


Carriers and Antigens: New Developments in Glycoconjugate Vaccines



Abstract
1. Introduction
2. Conjugate Vaccine Carriers
2.1. Traditional Protein Carriers
2.2. Other Protein Carriers
2.3. Recent Developments Regarding Protein Carriers
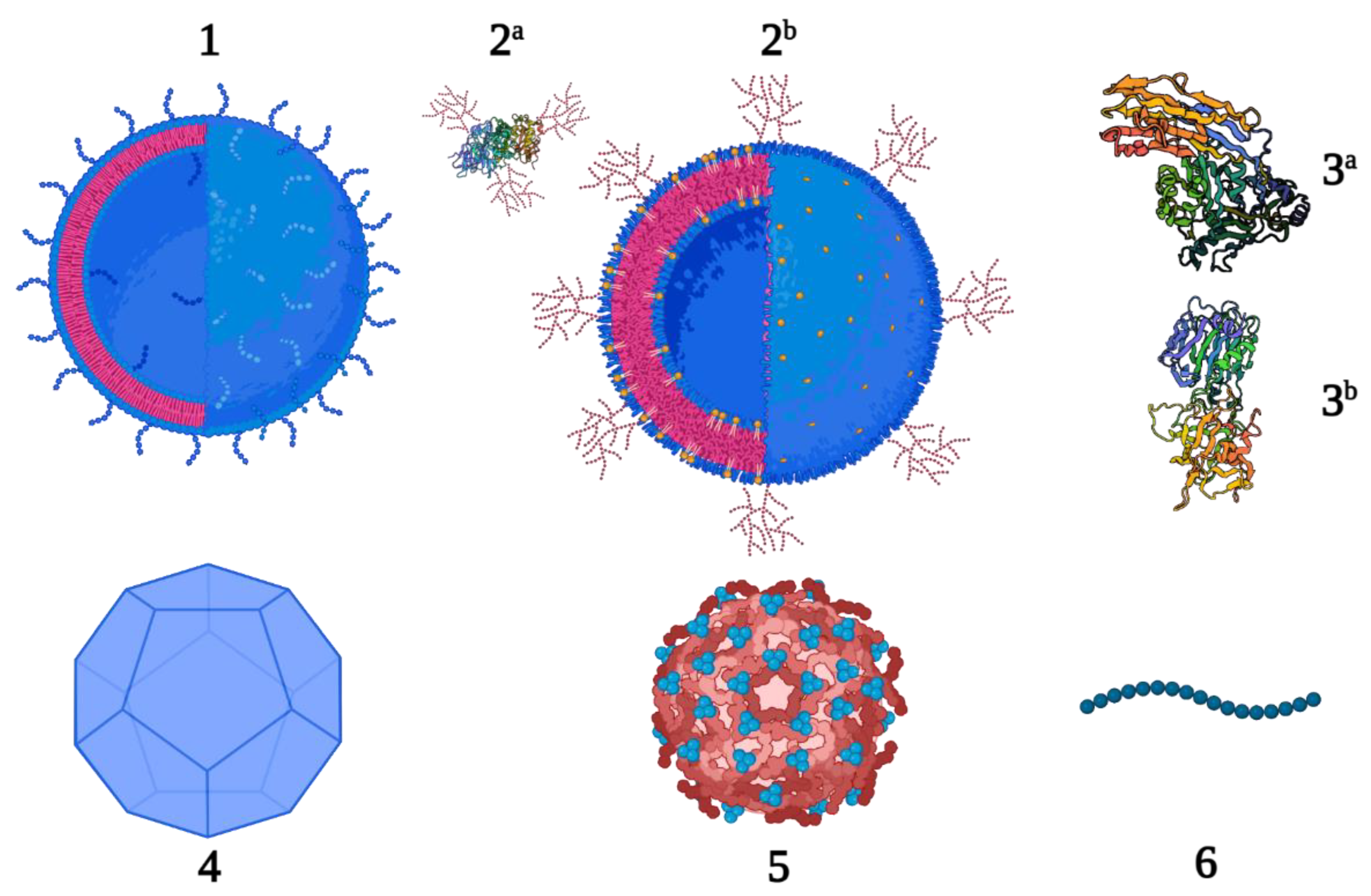
2.3.1. Outer Membrane Vesicles/Generalized Modules for Membrane Antigens
2.3.2. Glycoengineered OMVs and Proteins
2.3.3. Protein-Based Carriers
2.3.4. Virus-Like Particles
2.3.5. Protein Nanocages
2.3.6. Peptides
3. Conjugate Vaccine Antigens
3.1. Enterococcus faecium
3.2. Staphylococcus aureus
3.3. Klebsiella pneumoniae
3.4. Acinetobacter baumannii
3.5. Pseudomonas aeruginosa
3.6. Escherichia coli
4. Discussion
5. Conclusions
6. Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. History of Smallpox. Available online: https://www.cdc.gov/smallpox/history/history.html (accessed on 29 September 2022).
- Avery, O.T.; Goebel, W.F. Chemo-Immunological Studies on Conjugated Carbohydrate-Proteins: Ii. Immunological Specificity of Synthetic Sugar-Protein Antigens. J. Exp. Med. 1929, 50, 533–550. [Google Scholar] [CrossRef] [PubMed]
- Schneerson, R.; Robbins, J.B.; Parke, J.C., Jr.; Bell, C.; Schlesselman, J.J.; Sutton, A.; Wang, Z.; Schiffman, G.; Karpas, A.; Shiloach, J. Quantitative and qualitative analyses of serum antibodies elicited in adults by Haemophilus influenzae type b and pneumococcus type 6A capsular polysaccharide-tetanus toxoid conjugates. Infect. Immun. 1986, 52, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Schneerson, R.; Robbins, J.B.; Barrera, O.; Sutton, A.; Habig, W.B.; Hardegree, M.C.; Chaimovich, J. Haemophilus influenzae type B polysaccharide-protein conjugates: Model for a new generation of capsular polysaccharide vaccines. Prog. Clin. Biol. Res. 1980, 47, 77–94. [Google Scholar] [PubMed]
- Pollard, A.J.; Perrett, K.P.; Beverley, P.C. Maintaining protection against invasive bacteria with protein-polysaccharide conjugate vaccines. Nat. Rev. Immunol. 2009, 9, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.F.; Pollard, A.J.; Moxon, E.R. Immunological memory: The role of B cells in long-term protection against invasive bacterial pathogens. JAMA 2005, 294, 3019–3023. [Google Scholar] [CrossRef] [PubMed]
- Frasch, C.E. Preparation of bacterial polysaccharide-protein conjugates: Analytical and manufacturing challenges. Vaccine 2009, 27, 6468–6470. [Google Scholar] [CrossRef]
- Knuf, M.; Kowalzik, F.; Kieninger, D. Comparative effects of carrier proteins on vaccine-induced immune response. Vaccine 2011, 29, 4881–4890. [Google Scholar] [CrossRef]
- Peltola, H.; Kayhty, H.; Sivonen, A.; Makela, H. Haemophilus influenzae type b capsular polysaccharide vaccine in children: A double-blind field study of 100,000 vaccinees 3 months to 5 years of age in Finland. Pediatrics 1977, 60, 730–737. [Google Scholar] [CrossRef]
- Koskela, M.; Leinonen, M.; Haiva, V.M.; Timonen, M.; Makela, P.H. First and second dose antibody responses to pneumococcal polysaccharide vaccine in infants. Pediatr. Infect. Dis. 1986, 5, 45–50. [Google Scholar] [CrossRef]
- Costantino, P.; Rappuoli, R.; Berti, F. The design of semi-synthetic and synthetic glycoconjugate vaccines. Expert Opin. Drug Discov. 2011, 6, 1045–1066. [Google Scholar] [CrossRef]
- Pichichero, M.E. Protein carriers of conjugate vaccines: Characteristics, development, and clinical trials. Hum. Vaccin. Immunother. 2013, 9, 2505–2523. [Google Scholar] [CrossRef] [PubMed]
- Broker, M.; Berti, F.; Schneider, J.; Vojtek, I. Polysaccharide conjugate vaccine protein carriers as a "neglected valency" - Potential and limitations. Vaccine 2017, 35, 3286–3294. [Google Scholar] [CrossRef] [PubMed]
- Schutze, M.P.; Leclerc, C.; Vogel, F.R.; Chedid, L. Epitopic suppression in synthetic vaccine models: Analysis of the effector mechanisms. Cell. Immunol. 1987, 104, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Herzenberg, L.A.; Tokuhisa, T.; Herzenberg, L.A. Carrier-priming leads to hapten-specific suppression. Nature 1980, 285, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Barington, T.; Kristensen, K.; Henrichsen, J.; Heilmann, C. Influence of prevaccination immunity on the human B-lymphocyte response to a Haemophilus influenzae type b conjugate vaccine. Infect. Immun. 1991, 59, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Dagan, R.; Eskola, J.; Leclerc, C.; Leroy, O. Reduced response to multiple vaccines sharing common protein epitopes that are administered simultaneously to infants. Infect Immun 1998, 66, 2093–2098. [Google Scholar] [CrossRef]
- Dagan, R.; Poolman, J.; Siegrist, C.A. Glycoconjugate vaccines and immune interference: A review. Vaccine 2010, 28, 5513–5523. [Google Scholar] [CrossRef]
- Giannini, G.; Rappuoli, R.; Ratti, G. The amino-acid sequence of two non-toxic mutants of diphtheria toxin: CRM45 and CRM197. Nucleic Acids Res. 1984, 12, 4063–4069. [Google Scholar] [CrossRef]
- Broker, M.; Costantino, P.; DeTora, L.; McIntosh, E.D.; Rappuoli, R. Biochemical and biological characteristics of cross-reacting material 197 CRM197, a non-toxic mutant of diphtheria toxin: Use as a conjugation protein in vaccines and other potential clinical applications. Biologicals 2011, 39, 195–204. [Google Scholar] [CrossRef]
- Shinefield, H.R. Overview of the development and current use of CRM(197) conjugate vaccines for pediatric use. Vaccine 2010, 28, 4335–4339. [Google Scholar] [CrossRef]
- Forsgren, A.; Riesbeck, K.; Janson, H. Protein D of Haemophilus influenzae: A protective nontypeable H. influenzae antigen and a carrier for pneumococcal conjugate vaccines. Clin. Infect. Dis. 2008, 46, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Prymula, R.; Peeters, P.; Chrobok, V.; Kriz, P.; Novakova, E.; Kaliskova, E.; Kohl, I.; Lommel, P.; Poolman, J.; Prieels, J.P.; et al. Pneumococcal capsular polysaccharides conjugated to protein D for prevention of acute otitis media caused by both Streptococcus pneumoniae and non-typable Haemophilus influenzae: A randomised double-blind efficacy study. Lancet 2006, 367, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, J.J.; Deck, R.R.; Liu, M.A. Immunogenicity of a Haemophilus influenzae polysaccharide-Neisseria meningitidis outer membrane protein complex conjugate vaccine. J. Immunol. 1990, 145, 3071–3079. [Google Scholar] [CrossRef] [PubMed]
- Kilpi, T.; Ahman, H.; Jokinen, J.; Lankinen, K.S.; Palmu, A.; Savolainen, H.; Gronholm, M.; Leinonen, M.; Hovi, T.; Eskola, J.; et al. Protective efficacy of a second pneumococcal conjugate vaccine against pneumococcal acute otitis media in infants and children: Randomized, controlled trial of a 7-valent pneumococcal polysaccharide-meningococcal outer membrane protein complex conjugate vaccine in 1666 children. Clin. Infect. Dis. 2003, 37, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Adamo, R.; Costantino, P. Protein Carriers for Glycoconjugate Vaccines: History, Selection Criteria, Characterization and New Trends. Molecules 2018, 23, 1451. [Google Scholar] [CrossRef] [PubMed]
- Cryz, S.J., Jr.; Cross, A.S.; Sadoff, J.C.; Wegmann, A.; Que, J.U.; Furer, E. Safety and immunogenicity of Escherichia coli O18 O-specific polysaccharide (O-PS)-toxin A and O-PS-cholera toxin conjugate vaccines in humans. J. Infect. Dis. 1991, 163, 1040–1045. [Google Scholar] [CrossRef]
- Cross, A.; Artenstein, A.; Que, J.; Fredeking, T.; Furer, E.; Sadoff, J.C.; Cryz, S.J., Jr. Safety and immunogenicity of a polyvalent Escherichia coli vaccine in human volunteers. J. Infect. Dis. 1994, 170, 834–840. [Google Scholar] [CrossRef]
- Kossaczka, Z.; Lin, F.Y.; Ho, V.A.; Thuy, N.T.; Van Bay, P.; Thanh, T.C.; Khiem, H.B.; Trach, D.D.; Karpas, A.; Hunt, S.; et al. Safety and immunogenicity of Vi conjugate vaccines for typhoid fever in adults, teenagers, and 2- to 4-year-old children in Vietnam. Infect. Immun. 1999, 67, 5806–5810. [Google Scholar] [CrossRef]
- Fattom, A.; Schneerson, R.; Watson, D.C.; Karakawa, W.W.; Fitzgerald, D.; Pastan, I.; Li, X.; Shiloach, J.; Bryla, D.A.; Robbins, J.B. Laboratory and clinical evaluation of conjugate vaccines composed of Staphylococcus aureus type 5 and type 8 capsular polysaccharides bound to Pseudomonas aeruginosa recombinant exoprotein A. Infect. Immun. 1993, 61, 1023–1032. [Google Scholar] [CrossRef]
- Cohen, D.; Ashkenazi, S.; Green, M.S.; Gdalevich, M.; Robin, G.; Slepon, R.; Yavzori, M.; Orr, N.; Block, C.; Ashkenazi, I.; et al. Double-blind vaccine-controlled randomised efficacy trial of an investigational Shigella sonnei conjugate vaccine in young adults. Lancet 1997, 349, 155–159. [Google Scholar] [CrossRef]
- Szu, S.C.; Stone, A.L.; Robbins, J.D.; Schneerson, R.; Robbins, J.B. Vi capsular polysaccharide-protein conjugates for prevention of typhoid fever. Preparation, characterization, and immunogenicity in laboratory animals. J. Exp. Med. 1987, 166, 1510–1524. [Google Scholar] [CrossRef] [PubMed]
- Tontini, M.; Romano, M.R.; Proietti, D.; Balducci, E.; Micoli, F.; Balocchi, C.; Santini, L.; Masignani, V.; Berti, F.; Costantino, P. Preclinical studies on new proteins as carrier for glycoconjugate vaccines. Vaccine 2016, 34, 4235–4242. [Google Scholar] [CrossRef] [PubMed]
- Falugi, F.; Petracca, R.; Mariani, M.; Luzzi, E.; Mancianti, S.; Carinci, V.; Melli, M.L.; Finco, O.; Wack, A.; Di Tommaso, A.; et al. Rationally designed strings of promiscuous CD4(+) T cell epitopes provide help to Haemophilus influenzae type b oligosaccharide: A model for new conjugate vaccines. Eur. J. Immunol. 2001, 31, 3816–3824. [Google Scholar] [CrossRef] [PubMed]
- Baraldo, K.; Mori, E.; Bartoloni, A.; Norelli, F.; Grandi, G.; Rappuoli, R.; Finco, O.; Del Giudice, G. Combined conjugate vaccines: Enhanced immunogenicity with the N19 polyepitope as a carrier protein. Infect. Immun. 2005, 73, 5835–5841. [Google Scholar] [CrossRef]
- Bongat, A.F.; Saksena, R.; Adamo, R.; Fujimoto, Y.; Shiokawa, Z.; Peterson, D.C.; Fukase, K.; Vann, W.F.; Kovac, P. Multimeric bivalent immunogens from recombinant tetanus toxin HC fragment, synthetic hexasaccharides, and a glycopeptide adjuvant. Glycoconj. J. 2010, 27, 69–77. [Google Scholar] [CrossRef]
- Xin, H.; Dziadek, S.; Bundle, D.R.; Cutler, J.E. Synthetic glycopeptide vaccines combining beta-mannan and peptide epitopes induce protection against candidiasis. Proc. Natl. Acad. Sci. USA 2008, 105, 13526–13531. [Google Scholar] [CrossRef]
- Rothbard, J.B.; Taylor, W.R. A sequence pattern common to T cell epitopes. EMBO J. 1988, 7, 93–100. [Google Scholar] [CrossRef]
- Ingale, S.; Wolfert, M.A.; Gaekwad, J.; Buskas, T.; Boons, G.J. Robust immune responses elicited by a fully synthetic three-component vaccine. Nat. Chem. Biol. 2007, 3, 663–667. [Google Scholar] [CrossRef]
- Bixler, G.S., Jr.; Eby, R.; Dermody, K.M.; Woods, R.M.; Seid, R.C.; Pillai, S. Synthetic peptide representing a T-cell epitope of CRM197 substitutes as carrier molecule in a Haemophilus influenzae type B (Hib) conjugate vaccine. Adv. Exp. Med. Biol. 1989, 251, 175–180. [Google Scholar] [CrossRef]
- Belot, F.; Guerreiro, C.; Baleux, F.; Mulard, L.A. Synthesis of two linear PADRE conjugates bearing a deca- or pentadecasaccharide B epitope as potential synthetic vaccines against Shigella flexneri serotype 2a infection. Chemistry 2005, 11, 1625–1635. [Google Scholar] [CrossRef]
- Alexander, J.; del Guercio, M.F.; Maewal, A.; Qiao, L.; Fikes, J.; Chesnut, R.W.; Paulson, J.; Bundle, D.R.; DeFrees, S.; Sette, A. Linear PADRE T helper epitope and carbohydrate B cell epitope conjugates induce specific high titer IgG antibody responses. J. Immunol. 2000, 164, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; del Guercio, M.F.; Frame, B.; Maewal, A.; Sette, A.; Nahm, M.H.; Newman, M.J. Development of experimental carbohydrate-conjugate vaccines composed of Streptococcus pneumoniae capsular polysaccharides and the universal helper T-lymphocyte epitope (PADRE). Vaccine 2004, 22, 2362–2367. [Google Scholar] [CrossRef] [PubMed]
- Michon, F.; Fusco, P.C.; Minetti, C.A.; Laude-Sharp, M.; Uitz, C.; Huang, C.H.; D’Ambra, A.J.; Moore, S.; Remeta, D.P.; Heron, I.; et al. Multivalent pneumococcal capsular polysaccharide conjugate vaccines employing genetically detoxified pneumolysin as a carrier protein. Vaccine 1998, 16, 1732–1741. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, C.; Wilk, K.; Lee, J.C.; Gening, M.; Nifantiev, N.; Pier, G.B. Opsonic and protective properties of antibodies raised to conjugate vaccines targeting six Staphylococcus aureus antigens. PLoS ONE 2012, 7, e46648. [Google Scholar] [CrossRef] [PubMed]
- Romano, M.R.; Leuzzi, R.; Cappelletti, E.; Tontini, M.; Nilo, A.; Proietti, D.; Berti, F.; Costantino, P.; Adamo, R.; Scarselli, M. Recombinant Clostridium difficile toxin fragments as carrier protein for PSII surface polysaccharide preserve their neutralizing activity. Toxins 2014, 6, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Nilo, A.; Morelli, L.; Passalacqua, I.; Brogioni, B.; Allan, M.; Carboni, F.; Pezzicoli, A.; Zerbini, F.; Maione, D.; Fabbrini, M.; et al. Anti-Group B Streptococcus Glycan-Conjugate Vaccines Using Pilus Protein GBS80 As Carrier and Antigen: Comparing Lysine and Tyrosine-directed Conjugation. ACS Chem. Biol. 2015, 10, 1737–1746. [Google Scholar] [CrossRef]
- Nilo, A.; Passalacqua, I.; Fabbrini, M.; Allan, M.; Usera, A.; Carboni, F.; Brogioni, B.; Pezzicoli, A.; Cobb, J.; Romano, M.R.; et al. Exploring the Effect of Conjugation Site and Chemistry on the Immunogenicity of an anti-Group B Streptococcus Glycoconjugate Vaccine Based on GBS67 Pilus Protein and Type V Polysaccharide. Bioconjug. Chem. 2015, 26, 1839–1849. [Google Scholar] [CrossRef]
- Simon, R.; Tennant, S.M.; Wang, J.Y.; Schmidlein, P.J.; Lees, A.; Ernst, R.K.; Pasetti, M.F.; Galen, J.E.; Levine, M.M. Salmonella enterica serovar enteritidis core O polysaccharide conjugated to H:g,m flagellin as a candidate vaccine for protection against invasive infection with S. enteritidis. Infect. Immun. 2011, 79, 4240–4249. [Google Scholar] [CrossRef]
- Wang, L.X. Synthetic carbohydrate antigens for HIV vaccine design. Curr. Opin. Chem. Biol. 2013, 17, 997–1005. [Google Scholar] [CrossRef]
- Polonskaya, Z.; Deng, S.; Sarkar, A.; Kain, L.; Comellas-Aragones, M.; McKay, C.S.; Kaczanowska, K.; Holt, M.; McBride, R.; Palomo, V.; et al. T cells control the generation of nanomolar-affinity anti-glycan antibodies. J. Clin. Investig. 2017, 127, 1491–1504. [Google Scholar] [CrossRef]
- Valguarnera, E.; Feldman, M.F. Glycoengineered Outer Membrane Vesicles as a Platform for Vaccine Development. Methods Enzymol. 2017, 597, 285–310. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Alfini, R.; Di Benedetto, R.; Necchi, F.; Schiavo, F.; Mancini, F.; Carducci, M.; Palmieri, E.; Balocchi, C.; Gasperini, G.; et al. GMMA Is a Versatile Platform to Design Effective Multivalent Combination Vaccines. Vaccines 2020, 8, 540. [Google Scholar] [CrossRef] [PubMed]
- Jan, A.T. Outer Membrane Vesicles (OMVs) of Gram-negative Bacteria: A Perspective Update. Front. Microbiol. 2017, 8, 1053. [Google Scholar] [CrossRef] [PubMed]
- Launay, O.; Lewis, D.J.M.; Anemona, A.; Loulergue, P.; Leahy, J.; Scire, A.S.; Maugard, A.; Marchetti, E.; Zancan, S.; Huo, Z.; et al. Safety Profile and Immunologic Responses of a Novel Vaccine Against Shigella sonnei Administered Intramuscularly, Intradermally and Intranasally: Results From Two Parallel Randomized Phase 1 Clinical Studies in Healthy Adult Volunteers in Europe. EBioMedicine 2017, 22, 164–172. [Google Scholar] [CrossRef]
- de Kleijn, E.D.; de Groot, R.; Labadie, J.; Lafeber, A.B.; van den Dobbelsteen, G.; van Alphen, L.; van Dijken, H.; Kuipers, B.; van Omme, G.W.; Wala, M.; et al. Immunogenicity and safety of a hexavalent meningococcal outer-membrane-vesicle vaccine in children of 2-3 and 7-8 years of age. Vaccine 2000, 18, 1456–1466. [Google Scholar] [CrossRef]
- Cartwright, K.; Morris, R.; Rumke, H.; Fox, A.; Borrow, R.; Begg, N.; Richmond, P.; Poolman, J. Immunogenicity and reactogenicity in UK infants of a novel meningococcal vesicle vaccine containing multiple class 1 (PorA) outer membrane proteins. Vaccine 1999, 17, 2612–2619. [Google Scholar] [CrossRef]
- Skidmore, B.J.; Chiller, J.M.; Morrison, D.C.; Weigle, W.O. Immunologic properties of bacterial lipopolysaccharide (LPS): Correlation between the mitogenic, adjuvant, and immunogenic activities. J. Immunol. 1975, 114, 770–775. [Google Scholar] [CrossRef]
- Liu, Y.; Hammer, L.A.; Liu, W.; Hobbs, M.M.; Zielke, R.A.; Sikora, A.E.; Jerse, A.E.; Egilmez, N.K.; Russell, M.W. Experimental vaccine induces Th1-driven immune responses and resistance to Neisseria gonorrhoeae infection in a murine model. Mucosal Immunol. 2017, 10, 1594–1608. [Google Scholar] [CrossRef]
- Lehmann, A.K.; Halstensen, A.; Aaberge, I.S.; Holst, J.; Michaelsen, T.E.; Sornes, S.; Wetzler, L.M.; Guttormsen, H. Human opsonins induced during meningococcal disease recognize outer membrane proteins PorA and PorB. Infect. Immun. 1999, 67, 2552–2560. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- van der Pol, L.; Stork, M.; van der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef] [PubMed]
- Gerritzen, M.J.H.; Salverda, M.L.M.; Martens, D.E.; Wijffels, R.H.; Stork, M. Spontaneously released Neisseria meningitidis outer membrane vesicles as vaccine platform: Production and purification. Vaccine 2019, 37, 6978–6986. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Alfini, R.; Di Benedetto, R.; Necchi, F.; Schiavo, F.; Mancini, F.; Carducci, M.; Oldrini, D.; Pitirollo, O.; Gasperini, G.; et al. Generalized Modules for Membrane Antigens as Carrier for Polysaccharides: Impact of Sugar Length, Density, and Attachment Site on the Immune Response Elicited in Animal Models. Front. Immunol. 2021, 12, 719315. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, E.; Kis, Z.; Ozanne, J.; Di Benedetto, R.; Ricchetti, B.; Massai, L.; Carducci, M.; Oldrini, D.; Gasperini, G.; Aruta, M.G.; et al. GMMA as an Alternative Carrier for a Glycoconjugate Vaccine against Group A Streptococcus. Vaccines 2022, 10, 1034. [Google Scholar] [CrossRef]
- Jiang, L.; Driedonks, T.A.P.; Jong, W.S.P.; Dhakal, S.; Bart van den Berg van Saparoea, H.; Sitaras, I.; Zhou, R.; Caputo, C.; Littlefield, K.; Lowman, M.; et al. A bacterial extracellular vesicle-based intranasal vaccine against SARS-CoV-2 protects against disease and elicits neutralizing antibodies to wild-type and Delta variants. J. Extracell. Vesicles 2022, 11, e12192. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, R.; Alfini, R.; Carducci, M.; Aruta, M.G.; Lanzilao, L.; Acquaviva, A.; Palmieri, E.; Giannelli, C.; Necchi, F.; Saul, A.; et al. Novel Simple Conjugation Chemistries for Decoration of GMMA with Heterologous Antigens. Int. J. Mol. Sci. 2021, 22, 180. [Google Scholar] [CrossRef] [PubMed]
- Scaria, P.V.; Rowe, C.G.; Chen, B.B.; Muratova, O.V.; Fischer, E.R.; Barnafo, E.K.; Anderson, C.F.; Zaidi, I.U.; Lambert, L.E.; Lucas, B.J.; et al. Outer membrane protein complex as a carrier for malaria transmission blocking antigen Pfs230. NPJ Vaccines 2019, 4, 24. [Google Scholar] [CrossRef]
- Langdon, R.H.; Cuccui, J.; Wren, B.W. N-linked glycosylation in bacteria: An unexpected application. Future Microbiol. 2009, 4, 401–412. [Google Scholar] [CrossRef]
- Kay, E.; Cuccui, J.; Wren, B.W. Recent advances in the production of recombinant glycoconjugate vaccines. NPJ Vaccines 2019, 4, 16. [Google Scholar] [CrossRef]
- Wacker, M.; Linton, D.; Hitchen, P.G.; Nita-Lazar, M.; Haslam, S.M.; North, S.J.; Panico, M.; Morris, H.R.; Dell, A.; Wren, B.W.; et al. N-linked glycosylation in Campylobacter jejuni and its functional transfer into E. coli. Science 2002, 298, 1790–1793. [Google Scholar] [CrossRef]
- Nicolardi, S.; Danuser, R.; Dotz, V.; Dominguez-Vega, E.; Al Kaabi, A.; Beurret, M.; Anish, C.; Wuhrer, M. Glycan and Protein Analysis of Glycoengineered Bacterial E. coli Vaccines by MALDI-in-Source Decay FT-ICR Mass Spectrometry. Anal. Chem. 2022, 94, 4979–4987. [Google Scholar] [CrossRef] [PubMed]
- MacCalman, T.E.; Phillips-Jones, M.K.; Harding, S.E. Glycoconjugate vaccines: Some observations on carrier and production methods. Biotechnol. Genet. Eng. Rev. 2019, 35, 93–125. [Google Scholar] [CrossRef]
- Harding, C.M.; Nasr, M.A.; Scott, N.E.; Goyette-Desjardins, G.; Nothaft, H.; Mayer, A.E.; Chavez, S.M.; Huynh, J.P.; Kinsella, R.L.; Szymanski, C.M.; et al. A platform for glycoengineering a polyvalent pneumococcal bioconjugate vaccine using E. coli as a host. Nat. Commun. 2019, 10, 891. [Google Scholar] [CrossRef]
- Martin, P.; Alaimo, C. The Ongoing Journey of a Shigella Bioconjugate Vaccine. Vaccines 2022, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Ravenscroft, N.; Braun, M.; Schneider, J.; Dreyer, A.M.; Wetter, M.; Haeuptle, M.A.; Kemmler, S.; Steffen, M.; Sirena, D.; Herwig, S.; et al. Characterization and immunogenicity of a Shigella flexneri 2a O-antigen bioconjugate vaccine candidate. Glycobiology 2019, 29, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.F.; Mayer Bridwell, A.E.; Scott, N.E.; Vinogradov, E.; McKee, S.R.; Chavez, S.M.; Twentyman, J.; Stallings, C.L.; Rosen, D.A.; Harding, C.M. A promising bioconjugate vaccine against hypervirulent Klebsiella pneumoniae. Proc. Natl. Acad. Sci. USA 2019, 116, 18655–18663. [Google Scholar] [CrossRef]
- Reglinski, M.; Ercoli, G.; Plumptre, C.; Kay, E.; Petersen, F.C.; Paton, J.C.; Wren, B.W.; Brown, J.S. A recombinant conjugated pneumococcal vaccine that protects against murine infections with a similar efficacy to Prevnar-13. NPJ Vaccines 2018, 3, 53. [Google Scholar] [CrossRef]
- Stevenson, T.C.; Cywes-Bentley, C.; Moeller, T.D.; Weyant, K.B.; Putnam, D.; Chang, Y.F.; Jones, B.D.; Pier, G.B.; DeLisa, M.P. Immunization with outer membrane vesicles displaying conserved surface polysaccharide antigen elicits broadly antimicrobial antibodies. Proc. Natl. Acad. Sci. USA 2018, 115, E3106–E3115. [Google Scholar] [CrossRef]
- Sun, P.; Pan, C.; Zeng, M.; Liu, B.; Liang, H.; Wang, D.; Liu, X.; Wang, B.; Lyu, Y.; Wu, J.; et al. Design and production of conjugate vaccines against S. Paratyphi A using an O-linked glycosylation system in vivo. NPJ Vaccines 2018, 3, 4. [Google Scholar] [CrossRef]
- Marshall, L.E.; Nelson, M.; Davies, C.H.; Whelan, A.O.; Jenner, D.C.; Moule, M.G.; Denman, C.; Cuccui, J.; Atkins, T.P.; Wren, B.W.; et al. An O-Antigen Glycoconjugate Vaccine Produced Using Protein Glycan Coupling Technology Is Protective in an Inhalational Rat Model of Tularemia. J. Immunol. Res. 2018, 2018, 8087916. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Y.; Wang, G.; Feng, S.; Guo, Z.; Gu, G. Group A Streptococcus Cell Wall Oligosaccharide-Streptococcal C5a Peptidase Conjugates as Effective Antibacterial Vaccines. ACS Infect. Dis. 2020, 6, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, J.; Zhao, Y.; Wang, S.; Feng, S.; Gu, G. Immunogenicity Assessment of Different Segments and Domains of Group a Streptococcal C5a Peptidase and Their Application Potential as Carrier Protein for Glycoconjugate Vaccine Development. Vaccines 2021, 9, 139. [Google Scholar] [CrossRef]
- Kapoor, N.; Uchiyama, S.; Pill, L.; Bautista, L.; Sedra, A.; Yin, L.; Regan, M.; Chu, E.; Rabara, T.; Wong, M.; et al. Non-Native Amino Acid Click Chemistry-Based Technology for Site-Specific Polysaccharide Conjugation to a Bacterial Protein Serving as Both Carrier and Vaccine Antigen. ACS Omega 2022, 7, 24111–24120. [Google Scholar] [CrossRef] [PubMed]
- Romero-Saavedra, F.; Laverde, D.; Kalfopoulou, E.; Martini, C.; Torelli, R.; Martinez-Matamoros, D.; Sanguinetti, M.; Huebner, J. Conjugation of Different Immunogenic Enterococcal Vaccine Target Antigens Leads to Extended Strain Coverage. J. Infect. Dis. 2019, 220, 1589–1598. [Google Scholar] [CrossRef]
- Chang, M.J.; Ollivault-Shiflett, M.; Schuman, R.; Ngoc Nguyen, S.; Kaltashov, I.A.; Bobst, C.; Rajagopal, S.P.; Przedpelski, A.; Barbieri, J.T.; Lees, A. Genetically detoxified tetanus toxin as a vaccine and conjugate carrier protein. Vaccine 2022, 40, 5103–5113. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, J.; Sun, C.; Chi, H.; Luo, D.; Wang, R.; Qiu, H.; Zhang, Y.; Wu, M.; Zhang, X.; et al. Rational Development of a Polysaccharide-Protein-Conjugated Nanoparticle Vaccine Against SARS-CoV-2 Variants and Streptococcus pneumoniae. Adv. Mater. 2022, 34, e2200443. [Google Scholar] [CrossRef]
- Park, W.J.; Yoon, Y.K.; Park, J.S.; Pansuriya, R.; Seok, Y.J.; Ganapathy, R. Rotavirus spike protein DeltaVP8* as a novel carrier protein for conjugate vaccine platform with demonstrated antigenic potential for use as bivalent vaccine. Sci. Rep. 2021, 11, 22037. [Google Scholar] [CrossRef]
- Ahmadi, K.; Aslani, M.M.; Pouladfar, G.; Faezi, S.; Kalani, M.; Pourmand, M.R.; Ghaedi, T.; Havaei, S.A.; Mahdavi, M. Preparation and preclinical evaluation of two novel Staphylococcus aureus capsular polysaccharide 5 and 8-fusion protein (Hla-MntC-SACOL0723) immunoconjugates. IUBMB Life 2020, 72, 226–236. [Google Scholar] [CrossRef]
- Chiu, T.W.; Peng, C.J.; Chen, M.C.; Hsu, M.H.; Liang, Y.H.; Chiu, C.H.; Fang, J.M.; Lee, Y.C. Constructing conjugate vaccine against Salmonella Typhimurium using lipid-A free lipopolysaccharide. J. Biomed Sci. 2020, 27, 89. [Google Scholar] [CrossRef]
- Ou, L.; Kong, W.P.; Chuang, G.Y.; Ghosh, M.; Gulla, K.; O’Dell, S.; Varriale, J.; Barefoot, N.; Changela, A.; Chao, C.W.; et al. Preclinical Development of a Fusion Peptide Conjugate as an HIV Vaccine Immunogen. Sci. Rep. 2020, 10, 3032. [Google Scholar] [CrossRef]
- Qian, W.; Huang, Z.; Chen, Y.; Yang, J.; Wang, L.; Wu, K.; Chen, M.; Chen, N.; Duan, Y.; Shi, J.; et al. Elicitation of integrated immunity in mice by a novel pneumococcal polysaccharide vaccine conjugated with HBV surface antigen. Sci. Rep. 2020, 10, 6470. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.J.; Yang, Y.; Shaddeau, A.; Cai, C.X.; Li, Y.; Gulla, K.; Zhang, Y.; Ou, L.; Cooper, J.W.; Lei, Q.P. A unique algorithm for the determination of peptide-carrier protein conjugation ratio by amino acid analysis using intrinsic internal standard. Vaccine 2020, 38, 4507–4511. [Google Scholar] [CrossRef] [PubMed]
- An, S.J.; Scaria, P.V.; Chen, B.; Barnafo, E.; Muratova, O.; Anderson, C.; Lambert, L.; Chae, M.H.; Yang, J.S.; Duffy, P.E. Development of a bivalent conjugate vaccine candidate against malaria transmission and typhoid fever. Vaccine 2018, 36, 2978–2984. [Google Scholar] [CrossRef] [PubMed]
- Baliban, S.M.; Curtis, B.; Toema, D.; Tennant, S.M.; Levine, M.M.; Pasetti, M.F.; Simon, R. Immunogenicity and efficacy following sequential parenterally-administered doses of Salmonella Enteritidis COPS:FliC glycoconjugates in infant and adult mice. PLoS Negl. Trop Dis. 2018, 12, e0006522. [Google Scholar] [CrossRef]
- Baruffaldi, F.; Kelcher, A.H.; Laudenbach, M.; Gradinati, V.; Limkar, A.; Roslawski, M.; Birnbaum, A.; Lees, A.; Hassler, C.; Runyon, S.; et al. Preclinical Efficacy and Characterization of Candidate Vaccines for Treatment of Opioid Use Disorders Using Clinically Viable Carrier Proteins. Mol. Pharm. 2018, 15, 4947–4962. [Google Scholar] [CrossRef]
- Laird, R.M.; Ma, Z.; Dorabawila, N.; Pequegnat, B.; Omari, E.; Liu, Y.; Maue, A.C.; Poole, S.T.; Maciel, M.; Satish, K.; et al. Evaluation of a conjugate vaccine platform against enterotoxigenic Escherichia coli (ETEC), Campylobacter jejuni and Shigella. Vaccine 2018, 36, 6695–6702. [Google Scholar] [CrossRef]
- Fries, C.N.; Curvino, E.J.; Chen, J.L.; Permar, S.R.; Fouda, G.G.; Collier, J.H. Advances in nanomaterial vaccine strategies to address infectious diseases impacting global health. Nat. Nanotechnol. 2021, 16, 1–14. [Google Scholar] [CrossRef]
- Basu, R.; Zhai, L.; Contreras, A.; Tumban, E. Immunization with phage virus-like particles displaying Zika virus potential B-cell epitopes neutralizes Zika virus infection of monkey kidney cells. Vaccine 2018, 36, 1256–1264. [Google Scholar] [CrossRef]
- Basu, R.; Zhai, L.; Rosso, B.; Tumban, E. Bacteriophage Qbeta virus-like particles displaying Chikungunya virus B-cell epitopes elicit high-titer E2 protein antibodies but fail to neutralize a Thailand strain of Chikungunya virus. Vaccine 2020, 38, 2542–2550. [Google Scholar] [CrossRef]
- Warner, N.L.; Frietze, K.M. Development of Bacteriophage Virus-Like Particle Vaccines Displaying Conserved Epitopes of Dengue Virus Non-Structural Protein 1. Vaccines 2021, 9, 726. [Google Scholar] [CrossRef]
- Zha, L.; Chang, X.; Zhao, H.; Mohsen, M.O.; Hong, L.; Zhou, Y.; Chen, H.; Liu, X.; Zhang, J.; Li, D.; et al. Development of a Vaccine against SARS-CoV-2 Based on the Receptor-Binding Domain Displayed on Virus-Like Particles. Vaccines 2021, 9, 395. [Google Scholar] [CrossRef]
- Cabral-Miranda, G.; Lim, S.M.; Mohsen, M.O.; Pobelov, I.V.; Roesti, E.S.; Heath, M.D.; Skinner, M.A.; Kramer, M.F.; Martina, B.E.E.; Bachmann, M.F. Zika Virus-Derived E-DIII Protein Displayed on Immunologically Optimized VLPs Induces Neutralizing Antibodies without Causing Enhancement of Dengue Virus Infection. Vaccines 2019, 7, 72. [Google Scholar] [CrossRef]
- Sungsuwan, S.; Wu, X.; Shaw, V.; Kavunja, H.; McFall-Boegeman, H.; Rashidijahanabad, Z.; Tan, Z.; Lang, S.; Tahmasebi Nick, S.; Lin, P.H.; et al. Structure Guided Design of Bacteriophage Qbeta Mutants as Next Generation Carriers for Conjugate Vaccines. ACS Chem. Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, M.; Podsiadla-Bialoskorska, M.; Mielecki, D.; Ruffier, N.; Fateh, A.; Lambert, A.; Fanuel, M.; Camberlein, E.; Szolajska, E.; Grandjean, C. On the use of adenovirus dodecahedron as a carrier for glycoconjugate vaccines. Glycoconj. J. 2021, 38, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Zong, G.; Toonstra, C.; Yang, Q.; Zhang, R.; Wang, L.X. Chemoenzymatic Synthesis and Antibody Binding of HIV-1 V1/V2 Glycopeptide-Bacteriophage Qbeta Conjugates as a Vaccine Candidate. Int. J. Mol. Sci. 2021, 22, 1538. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Huo, C.X.; Lang, S.; Caution, K.; Nick, S.T.; Dubey, P.; Deora, R.; Huang, X. Chemical Synthesis and Immunological Evaluation of a Pentasaccharide Bearing Multiple Rare Sugars as a Potential Anti-pertussis Vaccine. Angew. Chem. Int. Ed. Engl. 2020, 59, 6451–6458. [Google Scholar] [CrossRef]
- Xu, L.; Li, Z.; Su, Z.; Yang, Y.; Ma, G.; Yu, R.; Zhang, S. Development of meningococcal polysaccharide conjugate vaccine that can elicit long-lasting and strong cellular immune response with hepatitis B core antigen virus-like particles as a novel carrier protein. Vaccine 2019, 37, 956–964. [Google Scholar] [CrossRef]
- Curley, S.M.; Putnam, D. Biological Nanoparticles in Vaccine Development. Front. Bioeng. Biotechnol. 2022, 10, 867119. [Google Scholar] [CrossRef]
- Flenniken, M.L.; Uchida, M.; Liepold, L.O.; Kang, S.; Young, M.J.; Douglas, T. A library of protein cage architectures as nanomaterials. Curr. Top Microbiol. Immunol. 2009, 327, 71–93. [Google Scholar] [CrossRef]
- Chakraborti, S.; Chakrabarti, P. Self-Assembly of Ferritin: Structure, Biological Function and Potential Applications in Nanotechnology. Adv. Exp. Med. Biol. 2019, 1174, 313–329. [Google Scholar] [CrossRef]
- Chen, R.R.R. Development of Receptor Binding Domain (RBD)-Conjugated Nanoparticle Vaccines with Broad Neutralization against SARS-CoV-2 Delta and Other Variants. Adv. Sci. 2022, 9, 5378. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhou, X.; Bian, Y.; Wang, S.; Chai, Q.; Guo, Z.; Wang, Z.; Zhu, P.; Peng, H.; Yan, X.; et al. Dual-targeting nanoparticle vaccine elicits a therapeutic antibody response against chronic hepatitis B. Nat. Nanotechnol. 2020, 15, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Zakeri, B.; Fierer, J.O.; Celik, E.; Chittock, E.C.; Schwarz-Linek, U.; Moy, V.T.; Howarth, M. Peptide tag forming a rapid covalent bond to a protein, through engineering a bacterial adhesin. Proc. Natl. Acad. Sci. USA 2012, 109, E690–E697. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Li, Z.; Yang, Y.; Ma, G.; Su, Z.; Zhang, S. An Apoferritin-Hemagglutinin Conjugate Vaccine with Encapsulated Nucleoprotein Antigen Peptide from Influenza Virus Confers Enhanced Cross Protection. Bioconjug. Chem. 2020, 31, 1948–1959. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.Y.Y. Apoferritin nanoparticle based dual-antigen influenza conjugate vaccine with potential cross-protective efficacy against heterosubtypic influenza virus. Particuology 2022, 64, 56–64. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef]
- Jiang, S.; Song, R.; Popov, S.; Mirshahidi, S.; Ruprecht, R.M. Overlapping synthetic peptides as vaccines. Vaccine 2006, 24, 6356–6365. [Google Scholar] [CrossRef]
- Zhang, H.; Hong, H.; Li, D.; Ma, S.; Di, Y.; Stoten, A.; Haig, N.; Di Gleria, K.; Yu, Z.; Xu, X.N.; et al. Comparing pooled peptides with intact protein for accessing cross-presentation pathways for protective CD8+ and CD4+ T cells. J. Biol. Chem. 2009, 284, 9184–9191. [Google Scholar] [CrossRef]
- He, Y.; Yu, W.; Shen, L.; Yan, W.; Xiao, L.; Qi, J.; Hu, T. A SARS-CoV-2 vaccine based on conjugation of SARS-CoV-2 RBD with IC28 peptide and mannan. Int. J. Biol. Macromol. 2022, 222, 661–670. [Google Scholar] [CrossRef]
- Azuar, A.; Shibu, M.A.; Adilbish, N.; Marasini, N.; Hung, H.; Yang, J.; Luo, Y.; Khalil, Z.G.; Capon, R.J.; Hussein, W.M.; et al. Poly(hydrophobic amino acid) Conjugates for the Delivery of Multiepitope Vaccine against Group A Streptococcus. Bioconjug. Chem. 2021, 32, 2307–2317. [Google Scholar] [CrossRef]
- Shalash, A.O.; Becker, L.; Yang, J.; Giacomin, P.; Pearson, M.; Hussein, W.M.; Loukas, A.; Skwarczynski, M.; Toth, I. Oral Peptide Vaccine against Hookworm Infection: Correlation of Antibody Titers with Protective Efficacy. Vaccines 2021, 9, 1034. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Sengupta, S.; Gupta, D.; Bhan, M.K.; Kumar, R.; Khan, A.; Jailkhani, B.S. Typhi derived OmpC peptide conjugated with Vi-polysaccharide evokes better immune response than free Vi-polysaccharide in mice. Biologicals 2019, 62, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Costantino, P.; Adamo, R. Potential targets for next generation antimicrobial glycoconjugate vaccines. FEMS Microbiol. Rev. 2018, 42, 388–423. [Google Scholar] [CrossRef] [PubMed]
- Del Bino, L.; Osterlid, K.E.; Wu, D.Y.; Nonne, F.; Romano, M.R.; Codee, J.; Adamo, R. Synthetic Glycans to Improve Current Glycoconjugate Vaccines and Fight Antimicrobial Resistance. Chem. Rev. 2022, 122, 15672–15716. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Shaikh, A.S.; Wang, F. Recent Advance in Tumor-associated Carbohydrate Antigens (TACAs)-based Antitumor Vaccines. ACS Chem. Biol. 2016, 11, 850–863. [Google Scholar] [CrossRef]
- Guo, C.; Manjili, M.H.; Subjeck, J.R.; Sarkar, D.; Fisher, P.B.; Wang, X.Y. Therapeutic cancer vaccines: Past, present, and future. Adv. Cancer Res. 2013, 119, 421–475. [Google Scholar] [CrossRef]
- Sorieul, C.; Papi, F.; Carboni, F.; Pecetta, S.; Phogat, S.; Adamo, R. Recent advances and future perspectives on carbohydrate-based cancer vaccines and therapeutics. Pharmacol. Ther. 2022, 235, 108158. [Google Scholar] [CrossRef]
- Shivatare, S.S.; Shivatare, V.S.; Wong, C.H. Glycoconjugates: Synthesis, Functional Studies, and Therapeutic Developments. Chem. Rev. 2022, 122, 15603–15671. [Google Scholar] [CrossRef]
- Hossain, F.; Andreana, P.R. Developments in Carbohydrate-Based Cancer Therapeutics. Pharmaceuticals 2019, 12, 84. [Google Scholar] [CrossRef]
- Bajad, N.G.; Swetha, R.; Gutti, G.; Singh, M.; Kumar, A.; Singh, S.K. A systematic review of carbohydrate-based bioactive molecules for Alzheimer’s disease. Future Med. Chem. 2021, 13, 1695–1711. [Google Scholar] [CrossRef]
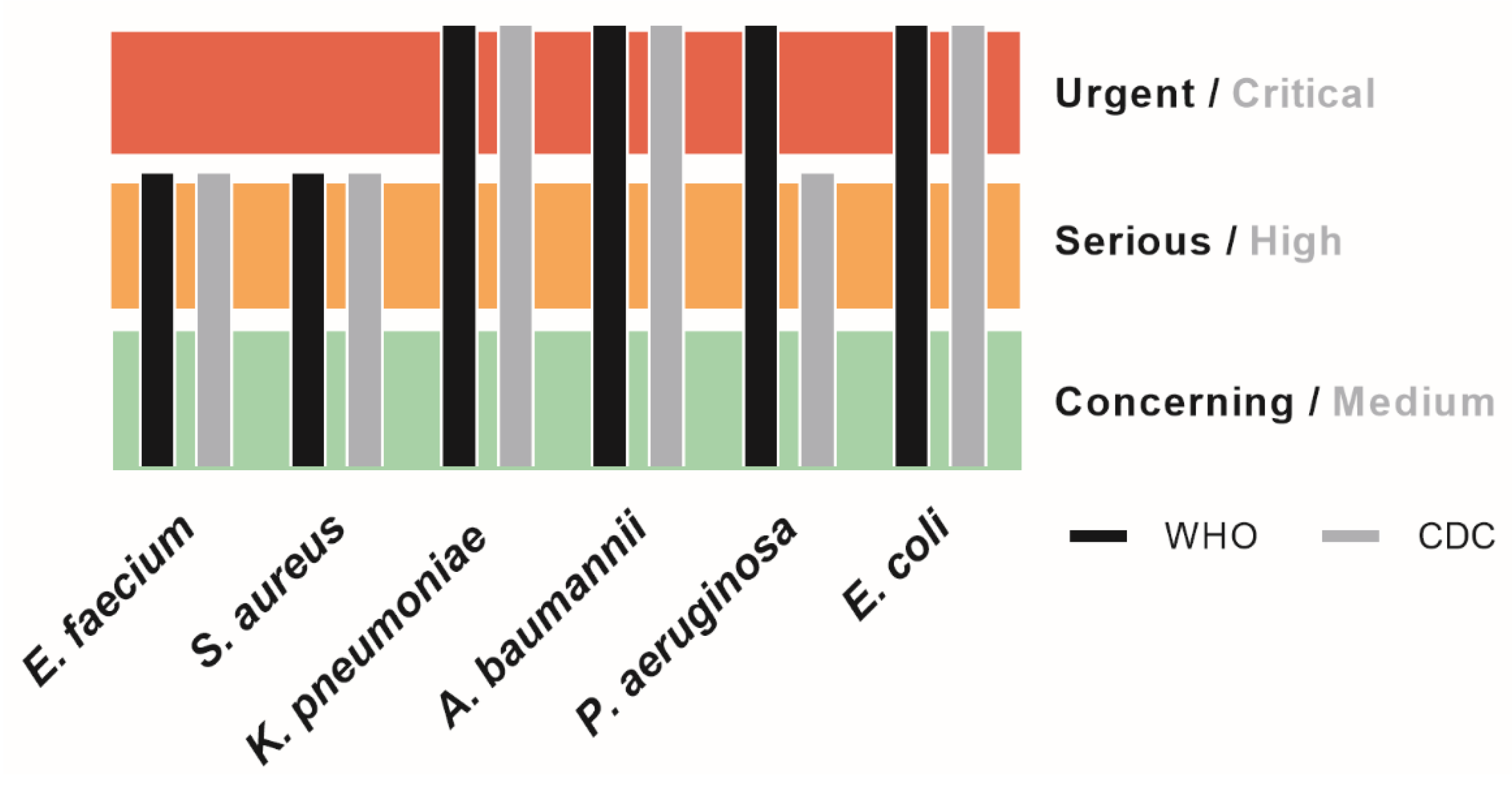
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance, C. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Laxminarayan, R. The overlooked pandemic of antimicrobial resistance. Lancet 2022, 399, 606–607. [Google Scholar] [CrossRef] [PubMed]
- Gautam, A. Antimicrobial Resistance: The Next Probable Pandemic. JNMA J. Nepal. Med. Assoc. 2022, 60, 225–228. [Google Scholar] [CrossRef] [PubMed]
- WHO. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, Including Tuberculosis. Available online: https://www.who.int/publications/i/item/WHO-EMP-IAU-2017.12 (accessed on 11 November 2022).
- CDC. 2019 AR Threats Report. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html#carp (accessed on 11 November 2022).
- European Antimicrobial Resistance, C. The burden of bacterial antimicrobial resistance in the WHO European region in 2019: A cross-country systematic analysis. Lancet Public Health 2022, 7, e897–e913. [Google Scholar] [CrossRef]
- Teng, F.; Singh, K.V.; Bourgogne, A.; Zeng, J.; Murray, B.E. Further characterization of the epa gene cluster and Epa polysaccharides of Enterococcus faecalis. Infect. Immun. 2009, 77, 3759–3767. [Google Scholar] [CrossRef]
- Kalfopoulou, E.; Laverde, D.; Miklic, K.; Romero-Saavedra, F.; Malic, S.; Carboni, F.; Adamo, R.; Lenac Rovis, T.; Jonjic, S.; Huebner, J. Development of Opsonic Mouse Monoclonal Antibodies against Multidrug-Resistant Enterococci. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef]
- Zhou, Z.; Ding, W.; Li, C.; Wu, Z. Synthesis and immunological study of a wall teichoic acid-based vaccine against E. faecium U0317. J. Carbohydr. Chem. 2017, 36, 205–219. [Google Scholar] [CrossRef]
- Zhao, M.; Qin, C.; Li, L.; Xie, H.; Ma, B.; Zhou, Z.; Yin, J.; Hu, J. Conjugation of Synthetic Trisaccharide of Staphylococcus aureus Type 8 Capsular Polysaccharide Elicits Antibodies Recognizing Intact Bacterium. Front. Chem. 2020, 8, 258. [Google Scholar] [CrossRef]
- Lin, T.L.; Yang, F.L.; Ren, C.T.; Pan, Y.J.; Liao, K.S.; Tu, I.F.; Chang, Y.P.; Cheng, Y.Y.; Wu, C.Y.; Wu, S.H.; et al. Development of Klebsiella pneumoniae Capsule Polysaccharide-Conjugated Vaccine Candidates Using Phage Depolymerases. Front. Immunol. 2022, 13, 843183. [Google Scholar] [CrossRef]
- Wei, R.; Yang, X.; Liu, H.; Wei, T.; Chen, S.; Li, X. Synthetic Pseudaminic-Acid-Based Antibacterial Vaccine Confers Effective Protection against Acinetobacter baumannii Infection. ACS Cent. Sci. 2021, 7, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Jamshidi, M.P.; Cairns, C.; Chong, S.; St Michael, F.; Vinogradov, E.V.; Cox, A.D.; Sauvageau, J. Synthesis and Immunogenicity of a Methyl Rhamnan Pentasaccharide Conjugate from Pseudomonas aeruginosa A-Band Polysaccharide. ACS Infect. Dis. 2022, 8, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Kowarik, M.; Wetter, M.; Haeuptle, M.A.; Braun, M.; Steffen, M.; Kemmler, S.; Ravenscroft, N.; De Benedetto, G.; Zuppiger, M.; Sirena, D.; et al. The development and characterization of an E. coli O25B bioconjugate vaccine. Glycoconj. J. 2021, 38, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Sorrell, T.C.; Packham, D.R.; Shanker, S.; Foldes, M.; Munro, R. Vancomycin therapy for methicillin-resistant Staphylococcus aureus. Ann. Intern. Med. 1982, 97, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Sievert, D.M.; Hageman, J.C.; Boulton, M.L.; Tenover, F.C.; Downes, F.P.; Shah, S.; Rudrik, J.T.; Pupp, G.R.; Brown, W.J.; et al. Infection with vancomycin-resistant Staphylococcus aureus containing the vanA resistance gene. N. Engl. J. Med. 2003, 348, 1342–1347. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar]
- O’Riordan, K.; Lee, J.C. Staphylococcus aureus capsular polysaccharides. Clin. Microbiol. Rev. 2004, 17, 218–234. [Google Scholar] [CrossRef]
- Gening, M.L.; Pier, G.B.; Nifantiev, N.E. Broadly protective semi-synthetic glycoconjugate vaccine against pathogens capable of producing poly-beta-(1-->6)-N-acetyl-d-glucosamine exopolysaccharide. Drug Discov. Today Technol. 2020, 35-36, 13–21. [Google Scholar] [CrossRef]
- Choi, M.; Hegerle, N.; Nkeze, J.; Sen, S.; Jamindar, S.; Nasrin, S.; Sen, S.; Permala-Booth, J.; Sinclair, J.; Tapia, M.D.; et al. The Diversity of Lipopolysaccharide (O) and Capsular Polysaccharide (K) Antigens of Invasive Klebsiella pneumoniae in a Multi-Country Collection. Front. Microbiol. 2020, 11, 1249. [Google Scholar] [CrossRef]
- Cryz, S.J. Progress in immunization against Klebsiella infections. Eur. J. Clin. Microbiol. 1983, 2, 523–528. [Google Scholar] [CrossRef]
- Ravinder, M.; Liao, K.S.; Cheng, Y.Y.; Pawar, S.; Lin, T.L.; Wang, J.T.; Wu, C.Y. A Synthetic Carbohydrate-Protein Conjugate Vaccine Candidate against Klebsiella pneumoniae Serotype K2. J. Org. Chem. 2020, 85, 15964–15997. [Google Scholar] [CrossRef] [PubMed]
- Ghaderinia, P.; Shapouri, R.; Rostamizadeh, K.; Khodavandi, A.; Mahdavi, M. Capsular K-antigen-PLGA Nano conjugated Vaccine against Klebsiella pneumoniea pneumoniae K2O1 Infection. J. Adv. Med. Biomed. Res. 2022, 30, 73–74. [Google Scholar] [CrossRef]
- Peng, Z.; Wu, J.; Wang, K.; Li, X.; Sun, P.; Zhang, L.; Huang, J.; Liu, Y.; Hua, X.; Yu, Y.; et al. Production of a Promising Biosynthetic Self-Assembled Nanoconjugate Vaccine against Klebsiella Pneumoniae Serotype O2 in a General Escherichia Coli Host. Adv. Sci. (Weinh) 2021, 8, e2100549. [Google Scholar] [CrossRef] [PubMed]
- Hegerle, N.; Choi, M.; Sinclair, J.; Amin, M.N.; Ollivault-Shiflett, M.; Curtis, B.; Laufer, R.S.; Shridhar, S.; Brammer, J.; Toapanta, F.R.; et al. Development of a broad spectrum glycoconjugate vaccine to prevent wound and disseminated infections with Klebsiella pneumoniae and Pseudomonas aeruginosa. PLoS ONE 2018, 13, e0203143. [Google Scholar] [CrossRef] [PubMed]
- Seeberger, P.H.; Pereira, C.L.; Khan, N.; Xiao, G.; Diago-Navarro, E.; Reppe, K.; Opitz, B.; Fries, B.C.; Witzenrath, M. A Semi-Synthetic Glycoconjugate Vaccine Candidate for Carbapenem-Resistant Klebsiella pneumoniae. Angew. Chem. Int. Ed. Engl. 2017, 56, 13973–13978. [Google Scholar] [CrossRef] [PubMed]
- Giguere, D. Surface polysaccharides from Acinetobacter baumannii: Structures and syntheses. Carbohydr. Res. 2015, 418, 29–43. [Google Scholar] [CrossRef]
- Rudenko, N.; Karatovskaya, A.; Zamyatina, A.; Shepelyakovskaya, A.; Semushina, S.; Brovko, F.; Shpirt, A.; Torgov, V.; Kolotyrkina, N.; Zinin, A.; et al. Immune Response to Conjugates of Fragments of the Type K9 Capsular Polysaccharide of Acinetobacter baumannii with Carrier Proteins. Microbiol. Spectr. 2022, 10, e0167422. [Google Scholar] [CrossRef]
- Li, X.; Pan, C.; Liu, Z.; Sun, P.; Hua, X.; Feng, E.; Yu, Y.; Wu, J.; Zhu, L.; Wang, H. Safety and immunogenicity of a new glycoengineered vaccine against Acinetobacter baumannii in mice. Microb. Biotechnol. 2022, 15, 703–716. [Google Scholar] [CrossRef]
- Lee, I.M.; Yang, F.L.; Chen, T.L.; Liao, K.S.; Ren, C.T.; Lin, N.T.; Chang, Y.P.; Wu, C.Y.; Wu, S.H. Pseudaminic Acid on Exopolysaccharide of Acinetobacter baumannii Plays a Critical Role in Phage-Assisted Preparation of Glycoconjugate Vaccine with High Antigenicity. J. Am. Chem. Soc. 2018, 140, 8639–8643. [Google Scholar] [CrossRef]
- Pier, G.B. The challenges and promises of new therapies for cystic fibrosis. J. Exp. Med. 2012, 209, 1235–1239. [Google Scholar] [CrossRef]
- Liu, P.V. Comparison of the Chinese schema and the International Antigenic Typing System for serotyping Pseudomonas aeruginosa. J. Clin. Microbiol. 1987, 25, 824–826. [Google Scholar] [CrossRef] [PubMed]
- Estahbanati, H.K.; Kashani, P.P.; Ghanaatpisheh, F. Frequency of Pseudomonas aeruginosa serotypes in burn wound infections and their resistance to antibiotics. Burns 2002, 28, 340–348. [Google Scholar] [CrossRef]
- Lu, Q.; Eggimann, P.; Luyt, C.E.; Wolff, M.; Tamm, M.; Francois, B.; Mercier, E.; Garbino, J.; Laterre, P.F.; Koch, H.; et al. Pseudomonas aeruginosa serotypes in nosocomial pneumonia: Prevalence and clinical outcomes. Crit. Care 2014, 18, R17. [Google Scholar] [CrossRef] [PubMed]
- Maleki, M.; Azimi, S.; Salouti, M. Protective effect of two new nanovaccines against Pseudomonas aeruginosa based on LPS and OPS: A comparison study. Immunobiology 2022, 227, 152278. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C. Biosynthesis and assembly of capsular polysaccharides in Escherichia coli. Annu. Rev. Biochem. 2006, 75, 39–68. [Google Scholar] [CrossRef]
- Saade, E.; Gravenstein, S.; Donskey, C.J.; Wilson, B.; Spiessens, B.; Abbanat, D.; Poolman, J.; de Palacios, P.I.; Hermans, P. Characterization of Escherichia coli isolates potentially covered by ExPEC4V and ExPEC10V, that were collected from post-transrectal ultrasound-guided prostate needle biopsy invasive urinary tract and bloodstream infections. Vaccine 2020, 38, 5100–5104. [Google Scholar] [CrossRef]
- McConnell, M.J.; Rumbo, C.; Bou, G.; Pachon, J. Outer membrane vesicles as an acellular vaccine against Acinetobacter baumannii. Vaccine 2011, 29, 5705–5710. [Google Scholar] [CrossRef]
- Roberts, R.; Moreno, G.; Bottero, D.; Gaillard, M.E.; Fingermann, M.; Graieb, A.; Rumbo, M.; Hozbor, D. Outer membrane vesicles as acellular vaccine against pertussis. Vaccine 2008, 26, 4639–4646. [Google Scholar] [CrossRef]
- Shang, E.S.; Champion, C.I.; Wu, X.Y.; Skare, J.T.; Blanco, D.R.; Miller, J.N.; Lovett, M.A. Comparison of protection in rabbits against host-adapted and cultivated Borrelia burgdorferi following infection-derived immunity or immunization with outer membrane vesicles or outer surface protein A. Infect. Immun. 2000, 68, 4189–4199. [Google Scholar] [CrossRef]
- Keenan, J.; Day, T.; Neal, S.; Cook, B.; Perez-Perez, G.; Allardyce, R.; Bagshaw, P. A role for the bacterial outer membrane in the pathogenesis of Helicobacter pylori infection. FEMS Microbiol. Lett. 2000, 182, 259–264. [Google Scholar] [CrossRef]
- Lee, J.C.; Lee, E.J.; Lee, J.H.; Jun, S.H.; Choi, C.W.; Kim, S.I.; Kang, S.S.; Hyun, S. Klebsiella pneumoniae secretes outer membrane vesicles that induce the innate immune response. FEMS Microbiol. Lett. 2012, 331, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Ellis, T.N.; Leiman, S.A.; Kuehn, M.J. Naturally produced outer membrane vesicles from Pseudomonas aeruginosa elicit a potent innate immune response via combined sensing of both lipopolysaccharide and protein components. Infect. Immun. 2010, 78, 3822–3831. [Google Scholar] [CrossRef] [PubMed]
- van der Put, R.M.F.; Smitsman, C.; de Haan, A.; Hamzink, M.; Timmermans, H.; Uittenbogaard, J.; Westdijk, J.; Stork, M.; Ophorst, O.; Thouron, F.; et al. The First-in-Human Synthetic Glycan-Based Conjugate Vaccine Candidate against Shigella. ACS Cent. Sci. 2022, 8, 449–460. [Google Scholar] [CrossRef] [PubMed]
- FDA. Vaccines Licensed for Use in the United States. Available online: https://www.fda.gov/vaccines-blood-biologics/vaccines/vaccines-licensed-use-united-states (accessed on 17 November 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| OMV | S. typhimurium | SARS-CoV-2 spike receptor-binding domain (RBD) | SpyCatcher-SpyTag [1] | Jiang et al., 2022 [66] |
| GMMA | S. typhimurium | Group A Streptococcus cell wall carbohydrate | Reductive amination | Palmieri et al., 2022 [65] |
| GMMA | S. typhimurium | Malaria transmission-blocking protein Pfs25 | Oxidation/reductive amination | Di Benedetto et al., 2021 [67] |
| GMMA | S. typhimurium | P. falciparum circumsporozoite protein | Thiol-maleimide | Micoli et al., 2020 [53] |
| S. sonnei | E. coli SslE | Reductive amination | ||
| N. meningitidis type B/S. typhimurium | N. meningitidis serogroups A and C oligosaccharides | SIDEA | ||
| OMV | N. meningitidis type B | Malaria transmission-blocking antigen Pfs230 | Thiol-maleimide | Scaria et al., 2019 [68] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Protein/Bi-valent | P. aeruginosa exotoxin A | E. coli O2, O6A and O25B | Bioconjugation | Nicolardi et al. [72] |
| Polysaccharide/Quadrivalent | P. aeruginosa exotoxin A | S. dysenteriae type O1, O antigen from S. flexneri 2a, 3a and 6 | Martin et al., 2022 [75] | |
| Polysaccharide/monovalent | P. aeruginosa exotoxin A | S. flexneri 2a O-polysaccharide | Ravenscroft et al., 2019 [76] | |
| Polysaccharide/protein Bi-valent | P. aeruginosa exotoxin A | K. pneumoniae K1 and K2 CPSs | Feldman et al., 2019 [77] | |
| Oligosaccharide/mono-, bi-, and trivalent | E. coli Acceptor protein ComP | S. pneumoniae CPS | Harding et al., 2019 [74] | |
| Polysaccharide/monovalent | S. pneumoniae NanA, PiuA, and Sp0148 | S. pneumoniae serotype 4 CPS | Reglinksy et al., 2018 [78] | |
| OMV | E. coli OMV | Poly-N-acetyl-d-glucosamine (rPNAG) | Stevenson et al., 2018 [79] | |
| Polysaccharide/monovalent | S. paratyphi A antigenic peptide (P2) | S. enterica serovar Paratyphi A O-polysaccharide | Sun et al., 2018 [80] | |
| Polysaccharide/monovalent | P. aeruginosa exotoxin A | F. tularensis O-antigen | Marshall et al., 2018 [81] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Bivalent | Genetically detoxified Tetanus toxin (8MTT) | Cattle tick fever peptide P0, H. influenzae type b (PRP) | Thiol-maleimide, CDAP | Chang et al., 2022 [86] |
| Bivalent | S. pneumoniae serotype type 14 CPS | recombinant SARS-CoV-2 RBD | Reductive amination | Deng et al., 2022 [87] |
| Monovalent | Group A Streptococcus Streptolysin O | Group A Streptococcus cell-wall oligosaccharides | Click chemistry | Kapoor et al., 2022 [84] |
| Bivalent | Rotavirus recombinant ΔVP8 protein | S. Typhi capsular polysaccharide (Vi) | EDAC-ADH | Park et al., 2021 [88] |
| Bivalent | Streptococcus C5a peptidase ScpA193, Fn and Fn2 | Group A Streptococcus cell-wall trisaccharide | di(N-succinimidyl) glutarate | Wang et al., 2021 [83] |
| Monovalent | S. aureus fusion protein (Hla-MntC-ACOL0723) | S. aureus 5 (CP5, Reynolds strain) and 8 (CP8, Becker strain) | Carbodiimide | Ahmadi et al., 2020 [89] |
| Monovalent | S. typhimurium flagellin | S. typhimurium lipid-A free lipopolysaccharide | Decarboxylative amidation | Chiu et al., 2020 [90] |
| Monovalent | Recombinant tetanus toxoid heavy chain fragment | HIV-1-fusion peptide (FP8) | Sulfo-SIAB | Ou et al., 2020 [91] |
| Bivalent | Hepatitis B virus surface antigen | Pneumococcal type 33 F-capsular polysaccharide | Carbodiimide | Qian et al., 2020 [92] |
| Bivalent | Recombinant Tetanus Toxoid Heavy Chain Fragment C | HIV-1 fusion peptide (FP) with eight amino acid residues (FP8) | Amine-to-sulfhydryl, Thiol-maleimide | Yang et al., 2020 [93] |
| Monovalent | Enterococcus secreted antigen A and the peptidyl-prolyl cis-trans isomerase | E. faecalis polysaccharide di-heteroglycan | CDAP | Romero-Saavedra et al., 2019 [85] |
| Bivalent | Plasmodium falciparum Pfs25 | S. Typhi Vi capsular polysaccharide | Carbodiimide | An et al., 2018 [94] |
| Monovalent | S. enteritidis homologous serovar phase 1 flagellin protein | S. enteritidis core and O-polysaccharide (COPS) | CDAP | Baliban et al., 2018 [95] |
| Monovalent | Recombinant Tetanus Toxoid Heavy Chain Fragment C | Oxycodone-based hapten | Carbodiimide | Baruffaldi et al., 2018 [96] |
| Monovalent, bivalent, and trivalent | ETEC adhesins CFA/I and CS6 | C. jejuni and Shigella polysaccharides and Shigella flexneri LPS | TEMPO-mediated oxidation and carbodiimide | Laird et al., 2018 [97] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Protein display | Qβ | Dengue virus synthetic peptides | Protein display | Warner et al., 2021 [101] |
| Protein display | Cucumber mosaic virus | Recombinantly expressed SARS-CoV-2 receptor-binding domain | Protein display | Zha et al., 2021 [102] |
| Protein display | Cucumber mosaic virus | Zika virus E-DIII protein | Protein display | Cabral-Miranda et al., 2019 [103] |
| Protein display | Qβ | Zika virus MS2 and PP7/Chikungunya virus B-cell synthetic peptides | Protein display | Basu et al., 2018, 2020 [99,100] |
| Chemical conjugation | Qβ | Thomsen-nouveau antigen, GD2 protein, SARS-CoV-2 peptides | diNHS ester adipate | Sungsuwan et al., 2022 [104] |
| Chemical conjugation | Recombinant adenoviral type 3 dodecahedron | S. pneumoniae serotype 14 CPS trisaccharide | Glutaraldehyde | Prasanna et al., 2021 [105] |
| Chemical conjugation | Qβ | HIV-1 V1V2 glycopeptide | Click chemistry | Zong et al., 2021 [106] |
| Chemical conjugation | Qβ | Synthetic Pertussis LPS-like pentasaccharide | diNHS ester adipate | Wang et al., 2020 [107] |
| Chemical conjugation | Full-length hepatitis B core antigen virus-like particles | Meningococcal group C polysaccharides | Amine-PEG-maleimide | Xu et al., 2019 [108] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Tri-valent | E. coli Sd-ferritin | SARS-CoV-2 RBD B.1.617.2, D614G and B.1.351 | SpyCatcher-SpyTag | Chen et al., 2022 [112] |
| Monovalent | Horse spleen apoFerritin | Influenza virus PR8 H1N1 hemagglutinin and M2e peptide | Thiol-maleimide | Sheng et al., 2022 [116] |
| Monovalent | E. coli SpyTag–ferritin | ΔN SpyCatcher-fused preS1 | SpyCatcher-SpyTag | Wang et al., 2020 [113] |
| Bivalent | Horse spleen apoFerritin | Influenza virus hemagglutinin | Thiol-maleimide | Wei et al., 2020 [115] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Semi-synthetic | IC28 peptide from bacterial flagellin | Recombinant SARS-CoV-2 RBD | Thiol-maleimide | He et al., 2022 [120] |
| Synthetic | PADRE and polyleucine | S. pyogenes (GAS) M-protein-derived B-cell epitopes J8, PL1, and 88/30 | Click-chemistry | Azuar et al., 2021 [121] |
| Synthetic | T-helper cell epitope P25 and polyleucine | Hookworm APR-1 B-cell epitope (p3) | Click-chemistry | Shalash et al., 2021 [122] |
| Semi-synthetic | S. Typhi OmpC synthetic peptide | S. Typhi Vi polysaccharide | ADH-EDC | Haque et al., 2019 [123] |
| Species | AMR |
|---|---|
| Enterococcus faecium | Vancomycin |
| Staphylococcus aureus | Methicillin and Vancomycin |
| Klebsiella pneumoniae | Carbapenem, ESBL [a] |
| Acinetobacter baumannii | Carbapenem |
| Pseudomonas aeruginosa | Carbapenem |
| Escherichia coli | Carbapenem, ESBL [a] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Traditional extracted | Enterococcus secreted antigen A | Di-heteroglycan | CDAP | Kalfopoulou et al., 2019 [140] |
| Traditional extracted | Enterococcus secreted antigen A and peptidyl-prolyl cis-trans isomerase | Di-heteroglycan | CDAP | Romero-Saavedra et al., 2019 [85] |
| Semi-synthetic | KLH and HSA | Cell wall teichoic acid | Disuccinimidyl glutarate | Zhou et al., 2017 [141] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Semi-synthetic | Tetanus toxoid, Shiga toxin 1b subunit (Stx1b) and S. aureus alpha-hemolysin (Hla H35L) | Synthetic penta- and nona-β-(1→6)-d-glucosamine (PNAG) | Thiol-maleimide | Gening et al., 2021 [151] |
| Semi-synthetic | CRM197 | Capsular polysaccharide type 8 trisaccharide | Bis(p-nitrophenyl adipate) | Zhao et al., 2020 [142] |
| Bioconjugate | E. coli OMV | Poly-N-acetyl-d-glucosamine (rPNAG) | Bioconjugation | Stevenson et al., 2018 [79] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Traditional extracted | Polylactic-co-glycolic acid (PLGA) | K. pneumoniae K2O1 capsule antigen | W/O/W emulsion | Ghaderinia et al., 2022 [155] |
| Traditional extracted | CRM197 | Capsular polysaccharide K1 and K2 | Thiol-maleimide | Lin et al., 2022 [143] |
| Bioconjugate | E. coli | K. pneumoniae serotype O2 polysaccharide | Bioconjugation | Peng et al., 2021 [156] |
| Semi-synthetic | CRM197 | K2 hexa-, hepta-, and octa-saccharide | Thiol-maleimide | Ravinder et al., 2020 [154] |
| Bioconjugate | P. aeruginosa exotoxin A | Capsular polysaccharide K1 and K2 | Bioconjugation | Feldman et al., 2019 [77] |
| Traditional extracted | P. aeruginosa rFlaA and rFlaB | O-polysaccharide O1, O2, O3 and O5 | Thiol-maleimide | Hegerle et al., 2018 [157] |
| Semi-synthetic | CRM197 | Synthetic hexasasaccharide | p-nitrophenyl adipate ester | Seeberger et al., 2017 [158] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Traditional extracted | BSA, OVA, KLH | CPS K9 di- and trimers | Reductive amination, squaric acid | Rudenko et al., 2022 [160] |
| Bioconjugate | cholera toxin B subunit | O-linked PgIS | Bioconjugation | Li et al., 2022 [161] |
| Semi-synthetic | CRM197 | Pseudaminic acid | OPA (ortho-phthalaldehyde) | Wei et al., 2021 [144] |
| Semi-synthetic | CRM197 | Pseudaminic acid | Thiol-maleimide | Lee et al., 2018 [162] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Semi-synthetic | PLGA | P. aeruginosa LPS and OPS | Carbodiimide | Maleki et al., 2022 [167] |
| Semi-synthetic | HSA | Methyl Rhamnan Pentasaccharide | Reductive amination | Jamshidi et al., 2022 [145] |
| Traditional extracted | P. aeruginosa rFlaA and rFlaB | O-polysaccharide O1, O2, O3 and O5 | Thiol-maleimide | Hegerle et al., 2018 [157] |
| Vaccine Type | Carrier | Antigen | Chemistry | References |
|---|---|---|---|---|
| Semi-synthetic | Tetanus toxoid, Shiga toxin 1b subunit (Stx1b) and S. aureus alpha-hemolysin (Hla H35L) | Synthetic penta- and nona-β-(1→6)-d-glucosamine (PNAG) | Thiol-maleimide | Gening et al., 2021 [151] |
| Bioconjugate | P. aeruginosa Exotoxin A | E. coli O25B | Bioconjugation | Kowarik et al., 2021 [146] |
| Bioconjugate | P. aeruginosa exotoxin A | O1, O2, O4, O6, O8, O15, O16, O18, O25 and O75 | Bioconjugation | Saade et al., 2020 [169] |
| Bioconjugate | P. aeruginosa Exotoxin A | E. coli O2, O6A and O25B | Bioconjugation | Nicolardi et al. [72] |
| Bioconjugate | E. coli OMV | Poly-N-acetyl-d-glucosamine (rPNAG) | Bioconjugation | Stevenson et al., 2018 [79] |
| Semi-synthetic | Tetanus toxoid, Shiga toxin 1b subunit (Stx1b) and S. aureus alpha-hemolysin (Hla H35L) | Synthetic penta- and nona-β-(1→6)-D-glucosamine (PNAG) | Thiol-maleimide | Gening et al., 2021 [151] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Put, R.M.F.; Metz, B.; Pieters, R.J. Carriers and Antigens: New Developments in Glycoconjugate Vaccines. Vaccines 2023, 11, 219. https://doi.org/10.3390/vaccines11020219
van der Put RMF, Metz B, Pieters RJ. Carriers and Antigens: New Developments in Glycoconjugate Vaccines. Vaccines. 2023; 11(2):219. https://doi.org/10.3390/vaccines11020219
Chicago/Turabian Stylevan der Put, Robert M.F., Bernard Metz, and Roland J. Pieters. 2023. "Carriers and Antigens: New Developments in Glycoconjugate Vaccines" Vaccines 11, no. 2: 219. https://doi.org/10.3390/vaccines11020219
APA Stylevan der Put, R. M. F., Metz, B., & Pieters, R. J. (2023). Carriers and Antigens: New Developments in Glycoconjugate Vaccines. Vaccines, 11(2), 219. https://doi.org/10.3390/vaccines11020219