
Comparative Immunoreactivity Analyses of Hantaan Virus Glycoprotein-Derived MHC-I Epitopes in Vaccination


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. HTNV GP Protein Sequences Retrieval
2.2. HTNV GP Pan-MHC-I Epitopes Prediction and Screening
2.3. Conservation Analysis
2.4. Immunogenicity Analysis
2.5. Docking of pMHC-I Molecules
2.6. HTNV GP Peptides and Pan-MHC-I Clustering
2.7. Sequence Alignment of HTNV Variants
2.8. Pan-MHC-I-Restricted HTNV GP Epitopes Application by Literature Review
2.9. Vaccine, Animals, and Immunization
2.10. Peptides and ELISpot Assay
3. Results
3.1. HTNV GP Epitopes for Mouse MHC-I and Major HLA-I Supertypes
3.2. Conservation Status of HTNV GP 9-Mer Dominant Epitopes
3.3. Immunogenicity of HTNV GP 9-Mer Peptides
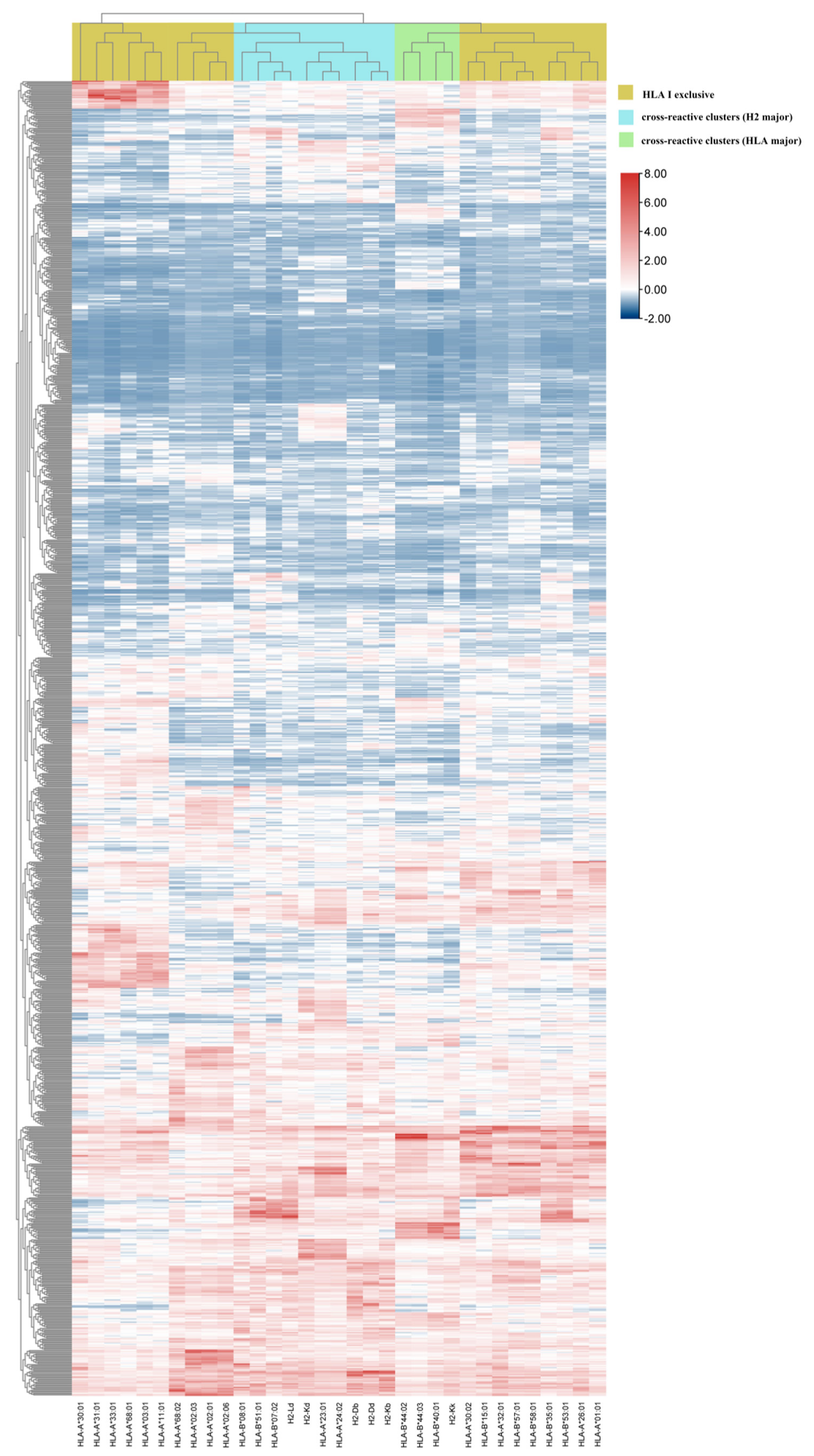
3.4. Hierarchical Clustering Showed Interaction between pan-MHC-I Molecules and HTNV 9-Mer Peptides
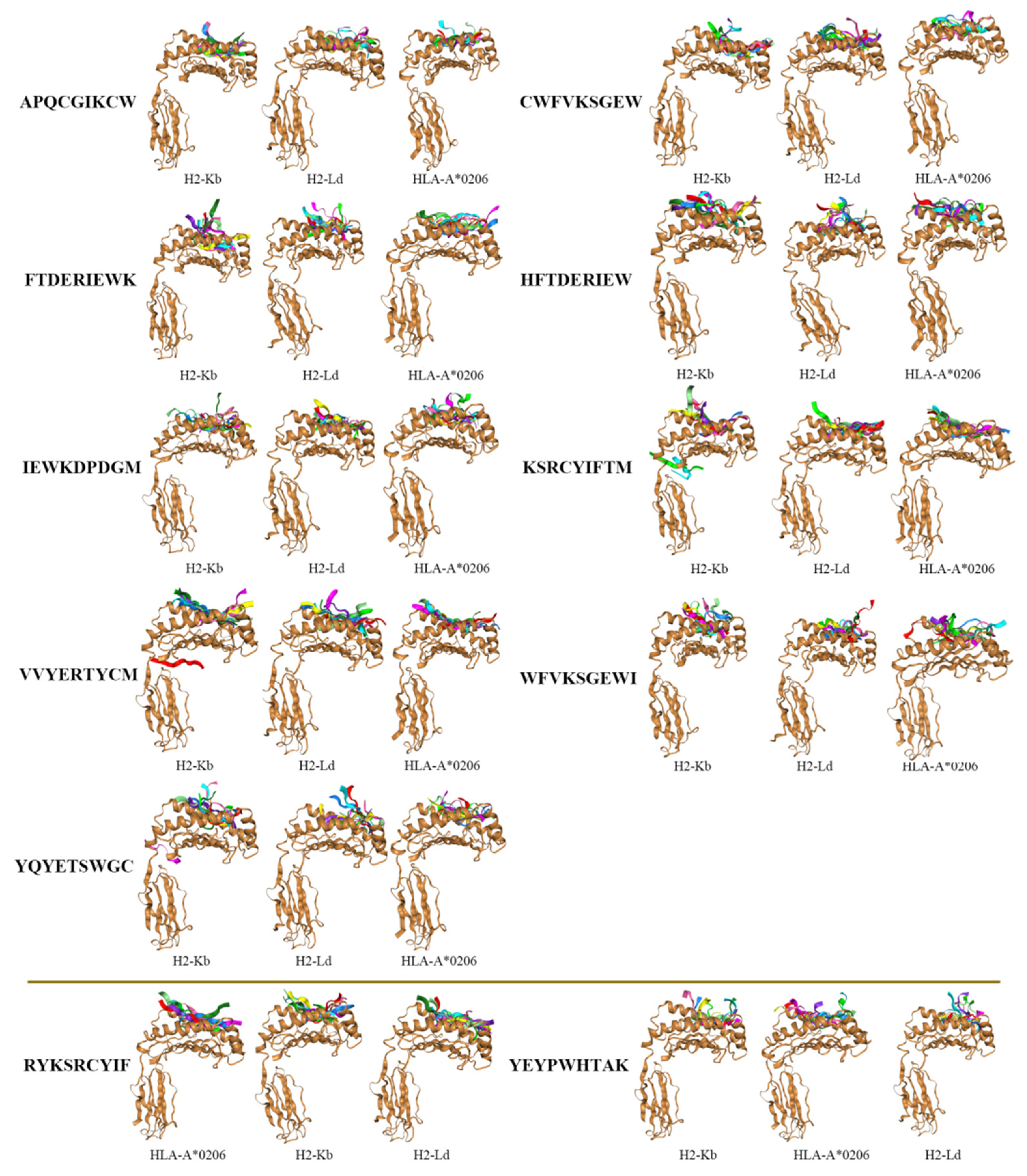
3.5. Docking of pMHC Molecules with the Dominant Epitopes
3.6. Comparison between the HTNV Strain 76-118 and the Other 147 Variants Based on Nine High-Affinity Segments
3.7. Affinity Differences of HTNV GP Hotspots between 76-118 and Variants
3.8. Applications HTNV GP-Derived CTL Epitopes by Literature Review
3.9. H2d Showed Cross-Immunoreactivity to HLA-A2-Restricted Epitopes in ELISpot Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Xu, C.; Wu, W.; Ren, J.; Li, Y.; Gui, L.; Yao, S. Time series analysis of temporal trends in hemorrhagic fever with renal syndrome morbidity rate in China from 2005 to 2019. Sci. Rep. 2020, 10, 9609. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Li, Y.; Li, Y.; Yu, C.; Bai, Y.; Wang, L.; Wang, Y. Estimating the Long-Term Epidemiological Trends and Seasonality of Hemorrhagic Fever with Renal Syndrome in China. Infect. Drug Resist. 2021, 14, 3849–3862. [Google Scholar] [CrossRef] [PubMed]
- Muyangwa, M.; Martynova, E.V.; Khaiboullina, S.F.; Morzunov, S.P.; Rizvanov, A.A. Hantaviral Proteins: Structure, Functions, and Role in Hantavirus Infection. Front. Microbiol. 2015, 6, 1326. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.L.; Wu, X.A.; Luo, W.; Bai, W.T.; Liu, Y.; Yan, Y.; Wang, H.T.; Xu, Z.K. The expression and genetic immunization of chimeric fragment of Hantaan virus M and S segments. Biochem. Biophys. Res. Commun. 2007, 354, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Bai, W.; Wu, X.; Zhang, L.; Zhang, L.; Li, P.; Wang, F.; Liu, Z.; Zhang, F.; Xu, Z. A recombinant pseudotyped lentivirus expressing the envelope glycoprotein of hantaan virus induced protective immunity in mice. Virol. J. 2013, 10, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manigold, T.; Mori, A.; Graumann, R.; Llop, E.; Simon, V.; Ferres, M.; Valdivieso, F.; Castillo, C.; Hjelle, B.; Vial, P. Highly differentiated, resting gn-specific memory CD8+ T cells persist years after infection by andes hantavirus. PLoS Pathog. 2010, 6, e1000779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, R.X.; Cheng, L.F.; Ying, Q.K.; Liu, R.R.; Ma, T.J.; Zhang, X.X.; Liu, Z.Y.; Zhang, L.; Ye, W.; Zhang, F.L.; et al. Screening and Identification of an H-2K(b)-Restricted CTL Epitope within the Glycoprotein of Hantaan Virus. Front. Cell Infect. Microbiol. 2016, 6, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, K.; Cheng, L.; Zhang, C.; Zhang, Y.; Zheng, X.; Zhang, Y.; Zhuang, R.; Jin, B.; Zhang, F.; Ma, Y. Novel Identified HLA-A*0201-Restricted Hantaan Virus Glycoprotein Cytotoxic T-Cell Epitopes Could Effectively Induce Protective Responses in HLA-A2.1/K(b) Transgenic Mice May Associate with the Severity of Hemorrhagic Fever with Renal Syndrome. Front. Immunol. 2017, 8, 1797. [Google Scholar] [CrossRef]
- Sette, A.; Sidney, A.J. Nine major HLA class I supertypes account for the vast preponderance of HLA-A and -B polymorphism. Immunogenetics 1999, 50, 201–212. [Google Scholar] [CrossRef]
- Kim, Y.; Ponomarenko, J.; Zhu, Z.; Tamang, D.; Wang, P.; Greenbaum, J.; Lundegaard, C.; Sette, A.; Lund, O.; Bourne, P.E.; et al. Immune epitope database analysis resource. Nucleic Acids Res. 2012, 40, W525–W530. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Sidney, J.; Pinilla, C.; Sette, A.; Peters, B. Derivation of an amino acid similarity matrix for peptide: MHC binding and its application as a Bayesian prior. BMC Bioinform. 2009, 10, 394. [Google Scholar] [CrossRef] [Green Version]
- Reynisson, B.; Alvarez, B.; Paul, S.; Peters, B.; Nielsen, M. NetMHCpan-4.1 and NetMHCIIpan-4.0: Improved predictions of MHC antigen presentation by concurrent motif deconvolution and integration of MS MHC eluted ligand data. Nucleic Acids Res. 2020, 48, W449–W454. [Google Scholar] [CrossRef] [PubMed]
- Rammensee, H.; Bachmann, J.; Emmerich, N.P.; Bachor, O.A.; Stevanovic, S. SYFPEITHI: Database for MHC ligands and peptide motifs. Immunogenetics 1999, 50, 213–219. [Google Scholar] [CrossRef]
- Reche, P.A.; Glutting, J.P.; Reinherz, E.L. Prediction of MHC class I binding peptides using profile motifs. Hum. Immunol. 2002, 63, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Reche, P.A.; Glutting, J.P.; Zhang, H.; Reinherz, E.L. Enhancement to the RANKPEP resource for the prediction of peptide binding to MHC molecules using profiles. Immunogenetics 2004, 56, 405–419. [Google Scholar] [CrossRef] [Green Version]
- Feltkamp, M.C.; Vierboom, M.P.; Kast, W.M.; Melief, C.J. Efficient MHC class I-peptide binding is required but does not ensure MHC class I-restricted immunogenicity. Mol. Immunol. 1994, 31, 1391–1401. [Google Scholar] [CrossRef]
- Soria-Guerra, R.E.; Nieto-Gomez, R.; Govea-Alonso, D.O.; Rosales-Mendoza, S. An overview of bioinformatics tools for epitope prediction: Implications on vaccine development. J. Biomed. Inform. 2015, 53, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Zaharieva, N.; Dimitrov, I.; Flower, D.R.; Doytchinova, I. VaxiJen Dataset of Bacterial Immunogens: An Update. Curr. Comput. Aided Drug Des. 2019, 15, 398–400. [Google Scholar] [CrossRef]
- Zhou, P.; Jin, B.; Li, H.; Huang, S.Y. HPEPDOCK: A web server for blind peptide-protein docking based on a hierarchical algorithm. Nucleic Acids Res. 2018, 46, W443–W450. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Tang, K.; Zhang, Y.; Zhang, C.; Cheng, L.; Zhang, F.; Zhuang, R.; Jin, B.; Zhang, Y. Protective CD8(+) T-cell response against Hantaan virus infection induced by immunization with designed linear multi-epitope peptides in HLA-A2.1/K(b) transgenic mice. Virol. J. 2020, 17, 146. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lu, Z.; Xuan, G.; Liu, N.; Wang, T.; Liu, Y.; Lan, M.; Xu, J.; Feng, Y.; Xu, S.; et al. Integrative Analysis of HTNV Glycoprotein Derived MHC II Epitopes by In Silico Prediction and Experimental Validation. Front. Cell Infect. Microbiol. 2021, 11, 671694. [Google Scholar] [CrossRef] [PubMed]
- Lamberth, K.; Roder, G.; Harndahl, M.; Nielsen, M.; Lundegaard, C.; Schafer-Nielsen, C.; Lund, O.; Buus, S. The peptide-binding specificity of HLA-A*3001 demonstrates membership of the HLA-A3 supertype. Immunogenetics 2008, 60, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Tang, K.; Zhang, Y.; Zhang, C.; Zhang, Y.; Jin, B.; Ma, Y. Design and synthesis of HLA-A*02-restricted Hantaan virus multiple-antigenic peptide for CD8(+) T cells. Virol. J. 2020, 17, 15. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Yoshimatsu, K.; Taruishi, M.; Tsuda, Y.; Arikawa, J. Involvement of CD8(+) T cells in the development of renal hemorrhage in a mouse model of hemorrhagic fever with renal syndrome. Arch. Virol. 2018, 163, 1577–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, G.J.; Chun, E.Y.; Kim, K.H.; Kim, W. Analysis of immune responses against nucleocapsid protein of the Hantaan virus elicited by virus infection or DNA vaccination. J. Microbiol. 2005, 43, 537–545. [Google Scholar] [PubMed]
- Araki, K.; Yoshimatsu, K.; Lee, B.H.; Kariwa, H.; Takashima, I.; Arikawa, J. Hantavirus-specific CD8(+)-T-cell responses in newborn mice persistently infected with Hantaan virus. J. Virol. 2003, 77, 8408–8417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perdomo-Celis, F.; Salvato, M.S.; Medina-Moreno, S.; Zapata, J.C. T-Cell Response to Viral Hemorrhagic Fevers. Vaccines 2019, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Xu, X.; Gu, R.; Wang, Z.; Chen, H.; Xu, K.; Zhang, M.; Hutton, J.; Yang, T. Prediction of HLA class I-restricted T-cell epitopes of islet autoantigen combined with binding and dissociation assays. Autoimmunity 2012, 45, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Hoof, I.; Costa, A.I.; van Baarle, D.; Kesmir, C. HLA class I allele promiscuity revisited. Immunogenetics 2011, 63, 691–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, S.; Chatziefthymiou, S.D.; Kolbe, M. Vaccines Targeting Numerous Coronavirus Antigens, Ensuring Broader Global Population Coverage: Multi-epitope and Multi-patch Vaccines. Methods Mol. Biol. 2022, 2410, 149–175. [Google Scholar] [PubMed]
- Yashvardhini, N.; Kumar, A.; Jha, D.K. Analysis of SARS-CoV-2 mutations in the main viral protease (NSP5) and its implications on the vaccine designing strategies. Vacunas 2021. [Google Scholar] [CrossRef] [PubMed]
- Bappy, S.S.; Sultana, S.; Adhikari, J.; Mahmud, S.; Khan, M.A.; Kibria, K.; Rahman, M.M.; Shibly, A.Z. Extensive immunoinformatics study for the prediction of novel peptide-based epitope vaccine with docking confirmation against envelope protein of Chikungunya virus: A computational biology approach. J. Biomol. Struct. Dyn. 2021, 39, 1139–1154. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, P.T.; Drexler, J.F.; Kallies, R.; Lickova, M.; Bokorova, S.; Maganga, G.D.; Szemes, T.; Leroy, E.M.; Kruger, D.H.; Drosten, C.; et al. Phylogenetic analysis of a newfound bat-borne hantavirus supports a laurasiatherian host association for ancestral mammalian hantaviruses. Infect. Genet. Evol. 2016, 41, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, R.; Gu, S.H.; Arai, S.; Kang, H.J.; Song, J.W. Hantaviruses: Rediscovery and new beginnings. Virus Res. 2014, 187, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Z. Discovery of hantaviruses in bats and insectivores and the evolution of the genus Hantavirus. Virus Res. 2014, 187, 15–21. [Google Scholar] [CrossRef]
- Forbes, K.M.; Sironen, T.; Plyusnin, A. Hantavirus maintenance and transmission in reservoir host populations. Curr. Opin. Virol. 2018, 28, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Salehi-Vaziri, M.; Sarvari, J.; Mansurnejadan, M.; Shiri, A.; Joharinia, N.; Khoshbakht, R.; Jaberi, O.; Pouriayevali, M.H.; Azad-Manjiri, S.; Jalali, T.; et al. Evidence of Hantavirus circulation among municipal street sweepers, southwest of Iran. Virusdisease 2021, 32, 251–254. [Google Scholar] [CrossRef]
- Engdahl, T.B.; Kuzmina, N.A.; Ronk, A.J.; Mire, C.E.; Hyde, M.A.; Kose, N.; Josleyn, M.D.; Sutton, R.E.; Mehta, A.; Wolters, R.M.; et al. Broad and potently neutralizing monoclonal antibodies isolated from human survivors of New World hantavirus infection. Cell Rep. 2021, 35, 109086. [Google Scholar] [CrossRef]
- Nguyen, A.; David, J.K.; Maden, S.K.; Wood, M.A.; Weeder, B.R.; Nellore, A.; Thompson, R.F. Human Leukocyte Antigen Susceptibility Map for Severe Acute Respiratory Syndrome Coronavirus 2. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, D.B.; Sun, L.J.; Cheng, L.F.; Zhang, J.P.; Xiao, S.B.; Sun, Y.J.; Yang, S.Y.; Wang, J.; Zhang, F.L.; Yang, K. Recombinant DNA vaccine of Hantavirus Gn and LAMP1 induced long-term immune protection in mice. Antivir. Res. 2017, 138, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Lai, J.H.; Zhu, Y.; Zhang, H.B.; Li, C.; Wang, J.P.; Li, Y.M.; Yang, A.G.; Jin, B.Q. Genetic susceptibility to haemorrhagic fever with renal syndrome caused by Hantaan virus in Chinese Han population. Int. J. Immunogenet. 2009, 36, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yuan, B.; Yi, J.; Zhuang, R.; Wang, J.; Zhang, Y.; Xu, Z.; Zhang, Y.; Liu, B.; Wei, C.; et al. The genetic polymorphisms of HLA are strongly correlated with the disease severity after Hantaan virus infection in the Chinese Han population. Clin. Dev. Immunol. 2012, 2012, 308237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Zhu, B.; Liu, M.; Li, S. Genetic polymorphisms at HLA-A, -B, and -DRB1 loci in Han population of Xi’an city in China. Croat Med. J. 2008, 49, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.M.; Zhu, B.F.; Ye, S.H.; Liu, M.L.; Yang, G.; Liu, S.; Qin, H.X.; Zhang, H.D.; Lucas, R.; Li, S.B. Allelic diversity and haplotype structure of HLA loci in the Chinese Han population living in the Guanzhong region of the Shaanxi province. Hum. Immunol. 2010, 71, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Korva, M.; Saksida, A.; Kunilo, S.; Vidan, J.B.; Avsic-Zupanc, T. HLA-associated hemorrhagic fever with renal syndrome disease progression in slovenian patients. Clin. Vaccine Immunol. 2011, 18, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.B.; Zhang, J.P.; Cheng, L.F.; Zhang, G.W.; Li, Y.; Li, Z.C.; Lu, Z.H.; Zhang, Z.X.; Lu, Y.C.; Zheng, L.H.; et al. Hantavirus Gc induces long-term immune protection via LAMP-targeting DNA vaccine strategy. Antivir. Res. 2018, 150, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Kariwa, H.; Lokugamage, K.; Lokugamage, N.; Miyamoto, H.; Yoshii, K.; Nakauchi, M.; Yoshimatsu, K.; Arikawa, J.; Ivanov, L.I.; Iwasaki, T.; et al. A comparative epidemiological study of hantavirus infection in Japan and Far East Russia. Jpn. J. Vet. Res. 2007, 54, 145–161. [Google Scholar]
- Dekonenko, A.E.; Tkachenko, E.A.; Lipskaia, G.; Dzagurova, T.K.; Ivanov, A.P.; Ivanov, L.I.; Slonova, R.A.; Markeshin, S.A.; Ivanidze, E.A.; Shutkova, T.M.; et al. Genetic differentiation of hantaviruses using the polymerase chain reaction and sequencing. Vopr. Virusol. 1996, 41, 24–27. [Google Scholar]
- Wang, Z.Q.; Wang, Y.L.; Fu, J.H.; Zhao, L.; Sun, C.Y.; Zhang, X.Q.; Zhang, Y.X.; Fan, S.Z.; Wang, N.D. Molecular analysis of hantavirus isolated from Shandong province. Chin. J. Exp. Clin. Virol. 2003, 17, 121–123. [Google Scholar]
- Zhou, J.H.; Zhang, H.L.; Wang, J.L.; Yang, W.H.; Mi, Z.Q.; Zhang, Y.Z.; Zhang, Y.Z.; Song, X.Y.; Hu, Q.L.; Dong, Y.K.; et al. Survey on host animal and molecular epidemiology of hantavirus in Chuxiong prefecture, Yunnan province. Zhonghua Liu Xing Bing Xue Za Zhi 2009, 30, 239–242. [Google Scholar] [PubMed]
- Lin, X.D.; Yang, P.F.; Liao, X.W.; Li, M.F.; Gao, N.; Chen, Y.; Zeng, S.D.; Wen, H.J.; Chen, L.P.; Li, M.H.; et al. The molecular epidemiologic investigation of hantavirus carried by rodent hosts in Wenzhou, Zhejiang province. Zhonghua Liu Xing Bing Xue Za Zhi 2008, 29, 891–894. [Google Scholar] [PubMed]
- Dheerasekara, K.; Sumathipala, S.; Muthugala, R. Hantavirus Infections-Treatment and Prevention. Curr. Treat. Options Infect. Dis. 2020, 12, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Chung, Y.S.; Yoon, C.H.; Lee, S.H.; Kim, S.S.; Kang, C.; Choi, B.S. Presentation of available CTL epitopes that induction of cell-mediated immune response against HIV-1 Koran clade B strain using computational technology. HIV Med. 2016, 17, 460–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Kolla, R.V.; Sidney, J.; Weiskopf, D.; Fleri, W.; Kim, Y.; Peters, B.; Sette, A. Evaluating the immunogenicity of protein drugs by applying in vitro MHC binding data and the immune epitope database and analysis resource. Clin. Dev. Immunol. 2013, 2013, 467852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Wang, J.; Yuan, B.; Wang, M.; Zhang, Y.; Xu, Z.; Zhang, C.; Zhang, Y.; Liu, B.; Yi, J.; et al. HLA-A2 and B35 restricted hantaan virus nucleoprotein CD8+ T-cell epitope-specific immune response correlates with milder disease in hemorrhagic fever with renal syndrome. PLoS Negl. Trop. Dis. 2013, 7, e2076. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.; Reche, P.; Pennisi, M.; Pappalardo, F. The combination of artificial intelligence and systems biology for intelligent vaccine design. Expert Opin. Drug Discov. 2020, 15, 1267–1281. [Google Scholar] [CrossRef] [PubMed]
- Michel-Todo, L.; Bigey, P.; Reche, P.A.; Pinazo, M.J.; Gascon, J.; Alonso-Padilla, J. Design of an Epitope-Based Vaccine Ensemble for Animal Trypanosomiasis by Computational Methods. Vaccines 2020, 8, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganaie, S.S.; Mir, M.A. The role of viral genomic RNA and nucleocapsid protein in the autophagic clearance of hantavirus glycoprotein Gn. Virus Res. 2014, 187, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Hussein, I.T.; Cheng, E.; Ganaie, S.S.; Werle, M.J.; Sheema, S.; Haque, A.; Mir, M.A. Autophagic clearance of Sin Nombre hantavirus glycoprotein Gn promotes virus replication in cells. J. Virol. 2012, 86, 7520–7529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Liu, W.; Hu, D.; Su, R.; Ji, M.; Huang, Y.; Shereen, M.A.; Xu, X.; Luo, Z.; Zhang, Q.; et al. HIV-1 Nef Interacts with LMP7 To Attenuate Immunoproteasome Formation and Major Histocompatibility Complex Class I Antigen Presentation. MBio 2020, 11, e02221-19. [Google Scholar] [CrossRef] [PubMed]
- Ressing, M.E.; Horst, D.; Griffin, B.D.; Tellam, J.; Zuo, J.; Khanna, R.; Rowe, M.; Wiertz, E.J. Epstein-Barr virus evasion of CD8(+) and CD4(+) T cell immunity via concerted actions of multiple gene products. Semin Cancer Biol. 2008, 18, 397–408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MHC-I Haplotypes | Prediction Tools | GP Epitopes | GP (Short-Listed) |
|---|---|---|---|
| HLA-A1 | IEDB | 32 | 32 |
| NetMHCpan | 32 | ||
| Rankpep | 0 | ||
| SMMPMBEC | 32 | ||
| SYFPEITHI | 10 | ||
| HLA-A2 | IEDB | 38 | 41 |
| NetMHCpan | 40 | ||
| Rankpep | 25 | ||
| SMMPMBEC | 28 | ||
| SYFPEITHI | 20 | ||
| HLA-A3 | IEDB | 52 | 57 |
| NetMHCpan | 53 | ||
| Rankpep | 35 | ||
| SMMPMBEC | 51 | ||
| SYFPEITHI | 37 | ||
| HLA-A24 | IEDB | 30 | 32 |
| NetMHCpan | 30 | ||
| Rankpep | 16 | ||
| SMMPMBEC | 24 | ||
| SYFPEITHI | 16 | ||
| HLA-B7 | IEDB | 40 | 41 |
| NetMHCpan | 41 | ||
| Rankpep | 27 | ||
| SMMPMBEC | 35 | ||
| SYFPEITHI | 25 | ||
| HLA-B8 | IEDB | 11 | 11 |
| NetMHCpan | 11 | ||
| Rankpep | 0 | ||
| SMMPMBEC | 9 | ||
| SYFPEITHI | 5 | ||
| HLA-B15 | IEDB | 31 | 33 |
| NetMHCpan | 32 | ||
| Rankpep | 3 | ||
| SMMPMBEC | 26 | ||
| SYFPEITHI | 21 | ||
| HLA-B44 | IEDB | 26 | 26 |
| NetMHCpan | 26 | ||
| Rankpep | 4 | ||
| SMMPMBEC | 23 | ||
| SYFPEITHI | 17 | ||
| HLA-B58 | IEDB | 24 | 25 |
| NetMHCpan | 24 | ||
| Rankpep | 19 | ||
| SMMPMBEC | 18 | ||
| SYFPEITHI | 17 |
| MHC-I Haplotypes | Prediction Tools | GP Epitopes | GP (Short-Listed) |
|---|---|---|---|
| H2-Db | IEDB | 17 | 18 |
| NetMHCpan | 17 | ||
| Rankpep | 7 | ||
| SMMPMBEC | 12 | ||
| SYFPEITHI | 14 | ||
| H2-Dd | IEDB | 10 | 12 |
| NetMHCpan | 10 | ||
| Rankpep | 8 | ||
| SMMPMBEC | 7 | ||
| SYFPEITHI | NA | ||
| H2-Kb | IEDB | 15 | 15 |
| NetMHCpan | 15 | ||
| Rankpep | 8 | ||
| SMMPMBEC | 11 | ||
| SYFPEITHI | NA | ||
| H2-Kd | IEDB | 16 | 17 |
| NetMHCpan | 16 | ||
| Rankpep | 10 | ||
| SMMPMBEC | 9 | ||
| SYFPEITHI | 13 | ||
| H2-Kk | IEDB | 12 | 15 |
| NetMHCpan | 12 | ||
| Rankpep | 5 | ||
| SMMPMBEC | 8 | ||
| SYFPEITHI | 8 | ||
| H2-Ld | IEDB | 12 | 15 |
| NetMHCpan | 13 | ||
| Rankpep | 7 | ||
| SMMPMBEC | 7 | ||
| SYFPEITHI | 6 |
| MHC-I Haplotypes | Interspecies- Intraspecies- | Interspecies- Intraspecies+ | Interspecies+ Intraspecies- | Interspecies+ Intraspecies+ |
|---|---|---|---|---|
| H2-Db | 16 | 2 | 0 | 0 |
| H2-Dd | 10 | 2 | 0 | 0 |
| H2-Kb | 12 | 2 | 0 | 1 |
| H2-Kd | 11 | 5 | 0 | 1 |
| H2-Kk | 13 | 2 | 0 | 0 |
| H2-Ld | 10 | 3 | 0 | 2 |
| HLA-A1 | 22 | 9 | 0 | 1 |
| HLA-A2 | 34 | 6 | 0 | 1 |
| HLA-A3 | 40 | 14 | 0 | 3 |
| HLA-A24 | 17 | 9 | 0 | 6 |
| HLA-B7 | 27 | 10 | 1 | 3 |
| HLA-B8 | 5 | 4 | 0 | 2 |
| HLA-B15 | 21 | 9 | 0 | 1 |
| HLA-B44 | 19 | 5 | 0 | 2 |
| HLA-B58 | 15 | 9 | 1 | 0 |
| Amino Acid Number | KT885048.1 | Variants | Dominant in KT885048.1 | Dominant in Variants | HLA-I Genotype |
|---|---|---|---|---|---|
| 214–222 | AVKGNTYKI | AVKGNTYKL | No | Yes | HLA-B07:02 |
| No | Yes | HLA-B08:01 | |||
| No | Yes | HLA-B15:01 | |||
| No | Yes | HLA-A30:02 | |||
| 218–226 | NTYKIFEQV | NTYKLFEQV | Yes | No | HLA-A32:01 |
| 221–229 | KIFEQVKKS | KLFEQVKKS | No | Yes | HLA-A02:01 |
| No | Yes | HLA-A02:03 | |||
| No | Yes | HLA-A02:06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, B.; Zhang, J.; Wang, J.; Liu, Y.; Sun, H.; Lu, Z.; Chen, L.; Ding, X.; Pan, J.; Hu, C.; et al. Comparative Immunoreactivity Analyses of Hantaan Virus Glycoprotein-Derived MHC-I Epitopes in Vaccination. Vaccines 2022, 10, 564. https://doi.org/10.3390/vaccines10040564
Sun B, Zhang J, Wang J, Liu Y, Sun H, Lu Z, Chen L, Ding X, Pan J, Hu C, et al. Comparative Immunoreactivity Analyses of Hantaan Virus Glycoprotein-Derived MHC-I Epitopes in Vaccination. Vaccines. 2022; 10(4):564. https://doi.org/10.3390/vaccines10040564
Chicago/Turabian StyleSun, Baozeng, Junqi Zhang, Jiawei Wang, Yang Liu, Hao Sun, Zhenhua Lu, Longyu Chen, Xushen Ding, Jingyu Pan, Chenchen Hu, and et al. 2022. "Comparative Immunoreactivity Analyses of Hantaan Virus Glycoprotein-Derived MHC-I Epitopes in Vaccination" Vaccines 10, no. 4: 564. https://doi.org/10.3390/vaccines10040564
APA StyleSun, B., Zhang, J., Wang, J., Liu, Y., Sun, H., Lu, Z., Chen, L., Ding, X., Pan, J., Hu, C., Yang, S., Jiang, D., & Yang, K. (2022). Comparative Immunoreactivity Analyses of Hantaan Virus Glycoprotein-Derived MHC-I Epitopes in Vaccination. Vaccines, 10(4), 564. https://doi.org/10.3390/vaccines10040564