
mRNA COVID-19 Vaccines—Facts and Hypotheses on Fragmentation and Encapsulation



Abstract
1. Introduction
2. Background
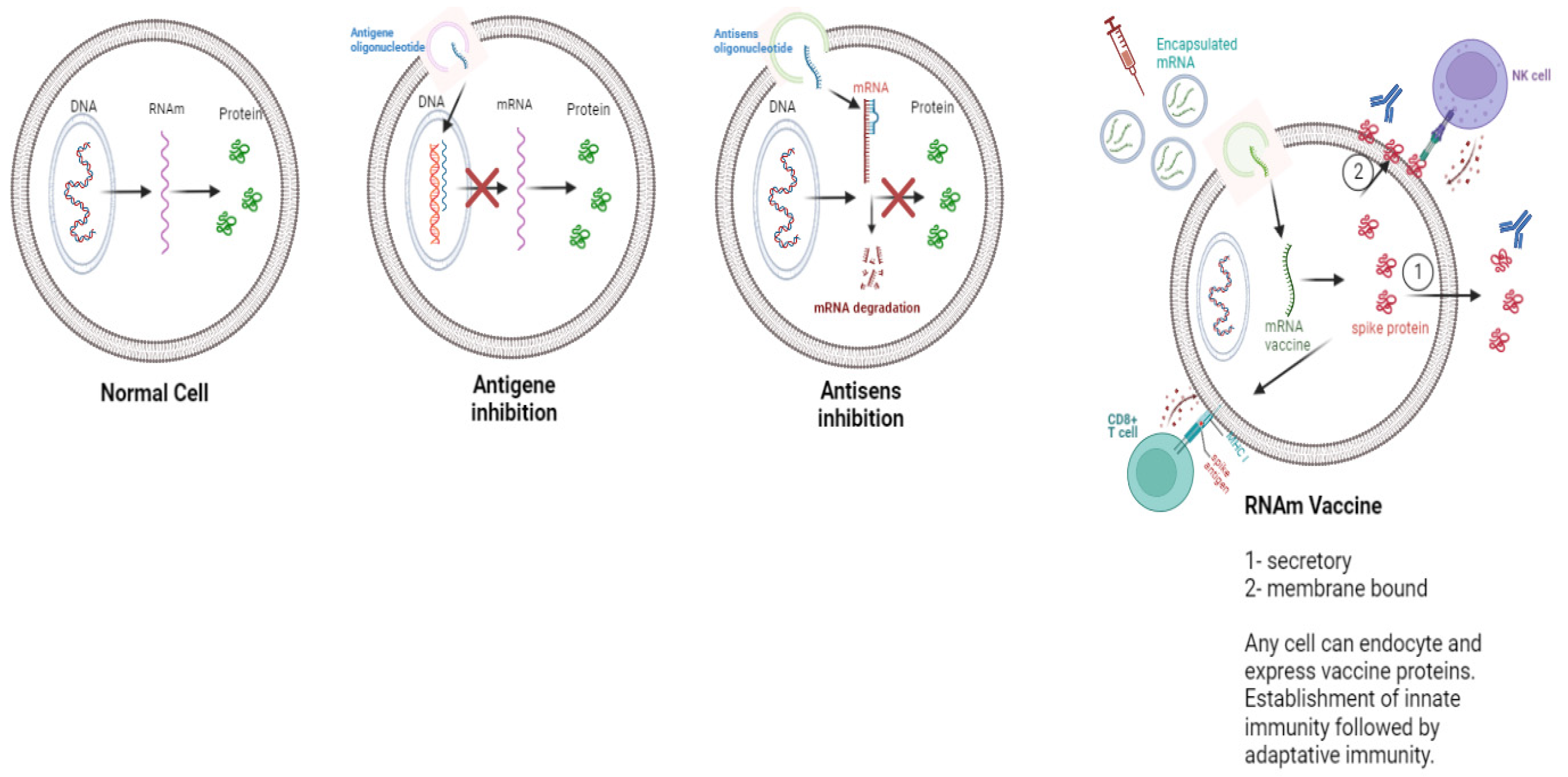
2.1. The Different RNA Strategies Considered in Therapy
2.2. Modifications in the mRNA Encoding the Spike Protein
2.3. Specific Modification of the mRNA Sequence of the Pfizer/BioNtech Vaccine®
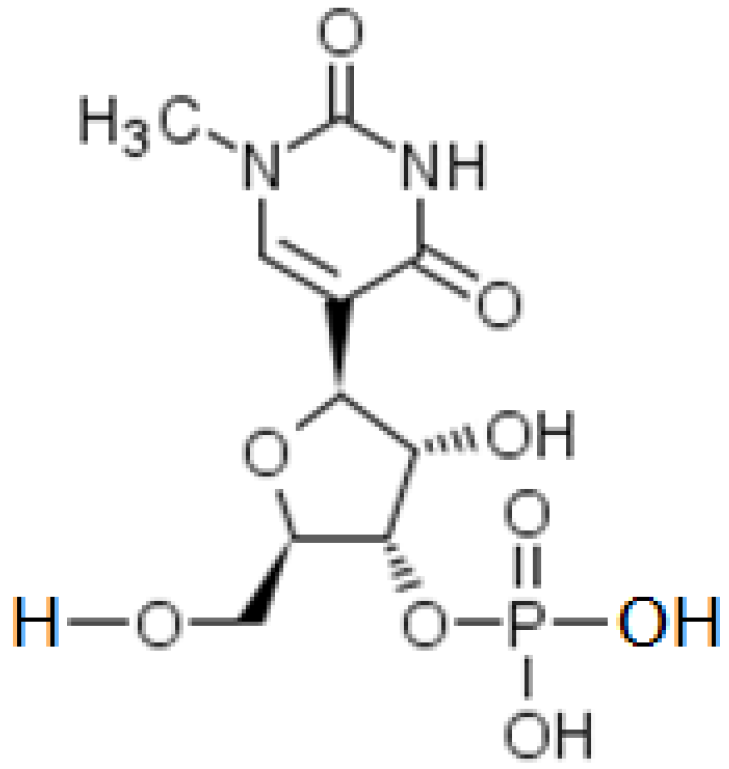
2.4. Substitution of Uridine by N1-Methyl-3′-pseudouridine
2.5. Modification of Codons and Substitution of 2 Amino Acids
2.6. mRNA and Poly(A) Tail Size
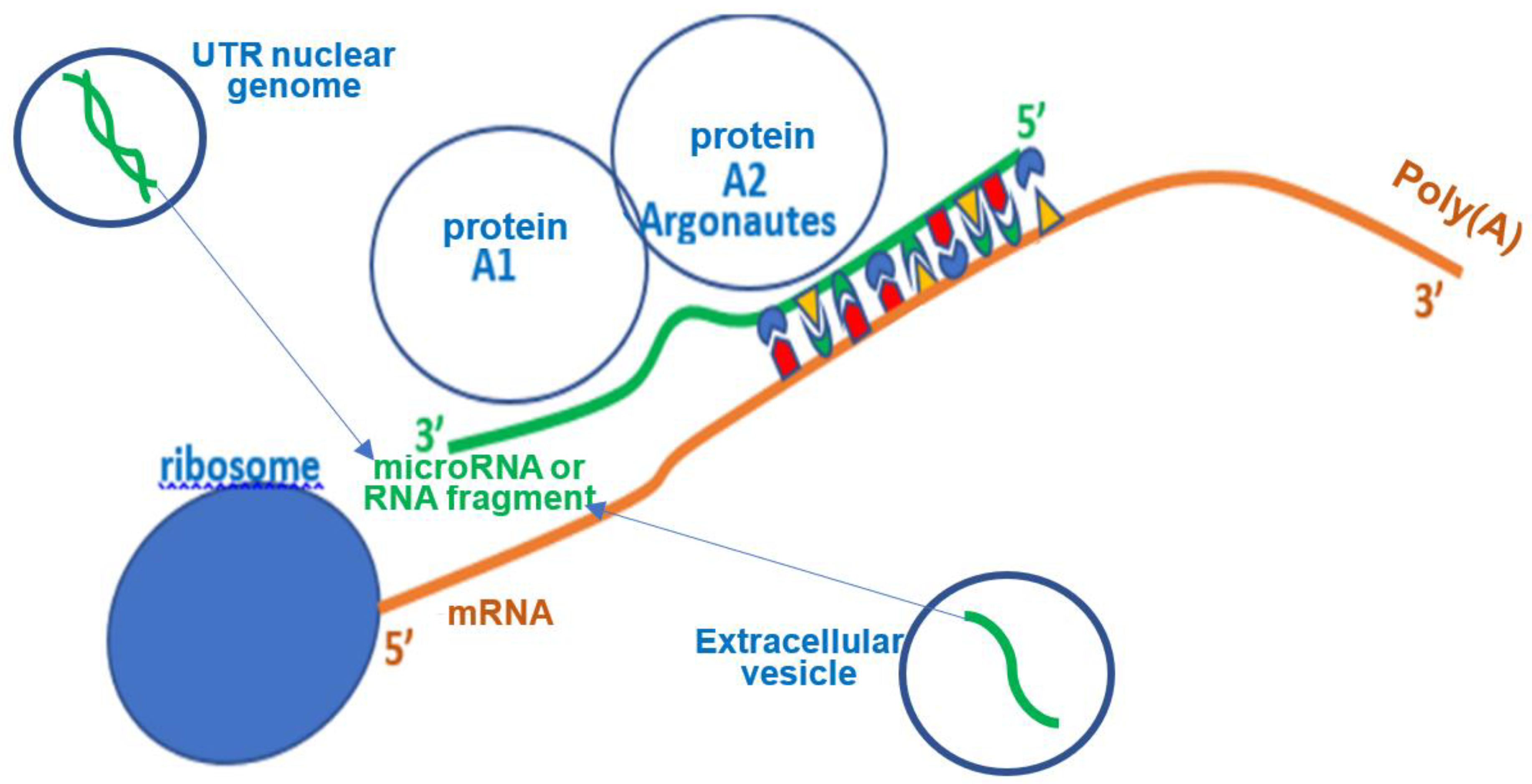
3. Micro-RNAs and Their Role
4. Results
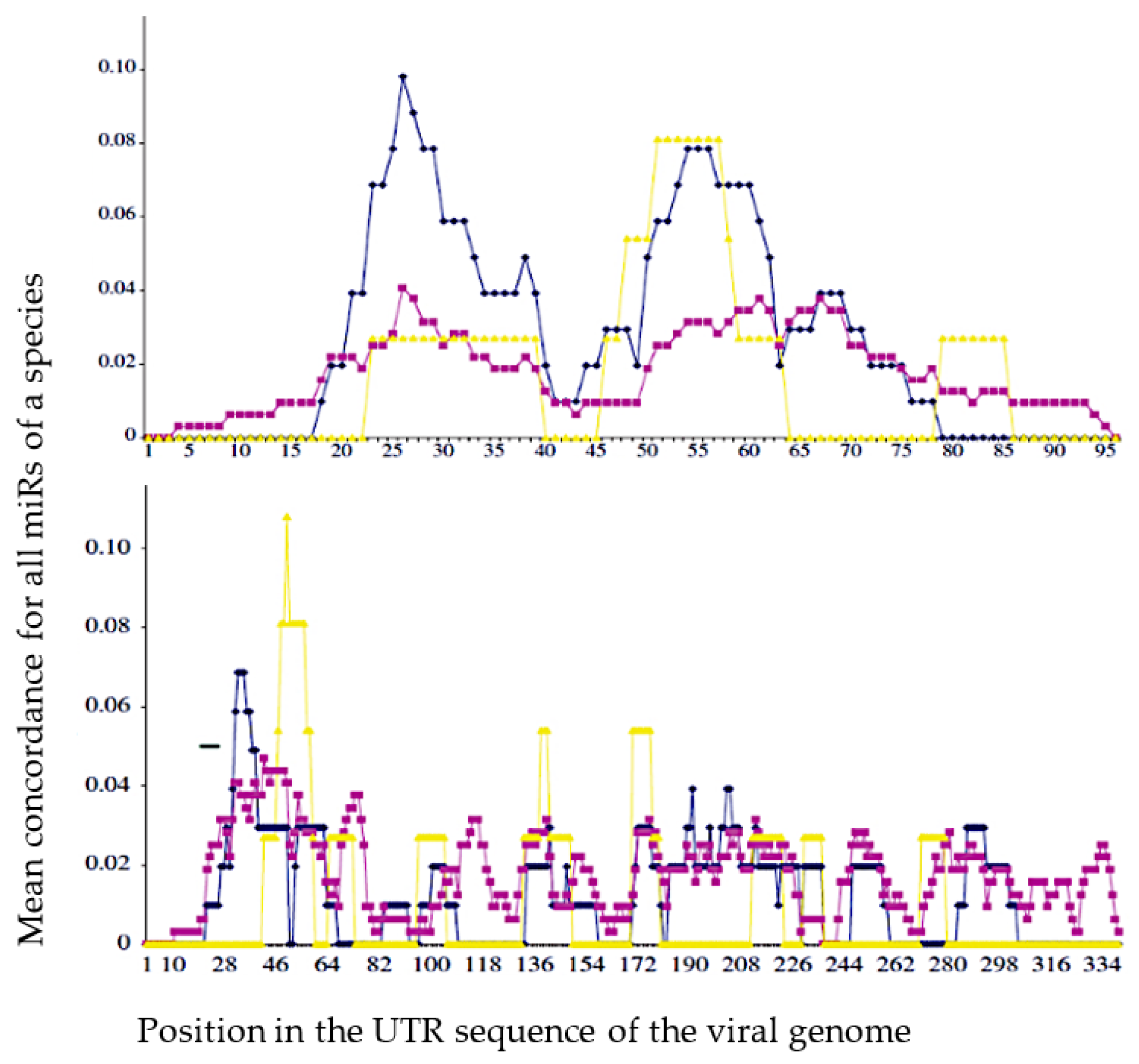
4.1. Viral and Host Genomes
4.2. Hybridizing Power of SARS-CoV-2 Viral RNA Fragments
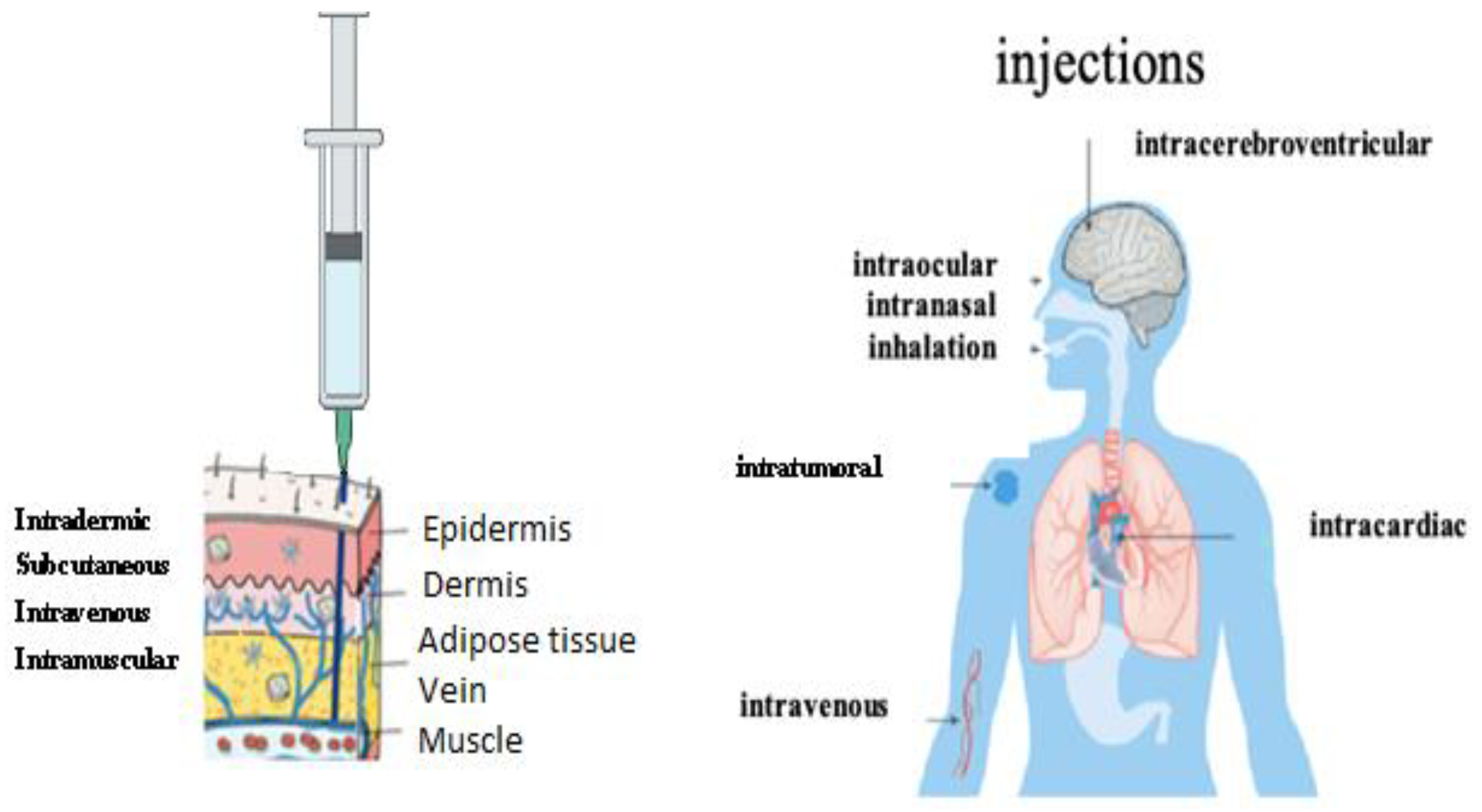
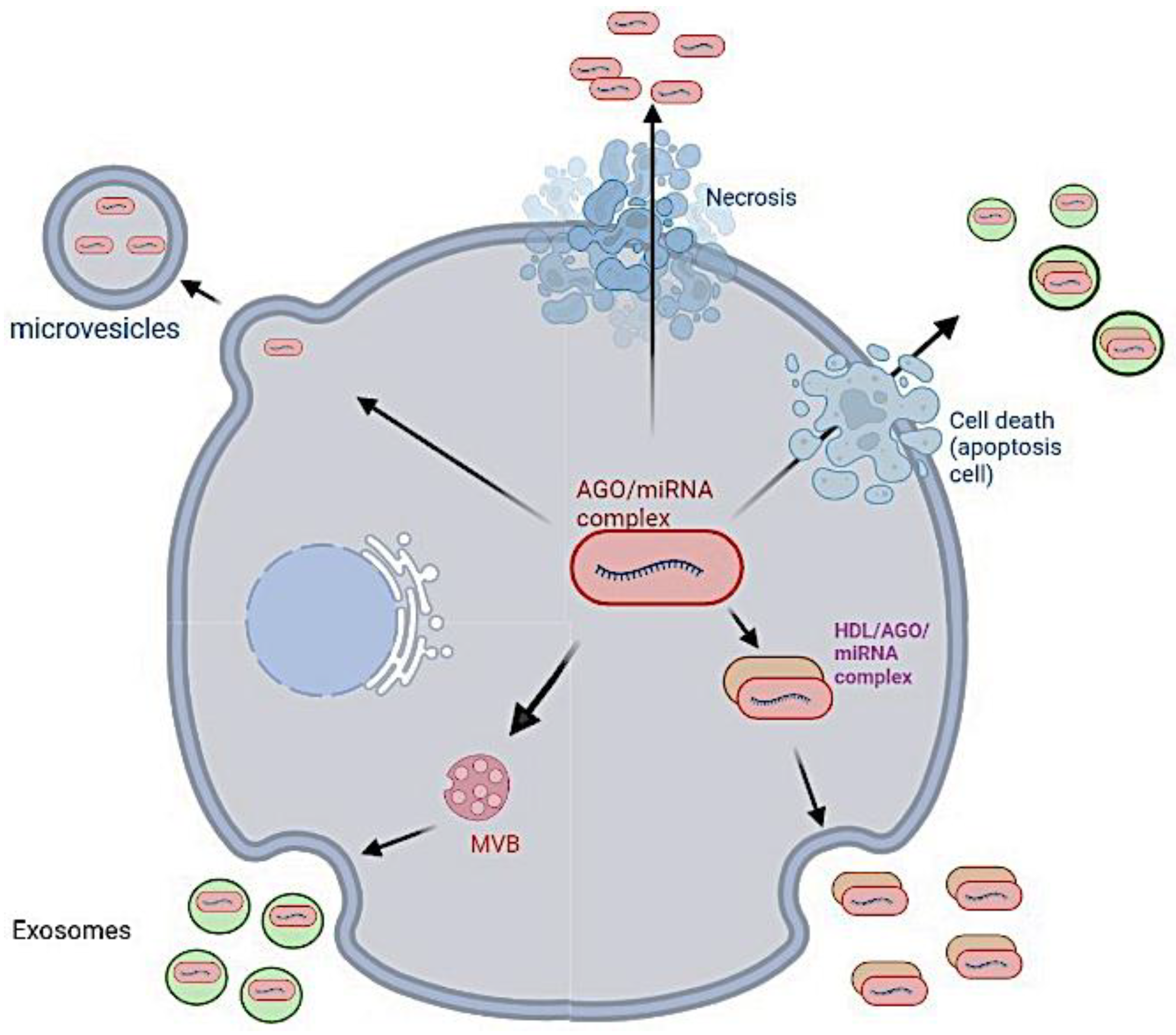
5. Biodistribution of mRNA Vaccine
6. Goal of Vectorization
- It must be easy to produce on an industrial scale by simple and safe methods. It should also be as inexpensive as possible.
- It must be non-immunogenic and thus allow repeated administration.
- It must be specific for the targeted cells.
- It must be able to adapt to the size of the oligonucleotide to be transported.
- It must be internalized by the cells and allow the release of the oligonucleotide into the cell (transfection phenomenon).
- It must break down and be easily eliminated from the body.
6.1. Differences between “Classic” Vaccines and New mRNA Vaccines Encapsulated in a Nanoparticle Liposome
6.2. Liposome Physical Properties
6.3. Liposome Chemistry Properties (LNP)

6.4. LNP Safety Profile
6.5. Conditional Marketing Authorization for the Pfizer/BioNtech® Vaccine [137]
6.6. Conditional Marketing Authorization for the Moderna® Vaccine [138]
7. Discussion
7.1. How to Overcome Some Obstacles in the Development of mRNA Vaccines for Diminishing the Adverse Effects
7.2. Optimizing the Development of Vaccines
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cobb, M. Who Discovered Messenger RNA? Curr. Biol. 2015, 25, R526–R532. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. MRNA Vaccines—A New Era in Vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Hajj, K.A.; Whitehead, K.A. Tools for Translation: Non-Viral Materials for Therapeutic MRNA Delivery. Nat. Rev. Mater. 2017, 2, 17056. [Google Scholar] [CrossRef]
- Xiong, Q.; Lee, G.Y.; Ding, J.; Li, W.; Shi, J. Biomedical Applications of MRNA Nanomedicine. Nano Res. 2018, 11, 5281–5309. [Google Scholar] [CrossRef]
- Guan, S.; Rosenecker, J. Nanotechnologies in Delivery of MRNA Therapeutics Using Nonviral Vector-Based Delivery Systems. Gene Ther. 2017, 24, 133–143. [Google Scholar] [CrossRef]
- Kowalski, P.S.; Rudra, A.; Miao, L.; Anderson, D.G. Delivering the Messenger: Advances in Technologies for Therapeutic MRNA Delivery. Mol. Ther. 2019, 27, 710–728. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Dong, Y. Nanoscale Platforms for Messenger RNA Delivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1530. [Google Scholar] [CrossRef]
- Zhao, W.; Hou, X.; Vick, O.G.; Dong, Y. RNA Delivery Biomaterials for the Treatment of Genetic and Rare Diseases. Biomaterials 2019, 217, 119291. [Google Scholar] [CrossRef] [PubMed]
- Uchida, S.; Perche, F.; Pichon, C.; Cabral, H. Nanomedicine-Based Approaches for MRNA Delivery. Mol. Pharm. 2020, 17, 3654–3684. [Google Scholar] [CrossRef]
- Meng, C.; Chen, Z.; Li, G.; Welte, T.; Shen, H. Nanoplatforms for MRNA Therapeutics. Adv. Ther. 2021, 4, 2000099. [Google Scholar] [CrossRef]
- Gebre, M.S.; Brito, L.A.; Tostanoski, L.H.; Edwards, D.K.; Carfi, A.; Barouch, D.H. Novel Approaches for Vaccine Development. Cell 2021, 184, 1589–1603. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Li, C.; Yang, T.; Hu, B.; Zhang, M.; Guo, S.; Xiao, H.; Liang, X.-J.; Huang, Y. The Challenge and Prospect of MRNA Therapeutics Landscape. Biotechnol. Adv. 2020, 40, 107534. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Eygeris, Y.; Gupta, M.; Sahay, G. Self-Assembled MRNA Vaccines. Adv. Drug Deliv. Rev. 2021, 170, 83–112. [Google Scholar] [CrossRef] [PubMed]
- Cazenave, C.; Frank, P.; Büsen, W. Characterization of Ribonuclease H Activities Present in Two Cell-Free Protein Synthesizing Systems, the Wheat Germ Extract and the Rabbit Reticulocyte Lysate. Biochimie 1993, 75, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Krishnan, S.; Lenzen, G.; Magné, R.; Gomard, E.; Guillet, J.G.; Lévy, J.P.; Meulien, P. Induction of Virus-Specific Cytotoxic T Lymphocytes in Vivo by Liposome-Entrapped MRNA. Eur. J. Immunol. 1993, 23, 1719–1722. [Google Scholar] [CrossRef]
- Akinc, A.; Maier, M.A.; Manoharan, M.; Fitzgerald, K.; Jayaraman, M.; Barros, S.; Ansell, S.; Du, X.; Hope, M.J.; Madden, T.D.; et al. The Onpattro Story and the Clinical Translation of Nanomedicines Containing Nucleic Acid-Based Drugs. Nat. Nanotechnol. 2019, 14, 1084–1087. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 MRNA-1273 Vaccine in Older Adults. N. Engl. J. Med. 2020, 383, 2427–2438. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Belikova, A.M.; Zarytova, V.F.; Grineva, N.I. Synthesis of Ribonucleosides and Diribonucleoside Phosphates Containing 2-Chloroethylamine and Nitrogen Mustard Residues. Tetrahedron Lett. 1967, 37, 3557–3562. [Google Scholar] [CrossRef]
- Stephenson, M.L.; Zamecnik, P.C. Inhibition of Rous Sarcoma Viral RNA Translation by a Specific Oligodeoxyribonucleotide. Proc. Natl. Acad. Sci. USA 1978, 75, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Ros, T.D.; Spalluto, G.; Prato, M.; Saison-Behmoaras, T.; Boutorine, A.; Cacciari, B. Oligonucleotides and Oligonucleotide Conjugates: A New Approach for Cancer Treatment. Curr. Med. Chem. 2005, 12, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.E.; Egholm, M.; Berg, R.H.; Buchardt, O. Sequence-Selective Recognition of DNA by Strand Displacement with a Thymine-Substituted Polyamide. Science 1991, 254, 1497–1500. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, E. RNA targeting using peptide nucleic acid. In RNA Towards Medicine; Springer: Berlin/Heidelberg, Germany, 2006; pp. 395–403. [Google Scholar] [CrossRef]
- Nielsen, P.E. Peptide Nucleic Acids as Therapeutic Agents. Curr. Opin. Struct. Biol. 1999, 9, 353–357. [Google Scholar] [CrossRef]
- Nielsen, P.E. Antisense Peptide Nucleic Acids. Curr. Opin. Mol. Ther. 2000, 2, 282–287. [Google Scholar]
- Nielsen, P.E. Peptide Nucleic Acid (PNA) From DNA Recognition to Antisense and DNA Structure. Biophys. Chem. 1997, 68, 103–108. [Google Scholar] [CrossRef]
- Nielsen, P.E. Applications of Peptide Nucleic Acids. Curr. Opin. Biotechnol. 1999, 10, 71–75. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 Spike Receptor-Binding Domain Bound to the ACE2 Receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural Basis for the Recognition of SARS-CoV-2 by Full-Length Human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 Facilitates SARS-CoV-2 Cell Entry and Infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- El-Arif, G.; Farhat, A.; Khazaal, S.; Annweiler, C.; Kovacic, H.; Wu, Y.; Cao, Z.; Fajloun, Z.; Khattar, Z.A.; Sabatier, J.M. The Renin-Angiotensin System: A Key Role in SARS-CoV-2-Induced COVID-19. Molecules 2021, 26, 6945. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Messenger RNA Encoding the Full-Length SARS-CoV-2 Spike Glycoprotein; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Nelson, J.; Sorensen, E.W.; Mintri, S.; Rabideau, A.E.; Zheng, W.; Besin, G.; Khatwani, N.; Su, S.V.; Miracco, E.J.; Issa, W.J.; et al. Impact of MRNA Chemistry and Manufacturing Process on Innate Immune Activation. Sci. Adv. 2020, 6, eaaz6893. [Google Scholar] [CrossRef] [PubMed]
- Pallesen, J.; Wang, N.; Corbett, K.S.; Wrapp, D.; Kirchdoerfer, R.N.; Turner, H.L.; Cottrell, C.A.; Becker, M.M.; Wang, L.; Shi, W.; et al. Immunogenicity and Structures of a Rationally Designed Prefusion MERS-CoV Spike Antigen. Proc. Natl. Acad. Sci. USA 2017, 114, E7348–E7357. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, M.W.; Thornton, J.M. Influence of Proline Residues on Protein Conformation. J. Mol. Biol. 1991, 218, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.M.; Hardré, R.; Edwards, P.J.B. The Impact of Pyrrolidine Hydroxylation on the Conformation of Proline-Containing Peptides. J. Org. Chem. 2005, 70, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Kudla, G.; Lipinski, L.; Caffin, F.; Helwak, A.; Zylicz, M. High Guanine and Cytosine Content Increases MRNA Levels in Mammalian Cells. PLoS Biol. 2006, 4, e180. [Google Scholar] [CrossRef]
- Kim, S.C.; Sekhon, S.S.; Shin, W.-R.; Ahn, G.; Cho, B.-K.; Ahn, J.-Y.; Kim, Y.-H. Modifications of MRNA Vaccine Structural Elements for Improving MRNA Stability and Translation Efficiency. Mol. Cell Toxicol. 2022, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Körner, C.G.; Wahle, E. Poly(A) Tail Shortening by a Mammalian Poly(A)-Specific 3′-Exoribonuclease. J. Biol. Chem. 1997, 272, 10448–10456. [Google Scholar] [CrossRef]
- Schlake, T.; Thess, A.; Fotin-Mleczek, M.; Kallen, K.-J. Developing MRNA-Vaccine Technologies. RNA Biol. 2012, 9, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Elango, N.; Elango, S.; Shivshankar, P.; Katz, M.S. Optimized Transfection of MRNA Transcribed from a d(A/T)100 Tail-Containing Vector. Biochem. Biophys. Res. Commun. 2005, 330, 958–966. [Google Scholar] [CrossRef]
- Demongeot, J.; Drouet, E.; Elena, A.; Moreira, A.; Rechoum, Y.; Sené, S. Micro-RNAs: Viral Genome and Robustness of Gene Expression in the Host. Philos. Trans. Math. Phys. Eng. Sci. 2009, 367, 4941–4965. [Google Scholar] [CrossRef] [PubMed]
- Gardes, J.; Maldivi, C.; Boisset, D.; Aubourg, T.; Vuillerme, N.; Demongeot, J. Maxwell®: An unsupervised learning approach for 5P medicine. In MEDINFO 2019: Health and Wellbeing e-Networks for All; IOS Press: Amsterdam, The Netherlands, 2019; pp. 1464–1465. [Google Scholar] [CrossRef]
- Mindell, D.P. Viruses and the Evolution of Life. By Luis P Villarreal. Q. Rev. Biol. 2005, 80, 478–479. [Google Scholar] [CrossRef]
- Pfeffer, S.; Sewer, A.; Lagos-Quintana, M.; Sheridan, R.; Sander, C.; Grässer, F.A.; van Dyk, L.F.; Ho, C.K.; Shuman, S.; Chien, M.; et al. Identification of MicroRNAs of the Herpesvirus Family. Nat. Methods 2005, 2, 269–276. [Google Scholar] [CrossRef]
- Kincaid, R.P.; Burke, J.M.; Sullivan, C.S. RNA Virus MicroRNA That Mimics a B-Cell OncomiR. Proc. Natl. Acad. Sci. USA 2012, 109, 3077–3082. [Google Scholar] [CrossRef] [PubMed]
- Cullen, B.R. Viruses and MicroRNAs. Nat. Genet. 2006, 38, S25–S30. [Google Scholar] [CrossRef]
- Yekta, S.; Shih, I.-H.; Bartel, D.P. MicroRNA-Directed Cleavage of HOXB8 MRNA. Science 2004, 304, 594–596. [Google Scholar] [CrossRef]
- Brennecke, J.; Stark, A.; Russell, R.B.; Cohen, S.M. Principles of MicroRNA–Target Recognition. PLoS Biol. 2005, 3, e85. [Google Scholar] [CrossRef]
- Lee Marzano, S.-Y.; Neupane, A.; Domier, L. Transcriptional and Small RNA Responses of the White Mold Fungus Sclerotinia Sclerotiorum to Infection by a Virulence-Attenuating Hypovirus. Viruses 2018, 10, 713. [Google Scholar] [CrossRef]
- Sakuma, T.; Tonne, J.M.; Squillace, K.A.; Ohmine, S.; Thatava, T.; Peng, K.-W.; Barry, M.A.; Ikeda, Y. Early Events in Retrovirus XMRV Infection of the Wild-Derived Mouse Mus Pahari. J. Virol. 2011, 85, 1205–1213. [Google Scholar] [CrossRef]
- H5N1 Influenza Virus-Specific MiRNA-like Small RNA Increases Cytokine Production and Mouse Mortality via Targeting Poly(RC)-Binding Protein 2. Cell Research. Available online: https://www.nature.com/articles/cr20183 (accessed on 15 November 2022).
- Bandiera, S.; Matégot, R.; Girard, M.; Demongeot, J.; Henrion-Caude, A. MitomiRs Delineating the Intracellular Localization of MicroRNAs at Mitochondria. Free. Radic. Biol. Med. 2013, 64, 12–19. [Google Scholar] [CrossRef]
- Demongeot, J.; Hazgui, H.; Bandiera, S.; Cohen, O.; Henrion-Caude, A. MitomiRs, ChloromiRs and Modelling of the MicroRNA Inhibition. Acta Biotheor. 2013, 61, 367–383. [Google Scholar] [CrossRef]
- Demongeot, J.; Cohen, O.; Henrion-Caude, A. MicroRNAs and robustness in biological regulatory networks. A Generic approach with applications at different levels: Physiologic metabolic and genetic. In Systems Biology of Metabolic and Signaling Networks; Springer: Berlin/Heidelberg, Germany, 2013; pp. 63–114. [Google Scholar]
- Demongeot, J.; Hazgui, H.; Escoffier, J.; Arnoult, C. Inhibitory Regulation by MicroRNAs and Circular RNAs. In XIII Mediterranean Conference on Medical and Biological Engineering and Computing 2013; Roa Romero, L.M., Ed.; Springer International Publishing: Cham, Switzerland, 2014; pp. 1250–1253. [Google Scholar]
- Demongeot, J.; Henrion-Caude, A. Footprints of a Singular 22-Nucleotide RNA Ring at the Origin of Life. Biology 2020, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Seligmann, H. Comparisons between Small Ribosomal RNA and Theoretical Minimal RNA Ring Secondary Structures Confirm Phylogenetic and Structural Accretion Histories. Sci. Rep. 2020, 10, 7693. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Seligmann, H. SARS-CoV-2 and MiRNA-like Inhibition Power. Med. Hypotheses 2020, 144, 110245. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.-Y.; Chen, M.; Wang, C.-D. Annotation of MiRNAs in COVID-19 Coronavirus. J. Electron. Sci. Technol. 2021, 19, 100060. [Google Scholar] [CrossRef]
- Mirzaei, R.; Mahdavi, F.; Badrzadeh, F.; Hosseini-Fard, S.R.; Heidary, M.; Jeda, A.S.; Mohammadi, T.; Roshani, M.; Yousefimashouf, R.; Keyvani, H.; et al. The Emerging Role of MicroRNAs in the Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection. Int. Immunopharmacol. 2021, 90, 107204. [Google Scholar] [CrossRef]
- Demirci, M.D.S.; Adan, A. Computational Analysis of MicroRNA-Mediated Interactions in SARS-CoV-2 Infection. PeerJ 2020, 8, e9369. [Google Scholar] [CrossRef] [PubMed]
- Abedi, F.; Rezaee, R.; Hayes, A.W.; Nasiripour, S.; Karimi, G. MicroRNAs and SARS-CoV-2 Life Cycle, Pathogenesis, and Mutations: Biomarkers or Therapeutic Agents? Cell Cycle 2021, 20, 143–153. [Google Scholar] [CrossRef]
- Gasparello, J.; Finotti, A.; Gambari, R. Tackling the COVID-19 “Cytokine Storm” with MicroRNA Mimics Directly Targeting the 3′UTR of pro-Inflammatory MRNAs. Med. Hypotheses 2021, 146, 110415. [Google Scholar] [CrossRef]
- Chauhan, N.; Jaggi, M.; Chauhan, S.C.; Yallapu, M.M. COVID-19: Fighting the Invisible Enemy with MicroRNAs. Expert Rev. Anti Infect. Ther. 2021, 19, 137–145. [Google Scholar] [CrossRef]
- Cellular Nanosponges Inhibit SARS-CoV-2 Infectivity. Nano Letters. Available online: https://pubs.acs.org/doi/full/10.1021/acs.nanolett.0c02278 (accessed on 15 November 2022).
- Nersisyan, S.; Engibaryan, N.; Gorbonos, A.; Kirdey, K.; Makhonin, A.; Tonevitsky, A. Potential Role of Cellular MiRNAs in Coronavirus-Host Interplay. PeerJ 2020, 8, e9994. [Google Scholar] [CrossRef] [PubMed]
- Satyam, R.; Bhardwaj, T.; Goel, S.; Jha, N.K.; Jha, S.K.; Nand, P.; Ruokolainen, J.; Kamal, M.A.; Kesari, K.K. MiRNAs in SARS-CoV 2: A Spoke in the Wheel of Pathogenesis. Curr. Pharm. Des. 2021, 27, 1628–1641. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.-A.-K.; Sany, M.R.U.; Islam, M.S.; Islam, A.B.M.M.K. Epigenetic Regulator MiRNA Pattern Differences Among SARS-CoV, SARS-CoV-2, and SARS-CoV-2 World-Wide Isolates Delineated the Mystery Behind the Epic Pathogenicity and Distinct Clinical Characteristics of Pandemic COVID-19. Front. Genet. 2020, 11, 765. [Google Scholar] [CrossRef] [PubMed]
- Bozgeyik, I. Therapeutic Potential of MiRNAs Targeting SARS-CoV-2 Host Cell Receptor ACE2. Meta Gene 2021, 27, 100831. [Google Scholar] [CrossRef] [PubMed]
- Faghih Dinevari, M.; Somi, M.H.; Sadeghi Majd, E.; Abbasalizad Farhangi, M.; Nikniaz, Z. Anemia Predicts Poor Outcomes of COVID-19 in Hospitalized Patients: A Prospective Study in Iran. BMC Infect. Dis. 2021, 21, 170. [Google Scholar] [CrossRef] [PubMed]
- Anai, M.; Akaike, K.; Iwagoe, H.; Akasaka, T.; Higuchi, T.; Miyazaki, A.; Naito, D.; Tajima, Y.; Takahashi, H.; Komatsu, T.; et al. Decrease in Hemoglobin Level Predicts Increased Risk for Severe Respiratory Failure in COVID-19 Patients with Pneumonia. Respir. Investig. 2021, 59, 187–193. [Google Scholar] [CrossRef]
- Li, D.; Dong, H.; Li, S.; Munir, M.; Chen, J.; Luo, Y.; Sun, Y.; Liu, L.; Qiu, H.J. Hemoglobin Subunit Beta Interacts with the Capsid Protein and Antagonizes the Growth of Classical Swine Fever Virus. J. Virol. 2013, 87, 5707–5717. [Google Scholar] [CrossRef]
- Read, R. Flawed Methods in “COVID-19: Attacks the 1-Beta Chain of Hemoglobin and Captures the Porphyrin to Inhibit Human Heme Metabolism”. ChemRxiv 2020. [Google Scholar] [CrossRef]
- McCarthy, K.R.; Rennick, L.J.; Nambulli, S.; Robinson-McCarthy, L.R.; Bain, W.G.; Haidar, G.; Duprex, W.P. Recurrent Deletions in the SARS-CoV-2 Spike Glycoprotein Drive Antibody Escape. Science 2021, 371, 1139–1142. [Google Scholar] [CrossRef]
- Choudhury, Y.; Cher, C.Y.; Wan, Z.Y.; Xie, C.; Lim, J.S.; Virk, R.K.; Tan, M.H.; Teo, A.K.J.; Hsu, L.Y. A Viral Fragmentation Signature for SARS-CoV-2 in Clinical Samples Correlating with Contagiousness. MedRxiv 2021. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence That D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827.e19. [Google Scholar] [CrossRef] [PubMed]
- Andreano, E.; Piccini, G.; Licastro, D.; Casalino, L.; Johnson, N.V.; Paciello, I.; Dal Monego, S.; Pantano, E.; Manganaro, N.; Manenti, A.; et al. SARS-CoV-2 Escape from a Highly Neutralizing COVID-19 Convalescent Plasma. Proc. Natl. Acad. Sci. USA 2021, 118, e2103154118. [Google Scholar] [CrossRef] [PubMed]
- Wise, J. COVID-19: New Coronavirus Variant Is Identified in UK. BMJ 2020, 371, m4857. [Google Scholar] [CrossRef]
- Iacobucci, G. COVID-19: Surge Testing for the South Africa Variant Begins in England. BMJ 2021, 372, n305. [Google Scholar] [CrossRef]
- Fournier, P.-E.; Colson, P.; Levasseur, A.; Devaux, C.A.; Gautret, P.; Bedotto, M.; Delerce, J.; Brechard, L.; Pinault, L.; Lagier, J.-C.; et al. Emergence and Outcomes of the SARS-CoV-2 ‘Marseille-4′ Variant. Int. J. Infect. Dis. 2021, 106, 228–236. [Google Scholar] [CrossRef]
- Hadfield, J.; Megill, C.; Bell, S.M.; Huddleston, J.; Potter, B.; Callender, C.; Sagulenko, P.; Bedford, T.; Neher, R.A. Nextstrain: Real-Time Tracking of Pathogen Evolution. Bioinformatics 2018, 34, 4121–4123. [Google Scholar] [CrossRef]
- Finnegan, E.J.; Matzke, M.A. The Small RNA World. J. Cell Sci. 2003, 116, 4689–4693. [Google Scholar] [CrossRef]
- Wang, X. Composition of Seed Sequence Is a Major Determinant of MicroRNA Targeting Patterns. Bioinformatics 2014, 30, 1377–1383. [Google Scholar] [CrossRef]
- Broughton, J.P.; Lovci, M.T.; Huang, J.L.; Yeo, G.W.; Pasquinelli, A.E. Pairing beyond the Seed Supports MicroRNA Targeting Specificity. Mol. Cell 2016, 64, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Gebetsberger, J.; Wyss, L.; Mleczko, A.M.; Reuther, J.; Polacek, N. A TRNA-Derived Fragment Competes with MRNA for Ribosome Binding and Regulates Translation during Stress. RNA Biol. 2017, 14, 1364–1373. [Google Scholar] [CrossRef]
- NCBI. Nucleotide. Available online: https://www.ncbi.nlm.nih.gov/nuccore (accessed on 15 November 2022).
- GeneCards. HBB Gene—Hemoglobin Subunit Beta. Available online: https://www.genecards.org/cgi-bin/carddisp.pl?gene=HBB (accessed on 16 November 2022).
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From MicroRNA Sequences to Function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Xu, P.; Palmer, L.E.; Lechauve, C.; Zhao, G.; Yao, Y.; Luan, J.; Vourekas, A.; Tan, H.; Peng, J.; Schuetz, J.D.; et al. Regulation of Gene Expression by MiR-144/451 during Mouse Erythropoiesis. Blood 2019, 133, 2518–2528. [Google Scholar] [CrossRef]
- ChangLin, W.; JianCheng, X.; XinTang, D. Detection of miR-144 gene in peripheral blood of children with β-thalassemia major and its significance. China Trop. Med. 2010, 10, 285–286. [Google Scholar]
- Saki, N.; Abroun, S.; Soleimani, M.; Kavianpour, M.; Shahjahani, M.; Mohammadi-Asl, J.; Hajizamani, S. MicroRNA Expression in β-Thalassemia and Sickle Cell Disease: A Role in The Induction of Fetal Hemoglobin. Cell J. 2016, 17, 583–592. [Google Scholar] [CrossRef]
- Lai, K.; Jia, S.; Yu, S.; Luo, J.; He, Y. Genome-Wide Analysis of Aberrantly Expressed LncRNAs and MiRNAs with Associated Co-Expression and CeRNA Networks in β-Thalassemia and Hereditary Persistence of Fetal Hemoglobin. Oncotarget 2017, 8, 49931–49943. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, S.; Kaviani, S.; Soleimani, M.; Kouhkan, F.; Pourfathollah, A.A.; Amirizadeh, N.; Abroun, S.; Noruzinia, M. Mir-155 Downregulation by MiRCURY LNATM MicroRNA Inhibitor Can Increase Alpha Chain Hemoglobins Expression in Erythroleukemic K562 Cell Line. Int. J. Hematol. Oncol. Stem Cell Res. 2010, 4, 4–9. [Google Scholar]
- Farkas, C.; Mella, A.; Turgeon, M.; Haigh, J.J. A Novel SARS-CoV-2 Viral Sequence Bioinformatic Pipeline Has Found Genetic Evidence That the Viral 3′ Untranslated Region (UTR) Is Evolving and Generating Increased Viral Diversity. Front. Microbiol. 2021, 12, 665041. [Google Scholar] [CrossRef]
- Nishiura, H.; Ito, K.; Anzai, A.; Kobayashi, T.; Piantham, C.; Rodríguez-Morales, A.J. Relative Reproduction Number of SARS-CoV-2 Omicron (B.1.1.529) Compared with Delta Variant in South Africa. J. Clin. Med. 2021, 11, 30. [Google Scholar] [CrossRef]
- Østergaard, S.D.; Schmidt, M.; Horváth-Puhó, E.; Thomsen, R.W.; Sørensen, H.T. Thromboembolism and the Oxford-AstraZeneca COVID-19 Vaccine: Side-Effect or Coincidence? Lancet 2021, 397, 1441–1443. [Google Scholar] [CrossRef]
- Challen, R.; Brooks-Pollock, E.; Read, J.M.; Dyson, L.; Tsaneva-Atanasova, K.; Danon, L. Risk of Mortality in Patients Infected with SARS-CoV-2 Variant of Concern 202012/1: Matched Cohort Study. BMJ 2021, 372, n579. [Google Scholar] [CrossRef]
- Ganesan, S.K.; Venkatratnam, P.; Mahendra, J.; Devarajan, N. Increased Mortality of COVID-19 Infected Diabetes Patients: Role of Furin Proteases. Int. J. Obes. 2020, 44, 2486–2488. [Google Scholar] [CrossRef] [PubMed]
- Laderoute, M.P.; Larocque, L.J.; Giulivi, A.; Diaz-Mitoma, F. Further Evidence That Human Endogenous Retrovirus K102 Is a Replication Competent Foamy Virus That May Antagonize HIV-1 Replication. Open AIDS J. 2015, 9, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Holdt, L.M.; Kohlmaier, A.; Teupser, D. Circular RNAs as Therapeutic Agents and Targets. Front. Physiol. 2018, 9, 1262. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, F.; Wu, W.; Zhang, Y.; Chang, W.; Ponnusamy, M.; Wang, K.; Li, P. Circular RNAs: A Novel Type of Non-Coding RNA and Their Potential Implications in Antiviral Immunity. Int. J. Biol. Sci. 2017, 13, 1497–1506. [Google Scholar] [CrossRef]
- Panda, A.C.; Grammatikakis, I.; Kim, K.M.; De, S.; Martindale, J.L.; Munk, R.; Yang, X.; Abdelmohsen, K.; Gorospe, M. Identification of Senescence-Associated Circular RNAs (SAC-RNAs) Reveals Senescence Suppressor CircPVT1. Nucleic Acids Res. 2017, 45, 4021–4035. [Google Scholar] [CrossRef]
- Ghetti, M.; Vannini, I.; Storlazzi, C.T.; Martinelli, G.; Simonetti, G. Linear and Circular PVT1 in Hematological Malignancies and Immune Response: Two Faces of the Same Coin. Mol. Cancer 2020, 19, 69. [Google Scholar] [CrossRef]
- Geier, M.R.; Geier, D.A. Respiratory Conditions in Coronavirus Disease 2019 (COVID-19): Important Considerations Regarding Novel Treatment Strategies to Reduce Mortality. Med. Hypotheses 2020, 140, 109760. [Google Scholar] [CrossRef]
- Li, H.; Liu, S.-M.; Yu, X.-H.; Tang, S.-L.; Tang, C.-K. Coronavirus Disease 2019 (COVID-19): Current Status and Future Perspectives. Int. J. Antimicrob. Agents 2020, 55, 105951. [Google Scholar] [CrossRef]
- Chen, Y.; Ding, Y.-Y.; Ren, Y.; Cao, L.; Xu, Q.-Q.; Sun, L.; Xu, M.-G.; Lv, H.-T. Identification of Differentially Expressed MicroRNAs in Acute Kawasaki Disease. Mol. Med. Rep. 2018, 17, 932–938. [Google Scholar] [CrossRef]
- Jones, V.G.; Mills, M.; Suarez, D.; Hogan, C.A.; Yeh, D.; Segal, J.B.; Nguyen, E.L.; Barsh, G.R.; Maskatia, S.; Mathew, R. COVID-19 and Kawasaki Disease: Novel Virus and Novel Case. Hosp. Pediatr. 2020, 10, 537–540. [Google Scholar] [CrossRef]
- Fani, M.; Zandi, M.; Ebrahimi, S.; Soltani, S.; Abbasi, S. The Role of MiRNAs in COVID-19 Disease. Future Virol. 2021, 16, 301–306. [Google Scholar] [CrossRef]
- Gangemi, S.; Tonacci, A. AntagomiRs: A Novel Therapeutic Strategy for Challenging COVID-19 Cytokine Storm. Cytokine Growth Factor Rev. 2021, 58, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Seligmann, H. Balanced Evaluation of Preliminary Data on a Candidate COVID-19 Hydroxychloroquine Treatment. Int. J. Antimicrob. Agents 2021, 57, 106292. [Google Scholar] [CrossRef] [PubMed]
- Gautret, P.; Hoang, V.T.; Colson, P.; Raoult, D. Effect of Hydroxychloroquine and Azithromycin on the Viral Clearance of SARS-CoV-2: Response to Hervé Seligmann. Int. J. Antimicrob. Agents 2021, 57, 106306. [Google Scholar] [CrossRef]
- European Medicines Agency. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/comirnaty (accessed on 15 November 2022).
- Glassman, P.M.; Muzykantov, V.R. Pharmacokinetic and Pharmacodynamic Properties of Drug Delivery Systems. J. Pharmacol. Exp. Ther. 2019, 370, 570–580. [Google Scholar] [CrossRef]
- Dykxhoorn, D.M.; Lieberman, J. The Silent Revolution: RNA Interference as Basic Biology, Research Tool, and Therapeutic. Annu. Rev. Med. 2005, 56, 401–423. [Google Scholar] [CrossRef]
- Di, J.; Du, Z.; Wu, K.; Jin, S.; Wang, X.; Li, T.; Xu, Y. Biodistribution and Non-Linear Gene Expression of MRNA LNPs Affected by Delivery Route and Particle Size. Pharm. Res. 2022, 39, 105–114. [Google Scholar] [CrossRef]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyártó, B.Z. The MRNA-LNP Platform’s Lipid Nanoparticle Component Used in Preclinical Vaccine Studies Is Highly Inflammatory. iScience 2021, 24, 103479. [Google Scholar] [CrossRef]
- Somia, N.; Verma, I.M. Gene Therapy: Trials and Tribulations. Nat. Rev. Genet. 2000, 1, 91–99. [Google Scholar] [CrossRef]
- Lozier, J.N.; Csako, G.; Mondoro, T.H.; Krizek, D.M.; Metzger, M.E.; Costello, R.; Vostal, J.G.; Rick, M.E.; Donahue, R.E.; Morgan, R.A. Toxicity of a First-Generation Adenoviral Vector in Rhesus Macaques. Hum. Gene Ther. 2002, 13, 113–124. [Google Scholar] [CrossRef]
- Cichon, G.; Schmidt, H.H.; Benhidjeb, T.; Löser, P.; Ziemer, S.; Haas, R.; Grewe, N.; Schnieders, F.; Heeren, J.; Manns, M.P.; et al. Intravenous Administration of Recombinant Adenoviruses Causes Thrombocytopenia, Anemia and Erythroblastosis in Rabbits. J. Gene Med. 1999, 1, 360–371. [Google Scholar] [CrossRef]
- Varnavski, A.N.; Calcedo, R.; Bove, M.; Gao, G.; Wilson, J.M. Evaluation of Toxicity from High-Dose Systemic Administration of Recombinant Adenovirus Vector in Vector-Naive and Pre-Immunized Mice. Gene Ther. 2005, 12, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, P.; Medin, J.; da Silva, H.G.; Yadavalli, M.; Shankar, N.K.; Mullerova, H.; Arnold, M.; Nord, M. Very Rare Thrombosis with Thrombocytopenia after Second AZD1222 Dose: A Global Safety Database Analysis. Lancet 2021, 398, 577–578. [Google Scholar] [CrossRef] [PubMed]
- Resnier, P.; Montier, T.; Mathieu, V.; Benoit, J.-P.; Passirani, C. A Review of the Current Status of SiRNA Nanomedicines in the Treatment of Cancer. Biomaterials 2013, 34, 6429–6443. [Google Scholar] [CrossRef]
- Rohner, E.; Yang, R.; Foo, K.S.; Goedel, A.; Chien, K.R. Unlocking the Promise of MRNA Therapeutics. Nat. Biotechnol. 2022, 40, 1586–1600. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid Nanoparticles for MRNA Delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef] [PubMed]
- Kranz, L.M.; Diken, M.; Haas, H.; Kreiter, S.; Loquai, C.; Reuter, K.C.; Meng, M.; Fritz, D.; Vascotto, F.; Hefesha, H.; et al. Systemic RNA Delivery to Dendritic Cells Exploits Antiviral Defence for Cancer Immunotherapy. Nature 2016, 534, 396–401. [Google Scholar] [CrossRef]
- Sato, Y. Development of Lipid Nanoparticles for the Delivery of Macromolecules Based on the Molecular Design of PH-Sensitive Cationic Lipids. Chem. Pharm. Bull. 2021, 69, 1141–1159. [Google Scholar] [CrossRef]
- Harvie, P.; Wong, F.M.; Bally, M.B. Use of Poly(Ethylene Glycol)-Lipid Conjugates to Regulate the Surface Attributes and Transfection Activity of Lipid-DNA Particles. J. Pharm. Sci. 2000, 89, 652–663. [Google Scholar] [CrossRef]
- Song, L.Y.; Ahkong, Q.F.; Rong, Q.; Wang, Z.; Ansell, S.; Hope, M.J.; Mui, B. Characterization of the Inhibitory Effect of PEG-Lipid Conjugates on the Intracellular Delivery of Plasmid and Antisense DNA Mediated by Cationic Lipid Liposomes. Biochim. Et Biophys. Acta Biomembr. 2002, 1558, 1–13. [Google Scholar] [CrossRef]
- Mui, B.L.; Tam, Y.K.; Jayaraman, M.; Ansell, S.M.; Du, X.; Tam, Y.Y.C.; Lin, P.J.; Chen, S.; Narayanannair, J.K.; Rajeev, K.G.; et al. Influence of Polyethylene Glycol Lipid Desorption Rates on Pharmacokinetics and Pharmacodynamics of SiRNA Lipid Nanoparticles. Mol. Ther. Nucleic Acids 2013, 2, e139. [Google Scholar] [CrossRef] [PubMed]
- Knop, K.; Hoogenboom, R.; Fischer, D.; Schubert, U.S. Poly(Ethylene Glycol) in Drug Delivery: Pros and Cons as Well as Potential Alternatives. Angew. Chem. Int. Ed. Engl. 2010, 49, 6288–6308. [Google Scholar] [CrossRef] [PubMed]
- Abu Lila, A.S.; Kiwada, H.; Ishida, T. The Accelerated Blood Clearance (ABC) Phenomenon: Clinical Challenge and Approaches to Manage. J. Control Release 2013, 172, 38–47. [Google Scholar] [CrossRef]
- Kedmi, R.; Ben-Arie, N.; Peer, D. The Systemic Toxicity of Positively Charged Lipid Nanoparticles and the Role of Toll-like Receptor 4 in Immune Activation. Biomaterials 2010, 31, 6867–6875. [Google Scholar] [CrossRef]
- Abrams, M.T.; Koser, M.L.; Seitzer, J.; Williams, S.C.; DiPietro, M.A.; Wang, W.; Shaw, A.W.; Mao, X.; Jadhav, V.; Davide, J.P.; et al. Evaluation of Efficacy, Biodistribution, and Inflammation for a Potent SiRNA Nanoparticle: Effect of Dexamethasone Co-Treatment. Mol. Ther. J. Am. Soc. Gene Ther. 2010, 18, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Zhang, S.; Wang, B.; Cui, S.; Yan, J. Toxicity of Cationic Lipids and Cationic Polymers in Gene Delivery. J. Control Release 2006, 114, 100–109. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency. Assessment Report. Available online: https://www.ema.europa.eu/en/documents/assessment-report/comirnaty-epar-public-assessment-report_en.pdf (accessed on 15 November 2022).
- European Medicines Agency. COVID-19 Vaccine Moderna. Available online: https://www.ema.europa.eu/en/documents/assessment-report/spikevax-previously-covid-19-vaccine-moderna-epar-public-assessment-report_en.pdf (accessed on 15 November 2022).
- Rapozzi, V.; Juarranz, A.; Habib, A.; Ihan, A.; Strgar, R. Is Haem the Real Target of COVID-19? Photodiagnosis Photodyn. Ther. 2021, 35, 102381. [Google Scholar] [CrossRef]
- Yoshikawa, F.S.Y.; Teixeira, F.M.E.; Sato, M.N.; Oliveira, L.M.d.S. Delivery of MicroRNAs by Extracellular Vesicles in Viral Infections: Could the News Be Packaged? Cells 2019, 8, 611. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Ratajczak, J. Horizontal Transfer of RNA and Proteins between Cells by Extracellular Microvesicles: 14 Years Later. Clin. Transl. Med. 2016, 5, 7. [Google Scholar] [CrossRef]
- Chahar, H.S.; Bao, X.; Casola, A. Exosomes and Their Role in the Life Cycle and Pathogenesis of RNA Viruses. Viruses 2015, 7, 3204–3225. [Google Scholar] [CrossRef]
- Garg, A.; Seeliger, B.; Derda, A.A.; Xiao, K.; Gietz, A.; Scherf, K.; Sonnenschein, K.; Pink, I.; Hoeper, M.M.; Welte, T.; et al. Circulating Cardiovascular MicroRNAs in Critically Ill COVID-19 Patients. Eur. J. Heart Fail 2021, 23, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Deciphering the TRNA-Derived Small RNAs: Origin, Development, and Future. Cell Death & Disease. Available online: https://www.nature.com/articles/s41419-021-04472-3 (accessed on 15 November 2022).
- Xiao, Q.; Gao, P.; Huang, X.; Chen, X.; Chen, Q.; Lv, X.; Fu, Y.; Song, Y.; Wang, Z. TRFTars: Predicting the Targets of TRNA-Derived Fragments. J. Transl. Med. 2021, 19, 88. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.K.; Madhry, D.; Ravi Kumar, Y.S.; Malvankar, S.; Sapra, L.; Srivastava, R.K.; Bhattacharyya, S.; Verma, B. Regulatory Roles of TRNA-Derived RNA Fragments in Human Pathophysiology. Mol. Ther. Nucleic Acids 2021, 26, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Shan, N.; Lu, L.; Wang, Z. TRFtarget: A Database for Transfer RNA-Derived Fragment Targets. Nucleic Acids Res. 2021, 49, D254–D260. [Google Scholar] [CrossRef]
- Seneff, S.; Nigh, G.; Kyriakopoulos, A.M.; McCullough, P.A. Innate Immune Suppression by SARS-CoV-2 MRNA Vaccinations: The Role of G-Quadruplexes, Exosomes, and MicroRNAs. Food Chem. Toxicol. 2022, 164, 113008. [Google Scholar] [CrossRef]
- Paul, S.; Bravo Vázquez, L.A.; Reyes-Pérez, P.R.; Estrada-Meza, C.; Aponte Alburquerque, R.A.; Pathak, S.; Banerjee, A.; Bandyopadhyay, A.; Chakraborty, S.; Srivastava, A. The Role of MicroRNAs in Solving COVID-19 Puzzle from Infection to Therapeutics: A Mini-Review. Virus Res. 2022, 308, 198631. [Google Scholar] [CrossRef]
- PubChem BPTF. Bromodomain PHD Finger Transcription Factor (Human). Available online: https://pubchem.ncbi.nlm.nih.gov/gene/BPTF/human (accessed on 16 November 2022).
- Bakhta, A.; Boiveau, T.; Maday, Y.; Mula, O. Epidemiological Forecasting with Model Reduction of Compartmental Models. Application to the COVID-19 Pandemic. Biology 2020, 10, 22. [Google Scholar] [CrossRef]
- Wurtzer, S.; Waldman, P.; Ferrier-Rembert, A.; Frenois-Veyrat, G.; Mouchel, J.M.; Boni, M.; Maday, Y.; Marechal, V.; Moulin, L. Several Forms of SARS-CoV-2 RNA Can Be Detected in Wastewaters: Implication for Wastewater-Based Epidemiology and Risk Assessment. Water Res. 2021, 198, 117183. [Google Scholar] [CrossRef]
- Packer, M.; Gyawali, D.; Yerabolu, R.; Schariter, J.; White, P. A Novel Mechanism for the Loss of MRNA Activity in Lipid Nanoparticle Delivery Systems. Nat. Commun. 2021, 12, 6777. [Google Scholar] [CrossRef]
- Zendjabil, M.; Favard, S.; Tse, C.; Abbou, O.; Hainque, B. The microRNAs as biomarkers: What prospects? C R Biol. 2017, 340, 114–131. [Google Scholar] [CrossRef]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids Are Single-Stranded Covalently Closed Circular RNA Molecules Existing as Highly Base-Paired Rod-like Structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs Are the Predominant Transcript Isoform from Hundreds of Human Genes in Diverse Cell Types. PLoS ONE 2012, 7, e30733. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0030733 (accessed on 15 November 2022). [CrossRef] [PubMed]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs Are a Large Class of Animal RNAs with Regulatory Potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs Are Abundant, Conserved, and Associated with ALU Repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef]
- Santer, L.; Bär, C.; Thum, T. Circular RNAs: A Novel Class of Functional RNA Molecules with a Therapeutic Perspective. Mol. Ther. 2019, 27, 1350–1363. [Google Scholar] [CrossRef]
- Yahi, N.; Chahinian, H.; Fantini, J. Infection-Enhancing Anti-SARS-CoV-2 Antibodies Recognize Both the Original Wuhan/D614G Strain and Delta Variants. A Potential Risk for Mass Vaccination? J. Infect. 2021, 83, 607–635. [Google Scholar] [CrossRef]
- Tombácz, I.; Laczkó, D.; Shahnawaz, H.; Muramatsu, H.; Natesan, A.; Yadegari, A.; Papp, T.E.; Alameh, M.-G.; Shuvaev, V.; Mui, B.L.; et al. Highly Efficient CD4+ T Cell Targeting and Genetic Recombination Using Engineered CD4+ Cell-Homing MRNA-LNPs. Mol. Ther. 2021, 29, 3293–3304. [Google Scholar] [CrossRef]
- Veiga, N.; Goldsmith, M.; Granot, Y.; Rosenblum, D.; Dammes, N.; Kedmi, R.; Ramishetti, S.; Peer, D. Cell Specific Delivery of Modified MRNA Expressing Therapeutic Proteins to Leukocytes. Nat. Commun. 2018, 9, 4493. [Google Scholar] [CrossRef]
- Simula, E.R.; Manca, M.A.; Jasemi, S.; Uzzau, S.; Rubino, S.; Manchia, P.; Bitti, A.; Palermo, M.; Sechi, L.A. HCoV-NL63 and SARS-CoV-2 Share Recognized Epitopes by the Humoral Response in Sera of People Collected Pre- and during CoV-2 Pandemic. Microorganisms 2020, 8, 1993. [Google Scholar] [CrossRef]
- Baldassarre, A.; Paolini, A.; Bruno, S.P.; Felli, C.; Tozzi, A.E.; Masotti, A. Potential Use of Noncoding RNAs and Innovative Therapeutic Strategies to Target the 5′UTR of SARS-CoV-2. Epigenomics 2020, 12, 1349–1361. [Google Scholar] [CrossRef]
- Suzuki, Y.J. The Viral Protein Fragment Theory of COVID-19 Pathogenesis. Med. Hypotheses 2020, 144, 110267. [Google Scholar] [CrossRef] [PubMed]
- Pozniak, T.; Shcharbin, D.; Bryszewska, M. Circulating MicroRNAs in Medicine. Int. J. Mol. Sci. 2022, 23, 3996. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.; Lareau, C.A.; Keshishian, H.; Ganskih, S.; Schneider, C.; Hennig, T.; Melanson, R.; Werner, S.; Wei, Y.; Zimmer, M.; et al. The SARS-CoV-2 RNA–Protein Interactome in Infected Human Cells. Nat. Microbiol. 2021, 6, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Richards, A.; Barrasa, M.I.; Hughes, S.H.; Young, R.A.; Jaenisch, R. Reverse-Transcribed SARS-CoV-2 RNA Can Integrate into the Genome of Cultured Human Cells and Can Be Expressed in Patient-Derived Tissues. Proc. Natl. Acad. Sci. USA 2021, 118, e2105968118. [Google Scholar] [CrossRef]
- Allen, S.E.; Hug, I.; Pabian, S.; Rzeszutek, I.; Hoehener, C.; Nowacki, M. Circular Concatemers of Ultra-Short DNA Segments Produce Regulatory RNAs. Cell 2017, 168, 990–999.e7. [Google Scholar] [CrossRef]
- Kwon, Y.; Nukala, S.B.; Srivastava, S.; Miyamoto, H.; Ismail, N.I.; Jousma, J.; Rehman, J.; Ong, S.-B.; Lee, W.H.; Ong, S.-G. Detection of Viral RNA Fragments in Human IPSC Cardiomyocytes Following Treatment with Extracellular Vesicles from SARS-CoV-2 Coding Sequence Overexpressing Lung Epithelial Cells. Stem Cell Res. Ther. 2020, 11, 514. [Google Scholar] [CrossRef]
- Wolff, G.; Melia, C.E.; Snijder, E.J.; Bárcena, M. Double-Membrane Vesicles as Platforms for Viral Replication. Trends Microbiol. 2020, 28, 1022–1033. [Google Scholar] [CrossRef]
- Marchi, R.; Sugita, B.; Centa, A.; Fonseca, A.S.; Bortoletto, S.; Fiorentin, K.; Ferreira, S.; Cavalli, L.R. The Role of MicroRNAs in Modulating SARS-CoV-2 Infection in Human Cells: A Systematic Review. Infect. Genet. Evol. 2021, 91, 104832. [Google Scholar] [CrossRef]
- Ying, H.; Ebrahimi, M.; Keivan, M.; Khoshnam, S.E.; Salahi, S.; Farzaneh, M. MiRNAs; a Novel Strategy for the Treatment of COVID-19. Cell Biol. Int. 2021, 45, 2045–2053. [Google Scholar] [CrossRef]
- Fani, M.; Namdar-Ahmadabad, H.; Azimian, A.; Ghasemzadeh-Moghaddam, H. Predicting MicroRNAs as Anti-Viral Agents in SARS-CoV-2 Infection Based on the Bioinformatics Approach: A Systematic Review. J. Cell. Mol. Anesth. 2021, 6, 141–147. [Google Scholar] [CrossRef]
- Martinez-Espinoza, I.; Banos-Lara, M.D.R.; Guerrero-Plata, A. The Importance of MiRNA Identification During Respiratory Viral Infections. J. Cell Immunol. 2021, 3, 207–214. [Google Scholar] [CrossRef]
- Eichmeier, A.; Kiss, T.; Kocanova, M.; Hakalova, E.; Spetik, M.; Cechova, J.; Tichy, B. Conserved MicroRNAs in Human Nasopharynx Tissue Samples from Swabs Are Differentially Expressed in Response to SARS-CoV-2. Genes 2022, 13, 348. [Google Scholar] [CrossRef] [PubMed]
- Naser, M.; Naser, M.M.; Shehata, L.H. MiRNAs and COVID-19 Therapy Review. Int. J. Progress. Sci. Technol. 2022, 30, 404–415. [Google Scholar] [CrossRef]
- Shimabukuro, T.T.; Cole, M.; Su, J.R. Reports of Anaphylaxis After Receipt of MRNA COVID-19 Vaccines in the US—December 14, 2020-January 18, 2021. JAMA 2021, 325, 1101–1102. [Google Scholar] [CrossRef]
- Nogueira, S.S.; Schlegel, A.; Maxeiner, K.; Weber, B.; Barz, M.; Schroer, M.A.; Blanchet, C.E.; Svergun, D.I.; Ramishetti, S.; Peer, D.; et al. Polysarcosine-Functionalized Lipid Nanoparticles for Therapeutic MRNA Delivery. ACS Appl. Nano Mater. 2020, 3, 10634–10645. [Google Scholar] [CrossRef]
- Xu, Z.; Yang, D.; Wang, L.; Demongeot, J. Statistical Analysis Supports UTR (Untranslated Region) Deletion Theory in SARS-CoV-2. Virulence 2022, 13, 1772–1789. [Google Scholar] [CrossRef] [PubMed]
- Experton, B.; Tetteh, H.A.; Lurie, N.; Walker, P.; Elena, A.; Hein, C.S.; Schwendiman, B.; Vincent, J.L.; Burrow, C.R. A Predictive Model for Severe COVID-19 in the Medicare Population: A Tool for Prioritizing Primary and Booster COVID-19 Vaccination. Biology 2021, 10, 1185. [Google Scholar] [CrossRef] [PubMed]
- Barda, N.; Canetti, M.; Gilboa, M.; Asraf, K.; Indenboim, V.; Weiss-Ottolenghi, Y.; Amit, S.; Zubli, D.; Doolman, R.; Mendelson, E.; et al. The Association Between Pre-Booster Vaccination Antibody Levels and the Risk of SARS-CoV-2 Infection. Clin. Infect. Dis. 2022, ciac886. [Google Scholar] [CrossRef]
- Demongeot, J.; Flet-Berliac, Y.; Seligmann, H. Temperature Decreases Spread Parameters of the New Covid-19 Case Dynamics. Biology 2020, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Griette, Q.; Demongeot, J.; Magal, P. A Robust Phenomenological Approach to Investigate COVID-19 Data for France. Math. Appl. Sci. Eng. 2021, 2, 149–218. [Google Scholar] [CrossRef]
- Demongeot, J.; Griette, Q.; Magal, P.; Webb, G. Modeling Vaccine Efficacy for COVID-19 Outbreak in New York City. Biology 2022, 11, 345. [Google Scholar] [CrossRef] [PubMed]
- Demongeot, J.; Griette, Q.; Magal, P. SI Epidemic Model Applied to COVID-19 Data in Mainland China. R. Soc. Open Sci. 2020, 7, 201878. [Google Scholar] [CrossRef]
- Oshinubi, K.; Amakor, A.; Peter, O.J.; Rachdi, M.; Demongeot, J.; Oshinubi, K.; Amakor, A.; Peter, O.J.; Rachdi, M.; Demongeot, J. Approach to COVID-19 Time Series Data Using Deep Learning and Spectral Analysis Methods. AIMS Bioeng. 2022, 9, 1–21. [Google Scholar] [CrossRef]
- Oshinubi, K.; Ibrahim, F.; Rachdi, M.; Demongeot, J.; Oshinubi, K.; Ibrahim, F.; Rachdi, M.; Demongeot, J. Functional Data Analysis: Application to Daily Observation of COVID-19 Prevalence in France. AIMS Math. 2022, 7, 5347–5385. [Google Scholar] [CrossRef]
- Griette, Q.; Demongeot, J.; Magal, P. What Can We Learn from COVID-19 Data by Using Epidemic Models with Unidentified Infectious Cases? Math. Biosci. Eng. 2022, 19, 537–594. [Google Scholar] [CrossRef] [PubMed]
- Oshinubi, K.; Buhamra, S.S.; Al-Kandari, N.M.; Waku, J.; Rachdi, M.; Demongeot, J. Age Dependent Epidemic Modeling of COVID-19 Outbreak in Kuwait, France, and Cameroon. Healthcare 2022, 10, 482. [Google Scholar] [CrossRef]
- Oshinubi, K.; Fougère, C.; Demongeot, J. A Model for the Lifespan Loss Due to a Viral Disease: Example of the COVID-19 Outbreak. Infect. Dis. Rep. 2022, 14, 321–340. [Google Scholar] [CrossRef]
- Waku, J.; Oshinubi, K.; Demongeot, J. Maximal Reproduction Number Estimation and Identification of Transmission Rate from the First Inflection Point of New Infectious Cases Waves: COVID-19 Outbreak Example. Math. Comput. Simul. 2022, 198, 47–64. [Google Scholar] [CrossRef]
- Experton, B.; Elena, A.; Hein, C.S.; Nordenberg, D.; Walker, P.; Schwendiman, B.; Burrow, C.R. Enhanced Vaccine Effectiveness During the Delta Phase of the COVID-19 Pandemic in the Medicare Population Supports a Multilayered Prevention Approach. Biology 2022, 11, 1700. [Google Scholar] [CrossRef]
- Demongeot, J.; Magal, P. Spectral Method in Epidemic Time Series: Application to COVID-19 Pandemic. Biology 2022, 11, 1825. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aminoacid | L | D | K | V | E | A | E | V | Q | I | D | R | L |
| Virus | CUU | GAC | AAA | GUU | GAG | GCU | GAA | GUG | CAA | AUU | GAU | AGG | UUG |
| Vaccine | CΨG | GAC | CCΨ | CCΨ | GAG | GCC | GAG | GΨG | CAG | AΨC | GAC | AGA | CΨG |
| Aminoacid | L | D | P | P | E | A | E | V | Q | I | D | R | L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demongeot, J.; Fougère, C. mRNA COVID-19 Vaccines—Facts and Hypotheses on Fragmentation and Encapsulation. Vaccines 2023, 11, 40. https://doi.org/10.3390/vaccines11010040
Demongeot J, Fougère C. mRNA COVID-19 Vaccines—Facts and Hypotheses on Fragmentation and Encapsulation. Vaccines. 2023; 11(1):40. https://doi.org/10.3390/vaccines11010040
Chicago/Turabian StyleDemongeot, Jacques, and Cécile Fougère. 2023. "mRNA COVID-19 Vaccines—Facts and Hypotheses on Fragmentation and Encapsulation" Vaccines 11, no. 1: 40. https://doi.org/10.3390/vaccines11010040
APA StyleDemongeot, J., & Fougère, C. (2023). mRNA COVID-19 Vaccines—Facts and Hypotheses on Fragmentation and Encapsulation. Vaccines, 11(1), 40. https://doi.org/10.3390/vaccines11010040