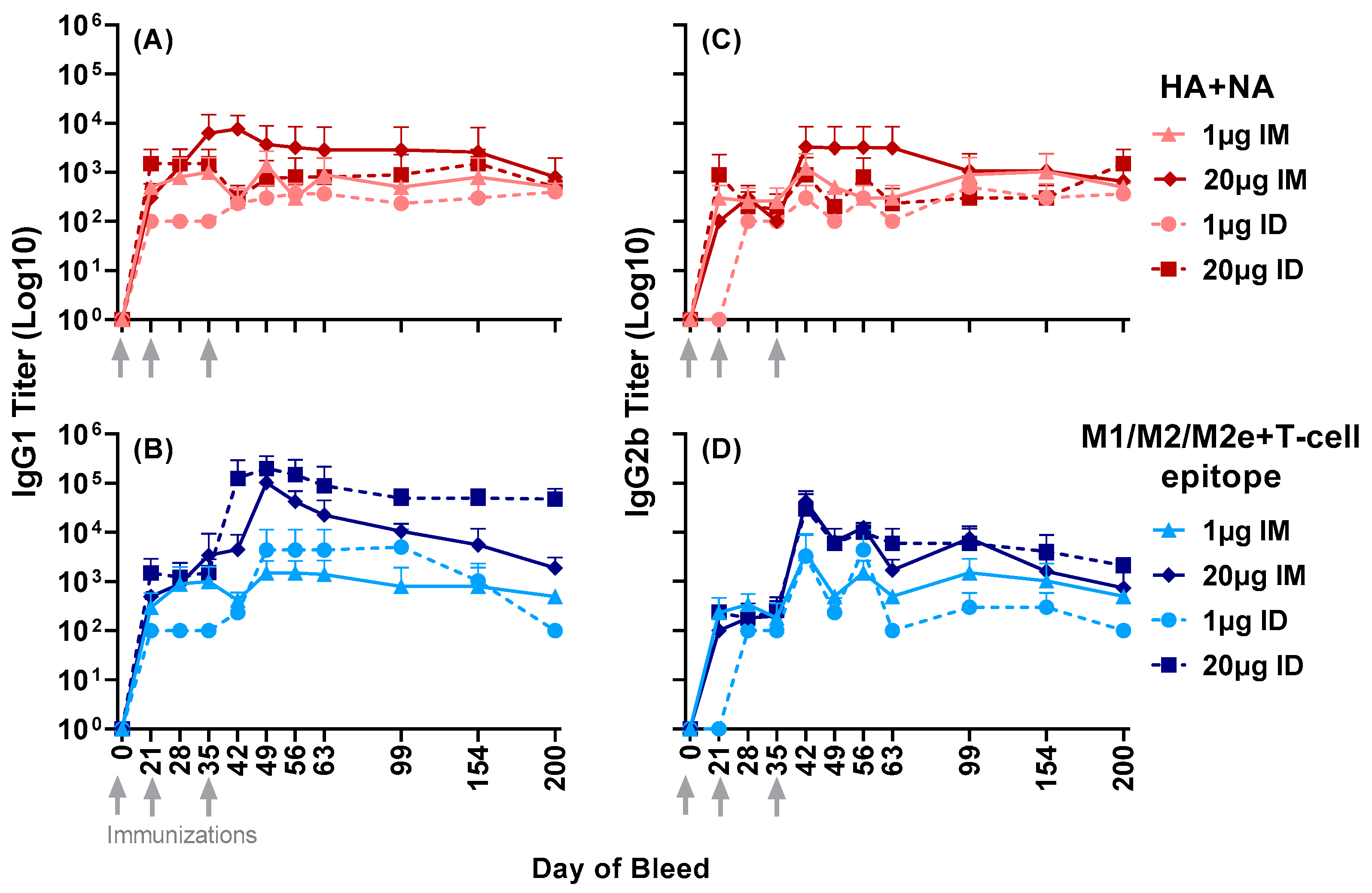


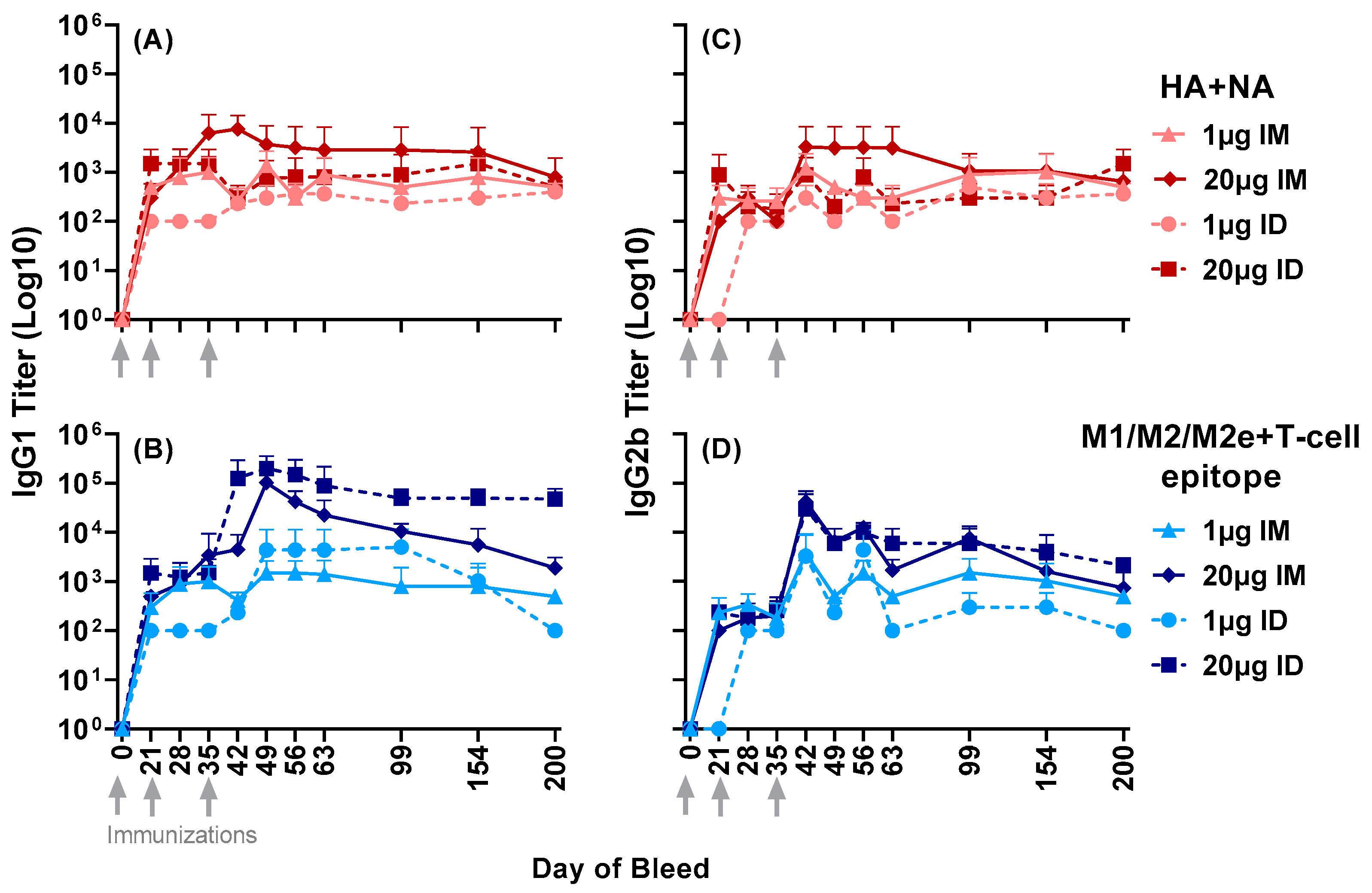
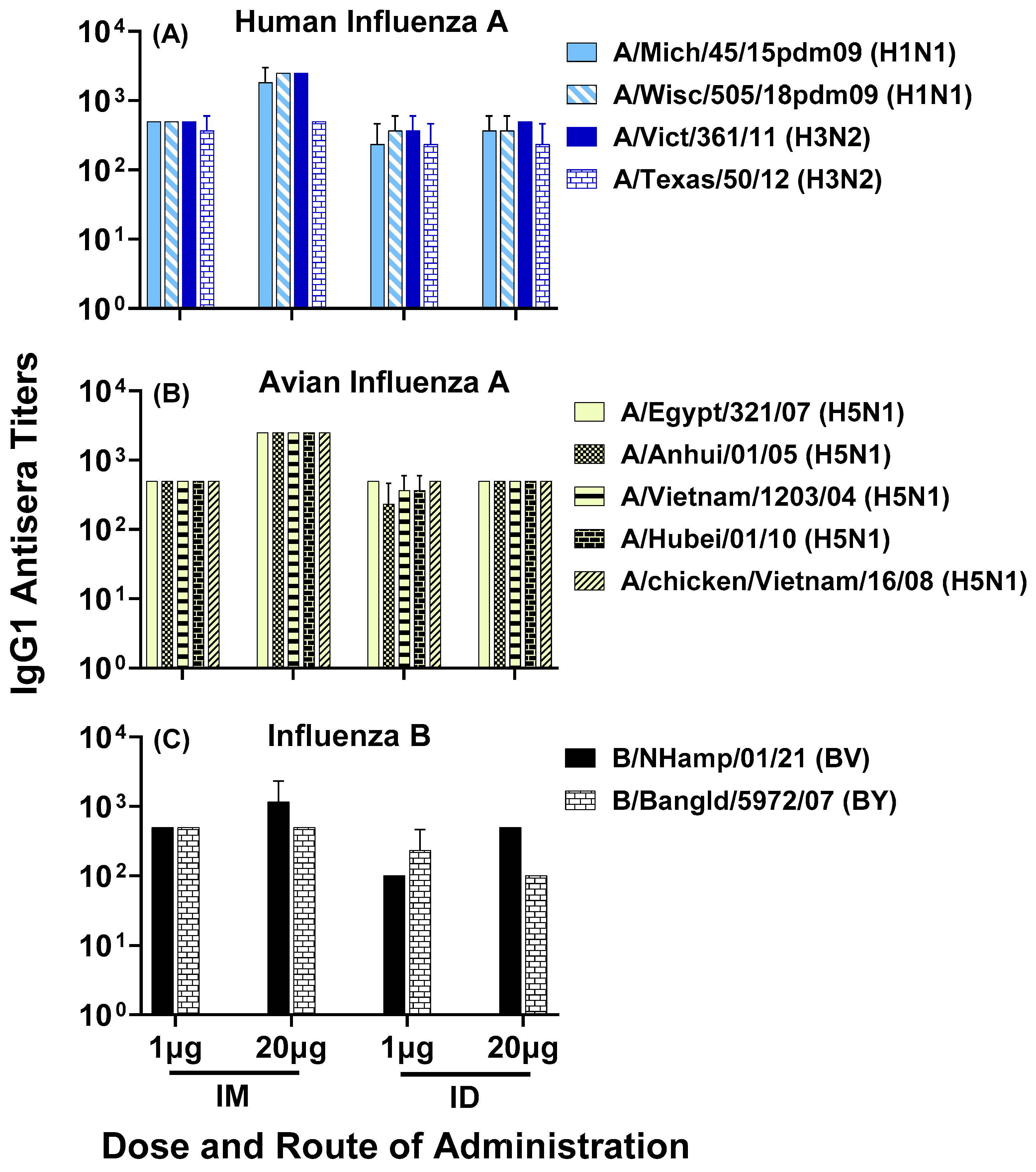
3.2. Antisera Titers to Influenza A and B Viruses

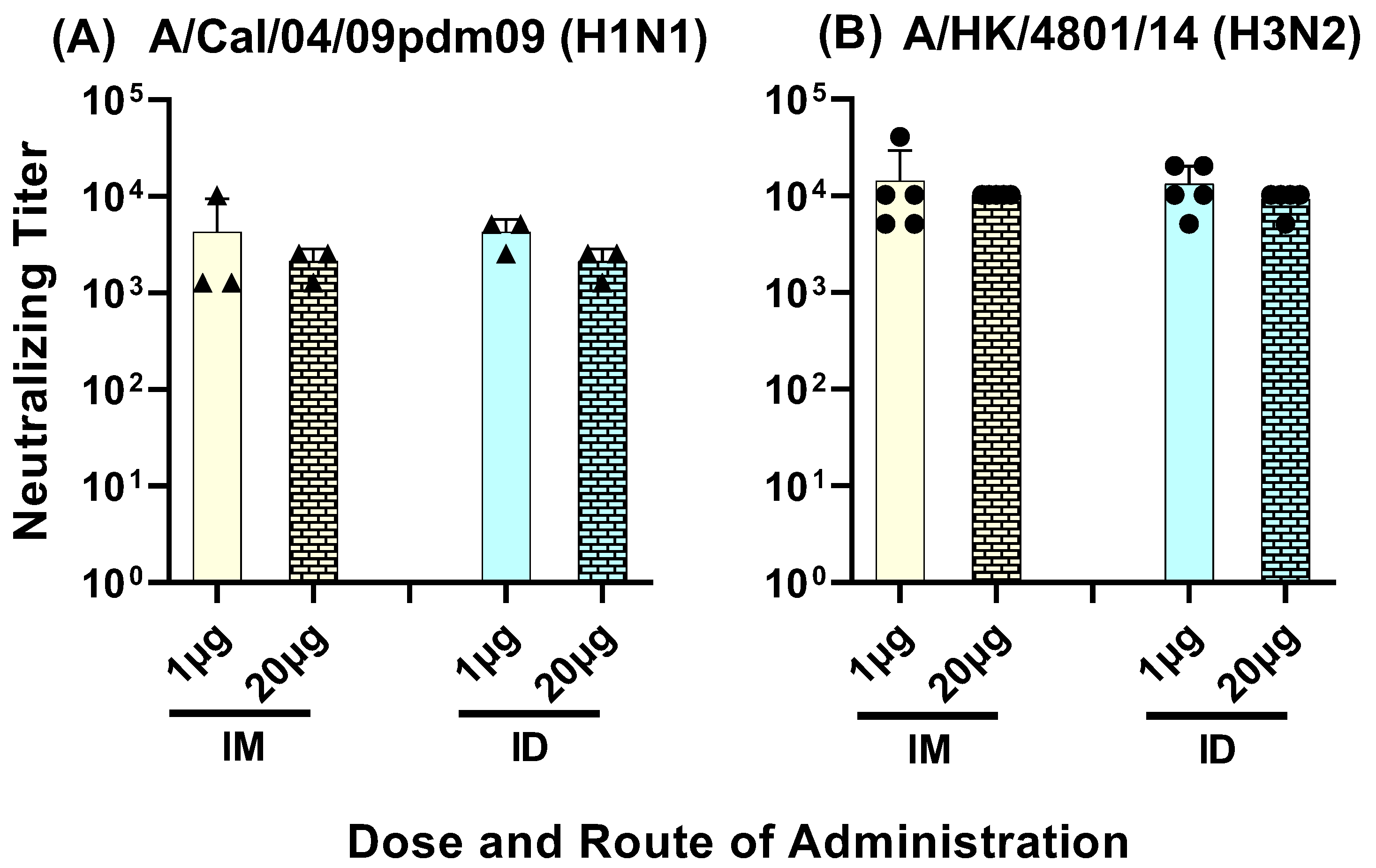
3.3. Serum Neutralizing Titers against Group 1 and 2 Influenza A Viruses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Agor, J.K.; Özaltın, O.Y. Models for predicting the evolution of influenza to inform vaccine strain selection. Hum. Vaccine Immunother. 2018, 14, 678–683. [Google Scholar] [CrossRef]
- Harding, A.; Heaton, N. Efforts to improve the seasonal influenza vaccine. Vaccines 2018, 6, 19. [Google Scholar] [CrossRef]
- Wang, W.-C.; Sayedahmed, E.E.; Sambhara, S.; Mittal, S.K. Progress towards the Development of a Universal Influenza Vaccine. Viruses 2022, 14, 1684. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Palese, P. Advances in the development of influenza virus vaccines. Nat. Rev. Drug Discov. 2015, 14, 167–182. [Google Scholar] [CrossRef]
- Romeli, S.; Hassan, S.S.; Yap, W.B. Multi-Epitope Peptide-Based and Vaccinia-Based Universal Influenza Vaccine Candidates Subjected to Clinical Trials. Malays. J. Med. Sci. 2020, 27, 10–20. [Google Scholar] [CrossRef]
- Pillet, S.; Couillard, J.; Trépanier, S.; Poulin, J.-F.; Yassine-Diab, B.; Guy, B.; Ward, B.J.; Landry, N. Immunogenicity and safety of a quadrivalent plant-derived virus like particle influenza vaccine candidate—Two randomized Phase II clinical trials in 18 to 49 and ≥50 years old adults. PLoS ONE 2019, 14, e0216533. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Hurst, B.; Shakya, A.K.; Uddin, M.J.; Ingrole, R.S.; Hernandez-Sanabria, M.; Arya, R.; Bimler, L.; Paust, S.; Tarbet, E.B.; et al. Consensus M2e peptide conjugated to gold nanoparticles confers protection against H1N1, H3N2 and H5N1 influenza A viruses. Antivir. Res. 2017, 141, 62–72. [Google Scholar] [CrossRef]
- Antrobus, R.D.; Coughlan, L.; Berthoud, T.K.; Dicks, M.D.; Hill, A.V.S.; Lambe, T.; Gilbert, S.C. Clinical Assessment of a Novel Recombinant Simian Adenovirus ChAdOx1 as a Vectored Vaccine Expressing Conserved Influenza A Antigens. Mol. Ther. 2014, 22, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Andersen, T.K.; Bodin, J.; Oftung, F.; Bogen, B.; Mjaaland, S.; Grødeland, G. Pandemic Preparedness Against Influenza: DNA Vaccine for Rapid Relief. Front. Immunol. 2021, 12, 747032. [Google Scholar] [CrossRef]
- Magini, D.; Giovani, C.; Mangiavacchi, S.; Maccari, S.; Cecchi, R.; Ulmer, J.B.; Gregorio, E.D.; Geall, A.J.; Brazzoli, M.; Bertholet, S. Self-Amplifying mRNA Vaccines Expressing Multiple Conserved Influenza Antigens Confer Protection against Homologous and Heterosubtypic Viral Challenge. PLoS ONE 2016, 11, e0161193. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Erbelding, E.J.; Post, D.J.; Stemmy, E.J.; Roberts, P.C.; Augustine, A.D.; Ferguson, S.; Paules, C.I.; Graham, B.S.; Fauci, A.S. A universal influenza vaccine: The strategic plan for the National Institute of Allergy and Infectious Diseases. J. Infect. Dis. 2018, 218, 347–354. [Google Scholar] [CrossRef]
- Francis, M.J. Recent Advances in Vaccine Technologies. Vet. Clin. Small Anim. 2018, 48, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Sei, C.J.; Rao, M.; Schuman, R.F.; Daum, L.T.; Matyas, G.R.; Rikhi, N.; Muema, K.; Anderson, A.; Jobe, O.; Kroscher, K.A.; et al. Conserved Influenza Hemagglutinin, Neuraminidase and Matrix Peptides Adjuvanted with ALFQ Induce Broadly Neutralizing Antibodies. Vaccines 2021, 9, 698. [Google Scholar] [CrossRef] [PubMed]
- Hermanson, G.T. Bioconjugate Techniques; Elsevier: Cambridge, MA, USA, 2013. [Google Scholar]
- Hutter, J.N.; Robben, P.M.; Lee, C.; Hamer, M.; Moon, J.E.; Merino, K.; Zhu, L.; Galli, H.; Quinn, X.; Brown, D.R.; et al. First-in-human assessment of safety and immunogenicity of low and high doses of Plasmodium falciparum malaria protein 013 (FMP013) administered intramuscularly with ALFQ adjuvant in healthy malaria-naïve adults. Vaccine 2022, 40, 5781–5790. [Google Scholar] [CrossRef] [PubMed]
- Alving, C.R.; Rao, M.; Matyas, G.R. Similarities and differences of chemical compositions and physical and functional properties of adjuvant system 01 and army liposome formulation with QS21. Front. Immunol. 2023, 14, 1102524. [Google Scholar] [CrossRef] [PubMed]
- Panina-Bordignon, P.; Tan, A.; Termijtelen, A.; Demotz, S.; Corradin, G.; Lanzavecchia, A. Universally immunogenic T cell epitopes: Promiscuous binding to human MHC class II and promiscuous recognition by T cells. Eur. J. Immunol. 1989, 19, 2237–2242. [Google Scholar] [CrossRef]
- Imai, M.; Herfst, S.; Sorrell, E.M.; Schrauwen, E.J.A.; Linster, M.; De Graaf, M.; Fouchier, R.A.M.; Kawaoka, Y. Transmission of influenza A/H5N1 viruses in mammals. Virus Res. 2013, 178, 15–20. [Google Scholar] [CrossRef]
- Tsybalova, L.M.; Stepanova, L.A.; Ramsay, E.S.; Vasin, A.V. Influenza B: Prospects for the Development of Cross-Protective Vaccines. Viruses 2022, 14, 1323. [Google Scholar] [CrossRef]
- Ran, Z.; Shen, H.; Lang, Y.; Kolb, E.A.; Turan, N.; Zhu, L.; Ma, J.; Bawa, B.; Liu, Q.; Liu, H.; et al. Domestic Pigs Are Susceptible to Infection with Influenza B Viruses. J. Virol. 2015, 89, 4818–4826. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-P.; Tsai, H.-J. Influenza B viruses in pigs, Taiwan. Influenza Other Respir Viruses. 2019, 13, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Sinha, N.K.; Roy, A.; Das, B.; Das, S.; Basak, S. Evolutionary complexities of swine flu H1N1 gene sequences of 2009. Biochem. Biophys. Res. Commun. 2009, 390, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Garten, R.J.; Davis, C.T.; Russell, C.A.; Shu, B.; Lindstrom, S.; Balish, A.; Sessions, W.M.; Xu, X.; Skepner, E.; Deyde, V.; et al. Antigenic and Genetic Characteristics of the Early Isolates of Swine-Origin 2009 A(H1N1) Influenza Viruses Circulating in Humans. Science 2009, 325, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Ma, W. Swine influenza virus: Current status and challenge. Virus Res. 2020, 288, 198118. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Xiao, Y.; Liu, J.; Wang, D.; Li, F.; Wang, C.; Li, C.; Zhu, J.; Song, J.; Sun, H.; et al. Prevalent Eurasian avian-like H1N1 swine influenza virus with 2009 pandemic viral genes facilitating human infection. Proc. Natl. Acad. Sci. USA 2020, 117, 17204–17210. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Per Cent Endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Beck, Z.; Matyas, G.R.; Jalah, R.; Rao, M.; Polonis, V.R.; Alving, C.R. Differential immune responses to HIV-1 envelope protein induced by liposomal adjuvant formulations containing monophosphoryl lipid A with or without QS21. Vaccine 2015, 33, 5578–5587. [Google Scholar] [CrossRef]
- Matyas, G.R.; Muderhwa, J.M.; Alving, C.R. Oil-in-water liposomal emulsions for vaccine delivery. Methods Enzym. 2003, 373, 34–50. [Google Scholar]
- Elbe, S.; Buckland-Merrett, G. Data, disease and diplomacy: GISAID’s innovative contribution to global health. Glob. Chall. 2017, 1, 33–46. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kenney, R.T.; Frech, S.A.; Muenz, L.R.; Villar, C.P.; Glenn, G.M. Dose Sparing with Intradermal Injection of Influenza Vaccine. N. Engl. J. Med. 2004, 351, 2295–2301. [Google Scholar] [CrossRef] [PubMed]
- Marra, F.; Young, F.; Richardson, K.; Marra, C.A. A Meta-analysis of intradermal versus intramuscular influenza vaccines: Immunogenicity and Adverse Events. Influenza Other Respir. Viruses 2013, 7, 584–603. [Google Scholar] [CrossRef] [PubMed]
- Egunsola, O.; Clement, F.; Taplin, J.; Mastikhina, L.; Li, J.W.; Lorenzetti, D.L.; Dowsett, L.E.; Noseworthy, T. Immunogenicity and Safety of Reduced-Dose Intradermal vs Intramuscular Influenza Vaccines: A Systematic Review and Meta-analysis. JAMA Netw. Open 2021, 4, e2035693. [Google Scholar] [CrossRef]
- Menon, I.; Bagwe, P.; Gomes, K.B.; Bajaj, L.; Gala, R.; Uddin, M.N.; D’Souza, M.J.; Zughaier, S.M. Microneedles: A New Generation Vaccine Delivery System. Micromachines 2021, 12, 435. [Google Scholar] [CrossRef]
- Hung, I.F.N.; Yuen, K.-Y. Immunogenicity, safety and tolerability of intradermal influenza vaccines. Hum. Vaccin. Immunother. 2018, 14, 565–570. [Google Scholar] [CrossRef]
- Zhao, G.; Lin, Y.; Du, L.; Guan, J.; Sun, S.; Sui, H.; Kou, Z.; Chan, C.C.-S.; Guo, Y.; Jiang, S.; et al. An M2e-based multiple antigenic peptide vaccine protects mice from lethal challenge with divergent H5N1 influenza viruses. Virol. J. 2010, 7, 9. [Google Scholar] [CrossRef]
- Zhao, G.; Sun, S.; Du, L.; Xiao, W.; Ru, Z.; Kou, Z.; Guo, Y.; Yu, H.; Jiang, S.; Lone, Y.; et al. An H5N1 M2e-based multiple antigenic peptide vaccine confers heterosubtypic protection from lethal infection with pandemic 2009 H1N1 virus. Virol. J. 2010, 7, 151. [Google Scholar] [CrossRef]
- Leung, H.-C.; Chan, C.C.-S.; Poon, V.K.-M.; Zhao, H.-J.; Cheung, C.-Y.; Ng, F.; Huang, J.-D.; Zheng, B.-J. An H5N1-based matrix protein 2 ectodomain tetrameric peptide vaccine provides cross-protection against lethal infection with H7N9 influenza virus. Emerg. Microbes Infect. 2015, 4, e22. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rikhi, N.; Sei, C.J.; Rao, M.; Schuman, R.F.; Kroscher, K.A.; Matyas, G.R.; Muema, K.; Lange, C.; Assiaw-Dufu, A.; Hussin, E.; et al. Unconjugated Multi-Epitope Peptides Adjuvanted with ALFQ Induce Durable and Broadly Reactive Antibodies to Human and Avian Influenza Viruses. Vaccines 2023, 11, 1468. https://doi.org/10.3390/vaccines11091468
Rikhi N, Sei CJ, Rao M, Schuman RF, Kroscher KA, Matyas GR, Muema K, Lange C, Assiaw-Dufu A, Hussin E, et al. Unconjugated Multi-Epitope Peptides Adjuvanted with ALFQ Induce Durable and Broadly Reactive Antibodies to Human and Avian Influenza Viruses. Vaccines. 2023; 11(9):1468. https://doi.org/10.3390/vaccines11091468
Chicago/Turabian StyleRikhi, Nimisha, Clara J. Sei, Mangala Rao, Richard F. Schuman, Kellie A. Kroscher, Gary R. Matyas, Kevin Muema, Camille Lange, Aba Assiaw-Dufu, Elizabeth Hussin, and et al. 2023. "Unconjugated Multi-Epitope Peptides Adjuvanted with ALFQ Induce Durable and Broadly Reactive Antibodies to Human and Avian Influenza Viruses" Vaccines 11, no. 9: 1468. https://doi.org/10.3390/vaccines11091468
APA StyleRikhi, N., Sei, C. J., Rao, M., Schuman, R. F., Kroscher, K. A., Matyas, G. R., Muema, K., Lange, C., Assiaw-Dufu, A., Hussin, E., Jobe, O., Alving, C. R., & Fischer, G. W. (2023). Unconjugated Multi-Epitope Peptides Adjuvanted with ALFQ Induce Durable and Broadly Reactive Antibodies to Human and Avian Influenza Viruses. Vaccines, 11(9), 1468. https://doi.org/10.3390/vaccines11091468