
Molecular Epidemiology of Enterovirus A71 in Surveillance of Acute Flaccid Paralysis Cases in Senegal, 2013–2020

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Sample Preparation and NPEV Classification
2.3. RNA Extraction and Molecular Detection
2.4. Sequencing of the VP1 Gene
2.5. Sequencing
2.6. Phylogenetic Analyses
3. Results
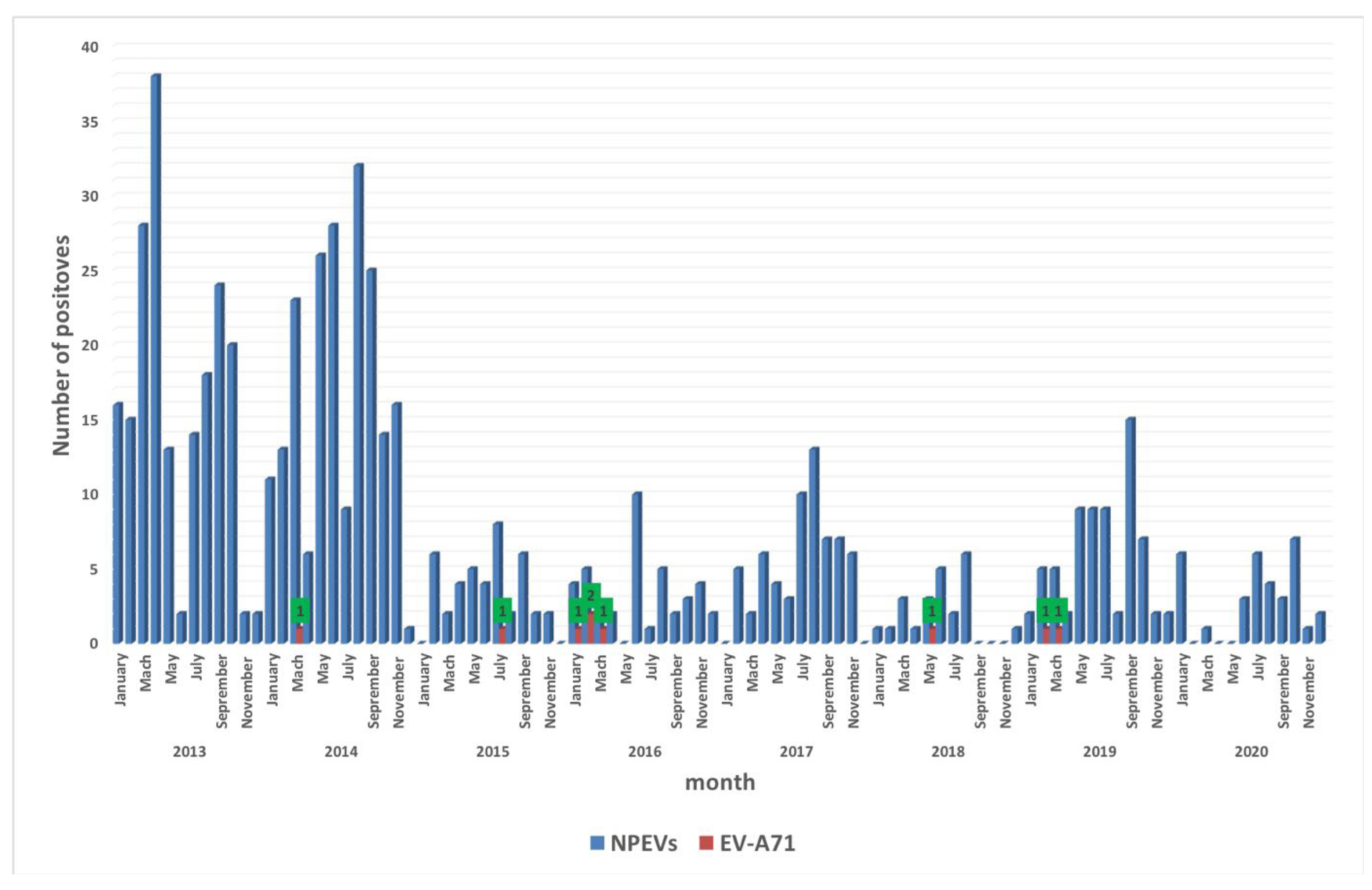
3.1. Temporal Distribution of EV-A71 Isolates
3.2. Geographical Distribution and Genomic Analysis
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Puenpa, J.; Wanlapakor, N.; Vongpunsawad, S.; Poovorawan, Y. The History of Enterovirus A71 Outbreaks and Molecular Epidemiology in the Asia-Pacific Region. J. Biomed. Sci. 2019, 26, 75. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Shih, R.S. Cell and tissue tropism of enterovirus 71 and others enteroviruses infections. J. Biomed. Sci. 2014, 21, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.T.C.; Wang, S.T.; Cheng, Y.T.; Yang, J.Y. Epidemiologic features of hand-foot-mouth disease and herpangina caused by enterovirus 71 in Taiwan, 1998–2005. Pediatrics 2007, 2, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.L.; Perera, D.; Cardosa, M.J.; McMinn, P.; Ooi, M.H. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet Infect. Dis. 2010, 11, 778–790. [Google Scholar] [CrossRef]
- Li, M.L.; Shih, S.; Tolbert, B.S.; Brewer, G. Enterovirus A71 Vaccines. Vaccines 2021, 9, 199. [Google Scholar] [CrossRef]
- Zhu, F.; Xu, W.; Xia, J.; Liang, Z.; Liu, Y.; Zhang, X.; Tan, X.; Wang, L.; Mao, Q.; Wu, J. Efficacy, safety, and immunogenicity of an enterovirus 71 vaccine in China. N. Engl. J. Med. 2014, 370, 818–828. [Google Scholar] [CrossRef]
- Huang, L.M.; Chiu, N.C.; Lin, C.Y.; Li, M.T.; Kuo, T.Y.; Weng, Y.J.; Hsieh, E.F.; Tai, I.C. Immunogenicity, safety, cross-reaction, and immune persistence of an inactivated enterovirus A71 vaccine in children aged from two months to 11 years in Taiwan. Vaccine 2019, 37, 1827–1835. [Google Scholar] [CrossRef]
- Volle, R.; Joffret, M.L.; Ndiaye, K. Development of a New Internally Controlled One-Step Real-Time RT-PCR for the Molecular Detection of Enterovirus A71 in Africa and Madagascar. Front. Microbiol. 2020, 11, 1907. [Google Scholar] [CrossRef]
- Yu, H.; Chen, W.; Chang, H. Genetic analysis of the VP1 region of enterovirus 71 reveals the emergence of genotype A in central China in 2008. Virus Genes 2010, 41, 1–4. [Google Scholar] [CrossRef]
- McMinn, P.C. Recent advances in the molecular epidemiology and control of human enterovirus 71 infection. Curr. Opin. Virol. 2012, 2, 199–205. [Google Scholar] [CrossRef]
- Saxena, V.K.; Sane, S.; Nadkarni, S.S.; Sharma, D.K.; Deshpande, J.M. Genetic diversity of enterovirus A71, India. Emerg. Infect. Dis. 2015, 21, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Tee, K.K.; Chan, Y.F.; Bibble, J.M.; Kamarulzaman, A.; Tong, C.Y.; Takebe, Y.; Pybus, O.G. Evolutionary genetics of human enterovirus 71: Origin, population dynamics, natural selection, and seasonal periodicity of the VP1 gene. J. Virol. 2010, 84, 3339–3350. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Enterovirus A-71. Available online: https://www.who.int/teams/health-product-policy-and-standards/standards-and-specifications/vaccine-standardization/enterovirus-71 (accessed on 22 March 2022).
- Gonzalez-Sanz, R.; Casas-Alba, D.; Launes, C.; Mirand, A.; Baily, J.L.; Otero, A.; Cabrerizo, M. Multirecombinant Enterovirus A71 Subgenogroup C1 Isolates Associated with Neurologic Disease, France, 2016–2017. Emerg. Infect. Dis. 2019, 25, 1204–1208. [Google Scholar]
- Wieczorek, M.; Purezynska, M.; Krzysztoszek, A.; Ciacka, A.; Figas, A.; Szenborn, L. Genetic characterization of enterovirus A71 isolates from severe neurological cases in Poland. J. Med. Virol. 2018, 90, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Casas-Alba, D.; de Selvilla, M.F.; Valero-Rello, A.; Fortuny, C.; MunozAlmagro, A.; Laune, C. Outbreak of brainstem encephalitis associated with enterovirus-A71 in Catalonia, Spain (2016): A clinical observational study in a children’s reference centre in Catalonia. Clin. Microbiol. Infect. 2017, 23, 874–881. [Google Scholar] [CrossRef] [Green Version]
- Bottcher, S.; Diedrich, S.; Keeren, K. The Laboratory Network for Enterovirus Diagnostic La NED. Increased detection of enterovirus A71 infections, Germany, 2019. Eurosurveillance 2019, 24, 39. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.S.; Miao, L.; Zhang, C.; Liu, Y. Molecular epidemiology and recombination of enterovirus A71 in mainland China from 1987 to 2017. Int. Microbiol. 2021, 24, 291–299. [Google Scholar] [CrossRef]
- Lee, M.S.; Tseng, F.C.; Wang, J.R.; Chi, C.Y.; Chong, P.; Su, I.J. Challenges to licensure of enterovirus 71 vaccines. PLoS Negl. Trop. Dis. 2012, 6, 1737. [Google Scholar] [CrossRef] [Green Version]
- Schufenecker, I.; Mirand, A.; Antona, D.; Henquell, C.; Chomel, J.J.; Archimbaud, C.; Billaud, G.; Peigue-Lafeuille, H.; Lina, B.; Bailly, J.L. Epidemiology of human enterovirus 71 infections in France, 2000–2009. J. Clin. Virol. 2011, 50, 56. [Google Scholar] [CrossRef]
- Witso, E.; Palacios, G.; Ronningen, K.S.; Cinek, O.; Janowitz, D.; Rewers, M.; Gri Grinde, B.; Lipkin, W.L. Asymptomatic circulation of HEV71 in Norway. Virus Res. 2007, 123, 19–29. [Google Scholar] [CrossRef]
- Silvia, M.; Sanna, G. Enterovirus 71 in Europe: A Briefing. Dev. Clin. Med. Pathol. 2018, 1, 1–3. [Google Scholar]
- Thao, N.T.T.; Donato, C.; Huong, V.T.Q.; Doorn, R.V.; Vijaykrishna, D. Evolution and Spatiotemporal Dynamics of Enterovirus A71 Subgenogroups in Vietnam. JID 2017, 216, 1371–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, R.; Iturriza-Gómara, M.; Musoke, R.; Palakudy, T.; D’Agostino, A.; Gray, J. An epidemic of enterovirus 71 infection among HIV-1-infected orphans in Nairobi. AIDS 2004, 18, 1968–1970. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, M.D.; Kebe, O.; Fall, A.D.; Delpeyroux, F.; Ndiaye, K. Enterovirus A71 Genogroups C and E in Children with Acute Flaccid Paralysis, West Africa. Emerg. Infect. Dis. 2016, 22, 753–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junttila, N.; Leveque, N.; Kabue, J.P.; Chomel, J.J.; Noder, H. New enteroviruses, EV-93 and EV-94, associated with acute flaccid paralysis in the Democratic Republic of the Congo. J. Med. Virol. 2007, 79, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Oyero, O.G.; Adu, F.D. Non-polio enteroviruses serotypes circulating in Nigeria. Afr. J. Med. Med. Sci. 2010, 39, 201–208. [Google Scholar] [PubMed]
- Bessaud, M.; Pillet, S.; Ibrahim, W.; Joffret, M.L.; Pozzetto, B.; Delpeyroux, F.; Vasilache, I.G. Molecular characterization of human enteroviruses in the Central African Republic: Uncovering wide diversity and identification of a new human enterovirus A71 genogroup. J. Clin. Microbiol. 2012, 50, 1650–1658. [Google Scholar] [CrossRef] [Green Version]
- Sadeuh-Mba, S.A.; Bessayd, M.; Massenet, D.; Rousset, D.; Delpeyroux, F. High frequency and diversity of species C enteroviruses in Cameroon and neighboring countries. J. Clin. Microbiol. 2013, 51, 759–770. [Google Scholar] [CrossRef] [Green Version]
- Gumede, N.; Okeibunor, J.; Diop, O. Progress on the Implementation of Environmental Surveillance in the African Region, 2011–2016. J. Immunol. Sci. 2018, 4, 24–30. [Google Scholar] [CrossRef]
- Fischer, T.K.; Simmond, P.; Harvala, H. The importance of enterovirus surveillance in a post-polio world. Lancet Infect. Dis. 2021, 20, 30852–30855. [Google Scholar] [CrossRef]
- Faleye, T.O.; Adewumi, M.O.; Coker, B.A.; Nudamajo, F.Y.; Adeniji, J.A. Direct detection and identification of enteroviruses from faeces of healthy Nigerian children using a cell-culture independent RT-seminested PCR assay. Adv. Virol. 2016, 2016, 1412838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Polio Laboratory Manual, Switzerland, 2004. Available online: https://apps.who.int/iris/handle/10665/68762 (accessed on 22 March 2022).
- Lewis, G.D.; Metcaf, T.G. Polyethylene glycol precipitation for recovery of pathogenic viruses, including hepatitis A virus and human rotavirus, from oyster, water, and sediment samples. Appl. Environ. Microbiol. 1988, 54, 1983–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunson, R.N. The Development, Implementation and Evaluation of a Real-Time PCR-Based Diagnostic Service for Viral Causes of Infectious Intestinal Disease. PhD Thesis, University of Glasgow, Scotland, UK, 2008. Available online: https://eleanor.lib.gla.ac.uk/record=b2617587 (accessed on 28 March 2022).
- Puenpa, J.; Suwannakarn, K.; Chansaenroj, J.; Vongpunsawad, S.; Poovorawan, Y. Development of single-step multiplex real-time RT-PCR assays for rapid diagnosis of enterovirus 71, coxsackievirus A6, and A16 in patients with hand, foot, and mouth disease. J. Virol. Methods 2017, 248, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Nix, W.A.; Oberste, M.S.; Pallansch, M.A. Sensitive, seminested PCR amplification of VP1 sequences for direct identification of all enterovirus serotypes from original clinical specimens. J. Clin. Microbiol. 2006, 44, 2698–2704. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Lo, C.C.; Anderson, J.J.; Davenport, K.W.; Bishop-Lilly, K.A.; Xu, Y.; Ahmed, S.; Feng, S.; Mokashi, V.P.; Chain, P.S.G. Enabling the democratization of the genomics revolution with a fully integrated web-based bioinformatics platform. Nucleic Acids Res. 2017, 45, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Garcia, M.D.; Volle, R.; Joffret, M.L.; Ndiaye, K.; Bessaud, M. Genetic Characterization of Enterovirus A71 Circulating in Africa. Emerg. Infect. Dis. 2018, 24, 754–757. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.L.; Cheng, C.P.; Su, H.J.; Liao, C.M.; Lin, C.Y.; Chou, Y.L.; Li, C.Y.; Chen, K.L.; Chen, K.T. The Association between Enterovirus 71 Infections and Meteorological Parameters in Taiwan. PLoS ONE 2012, 7, e46845. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, J.X.; Jin, P.F.; Wang, Y.X.; Zhu, F.C. Enterovirus 71: A whole virion inactivated enterovirus 71 vaccine. Expert Rev. Vaccines 2016, 15, 803–813. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Healthcare Environmental Infection Prevention and Control. Available online: https://www.cdc.gov/hai/prevent/environment/index.html (accessed on 5 April 2022).
- Mencattelli, G.; Ndione, M.H.D.; Rosa, R.; Giovanni, M.; Diagne, C.T.; Fall, G.; Faye, O.; Diallo, M.; Faye, O.; Savini, G.; et al. Epidemiology of West Nile virus in Africa: An underestimated threat. PLoS Negl. Trop. Dis. 2022, 16, 0010075. [Google Scholar] [CrossRef]
- Dia, N.; Lakh, N.; Diagne, M.M.; Diallo, K.; Fall, N.M.; Faye, O.; Jallow, M.M.; Dieng, I.; Diop, M.; Loucoubar, C.; et al. COVID-19 Outbreak, Senegal, 2020. Emerg. Infect. Dis. 2020, 26, 2772–2774. [Google Scholar] [CrossRef]
- Huang, K.; Zang, Y.; Han, Z.; Zhou, X.; Song, Y.; Wang, D.; Zhu, G.; Yan, D.; Xu, W.; Xu, W. Global Spread of the B5 Subgenotype EV-A71 and the Phylogeographical Analysis of Chinese Migration Events 2020. Front. Cell. Infect. Microbiol. 2020, 10, 475. [Google Scholar] [CrossRef] [PubMed]
- Adebisi, Y.A.; Rabe, A.; Prisno, D.E.L. COVID-19 surveillance systems in African countries. Health Promot. Perspect. 2021, 11, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.Y.; Wang, Y.; Bian, L.; Xu, M.; Liang, Z. EV-71 vaccine, a new tool to control outbreaks of hand, foot and mouth disease (HFMD). Expert Rev. Vaccines 2016, 15, 599–606. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Year of Isolation | Number of NPEV Isolates | Number of EV-A71 Isolates | Ratio of EV-A71-Positive Isolates (%) | |||
|---|---|---|---|---|---|---|
| AFP | ES | AFP | ES | AFP | ES | |
| 2013 | 36 | - | - | - | - | - |
| 2014 | 68 | - | 1 | - | 1.5 | - |
| 2015 | 41 | 24 | 1 | - | 2.4 | - |
| 2016 | 40 | 11 | 4 | - | 10 | - |
| 2017 | 63 | 35 | - | - | - | - |
| 2018 | 23 | 27 | 1 | - | 4.3 | - |
| 2019 | 69 | 31 | 2 | - | 2.9 | - |
| 2020 | 33 | 20 | - | - | - | - |
| Total | 373 | 148 | 9 | - | 2.4 | - |
| Strain | Years | Locality | Patient’s Gender | Case or Contact | Genogroup (Subgenotype) | Genbank Accession Number | Region | Reference |
|---|---|---|---|---|---|---|---|---|
| 14-157 | 2014 | Thies | Female | Case | C (c2) | MG672480 | Complete polyprotein | [39] |
| 15-355 | 2015 | Kaffrine | Female | Case | C (c2) | MG013988 | Complete polyprotein | [39] |
| 16-085 | 2016 | Diourbel | Male | Case | C (c2) | ON191506 | Partial VP1 gene | This study |
| 16-200 | 2016 | Dakar | Male | Case | C (c2) | ON191507 | Complete polyprotein | This study |
| 16-248 | 2016 | Saint Louis | Male | Case | C (c2) | ON191508 | Complete polyprotein | This study |
| 16-419 | 2016 | Kaolack | Male | Case | C (c1) | ON191509 | Complete polyprotein | This study |
| 18-1059 | 2018 | Tambacounda | Male | Case | C (c2) | ON191510 | Complete polyprotein | This study |
| 19-394 | 2019 | Diourbel | Male | Case | C (c2) | ON191511 | Complete polyprotein | This study |
| 19-196 | 2019 | Diourbel | Female | Case | C (c2) | ON191512 | Complete polyprotein | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndiaye, N.; Thiaw, F.D.; Fall, A.; Kébé, O.; Diatta, K.L.; Dia, N.; Fall, M.; Sall, A.A.; Faye, M.; Faye, O. Molecular Epidemiology of Enterovirus A71 in Surveillance of Acute Flaccid Paralysis Cases in Senegal, 2013–2020. Vaccines 2022, 10, 843. https://doi.org/10.3390/vaccines10060843
Ndiaye N, Thiaw FD, Fall A, Kébé O, Diatta KL, Dia N, Fall M, Sall AA, Faye M, Faye O. Molecular Epidemiology of Enterovirus A71 in Surveillance of Acute Flaccid Paralysis Cases in Senegal, 2013–2020. Vaccines. 2022; 10(6):843. https://doi.org/10.3390/vaccines10060843
Chicago/Turabian StyleNdiaye, Ndack, Fatou Diène Thiaw, Amary Fall, Ousmane Kébé, Khadija Leila Diatta, Ndongo Dia, Malick Fall, Amadou Alpha Sall, Martin Faye, and Ousmane Faye. 2022. "Molecular Epidemiology of Enterovirus A71 in Surveillance of Acute Flaccid Paralysis Cases in Senegal, 2013–2020" Vaccines 10, no. 6: 843. https://doi.org/10.3390/vaccines10060843
APA StyleNdiaye, N., Thiaw, F. D., Fall, A., Kébé, O., Diatta, K. L., Dia, N., Fall, M., Sall, A. A., Faye, M., & Faye, O. (2022). Molecular Epidemiology of Enterovirus A71 in Surveillance of Acute Flaccid Paralysis Cases in Senegal, 2013–2020. Vaccines, 10(6), 843. https://doi.org/10.3390/vaccines10060843