
Augmenting Vaccine Efficacy against Delta Variant with ‘Mycobacterium-w’-Mediated Modulation of NK-ADCC and TLR-MYD88 Pathways

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Immunological Monitoring
2.2. Statistics
2.3. RNA Sequencing (RNA-Seq) Analysis
2.4. Gene Ontology (GO) Pathway Analysis
3. Results
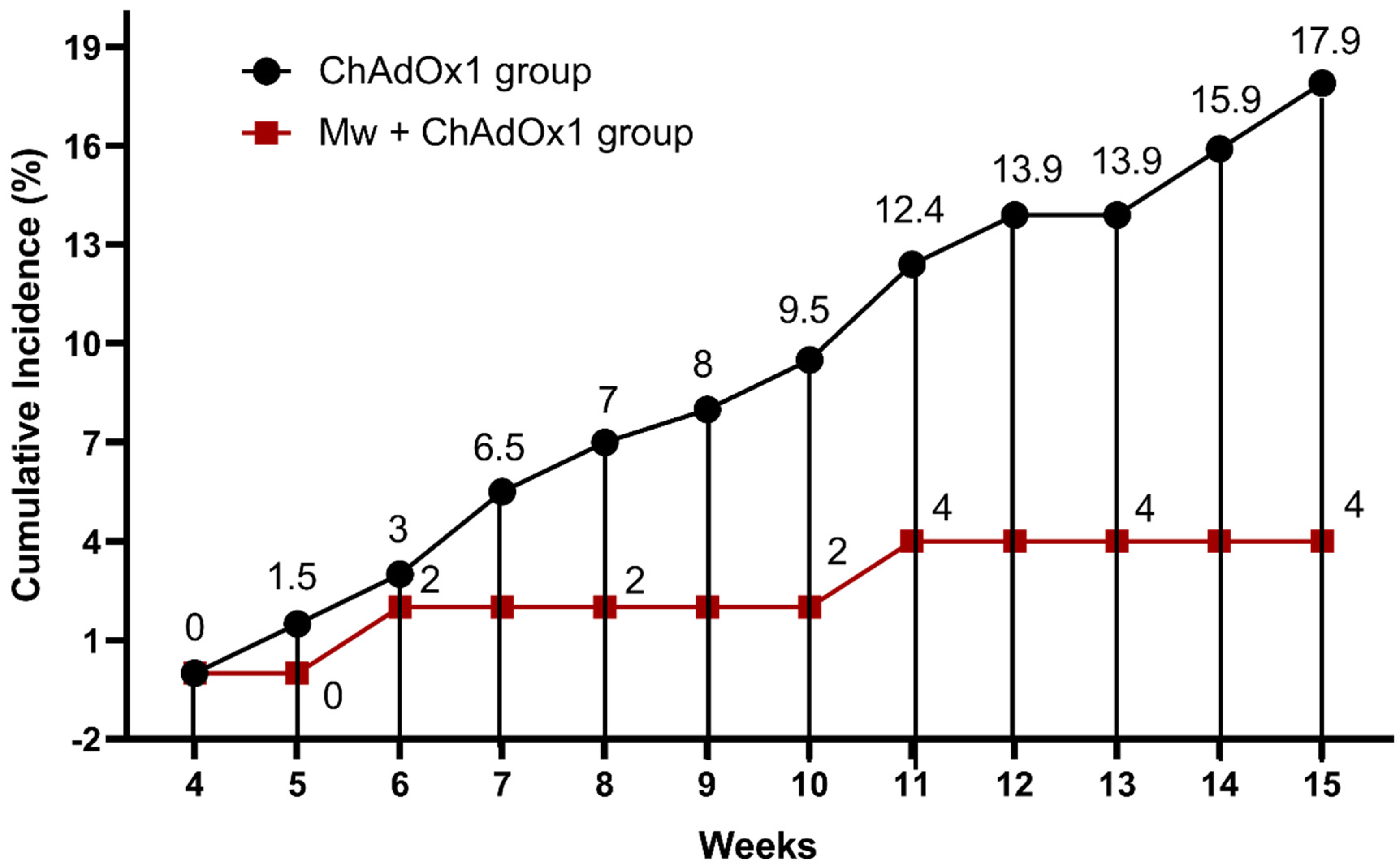
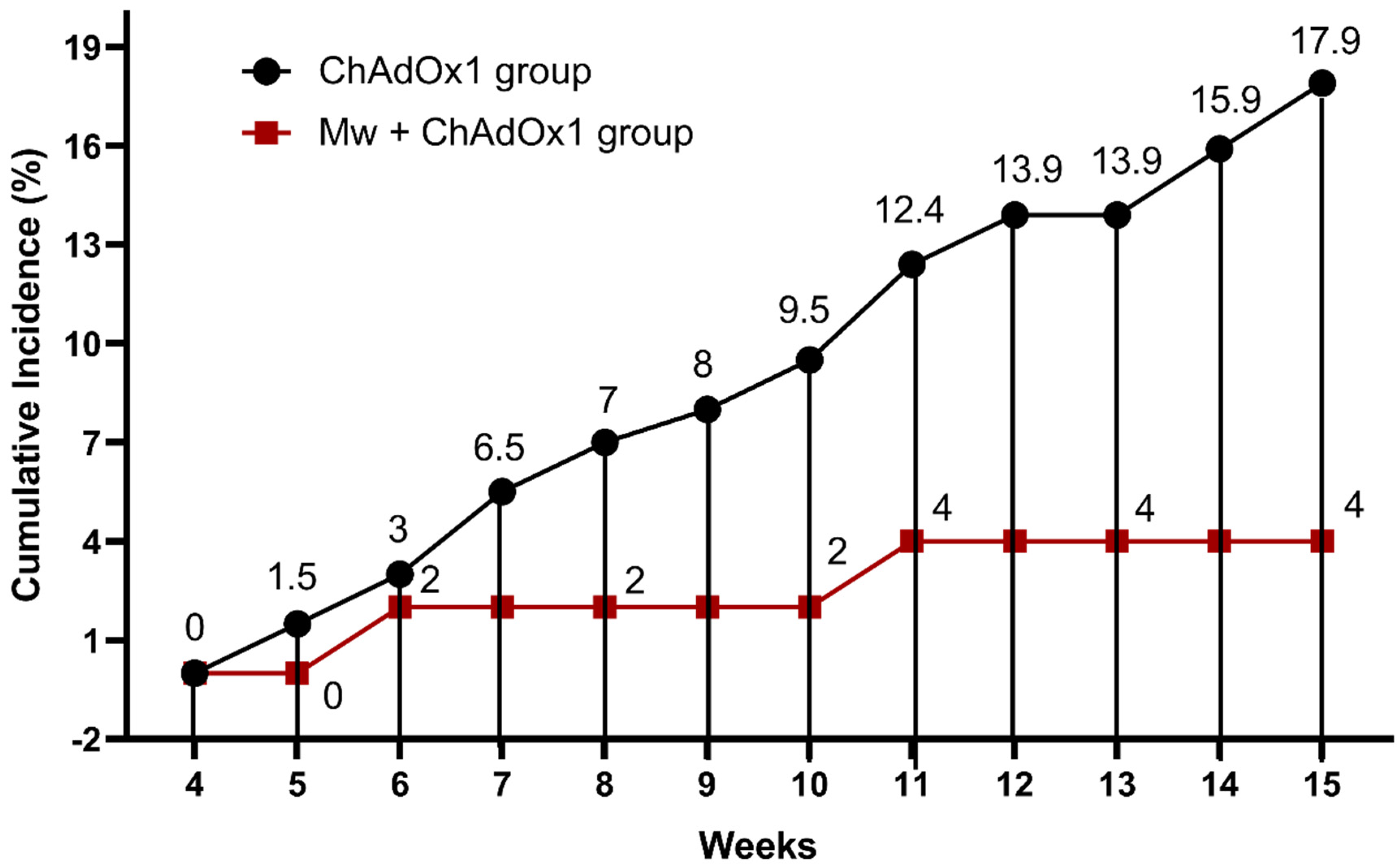
3.1. Effect of Mw Priming on Incidence and Outcome of COVID-19
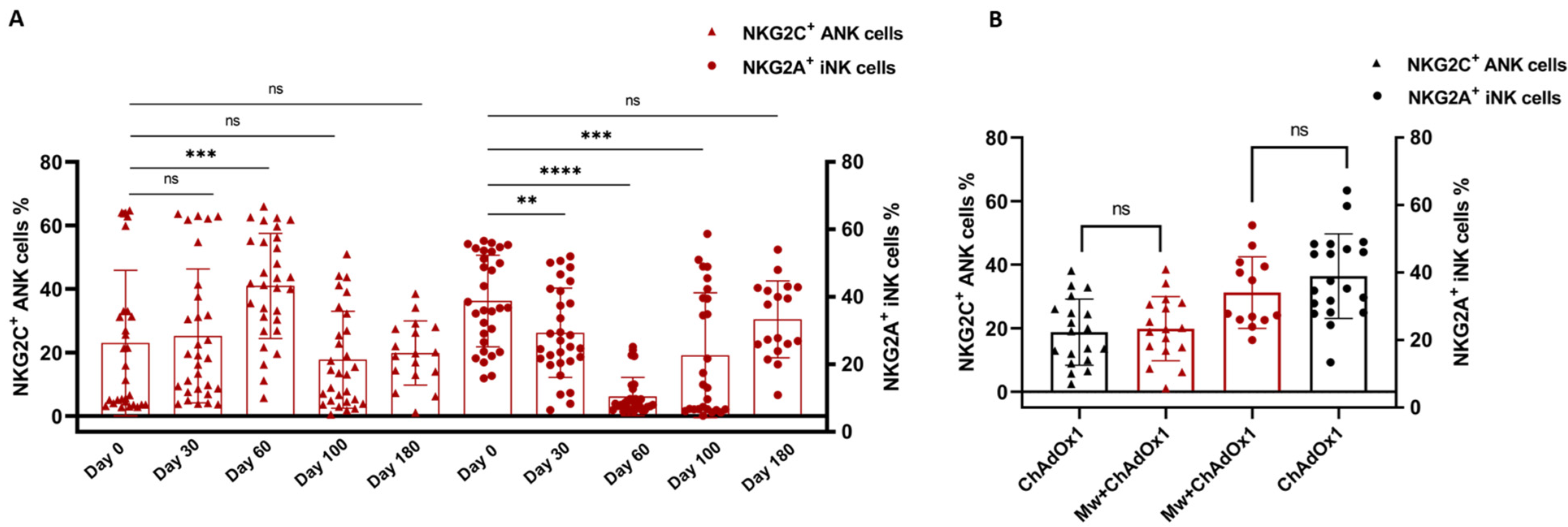
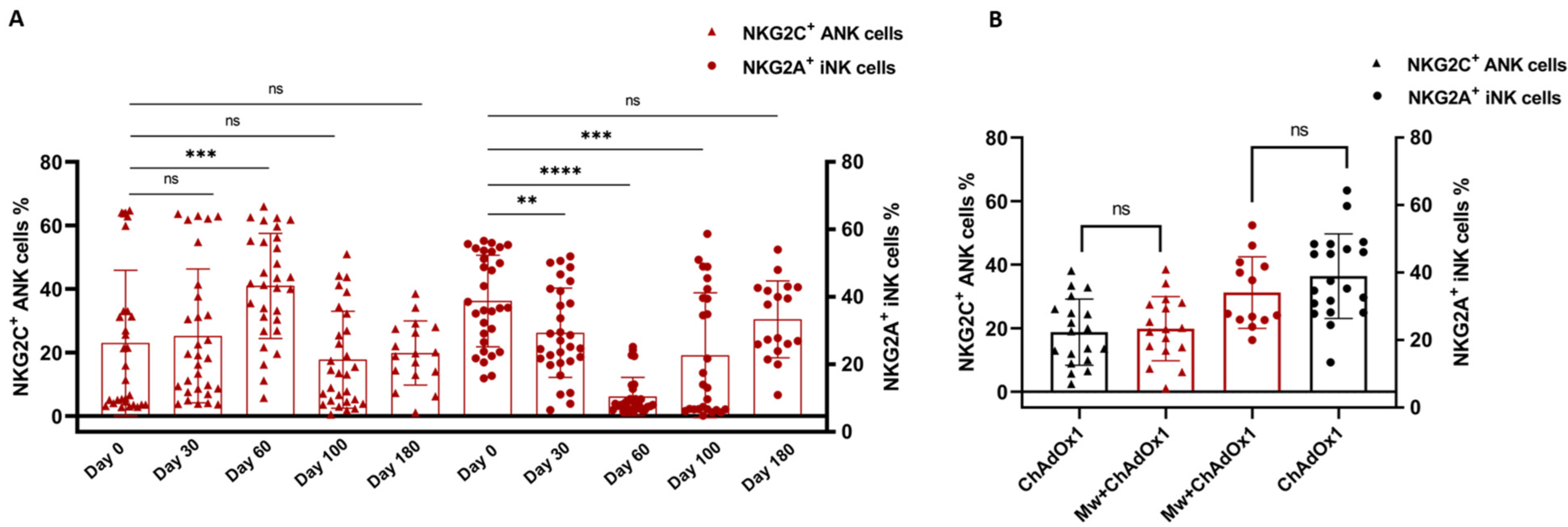
3.2. NK and T Cell Subset Analysis
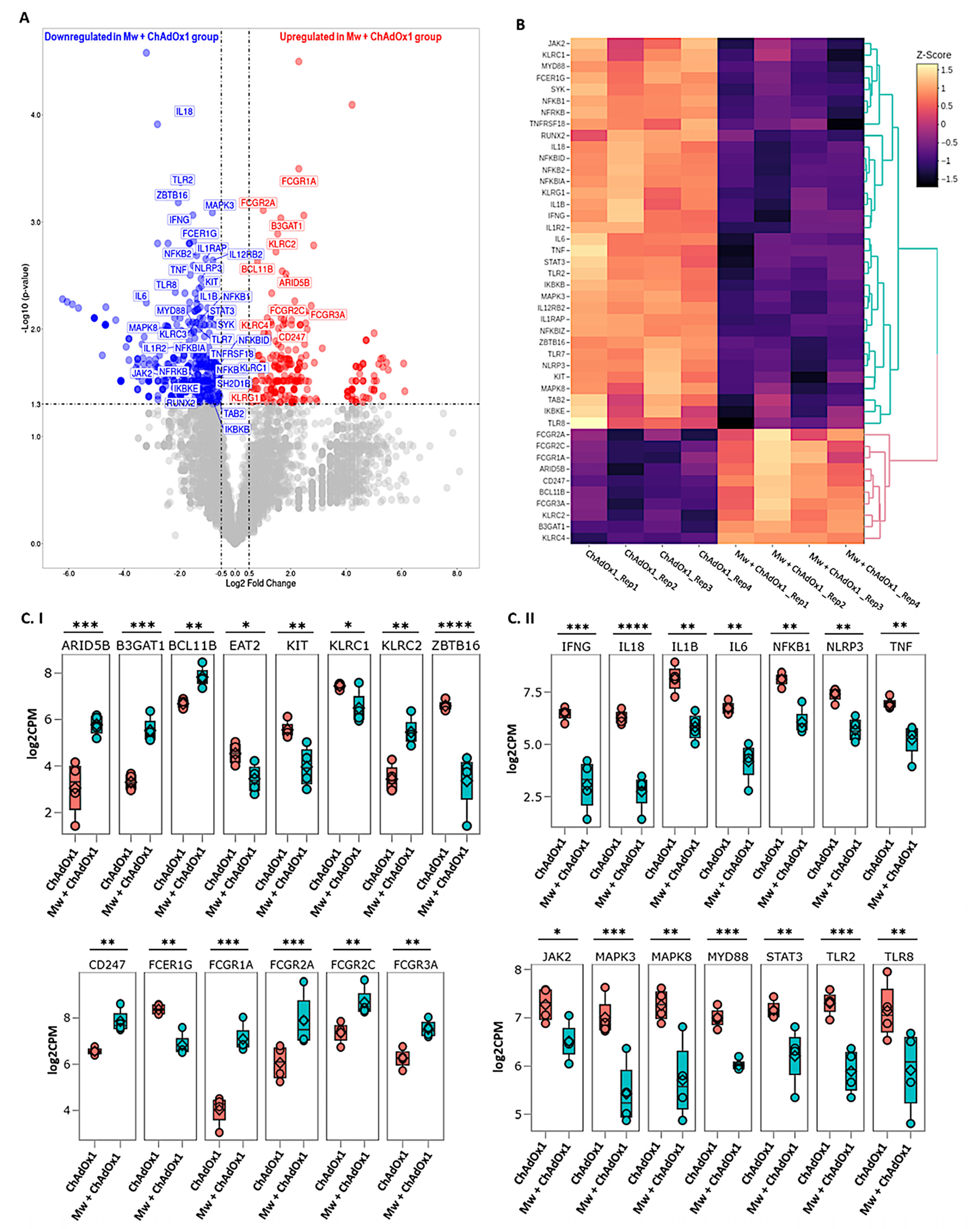
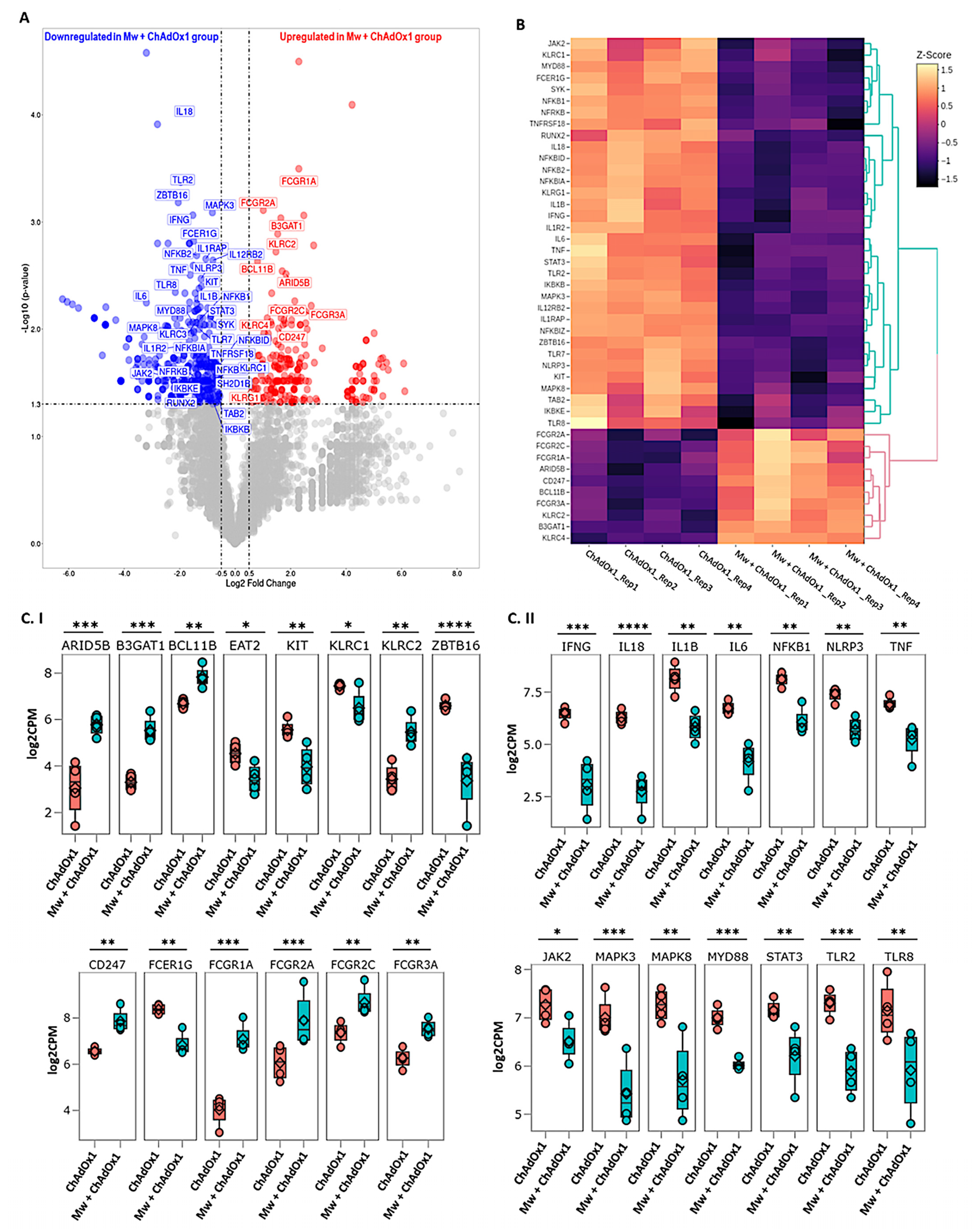
3.3. DGE Analysis
3.4. ANK-ADCC Pathway Genes
3.5. Innate Immune Inflammatory Pathway Genes
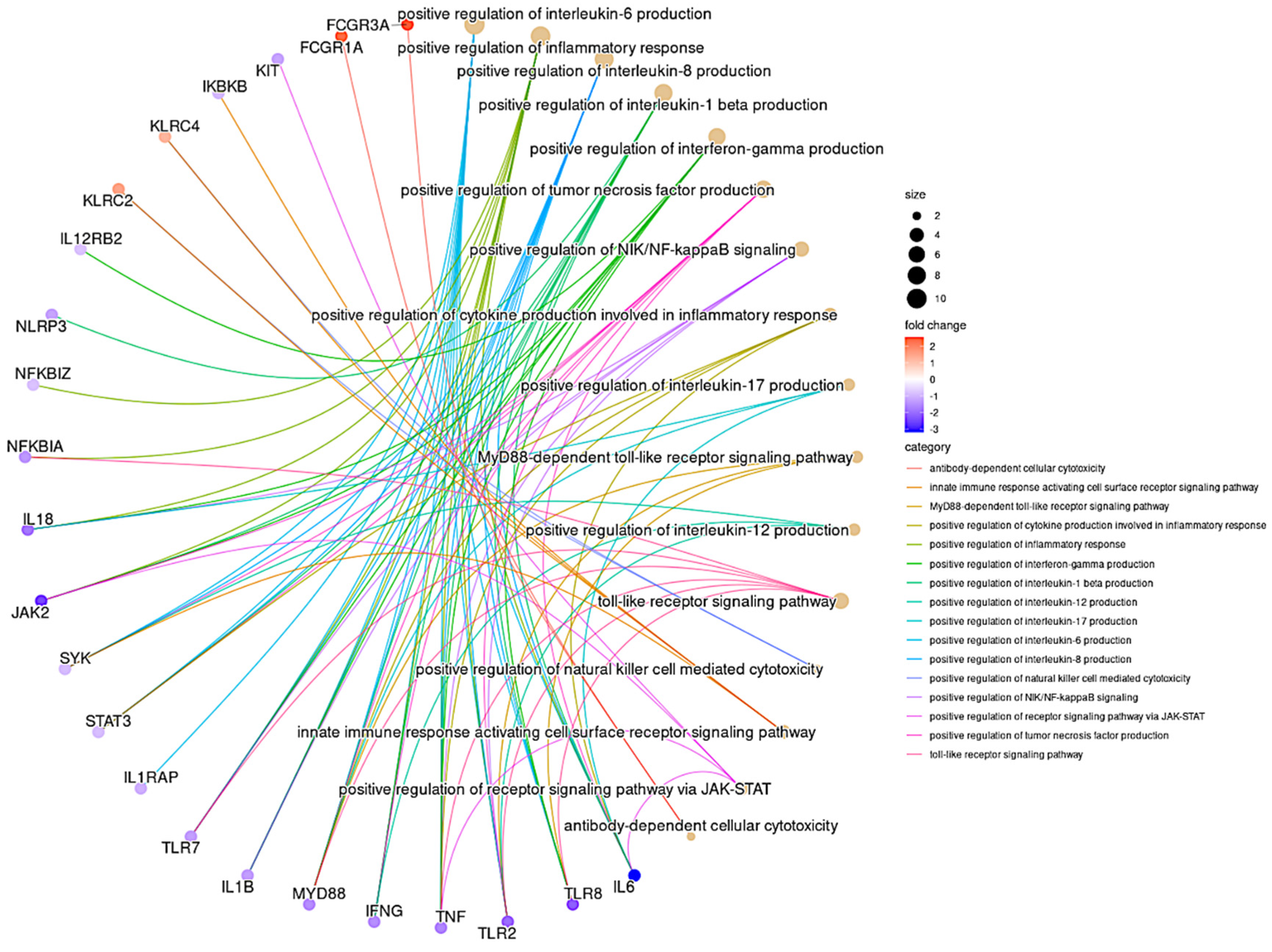
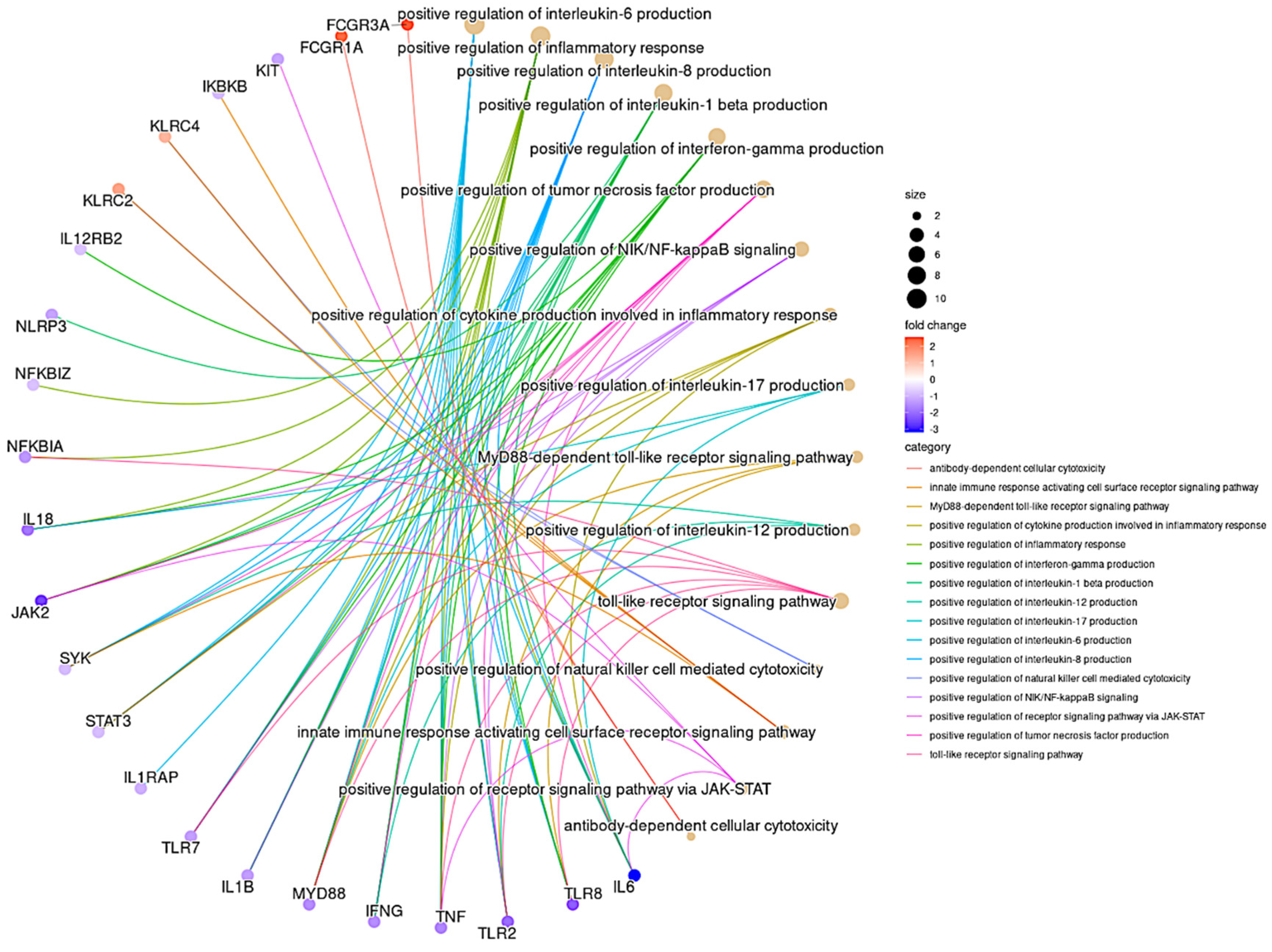
3.6. GO Pathway Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barouch, D.H. COVID-19 Vaccines—Immunity, Variants, Boosters. N. Engl. J. Med. 2022, 387, 1011–1020. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827.e819. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Iketani, S.; Guo, Y.; Chan, J.F.; Wang, M.; Liu, L.; Luo, Y.; Chu, H.; Huang, Y.; Nair, M.S.; et al. Striking antibody evasion manifested by the Omicron variant of SARS-CoV-2. Nature 2022, 602, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Goldberg, Y.; Milo, R. Protection by a Fourth Dose of BNT162b2 against Omicron in Israel. Reply. N. Engl. J. Med. 2022, 386, 2441–2442. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, Y.; Mandel, M.; Bar-On, Y.M.; Bodenheimer, O.; Freedman, L.S.; Ash, N.; Alroy-Preis, S.; Huppert, A.; Milo, R. Protection and Waning of Natural and Hybrid Immunity to SARS-CoV-2. N. Engl. J. Med. 2022, 386, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Munro, A.P.S.; Feng, S.; Janani, L.; Aley, P.K.; Babbage, G.; Baxter, D.; Bula, M.; Cathie, K.; Chatterjee, K.; et al. Persistence of immunogenicity after seven COVID-19 vaccines given as third dose boosters following two doses of ChAdOx1 nCov-19 or BNT162b2 in the UK: Three month analyses of the COV-BOOST trial. J. Infect. 2022, 84, 795–813. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.S.; Collier, A.Y.; Yu, J.; Liu, J.; Chandrashekar, A.; McMahan, K.; Jacob-Dolan, C.; He, X.; Roy, V.; Hauser, B.M.; et al. Durability of Heterologous and Homologous COVID-19 Vaccine Boosts. JAMA Netw. Open 2022, 5, e2226335. [Google Scholar] [CrossRef]
- Jaiswal, S.R.; Arunachalam, J.; Saifullah, A.; Lakhchaura, R.; Tailor, D.; Mehta, A.; Bhagawati, G.; Aiyer, H.; Khamar, B.; Malhotra, S.V.; et al. Impact of an Immune Modulator Mycobacterium-w on Adaptive Natural Killer Cells and Protection Against COVID-19. Front. Immunol. 2022, 13, 887230. [Google Scholar] [CrossRef]
- Kumar, P.; Tyagi, R.; Das, G.; Bhaskar, S. Mycobacterium indicus pranii and Mycobacterium bovis BCG lead to differential macrophage activation in Toll-like receptor-dependent manner. Immunology 2014, 143, 258–268. [Google Scholar] [CrossRef]
- Kumar, P.; Das, G.; Bhaskar, S. Mycobacterium indicus pranii therapy induces tumor regression in MyD88- and TLR2-dependent manner. BMC Res. Notes 2019, 12, 648. [Google Scholar] [CrossRef]
- Sharma, P.; Misra, R.S.; Kar, H.K.; Mukherjee, A.; Poricha, D.; Kaur, H.; Mukherjee, R.; Rani, R. Mycobacterium w vaccine, a useful adjuvant to multidrug therapy in multibacillary leprosy: A report on hospital based immunotherapeutic clinical trials with a follow-up of 1–7 years after treatment. Lepr. Rev. 2000, 71, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Trapathi, S.B. Improved cure rates in pulmonary tuberculosis category II (retreatment) with mycobacterium w. J. Indian Med. Assoc. 2003, 101, 680–682. [Google Scholar] [PubMed]
- Kumar, P.; Dar, L.; Saldiwal, S.; Varma, S.; Datt Upadhyay, A.; Talwar, D.; Sharma, V.K.; Verma, K.K.; Dwivedi, S.N.; Raj, R.; et al. Intralesional injection of Mycobacterium w vaccine vs imiquimod, 5%, cream in patients with anogenital warts: A randomized clinical trial. JAMA Dermatol. 2014, 150, 1072–1078. [Google Scholar] [CrossRef]
- Thiruvengadam, R.; Awasthi, A.; Medigeshi, G.; Bhattacharya, S.; Mani, S.; Sivasubbu, S.; Shrivastava, T.; Samal, S.; Rathna Murugesan, D.; Koundinya Desiraju, B.; et al. Effectiveness of ChAdOx1 nCoV-19 vaccine against SARS-CoV-2 infection during the delta (B.1.617.2) variant surge in India: A test-negative, case-control study and a mechanistic study of post-vaccination immune responses. Lancet Infect. Dis. 2022, 22, 473–482. [Google Scholar] [CrossRef]
- Jaiswal, S.R.; Arunachalam, J.; Bhardwaj, A.; Saifullah, A.; Lakhchaura, R.; Soni, M.; Bhagawati, G.; Chakrabarti, S. Impact of adaptive natural killer cells, KLRC2 genotype and cytomegalovirus reactivation on late mortality in patients with severe COVID-19 lung disease. Clin. Transl. Immunol. 2022, 11, e1359. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72314 Cases From the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Jaiswal, S.R.; Chakraborty, S.; Lakhchaura, R.; Shashi, P.; Mehta, A.; Soni, M.; Chakrabarti, S. Early and Sustained Expansion of Adaptive Natural Killer Cells Following Haploidentical Transplantation and CTLA4Ig-Primed Donor Lymphocyte Infusions Dissociate Graft-versus-Leukemia and Graft-versus-Host Effects. Transplant. Cell. Ther. 2021, 27, 144–151. [Google Scholar] [CrossRef]
- Yang, W.; Shaman, J. COVID-19 pandemic dynamics in India, the SARS-CoV-2 Delta variant and implications for vaccination. J. R. Soc. Interface 2022, 19, 20210900. [Google Scholar] [CrossRef]
- Salvatore, M.; Purkayastha, S.; Ganapathi, L.; Bhattacharyya, R.; Kundu, R.; Zimmermann, L.; Ray, D.; Hazra, A.; Kleinsasser, M.; Solomon, S.; et al. Lessons from SARS-CoV-2 in India: A data-driven framework for pandemic resilience. Sci. Adv. 2022, 8, eabp8621. [Google Scholar] [CrossRef]
- Krishna, B.; Gupta, A.; Meena, K.; Gaba, A.; Krishna, S.; Jyoti, R.; Aeron, N.; Prashanth, S.; Samriti; Ganapathy, U. Prevalence, severity, and risk factor of breakthrough infection after vaccination with either the Covaxin or the Covishield among healthcare workers: A nationwide cross-sectional study. J. Anaesthesiol. Clin. Pharmacol. 2022, 38, S66–S78. [Google Scholar] [CrossRef]
- Grunst, M.W.; Uchil, P.D. Fc effector cross-reactivity: A hidden arsenal against SARS-CoV-2’s evasive maneuvering. Cell Rep. Med. 2022, 3, 100540. [Google Scholar] [CrossRef]
- Richardson, S.I.; Manamela, N.P.; Motsoeneng, B.M.; Kaldine, H.; Ayres, F.; Makhado, Z.; Mennen, M.; Skelem, S.; Williams, N.; Sullivan, N.J.; et al. SARS-CoV-2 Beta and Delta variants trigger Fc effector function with increased cross-reactivity. Cell Rep. Med. 2022, 3, 100510. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, K.; Riecken, K.; Jung, J.M.; Hildebrandt, H.; Menzel, S.; Bunders, M.J.; Fehse, B.; Koch-Nolte, F.; Heinrich, F.; Peine, S.; et al. Natural killer cell-mediated ADCC in SARS-CoV-2-infected individuals and vaccine recipients. Eur. J. Immunol. 2022, 52, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.; Nayrac, M.; Benlarbi, M.; Gong, S.Y.; Gasser, R.; Beaudoin-Bussieres, G.; Brassard, N.; Laumaea, A.; Vezina, D.; Prevost, J.; et al. A single dose of the SARS-CoV-2 vaccine BNT162b2 elicits Fc-mediated antibody effector functions and T cell responses. Cell Host Microbe 2021, 29, 1137–1150.e1136. [Google Scholar] [CrossRef] [PubMed]
- Rieke, G.J.; van Bremen, K.; Bischoff, J.; ToVinh, M.; Monin, M.B.; Schlabe, S.; Raabe, J.; Kaiser, K.M.; Finnemann, C.; Odainic, A.; et al. Natural Killer Cell-Mediated Antibody-Dependent Cellular Cytotoxicity Against SARS-CoV-2 After Natural Infection Is More Potent Than After Vaccination. J. Infect. Dis. 2022, 225, 1688–1693. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, M.; Zhang, X.; Li, S.; Lu, Q.; Zeng, H.; Hou, H.; Li, H.; Zhang, M.; Jiang, F.; et al. Antibody-dependent cellular cytotoxicity response to SARS-CoV-2 in COVID-19 patients. Signal Transduct. Target. Ther. 2021, 6, 346. [Google Scholar] [CrossRef]
- Maucourant, C.; Filipovic, I.; Ponzetta, A.; Aleman, S.; Cornillet, M.; Hertwig, L.; Strunz, B.; Lentini, A.; Reinius, B.; Brownlie, D.; et al. Natural killer cell immunotypes related to COVID-19 disease severity. Sci. Immunol. 2020, 5, eabd6832. [Google Scholar] [CrossRef]
- Hwang, I.; Zhang, T.; Scott, J.M.; Kim, A.R.; Lee, T.; Kakarla, T.; Kim, A.; Sunwoo, J.B.; Kim, S. Identification of human NK cells that are deficient for signaling adaptor FcRgamma and specialized for antibody-dependent immune functions. Int. Immunol. 2012, 24, 793–802. [Google Scholar] [CrossRef]
- Liu, W.; Scott, J.M.; Langguth, E.; Chang, H.; Park, P.H.; Kim, S. FcRgamma Gene Editing Reprograms Conventional NK Cells to Display Key Features of Adaptive Human NK Cells. iScience 2020, 23, 101709. [Google Scholar] [CrossRef]
- Sariol, A.; Perlman, S. SARS-CoV-2 takes its Toll. Nat. Immunol. 2021, 22, 801–802. [Google Scholar] [CrossRef]
- Zheng, M.; Karki, R.; Williams, E.P.; Yang, D.; Fitzpatrick, E.; Vogel, P.; Jonsson, C.B.; Kanneganti, T.D. TLR2 senses the SARS-CoV-2 envelope protein to produce inflammatory cytokines. Nat. Immunol. 2021, 22, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Freeman, T.L.; Swartz, T.H. Targeting the NLRP3 Inflammasome in Severe COVID-19. Front. Immunol. 2020, 11, 1518. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ChAdOx1 Group (N = 201) | Mw + ChAdOx1 Group (N = 50) | p-Value | |
|---|---|---|---|
| Age at vaccination, median (range), years | 29 (21–53) | 28 (22–56) | 0.46 |
| Gender | 0.43 | ||
| Male | 103 (51.24%) | 28 (56%) | |
| Female | 98 (48.75%) | 22 (44%) | |
| ChAdOx1 nCoV-19 vaccine both doses | 201 (100%) | 24 (48%) | 0.0001 |
| SARS-CoV-2 infection | 36 (17.9%) | 2 (4%) | 0.01 |
| Mild | 27 (13.4%) | 2 (4%) | |
| Moderate | 9 (4.4%) | 0 | |
| Severe | 0 | 0 | |
| Reinfection | 13 | 1 | 0.3 |
| Sub-Group | SARS-CoV-2 Infection | No SARS-CoV-2 Infection | p-Value |
|---|---|---|---|
| Mw + ChAdOx1 both doses | 2 | 22 | 0.22 |
| Mw + ChAdOx1 single dose | 0 | 26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaiswal, S.R.; Saifullah, A.; Arunachalam, J.; Lakhchaura, R.; Tailor, D.; Mehta, A.; Bhagawati, G.; Aiyer, H.; Biswas, S.; Khamar, B.; et al. Augmenting Vaccine Efficacy against Delta Variant with ‘Mycobacterium-w’-Mediated Modulation of NK-ADCC and TLR-MYD88 Pathways. Vaccines 2023, 11, 328. https://doi.org/10.3390/vaccines11020328
Jaiswal SR, Saifullah A, Arunachalam J, Lakhchaura R, Tailor D, Mehta A, Bhagawati G, Aiyer H, Biswas S, Khamar B, et al. Augmenting Vaccine Efficacy against Delta Variant with ‘Mycobacterium-w’-Mediated Modulation of NK-ADCC and TLR-MYD88 Pathways. Vaccines. 2023; 11(2):328. https://doi.org/10.3390/vaccines11020328
Chicago/Turabian StyleJaiswal, Sarita Rani, Ashraf Saifullah, Jaganath Arunachalam, Rohit Lakhchaura, Dhanir Tailor, Anupama Mehta, Gitali Bhagawati, Hemamalini Aiyer, Subhrajit Biswas, Bakulesh Khamar, and et al. 2023. "Augmenting Vaccine Efficacy against Delta Variant with ‘Mycobacterium-w’-Mediated Modulation of NK-ADCC and TLR-MYD88 Pathways" Vaccines 11, no. 2: 328. https://doi.org/10.3390/vaccines11020328
APA StyleJaiswal, S. R., Saifullah, A., Arunachalam, J., Lakhchaura, R., Tailor, D., Mehta, A., Bhagawati, G., Aiyer, H., Biswas, S., Khamar, B., Malhotra, S. V., & Chakrabarti, S. (2023). Augmenting Vaccine Efficacy against Delta Variant with ‘Mycobacterium-w’-Mediated Modulation of NK-ADCC and TLR-MYD88 Pathways. Vaccines, 11(2), 328. https://doi.org/10.3390/vaccines11020328