
GlnH, a Novel Antigen That Offers Partial Protection against Verocytotoxigenic Escherichia coli Infection
 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Human Cell Lines and Maintenance
2.3. Attachment of E. coli Strains to Gastrointestinal Epithelial Cells
2.4. Outer Membrane Protein (OMP) Preparation and Quantification
2.5. Protein Separation by 2-D Gel Electrophoresis
2.6. Gastrointestinal Epithelial Cell-Probed Blots
2.7. Tryptic Digestion of Protein Samples and Preparation for LC/MS
2.8. PCR Analysis of Bovine and Human VTEC Isolates
2.9. Cloning of glnH Gene
2.10. Host Cell Attachment of E. coli BL21 Strain Expressing Recombinant GlnH
2.11. GlnH Expression and Purification
2.12. Preparation of Nalr-Resistant Prototype Strain E. coli O157 Strain NCTC 12900
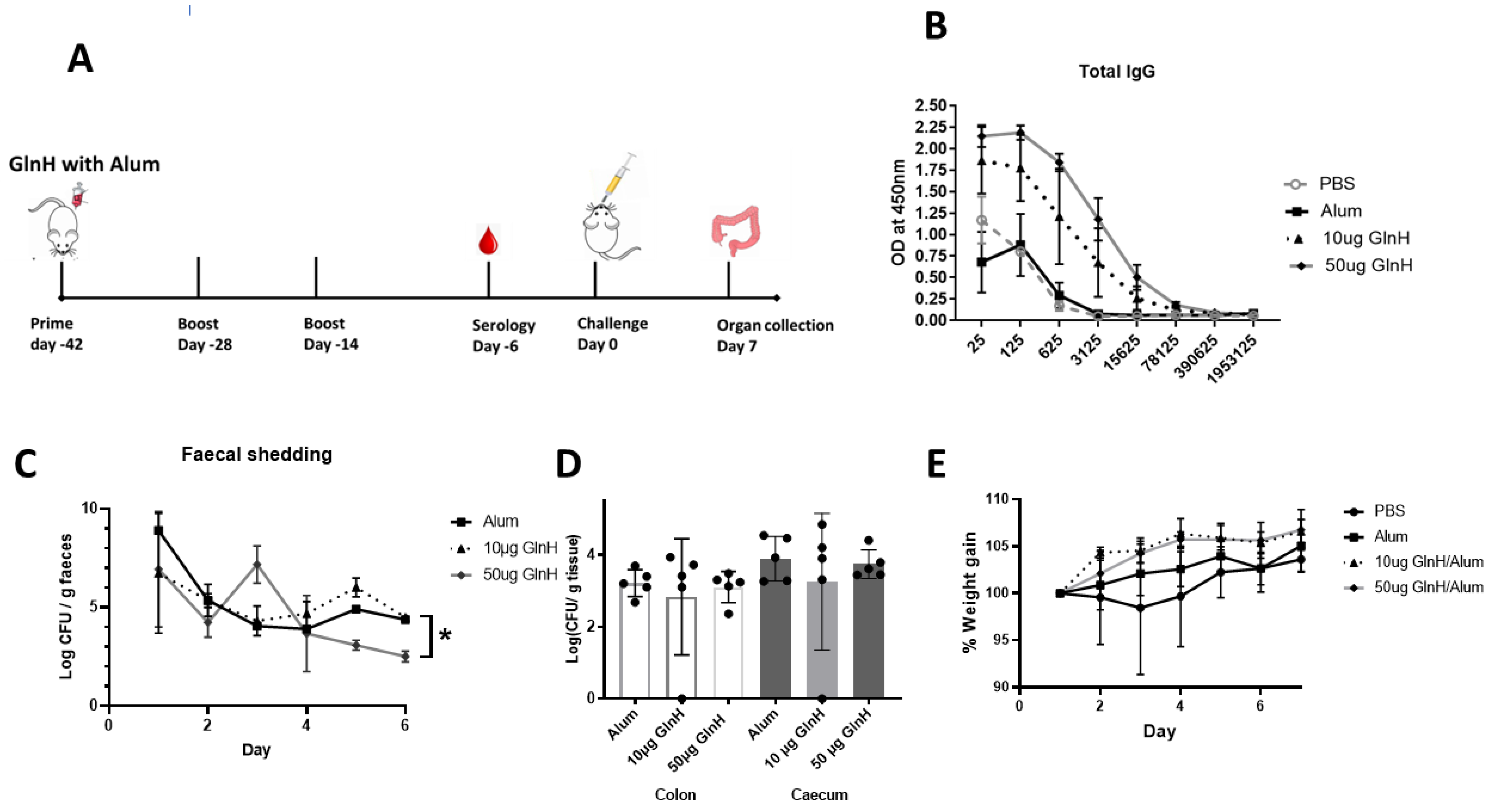
2.13. Immunisation with GlnH and Bacterial Challenge in BALB/c Mice
2.14. Serum IgG Antibodies Determination
2.15. Statistical Analysis
3. Results
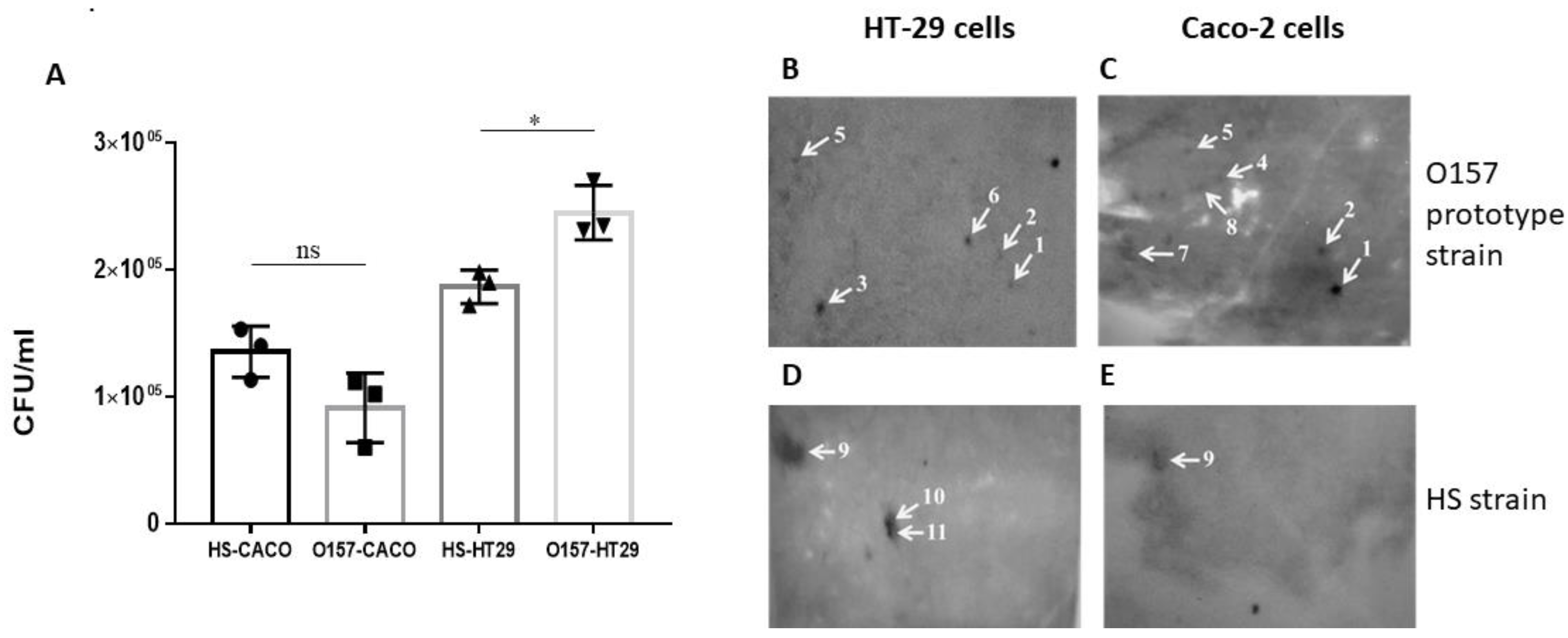
3.1. Comparison of Attachment of Commensal and VTEC Prototype Strains to Gastrointestinal Epithelial Cells
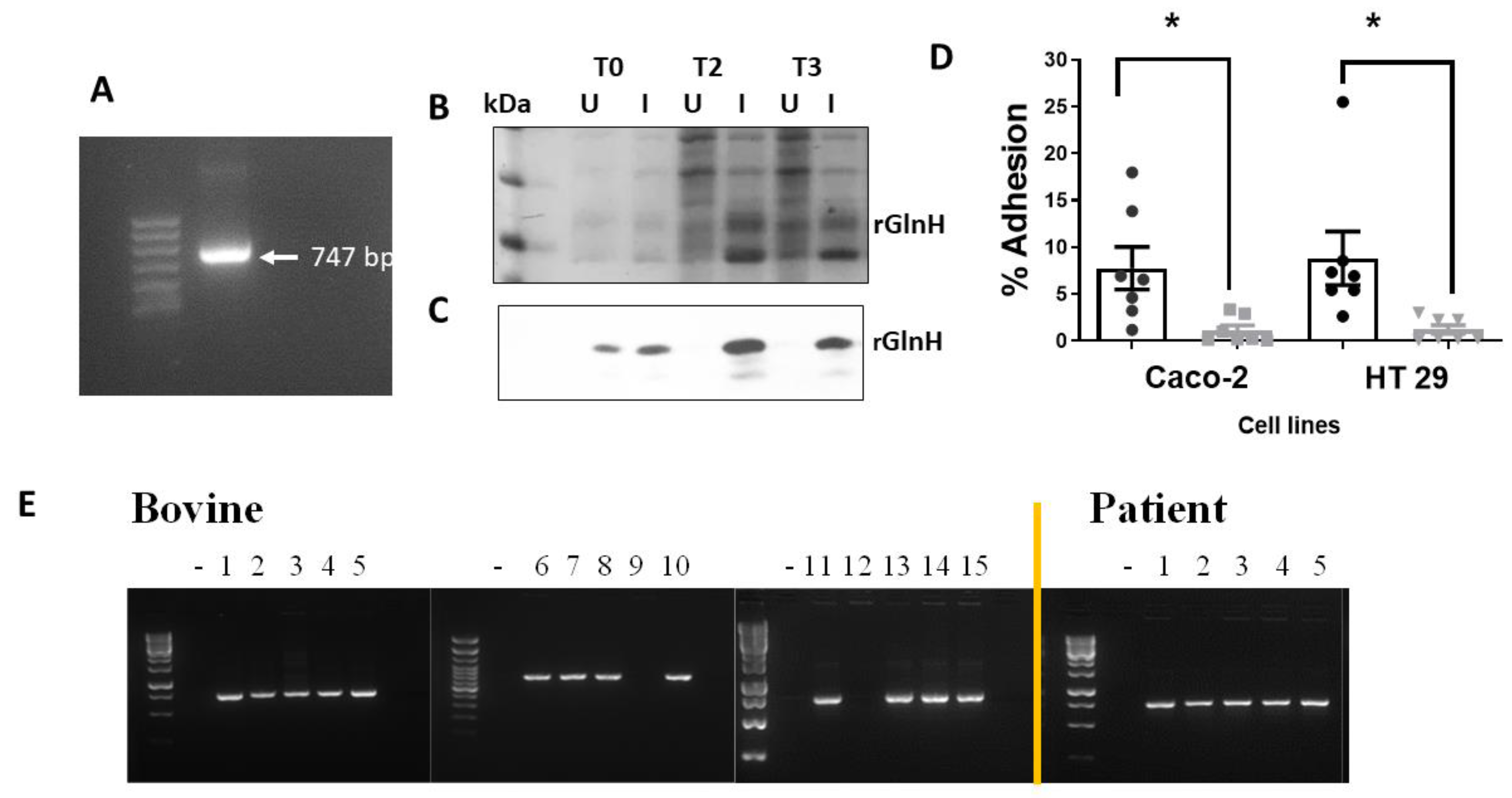
3.2. Identification of Proteins Involved in Gastrointestinal Cell Attachment
3.3. The GlnH Protein Plays a Role in E. coli Attachment
3.4. Presence in Bovine and Human Patient Samples
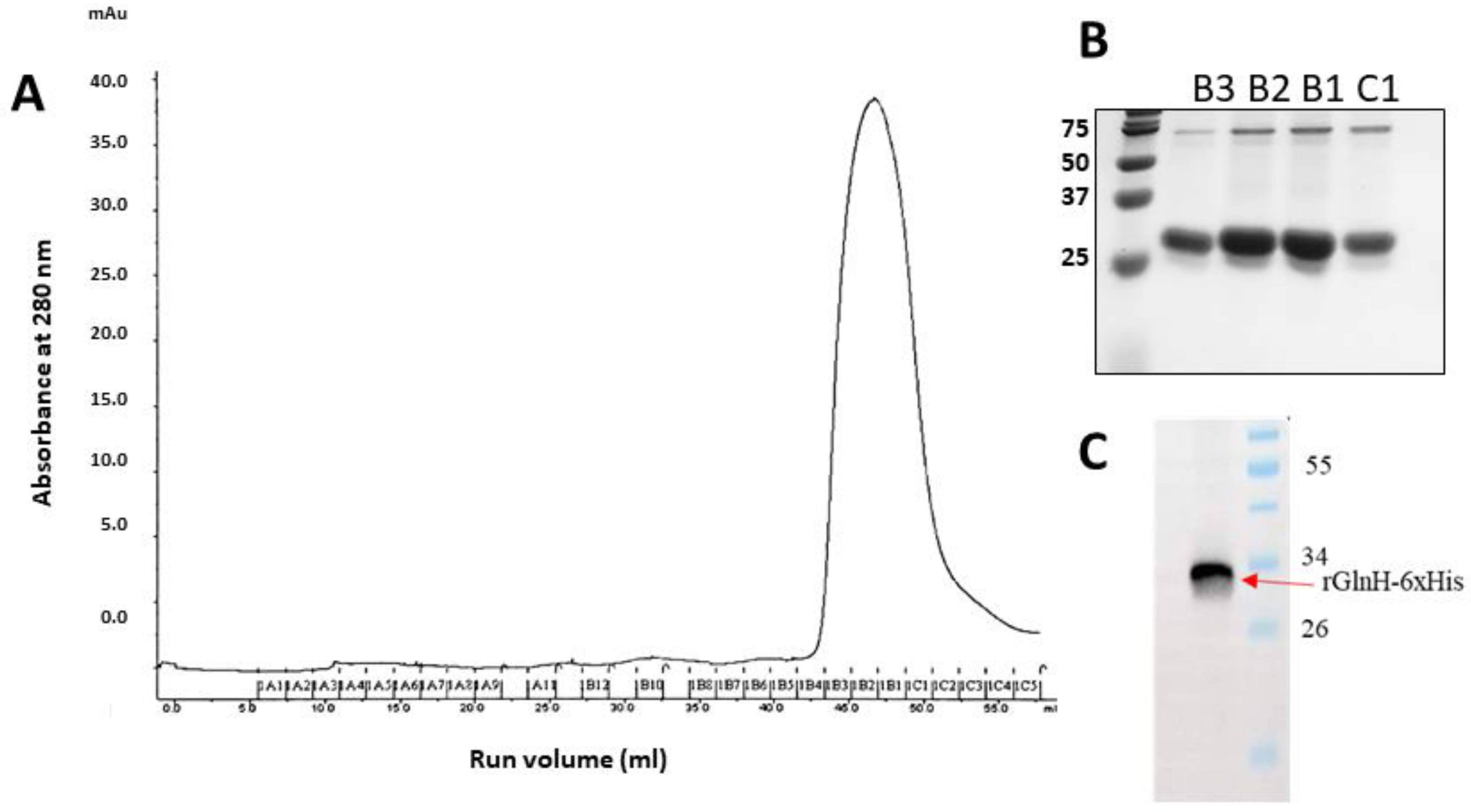
3.5. Purification of the Recombinant GlnH Protein for Preclinical Evaluation
3.6. GlnH Is Immunogenic in Immunised Mice
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karmali, M.A.; Gannon, V.; Sargeant, J.M. Verocytotoxin-producing Escherichia coli (VTEC). Vet. Microbiol. 2010, 140, 360–370. [Google Scholar] [CrossRef]
- Piérard, D.; De Greve, H.; Haesebrouck, F.; Mainil, J. O157:H7 and O104:H4 Vero/Shiga toxin-producing Escherichia coli outbreaks: Respective role of cattle and humans. Veter-Res. 2012, 43, 13. [Google Scholar] [CrossRef] [PubMed]
- Hall, G.; Kurosawa, S.; Stearns-Kurosawa, D.J.; Deborah, S.-K.; Shinichiro, K.; Gregory, H. Shiga toxin therapeutics: Beyond neutralization. Toxins 2017, 9, 291. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Andreoli, S. Extrarenal manifestations of the hemolytic uremic syndrome associated with Shiga toxin-producing Escherichia coli (STEC HUS). Pediatr. Nephrol. 2019, 34, 2495–2507. [Google Scholar] [CrossRef]
- Eriksson, K.J.; Boyd, S.G.; Tasker, R.C. Acute neurology and neurophysiology of haemolytic–uraemic syndrome. Arch. Dis. Child. 2001, 84, 434–435. [Google Scholar] [CrossRef]
- Garg, A.X.; Suri, R.S.; Barrowman, N.; Rehman, F.; Matsell, D.; Rosas-Arellano, M.P.; Salvadori, M.; Haynes, R.B.; Clark, W.F. Long-term Renal Prognosis of Diarrhea-Associated Hemolytic Uremic Syndrome: A Systematic Review, Meta-analysis, and Meta-regression. JAMA J. Am. Med. Assoc. 2003, 290, 1360–1370. [Google Scholar] [CrossRef] [PubMed]
- Kakoullis, L.; Papachristodoulou, E.; Chra, P.; Panos, G. Shiga toxin-induced haemolytic uraemic syndrome and the role of antibiotics: A global overview. J. Infect. 2019, 79, 75–94. [Google Scholar] [CrossRef] [PubMed]
- Nathanson, S.; Kwon, T.; Elmaleh, M.; Charbit, M.; Launay, E.A.; Harambat, J.; Brun, M.; Ranchin, B.; Bandin, F.; Cloarec, S.; et al. Acute neurological involvement in diarrhea-associated hemolytic uremic syndrome. Clin. J. Am. Soc. Nephrol. 2010, 5, 1218–1228. [Google Scholar] [CrossRef]
- Oakes, R.S.; Siegler, R.L.; McReynolds, M.A.; Pysher, T.; Pavia, A.T. Predictors of Fatality in Postdiarrheal Hemolytic Uremic Syndrome. Pediatrics 2006, 117, 1656–1662. [Google Scholar] [CrossRef]
- Spinale, J.M.; Ruebner, R.L.; Copelovitch, L.; Kaplan, B.S. Long-term outcomes of Shiga toxin hemolytic uremic syndrome. Pediatr. Nephrol. 2013, 28, 2097–2105. [Google Scholar] [CrossRef]
- Suri, R.S.; Mahon, J.L.; Clark, W.F.; Moist, L.M.; Salvadori, M.; Garg, A.X. Relationship between Escherichia coli O157:H7 and diabetes mellitus. Kidney Int. 2009, 75, S44–S46. [Google Scholar] [CrossRef]
- Suri, R.S.; Clark, W.F.; Barrowman, N.; Mahon, J.L.; Thiessen-Philbrook, H.R.; Rosas-Arellano, M.P.; Zarnke, K.; Garland, J.S.; Garg, A.X. Diabetes during diarrhea-associated hemolytic uremic syndrome: A systematic review and meta-analysis. Diabetes Care 2005, 28, 2556–2562. [Google Scholar] [CrossRef] [PubMed]
- Ylinen, E.; Salmenlinna, S.; Halkilahti, J.; Jahnukainen, T.; Korhonen, L.; Virkkala, T.; Rimhanen-Finne, R.; Nuutinen, M.; Kataja, J.; Arikoski, P.; et al. Hemolytic uremic syndrome caused by Shiga toxin–producing Escherichia coli in children: Incidence, risk factors, and clinical outcome. Pediatr. Nephrol. 2020, 35, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Ageorges, V.; Rojas-Lopez, M.; Schmidt, H.; Weiss, A.; Bertin, Y.; Forano, E.; Jubelin, G.; Henderson, I.R.; Livrelli, V.; et al. A secretome view of colonisation factors in Shiga toxin-encoding Escherichia coli (STEC): From enterohaemorrhagic E. coli (EHEC) to related enteropathotypes. FEMS Microbiol. Lett. 2016, 363, fnw179. [Google Scholar] [CrossRef]
- Rojas-Lopez, M.; Monterio, R.; Pizza, M.; Desvaux, M.; Rosini, R. Intestinal Pathogenic Escherichia coli: Insights for Vaccine Development. Front. Microbiol. 2018, 9, 440. [Google Scholar] [CrossRef]
- Garcia-Angulo, V.A.; Kalita, A.; Torres, A.G. Advances in the development of enterohemorrhagic Escherichia coli vaccines using murine models of infection. Vaccine 2013, 31, 3229–3235. [Google Scholar] [CrossRef] [PubMed]
- Walle, K.V.; Vanrompay, D.; Cox, E. Bovine innate and adaptive immune responses against Escherichia coli O157:H7 and vaccination strategies to reduce faecal shedding in ruminants. Vet. Immunol. Immunopathol. 2013, 152, 109–120. [Google Scholar] [CrossRef]
- Moxley, R.A. Escherichia coli 0157:H7: An update on intestinal colonization and virulence mechanisms. Anim. Health Res. Rev. 2004, 5, 15–33. [Google Scholar] [CrossRef]
- Potter, A.A.; Klashinsky, S.; Li, Y.; Frey, E.; Townsend, H.; Rogan, D.; Erickson, G.; Hinkley, S.; Klopfenstein, T.; Moxley, R.A.; et al. Decreased shedding of Escherichia coli O157:H7 by cattle following vaccination with type III secreted proteins. Vaccine 2004, 22, 362–369. [Google Scholar] [CrossRef]
- Allen, K.J.; Rogan, D.; Finlay, B.B.; Potter, A.A.; Asper, D.J. Vaccination with type III secreted proteins leads to decreased shedding in calves after experimental infection with Escherichia coli O157. Can. J. Vet. Res. 2011, 75, 98–105. [Google Scholar]
- Wizemann, T.M.; Adamou, J.E.; Langermann, S. Adhesins as targets for vaccine development. Emerg. Infect. Dis. 1999, 5, 395–403. [Google Scholar] [CrossRef]
- van Diemen, P.M.; Dziva, F.; Abu-Median, A.; Wallis, T.S.; van den Bosch, H.; Dougan, G.; Chanter, N.; Frankel, G.; Stevens, M.P. Subunit vaccines based on intimin and Efa-1 polypeptides induce humoral immunity in cattle but do not protect against intestinal colonisation by enterohaemorrhagic Escherichia coli O157:H7 or O26:H. Vet. Immunol. Immunopathol. 2007, 116, 47–58. [Google Scholar] [CrossRef]
- Amani, J.; Salmanian, A.H.; Rafati, S.; Mousavi, S.L. Immunogenic properties of chimeric protein from espA, eae and tir genes of Escherichia coli O157:H7. Vaccine 2010, 28, 6923–6929. [Google Scholar] [CrossRef]
- Jenkins, C.; Chart, H.; Smith, H.R.; Hartland, E.L.; Batchelor, M.; Delahay, R.M.; Dougan, G.; Frankel, G. Antibody response of patients infected with verocytotoxin-producing Escherichia coli to protein antigens encoded on the LEE locus. J. Med. Microbiol. 2000, 49, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Frey, E.; Mackenzie, A.M.; Finlay, B.B. Human response to Escherichia coli O157:H7 infection: Antibodies to secreted virulence factors. Infect. Immun. 2000, 68, 5090–5095. [Google Scholar] [CrossRef] [PubMed]
- McWilliams, B.D.; Torres, A.G. Enterohemorrhagic Escherichia coli Adhesins. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Ghaem-Maghami, M.; Simmons, C.P.; Daniell, S.; Pizza, M.; Lewis, D.; Frankel, G.; Dougan, G. Intimin-specific immune responses prevent bacterial colonization by the attaching-effacing pathogen Citrobacter rodentium. Infect. Immun. 2001, 69, 5597–5605. [Google Scholar] [CrossRef]
- García-Angulo, V.A.; Kalita, A.; Kalita, M.; Lozano, L.; Torres, A.G. Comparative genomics and immunoinformatics approach for the identification of vaccine candidates for enterohemorrhagic Escherichia coli O157:H7. Infect. Immun. 2014, 82, 2016–2026. [Google Scholar] [CrossRef]
- Sanchez-Villamil, J.I.; Tapia, D.; Torres, A.G. Development of a Gold Nanoparticle Vaccine against Enterohemorrhagic Escherichia coli O157:H7. mBio 2019, 10, e01869-19. [Google Scholar] [CrossRef]
- Uriza, P.J.; Trautman, C.; Palomino, M.M.; Fina Martin, J.; Ruzal, S.M.; Roset, M.S.; Briones, G. Development of an Antigen Delivery Platform Using Lactobacillus acidophilus Decorated With Heterologous Proteins: A Sheep in Wolf’s Clothing Story. Front. Microbiol. 2020, 11, 509380. [Google Scholar] [CrossRef]
- Iannino, F.; Uriza, P.J.; Duarte, C.M.; Pepe, M.V.; Roset, M.S.; Briones, G. Development of a Salmonella-based oral vaccine to control intestinal colonization of Shiga-toxin-producing Escherichia coli (STEC) in animals. Vaccine 2022, 40, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Fingermann, M.; Avila, L.; De Marco, M.B.; Vázquez, L.; Di Biase, D.N.; Müller, A.V.; Lescano, M.; Dokmetjian, J.C.; Fernández Castillo, S.; Pérez Quiñoy, J.L. OMV-based vaccine formulations against Shiga toxin producing Escherichia coli strains are both protective in mice and immunogenic in calves. Hum. Vaccin. Immunother. 2018, 14, 2208–2213. [Google Scholar] [CrossRef] [PubMed]
- Tobe, T.; Beatson, S.A.; Taniguchi, H.; Abe, H.; Bailey, C.M.; Fivian, A.; Younis, R.; Matthews, S.; Marches, O.; Frankel, G.; et al. An Extensive Repertoire of Type III Secretion Effectors in Escherichia coli O157 and the Role of Lambdoid Phages in Their Dissemination. Proc. Natl. Acad. Sci. USA 2006, 103, 14941–14946. [Google Scholar] [CrossRef] [PubMed]
- McClean, S.; Healy, M.E.; Collins, C.; Carberry, S.; O’Shaughnessy, L.; Dennehy, R.; Adams, Á.; Kennelly, H.; Corbett, J.M.; Carty, F.; et al. Linocin and OmpW Are Involved in Attachment of the Cystic Fibrosis-Associated Pathogen Burkholderia cepacia Complex to Lung Epithelial Cells and Protect Mice against Infection. Infect. Immun. 2016, 84, 1424–1437. [Google Scholar] [CrossRef]
- Casey, W.T.; Spink, N.; Cia, F.; Collins, C.; Romano, M.; Berisio, R.; Bancroft, G.J.; McClean, S. Identification of an OmpW homologue in Burkholderia pseudomallei, a protective vaccine antigen against melioidosis. Vaccine 2016, 34, 2616–2621. [Google Scholar] [CrossRef]
- Tomás-Cortázar, J.; Bossi, L.; Quinn, C.; Reynolds, C.J.; Butler, D.K.; Corcoran, N.; Murchú, M.; McMahon, E.; Singh, M.; Rongkard, P.; et al. BpOmpW Antigen Stimulates the Necessary Protective T-Cell Responses Against Melioidosis. Front. Immunol. 2021, 12, 767359. [Google Scholar] [CrossRef]
- La Ragione, R.M.; Ahmed, N.M.; Best, A.; Clifford, D.; Weyer, U.; Cooley, W.A.; Johnson, L.; Pearson, G.R.; Woodward, M.J. Colonization of 8-week-old conventionally reared goats by Escherichia coli O157: H7 after oral inoculation. J. Med. Microbiol. 2005, 54, 485–492. [Google Scholar] [CrossRef][Green Version]
- Mahajan, A.; Currie, C.G.; Mackie, S.; Tree, J.; McAteer, S.; McKendrick, I.; McNeilly, T.N.; Roe, A.; La Ragione, R.M.; Woodward, M.J.; et al. An investigation of the expression and adhesin function of H7 flagella in the interaction of Escherichia coli O157:H7 with bovine intestinal epithelium. Cell. Microbiol. 2009, 11, 121–137. [Google Scholar] [CrossRef]
- Montero, D.; Orellana, P.; Gutiérrez, D.; Araya, D.; Salazar, J.C.; Prado, V.; Oñate, A.; Del Canto, F.; Vidal, R. Immunoproteomic analysis to identify Shiga toxin-producing Escherichia coli outer membrane proteins expressed during human infection. Infect. Immun. 2014, 82, 4767–4777. [Google Scholar] [CrossRef]
- Shevchenko, A.; Mann, M.; Tomas, H.; Olsen, J.V.; Havli, J. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2007, 1, 2856–2860. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized ppb-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- Delvecchio, V.G.; Connolly, J.P.; Alefantis, T.G.; Walz, A.; Quan, M.A.; Patra, G.; Ashton, J.M.; Whittington, J.T.; Chafin, R.D.; Liang, X.; et al. Proteomic profiling and identification of immunodominant spore antigens of Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis. Appl. Environ. Microbiol. 2006, 72, 6355–6363. [Google Scholar] [CrossRef] [PubMed]
- Al-Maleki, A.R.; Mariappan, V.; Vellasamy, K.M.; Shankar, E.M.; Tay, S.T.; Vadivelu, J. Enhanced intracellular survival and epithelial cell adherence abilities of Burkholderia pseudomallei morphotypes are dependent on differential expression of virulence-associated proteins during mid-logarithmic growth phase. J. Proteom. 2014, 106, 205–220. [Google Scholar] [CrossRef]
- Reynolds, C.; Goudet, A.; Jenjaroen, K.; Sumonwiriya, M.; Rinchai, D.; Musson, J.; Overbeek, S.; Makinde, J.; Quigley, K.; Manji, J.; et al. T Cell Immunity to the Alkyl Hydroperoxide Reductase of Burkholderia pseudomallei: A Correlate of Disease Outcome in Acute Melioidosis. J. Immunol. 2015, 194, 4814–4824. [Google Scholar] [CrossRef] [PubMed]
- Asakura, H.; Panutdaporn, N.; Kawamoto, K.; Igimi, S.; Yamamoto, S.; Makino, S. Proteomic characterization of enterohemorrhagic escherichia coli O157:H7 in the oxidation-induced viable but non-culturable state. Microbiol. Immunol. 2007, 51, 875–881. [Google Scholar] [CrossRef]
- Malyala, P.; Singh, M. Endotoxin limits in formulations for preclinical research. J. Pharm. Sci. 2008, 97, 2041–2044. [Google Scholar] [CrossRef]
- Toledo, A.; Coleman, J.L.; Kuhlow, C.J.; Crowley, J.T.; Benach, J.L. The enolase of Borrelia burgdorferi is a plasminogen receptor released in outer membrane vesicles. Infect. Immun. 2011, 80, 359–368. [Google Scholar] [CrossRef]
- Capello, M.; Ferri-Borgogno, S.; Cappello, P.; Novelli, F. α-enolase: A promising therapeutic and diagnostic tumor target. FEBS J. 2011, 278, 1064–1074. [Google Scholar] [CrossRef]
- Pancholi, V. Multifunctional [alpha]-enolase: Its role in diseases. Cell. Mol. Life Sci. CMLS 2001, 58, 902. [Google Scholar] [CrossRef]
- Henderson, B.; Martin, A. Bacterial virulence in the moonlight: Multitasking bacterial moonlighting proteins are virulence determinants in infectious disease. Infect. Immun. 2011, 79, 3476–3491. [Google Scholar] [CrossRef]
- Choi, C.W.; Lee, Y.G.; Kwon, S.O.; Kim, H.Y.; Lee, J.C.; Chung, Y.H.; Yun, C.Y.; Kim, S.I. Analysis of Streptococcus pneumoniae secreted antigens by immuno-proteomic approach. Diagn. Microbiol. Infect. Dis. 2012, 72, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Ling, E.; Feldman, G.; Portnoi, M.; Dagan, R.; Overweg, K.; Mulholland, F.; Chalifa-Caspi, V.; Wells, J.; Mizrachi-Nebenzahl, Y. Glycolytic enzymes associated with the cell surface of Streptococcus pneumoniae are antigenic in humans and elicit protective immune responses in the mouse. Clin. Exp. Immunol. 2004, 138, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Kovach, Z.; Kaakoush, N.O.; Lamb, S.; Zhang, L.; Raftery, M.J.; Mitchell, H. Immunoreactive proteins of Campylobacter concisus, an emergent intestinal pathogen. FEMS Immunol. Med. Microbiol. 2011, 63, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Nowalk, A.J.; Nolder, C.; Clifton, D.R.; Carroll, J.A. Comparative proteome analysis of subcellular fractions from Borrelia burgdorferi by NEPHGE and IPG. Proteomics 2006, 6, 2121–2134. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, S.; Scire, A.; Varriale, A.; Scognamiglio, V.; Staiano, M.; Ausili, A.; Marabotti, A.; Rossi, M.; Tanfani, F. Binding of glutamine to glutamine-binding protein from Escherichia coli induces changes in protein structure and increases protein stability. Proteins 2005, 58, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Röhm, K.-H. A new subfamily of bacterial glutamate/aspartate receptors. Biol. Chem. 2008, 389, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Tam, R.; Saier, M.H., Jr. Structural, functional, and evolutionary relationships among extracellular solute-binding receptors of bacteria. Microbiol. Rev. 1993, 57, 320–346. [Google Scholar] [CrossRef]
- Herman, P.; Vecer, J.; Scognamiglio, V.; Staiano, M.; Rossi, M.; D’Auria, S. A Recombinant Glutamine-Binding Protein from Escherichia coli: Effect of Ligand-Binding on Protein Conformational Dynamics. Biotechnol. Prog. 2004, 20, 1847–1854. [Google Scholar] [CrossRef]
- Bai, J.; Kim, S.I.; Ryu, S.; Yoon, H. Identification and characterization of outer membrane vesicle-associated proteins in Salmonella enterica serovar Typhimurium. Infect. Immun. 2014, 82, 4001–4010. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.; Leon-Kempis, M.; Dodson, E.; Wilson, K.; Wilkinson, A.; Kelly, D. A bacterial virulence factor with a dual role as an adhesin and a solute-binding protein: The crystal structure at 1.5 angstrom resolution of the PEBIa protein from the food-borne human pathogen Campylobacter jejuni. J. Mol. Biol. 2007, 372, 160–171. [Google Scholar] [CrossRef]
- Du, L.F.; Li, Z.J.; Tang, X.Y.; Huang, J.Q.; Sun, W.B. Immunogenicity and immunoprotection of recombinant PEB1 in Campylobacter-jejuni-infected mice. World J. Gastroenterol. 2008, 14, 6244–6248. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.M.; Wang, J.; Hudson, D.L.; Grant, A.J.; Jones, M.A.; Maskell, D.J.; Stevens, M.P. Evaluation of live-attenuated Salmonella vaccines expressing Campylobacter antigens for control of C. jejuni in poultry. Vaccine 2010, 28, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.; Chafsey, I.; Ageorges, V.; Leroy, S.; Chambon, C.; Hébraud, M.; Livrelli, V.; Pizza, M.; Pezzicoli, A.; Desvaux, M. The Secretome landscape of Escherichia coli O157:H7: Deciphering the cell-surface, outer membrane vesicle and extracellular subproteomes. J. Proteom. 2021, 232, 104025. [Google Scholar] [CrossRef]
- Dziva, F.; Vlisidou, I.; Crepin, V.F.; Wallis, T.S.; Frankel, G.; Stevens, M.P. Vaccination of calves with EspA, a key colonisation factor of Escherichia coli O157:H7, induces antigen-specific humoral responses but does not confer protection against intestinal colonisation. Vet. Microbiol. 2007, 123, 254–261. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, T.N.; Mitchell, M.C.; Corbishley, A.; Nath, M.; Simmonds, H.; McAteer, S.P.; Mahajan, A.; Low, J.C.; Smith, D.G.E.; Huntley, J.F.; et al. Optimizing the protection of Cattle against Escherichia coli O157:H7 colonization through immunization with different combinations of H7 flagellin, Tir, intimin-531 or EspA. PLoS ONE 2015, 10, e0128391. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Milflores-Flores, L.; Garcia-Gallegos, J.G.; Patel, S.D.; Best, A.; La Ragione, R.M.; Martinez-Laguna, Y.; Woodward, M.J. Environmental regulation and colonization attributes of the long polar fimbriae (LPF) of Escherichia coli O157:H7. Int. J. Med. Microbiol. 2007, 297, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Shekar, A.; Ramlal, S.; Jeyabalaji, J.K.; Sripathy, M.H. Intranasal co-administration of recombinant active fragment of Zonula occludens toxin and truncated recombinant EspB triggers potent systemic, mucosal immune responses and reduces span of E. coli O157:H7 fecal shedding in BALB/c mice. Med. Microbiol. Immunol. 2019, 208, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.A.; Del Canto, F.; Salazar, J.C.; Céspedes, S.; Cádiz, L.; Arenas-Salinas, M.; Reyes, J.; Oñate, Á.; Vidal, R.M. Immunization of mice with chimeric antigens displaying selected epitopes confers protection against intestinal colonization and renal damage caused by Shiga toxin-producing Escherichia coli. NPJ Vaccines 2020, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Davitt, C.J.; Lavelle, E.C. Delivery strategies to enhance oral vaccination against enteric infections. Adv. Drug Deliv. Rev. 2015, 91, 52–69. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Protein ID 1 | Protein Name | Sequence Coverage (%) | Unique Peptides 2 | pI 3 | MW kDa 4 | Score 5 | Cell Line |
|---|---|---|---|---|---|---|---|---|
| 1 | P0AEQ5 | Glutamine-binding periplasmic protein (GlnH) | 77.8 | 21 | 8.44 | 27.19 | 358.58 | HT29 & Caco-2 |
| 2 | P65765 | FKBP-type peptidyl-prolyl cis-trans isomerase FkpA | 48.5 | 15 | 8.39 | 28.9 | 260.58 | HT29 & Caco-2 |
| 3 | P0AE10 | Alkyl hydroperoxide reductase C (AhpC) | 72.1 | 12 | 5.03 | 20.8 | 214.97 | HT29 only |
| 4 | P0A6Q1 | Enolase (Eno) | 41 | 22 | 5.32 | 45.7 | 323.31 | Caco-2 only |
| 5 | Q8XBL3 | Phosphoenolpyruvate-protein phosphotransferase (PPP) | 23.1 | 15 | 4.78 | 63.7 | 323.31 | Caco-2 only |
| 6 | P0AGF1 | Succinate—CoA ligase (ADP-forming) subunit alpha (SucD) | 66.4 | 14 | 6.32 | 30.1 | 247.46 | HT29 only |
| 7 | Q7ABI1 | Protein GrpE | 67.5 | 17 | 4.68 | 21.8 | 258.66 | HT29 & Caco-2 |
| 8 | Q8XD03 | Phosphoglycerate kinase (PgK) | 33.1 | 15 | 4.97 | 41.13 | 323.21 | Caco-2 only |
| 9 | Q8XE41 | Outer membrane protein C | 14.4 | 10 | 4.6 | 40.51 | 260.11 | HT29 & Caco-2 |
| 9a | P0A911 | Outer membrane protein A | 7.2 | 9 | 5.9 | 39.31 | 221.29 | HT29 & Caco-2 |
| 10 | Q8XAW6 | D-Ribose periplasmic binding protein (RbsB) | 67.9 | 16 | 6.85 | 30.9 | 304.08 | HT29 only |
| 11 | Q8X8L3 | Uridine phosphorylase (Udp) | 83.3 | 12 | 5.71 | 27.3 | 218.33 | HT29 only |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quinn, C.; Tomás-Cortázar, J.; Ofioritse, O.; Cosgrave, J.; Purcell, C.; McAloon, C.; Frost, S.; McClean, S. GlnH, a Novel Antigen That Offers Partial Protection against Verocytotoxigenic Escherichia coli Infection. Vaccines 2023, 11, 175. https://doi.org/10.3390/vaccines11010175
Quinn C, Tomás-Cortázar J, Ofioritse O, Cosgrave J, Purcell C, McAloon C, Frost S, McClean S. GlnH, a Novel Antigen That Offers Partial Protection against Verocytotoxigenic Escherichia coli Infection. Vaccines. 2023; 11(1):175. https://doi.org/10.3390/vaccines11010175
Chicago/Turabian StyleQuinn, Conor, Julen Tomás-Cortázar, Oritsejolomi Ofioritse, Joanne Cosgrave, Claire Purcell, Catherine McAloon, Susanna Frost, and Siobhán McClean. 2023. "GlnH, a Novel Antigen That Offers Partial Protection against Verocytotoxigenic Escherichia coli Infection" Vaccines 11, no. 1: 175. https://doi.org/10.3390/vaccines11010175
APA StyleQuinn, C., Tomás-Cortázar, J., Ofioritse, O., Cosgrave, J., Purcell, C., McAloon, C., Frost, S., & McClean, S. (2023). GlnH, a Novel Antigen That Offers Partial Protection against Verocytotoxigenic Escherichia coli Infection. Vaccines, 11(1), 175. https://doi.org/10.3390/vaccines11010175