
SARS-CoV-2 Antibody Profiles in Maternal Serum and Breast Milk Following mRNA COVID-19 Vaccination: A Longitudinal Prospective Observational Cohort Study

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participant Enrollment and Sample Collection and Processing
2.2. Meso Scale Discovery (MSD) Assays to Detect IgG and IgA Levels in Serum and Breast Milk
2.3. Production of Pseudovirus and Titration Assay
2.4. Ancestral SARS-CoV-2 and Omicron BA.4/5 Pseudovirus-Based Neutralization Assay
2.5. Statistical Analyses
3. Results
3.1. Clinical Cohort Characteristics
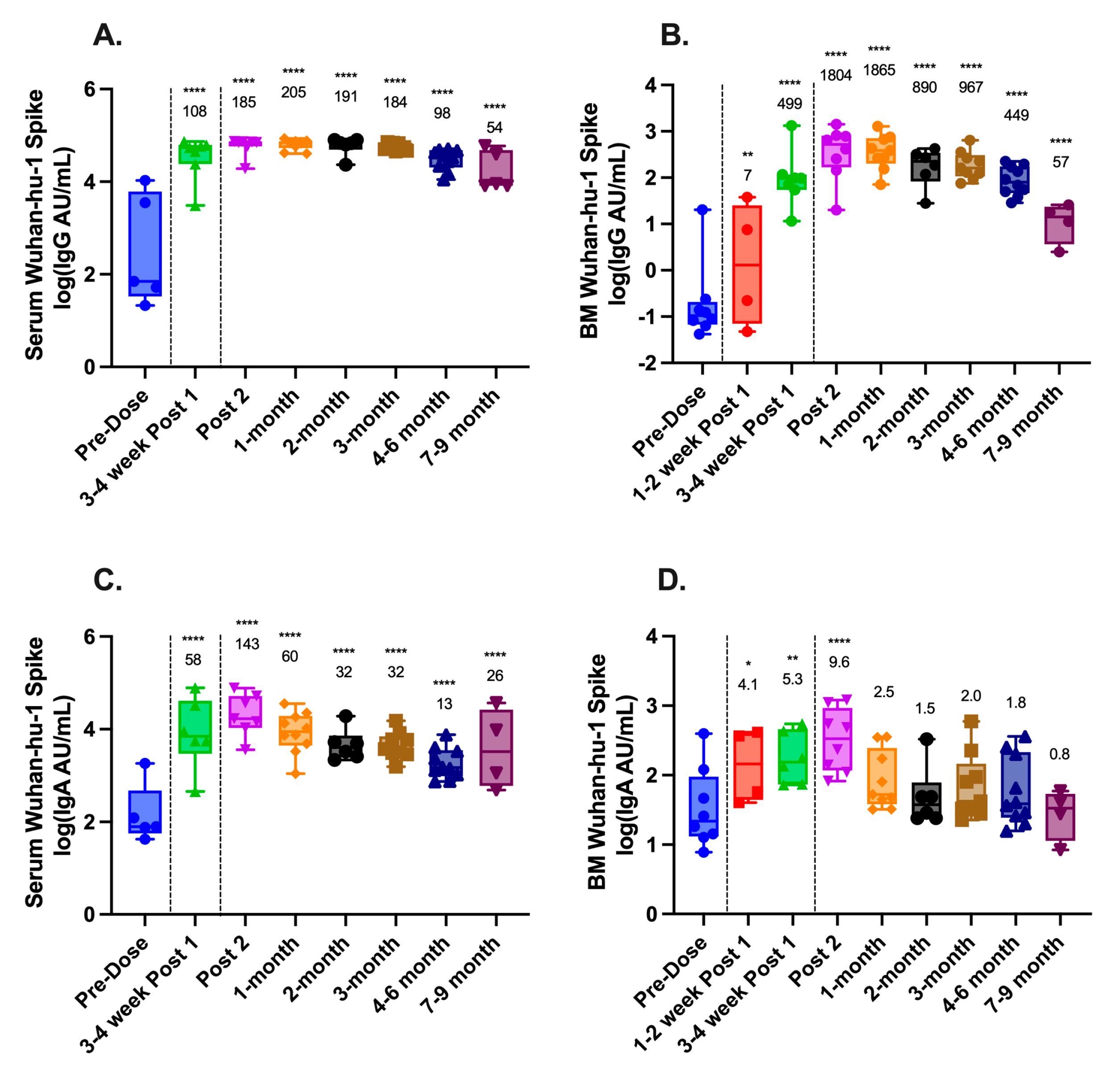
3.2. SARS-CoV-2-Binding Antibodies in Serum and Breast Milk
3.3. SARS-CoV-2 Pseudovirus-Neutralizing Antibodies in Serum and Breast Milk
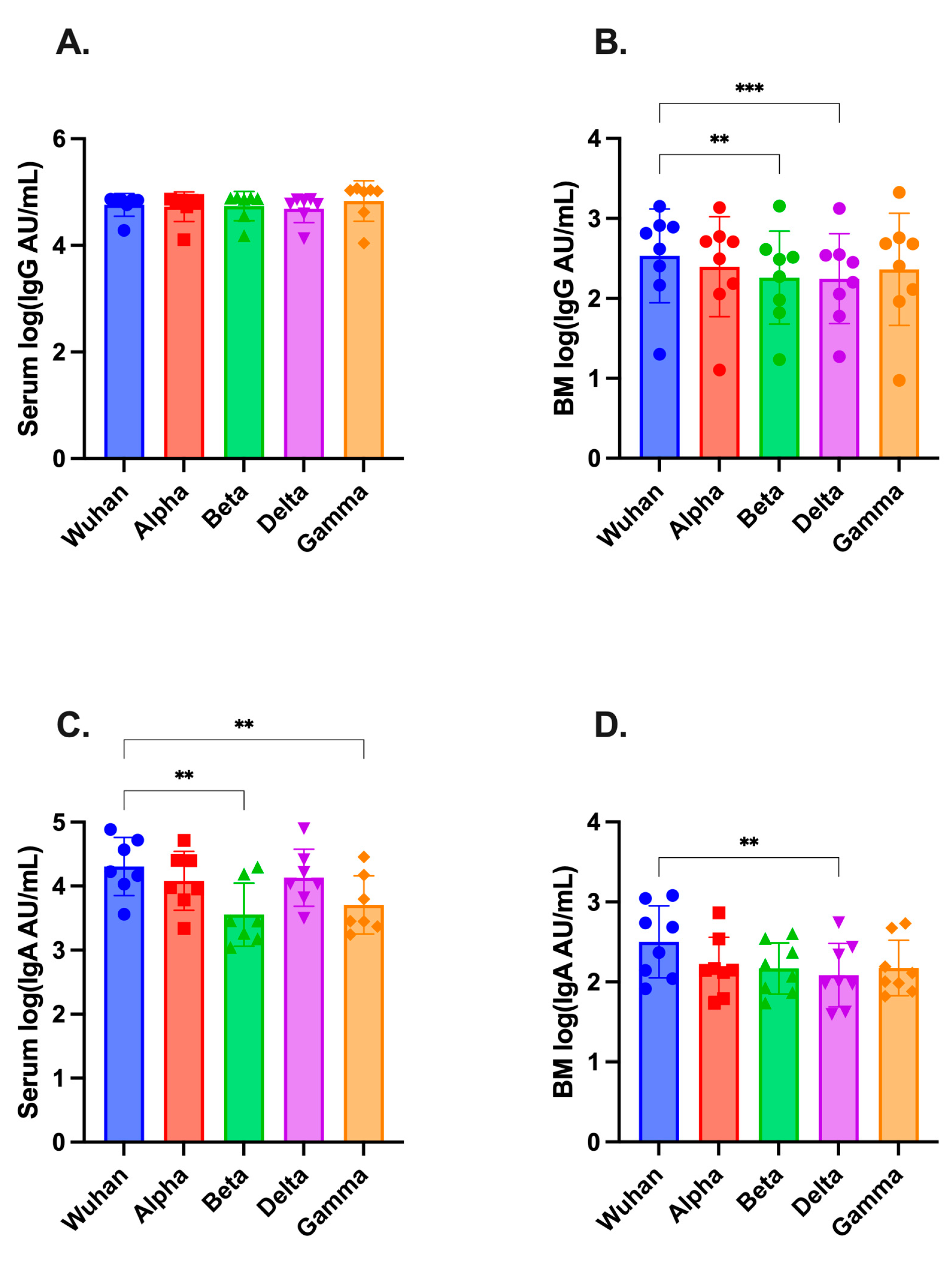
3.4. SARS-CoV-2-Variant Antibodies in Serum and Breast Milk
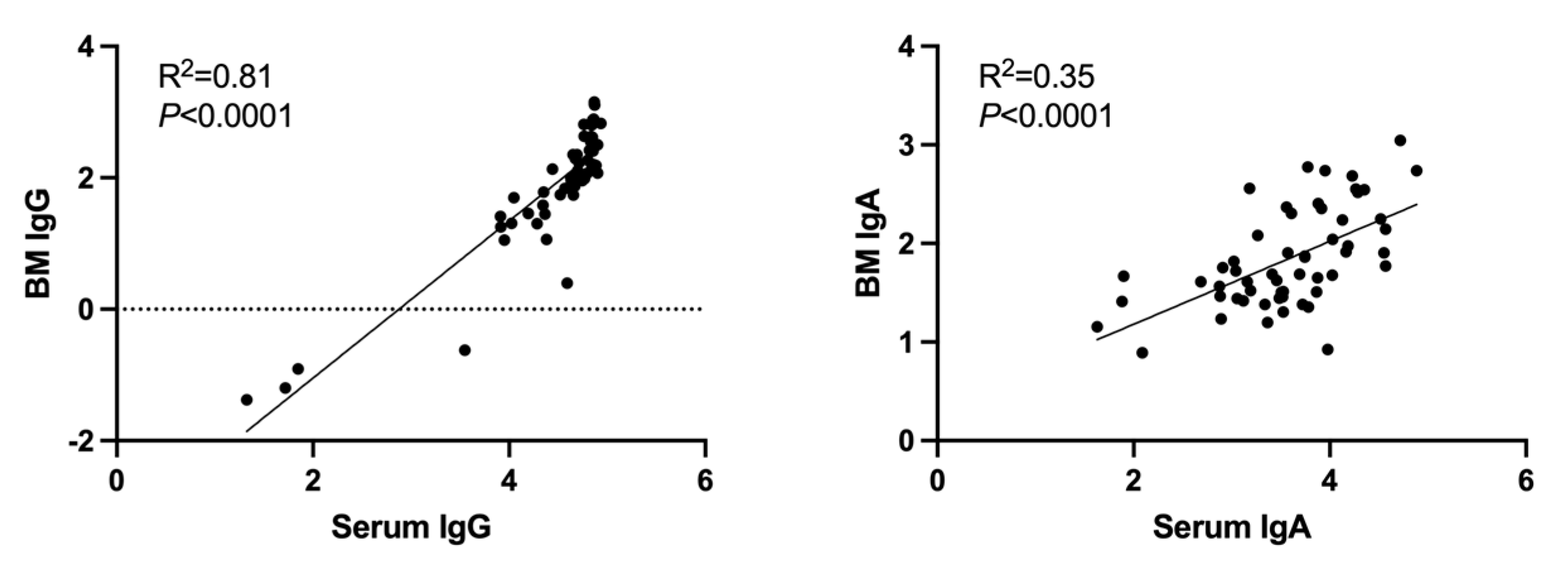
3.5. Correlation between SARS-CoV-2 Serum and Breast-Milk Antibodies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: A mathematical modelling study. Lancet Infect. Dis. 2022, 22, 1293–1302. [Google Scholar] [CrossRef]
- Children and COVID-19 Vaccination Trends: Summary of Data Publicly Reported by the Centers for Disease Control and Prevention. Available online: https://www.aap.org/en/pages/2019-novel-coronavirus-covid-19-infections/children-and-covid-19-vaccination-trends/ (accessed on 22 August 2023).
- COVID-19 Vaccinations in the United States. Available online: https://covid.cdc.gov/covid-data-tracker/#vaccinations_vacc-people-booster-percent-pop5 (accessed on 22 August 2023).
- Hospitalization of Infants and Children Aged 0–4 Years with Laboratory-Confirmed COVID-19—COVID-NET, 14 States, March 2020–February 2022. Available online: https://www.cdc.gov/mmwr/volumes/71/wr/mm7111e2.htm (accessed on 22 August 2023).
- Hunagund, S.; Golan, Y.; Asiodu, I.V.; Prahl, M.; Gaw, S.L. Effects of Vaccination Against Influenza, Pertussis, and COVID-19 on Human Milk Antibodies: Current Evidence and Implications for Health Equity. Front. Immunol. 2022, 13, 910383. [Google Scholar] [CrossRef]
- Perl, S.H.; Uzan-Yulzari, A.; Klainer, H.; Asiskovich, L.; Youngster, M.; Rinott, E.; Youngster, I. SARS-CoV-2-Specific Antibodies in Breast Milk After COVID-19 Vaccination of Breastfeeding Women. JAMA 2021, 325, 2013–2014. [Google Scholar] [CrossRef]
- Scrimin, F.; Campisciano, G.; Comar, M.; Ragazzon, C.; Davanzo, R.; Quadrifoglio, M.; Giangreco, M.; Stabile, G.; Ricci, G. IgG and IgA Antibodies Post SARS-CoV-2 Vaccine in the Breast Milk and Sera of Breastfeeding Women. Vaccines 2022, 10, 125. [Google Scholar] [CrossRef]
- Rostad, C.A.; Chen, X.; Sun, H.Y.; Hussaini, L.; Lu, A.; Perez, M.A.; Hsiao, H.M.; Anderson, L.J.; Anderson, E.J. Functional Antibody Responses to Severe Acute Respiratory Syndrome Coronavirus 2 Variants in Children with Coronavirus Disease 2019, Multisystem Inflammatory Syndrome in Children, and After Two Doses of BNT162b2 Vaccination. J. Infect. Dis. 2022, 226, 1237–1242. [Google Scholar] [CrossRef]
- Chen, X.; Rostad, C.A.; Anderson, L.J.; Sun, H.Y.; Lapp, S.A.; Stephens, K.; Hussaini, L.; Gibson, T.; Rouphael, N.; Anderson, E.J. The development and kinetics of functional antibody-dependent cell-mediated cytotoxicity (ADCC) to SARS-CoV-2 spike protein. Virology 2021, 559, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Olearo, F.; Radmanesh, L.S.; Felber, N.; von Possel, R.; Emmerich, P.; Pekarek, N.; Pfefferle, S.; Norz, D.; Hansen, G.; Diemert, A.; et al. Anti-SARS-CoV-2 antibodies in breast milk during lactation after infection or vaccination: A cohort study. J. Reprod Immunol. 2022, 153, 103685. [Google Scholar] [CrossRef]
- Pace, R.M.; Williams, J.E.; Jarvinen, K.M.; Belfort, M.B.; Pace, C.D.W.; Lackey, K.A.; Gogel, A.C.; Nguyen-Contant, P.; Kanagaiah, P.; Fitzgerald, T.; et al. Characterization of SARS-CoV-2 RNA, Antibodies, and Neutralizing Capacity in Milk Produced by Women with COVID-19. mBio 2021, 12, e03192-20. [Google Scholar] [CrossRef] [PubMed]
- Young, B.E.; Seppo, A.E.; Diaz, N.; Rosen-Carole, C.; Nowak-Wegrzyn, A.; Cruz Vasquez, J.M.; Ferri-Huerta, R.; Nguyen-Contant, P.; Fitzgerald, T.; Sangster, M.Y.; et al. Association of Human Milk Antibody Induction, Persistence, and Neutralizing Capacity With SARS-CoV-2 Infection vs. mRNA Vaccination. JAMA Pediatr. 2022, 176, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Charepe, N.; Goncalves, J.; Juliano, A.M.; Lopes, D.G.; Canhao, H.; Soares, H.; Serrano, E.F. COVID-19 mRNA vaccine and antibody response in lactating women: A prospective cohort study. BMC Pregnancy Childbirth 2021, 21, 632. [Google Scholar] [CrossRef]
- Collier, A.Y.; McMahan, K.; Yu, J.; Tostanoski, L.H.; Aguayo, R.; Ansel, J.; Chandrashekar, A.; Patel, S.; Apraku Bondzie, E.; Sellers, D.; et al. Immunogenicity of COVID-19 mRNA Vaccines in Pregnant and Lactating Women. JAMA 2021, 325, 2370–2380. [Google Scholar] [CrossRef]
- Guida, M.; Terracciano, D.; Cennamo, M.; Aiello, F.; La Civita, E.; Esposito, G.; Gargiulo, V.; Maruotti, G.M.; Portella, G.; Sarno, L. COVID-19 Vaccine mRNABNT162b2 Elicits Human Antibody Response in Milk of Breastfeeding Women. Vaccines 2021, 9, 785. [Google Scholar] [CrossRef] [PubMed]
- Jakuszko, K.; Koscielska-Kasprzak, K.; Zabinska, M.; Bartoszek, D.; Poznanski, P.; Rukasz, D.; Klak, R.; Krolak-Olejnik, B.; Krajewska, M. Immune Response to Vaccination against COVID-19 in Breastfeeding Health Workers. Vaccines 2021, 9, 663. [Google Scholar] [CrossRef]
- Juncker, H.G.; Mulleners, S.J.; Coenen, E.R.M.; van Goudoever, J.B.; van Gils, M.J.; van Keulen, B.J. Comparing Human Milk Antibody Response After 4 Different Vaccines for COVID-19. JAMA Pediatr. 2022, 176, 611–612. [Google Scholar] [CrossRef] [PubMed]
- Juncker, H.G.; Mulleners, S.J.; van Gils, M.J.; de Groot, C.J.M.; Pajkrt, D.; Korosi, A.; van Goudoever, J.B.; van Keulen, B.J. The Levels of SARS-CoV-2 Specific Antibodies in Human Milk Following Vaccination. J. Hum. Lact. 2021, 37, 477–484. [Google Scholar] [CrossRef]
- Narayanaswamy, V.; Pentecost, B.T.; Schoen, C.N.; Alfandari, D.; Schneider, S.S.; Baker, R.; Arcaro, K.F. Neutralizing Antibodies and Cytokines in Breast Milk After Coronavirus Disease 2019 (COVID-19) mRNA Vaccination. Obstet. Gynecol. 2022, 139, 181–191. [Google Scholar] [CrossRef]
- Romero Ramírez, D.S.; Lara Pérez, M.M.; Carretero Pérez, M.; Suárez Hernández, M.I.; Martín Pulido, S.; Pera Villacampa, L.; Fernández Vilar, A.M.; Rivero Falero, M.; González Carretero, P.; Reyes Millán, B.; et al. SARS-CoV-2 Antibodies in Breast Milk After Vaccination. Pediatrics 2021, 148, e2021052286. [Google Scholar] [CrossRef] [PubMed]
- Yeo, K.T.; Chia, W.N.; Tan, C.W.; Ong, C.; Yeo, J.G.; Zhang, J.; Poh, S.L.; Lim, A.J.M.; Sim, K.H.Z.; Sutamam, N.; et al. Neutralizing Activity and SARS-CoV-2 Vaccine mRNA Persistence in Serum and Breastmilk After BNT162b2 Vaccination in Lactating Women. Front. Immunol. 2021, 12, 783975. [Google Scholar] [CrossRef] [PubMed]
- Golan, Y.; Ilala, M.; Li, L.; Gay, C.; Hunagund, S.; Lin, C.Y.; Cassidy, A.G.; Jigmeddagva, U.; Matsui, Y.; Ozarslan, N.; et al. Milk antibody response after 3(rd) COVID-19 vaccine and SARS-CoV-2 infection and implications for infant protection. iScience 2023, 26, 107767. [Google Scholar] [CrossRef]
- Szczygioł, P.; Łukianowski, B.; Kościelska-Kasprzak, K.; Jakuszko, K.; Bartoszek, D.; Krajewska, M.; Królak-Olejnik, B. Antibodies in the breastmilk of COVID-19 recovered women. BMC Pregnancy Childbirth 2022, 22, 635. [Google Scholar] [CrossRef]
- Selma-Royo, M.; Bauerl, C.; Mena-Tudela, D.; Aguilar-Camprubi, L.; Perez-Cano, F.J.; Parra-Llorca, A.; Lerin, C.; Martinez-Costa, C.; Collado, M.C. Anti-SARS-CoV-2 IgA and IgG in human milk after vaccination is dependent on vaccine type and previous SARS-CoV-2 exposure: A longitudinal study. Genome Med. 2022, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Perez, S.E.; Luna Centeno, L.D.; Cheng, W.A.; Marentes Ruiz, C.J.; Lee, Y.; Congrave-Wilson, Z.; Powell, R.L.; Stellwagen, L.; Pannaraj, P.S. Human Milk SARS-CoV-2 Antibodies up to 6 Months After Vaccination. Pediatrics 2022, 149, e2021054260. [Google Scholar] [CrossRef] [PubMed]
- Bauerl, C.; Zulaica, J.; Rusu, L.; Moreno, A.R.; Perez-Cano, F.J.; Lerin, C.; Mena-Tudela, D.; Aguilar-Camprubi, L.; Parra-Llorca, A.; Martinez-Costa, C.; et al. Assessment of SARS-CoV-2 neutralizing antibody titers in breastmilk from convalescent and vaccinated mothers. iScience 2023, 26, 106802. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas-Bernal, O.; Cervantes-Luevano, K.; Flores-Acosta, G.I.; Bernaldez-Sarabia, J.; Licea-Navarro, A.F. COVID-19 Neutralizing Antibodies in Breast Milk of Mothers Vaccinated with Three Different Vaccines in Mexico. Vaccines 2022, 10, 629. [Google Scholar] [CrossRef]
- Low, J.M.; Gu, Y.; Ng, M.S.F.; Wang, L.W.; Amin, Z.; Zhong, Y.; MacAry, P.A. Human Milk Antibodies after BNT162b2 Vaccination Exhibit Reduced Binding against SARS-CoV-2 Variants of Concern. Vaccines 2022, 10, 225. [Google Scholar] [CrossRef]
- Chatterjee, S.; Bhattacharya, M.; Nag, S.; Dhama, K.; Chakraborty, C. A Detailed Overview of SARS-CoV-2 Omicron: Its Sub-Variants, Mutations and Pathophysiology, Clinical Characteristics, Immunological Landscape, Immune Escape, and Therapies. Viruses 2023, 15, 167. [Google Scholar] [CrossRef]
- Schwartz, A.; Nir, O.; Toussia-Cohen, S.; Leibovich, L.; Strauss, T.; Asraf, K.; Doolman, R.; Sharabi, S.; Cohen, C.; Levin, E.G.; et al. Presence of SARS-CoV-2 antibodies in lactating women and their infants following BNT162b2 messenger RNA vaccine. Am. J. Obstet. Gynecol. 2021, 225, 577–579. [Google Scholar] [CrossRef]
- Breast Feeding Report Card. Available online: https://www.cdc.gov/breastfeeding/data/reportcard.htm#:~:text=At%206%20months%2C%2055.8%25%20of,breast%20milk%20at%206%20months (accessed on 22 August 2023).
- Regan, A.K.; Kaur, R.; Nosek, M.; Swathi, P.A.; Gu, N.Y. COVID-19 vaccine acceptance and coverage among pregnant persons in the United States. Prev. Med. Rep. 2022, 29, 101977. [Google Scholar] [CrossRef]
- Villar, J.; Ariff, S.; Gunier, R.B.; Thiruvengadam, R.; Rauch, S.; Kholin, A.; Roggero, P.; Prefumo, F.; do Vale, M.S.; Cardona-Perez, J.A.; et al. Maternal and Neonatal Morbidity and Mortality Among Pregnant Women with and without COVID-19 Infection: The INTERCOVID Multinational Cohort Study. JAMA Pediatr. 2021, 175, 817–826. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Mean (SD) [Range] or n (%) |
|---|---|
| Age, years |
|
Race
|
|
Ethnicity
|
|
| Pregnant at time of first dose |
|
| Gestational age at delivery, weeks |
|
Fetus/Infant age at first dose, weeks
|
|
Vaccine manufacturer
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiao, H.-M.; DiMaggio, L.S.; Perez, M.A.; Chen, X.; Stephens, K.; Gibson, T.; Anderson, E.J.; Rostad, C.A. SARS-CoV-2 Antibody Profiles in Maternal Serum and Breast Milk Following mRNA COVID-19 Vaccination: A Longitudinal Prospective Observational Cohort Study. Vaccines 2023, 11, 1643. https://doi.org/10.3390/vaccines11111643
Hsiao H-M, DiMaggio LS, Perez MA, Chen X, Stephens K, Gibson T, Anderson EJ, Rostad CA. SARS-CoV-2 Antibody Profiles in Maternal Serum and Breast Milk Following mRNA COVID-19 Vaccination: A Longitudinal Prospective Observational Cohort Study. Vaccines. 2023; 11(11):1643. https://doi.org/10.3390/vaccines11111643
Chicago/Turabian StyleHsiao, Hui-Mien, Langdon S. DiMaggio, Maria A. Perez, Xuemin Chen, Kathleen Stephens, Theda Gibson, Evan J. Anderson, and Christina A. Rostad. 2023. "SARS-CoV-2 Antibody Profiles in Maternal Serum and Breast Milk Following mRNA COVID-19 Vaccination: A Longitudinal Prospective Observational Cohort Study" Vaccines 11, no. 11: 1643. https://doi.org/10.3390/vaccines11111643
APA StyleHsiao, H.-M., DiMaggio, L. S., Perez, M. A., Chen, X., Stephens, K., Gibson, T., Anderson, E. J., & Rostad, C. A. (2023). SARS-CoV-2 Antibody Profiles in Maternal Serum and Breast Milk Following mRNA COVID-19 Vaccination: A Longitudinal Prospective Observational Cohort Study. Vaccines, 11(11), 1643. https://doi.org/10.3390/vaccines11111643