
Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies


Abstract
1. Introduction
2. Coccidiosis in Chickens
3. Host Immune Response to Coccidiosis
3.1. Role of Gut-Associated Lymphoid Tissues (GALT) and Mucosal-Associated Lymphoid Tissues (MALT)
3.2. Role of Cell-Mediated Immune Response
3.3. Role of Cytokines and Chemokines
3.4. Role of the Gut Microbiome in Host Response
4. Recombinant Vaccine Antigens of Eimeria
4.1. Candidate Antigens for Vaccine Development
4.2. Molecular Vaccines against Coccidiosis
4.3. Adjuvants, Cytokines and the Modes of Recombinant Vaccine Immunization
4.4. Delivery Vectors for Recombinant Vaccines
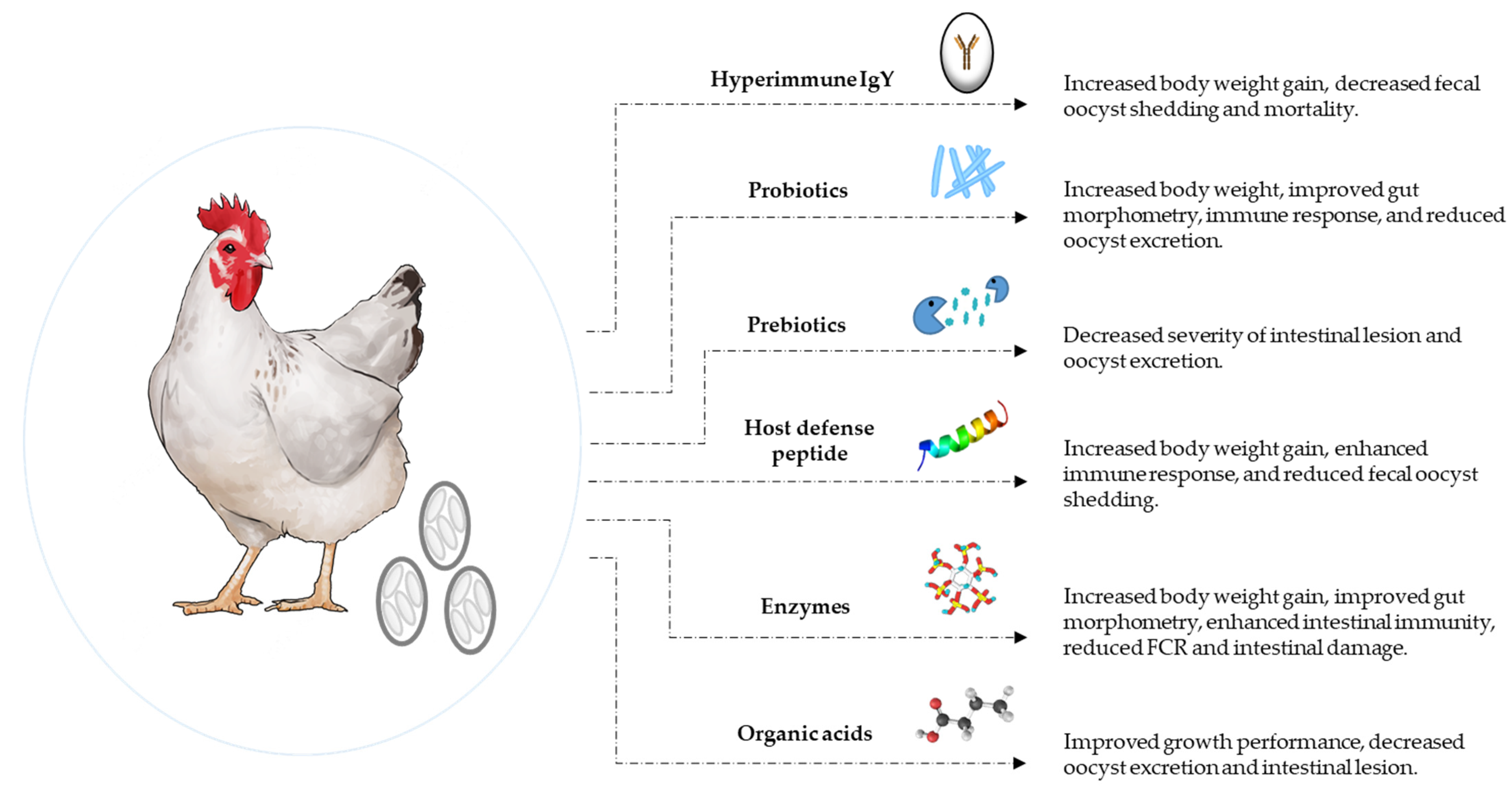
5. Alternatives to Antibiotic (ATA) Strategies to Control Coccidiosis
5.1. Hyperimmune Egg Yolk Antibodies
5.2. Probiotics
5.3. Prebiotics
5.4. Host Defense Peptides
5.5. Organic Acids and other Feed Additives
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.S.; Ding, X.; Quiroz, M.A.; Bevensee, E.; Lillehoj, E.P. Resistance to intestinal coccidiosis following DNA immunization with the cloned 3-1E Eimeria gene plus IL-2, IL-15, and IFN-γ. Avian Dis. 2005, 49, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Soutter, F.; Werling, D.; Tomley, F.M.; Blake, D.P. Poultry Coccidiosis: Design and interpretation of vaccine Studies. Front. Vet. Sci. 2020, 7, 101. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B. Anticoccidial vaccines for broiler chickens: Pathways to success. Avian Pathol. 2002, 31, 317–353. [Google Scholar] [CrossRef]
- Wickramasuriya, S.S.; Park, I.; Lee, Y.; Kim, W.H.; Przybyszewski, C.; Gay, C.G.; Oosterwijk, J.G.; Lillehoj, H.S. Oral delivery of Bacillus subtilis expressing chicken NK-2 peptide protects against Eimeria acervulina infection in broiler chickens. Front. Vet. Sci. 2021, 8, 562. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B. Epidemiological aspects of the use of live anticoccidial vaccines for chickens. Int. J. Parasitol. 1998, 28, 1089–1098. [Google Scholar] [CrossRef]
- Blake, D.P.; Clark, E.L.; Macdonald, S.E.; Thenmozhi, V.; Kundu, K.; Garg, R.; Jatau, I.D.; Ayoade, S.; Kawahara, F.; Moftah, A.; et al. Population, genetic, and antigenic diversity of the apicomplexan Eimeria tenella and their relevance to vaccine development. Proc. Natl. Acad. Sci. USA 2015, 112, E5343–E5350. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Lillehoj, E.P.; Hong, Y.H.; An, D.J.; Jeoung, H.Y.; Chun, J.E. Relative disease susceptibility and clostridial toxin antibody responses in three commercial broiler lines coinfected with clostridium perfringens and eimeria maxima using an experimental model of necrotic enteritis. Avian Dis. 2013, 57, 684–687. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, S.H.; Gadde, U.D.; Oh, S.T.; Lee, S.J.; Lillehoj, H.S. Allium hookeri supplementation improves intestinal immune response against necrotic enteritis in young broiler chickens. Poult. Sci. 2018, 97, 1899–1908. [Google Scholar] [CrossRef]
- Park, S.S.; Lillehoj, H.S.; Allen, P.C.; Park, D.W.; FitzCoy, S.; Bautista, D.A.; Lillehoj, E.P. Immunopathology and cytokine responses in broiler chickens coinfected with Eimeria maxima and Clostridium perfringens with the use of an animal model of necrotic enteritis. Avian Dis. 2008, 52, 14–22. [Google Scholar] [CrossRef]
- Lillehoj, H.; Okamura, M. Host immunity and vaccine development to coccidia and salmonella infections in chickens. J. Poult. Sci. 2003, 40, 151–193. [Google Scholar] [CrossRef]
- Chaudhari, A.A.; Lee, Y.; Lillehoj, H.S. Beneficial effects of dietary supplementation of Bacillus strains on growth performance and gut health in chickens with mixed coccidiosis infection. Vet. Parasitol. 2020, 277, 109009. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Lillehoj, E.P.; Bertrand, F.; Dupuis, L.; Deville, S. Mucosal immunity against Eimeria acervulina infection in broiler chickens following oral immunization with profilin in MontanideTM adjuvants. Exp. Parasitol. 2011, 129, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Mayer, L. Review article: Local and systemic regulation of mucosal immunity. Aliment. Pharmacol. Ther. 1997, 11, 81–88. [Google Scholar] [CrossRef]
- Matsumoto, R.; Hashimoto, Y. Distribution and developmental change of lymphoid tissues in the chicken proventriculus. J. Vet. Med. Sci. 2000, 62, 161–167. [Google Scholar] [CrossRef]
- Tellez, G.; Shivaramaiah, S.; Barta, J.; Hernandez-Velasco, X.; Hargis, B. Coccidiosis: Recent advancements in the immunobiology of Eimeria species, preventive measures, and the importance of vaccination as a control tool against these Apicomplexan parasites. Vet. Med. Res. Rep. 2014, 5, 23–34. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Trout, J.M. Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev. 1996, 9, 349–360. [Google Scholar] [CrossRef]
- Lillehoj, H.S. Role of T lymphocytes and cytokines in coccidiosis. Int. J. Parasitol. 1998, 28, 1071–1081. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Lillehoj, H.S. Poultry coccidiosis: Recent advancements in control measures and vaccine development. Expert Rev. Vaccines 2006, 5, 143–163. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Lillehoj, E.P. Avian Coccidiosis. A Review of acquired intestinal immunity and vaccination Strategies. Avian Dis. 2000, 44, 408–425. [Google Scholar] [CrossRef]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and regulation of the mucus barrier in the gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Nguyen, T.; Liu, L.; Sacco, R.E.; Brogden, K.A.; Lehrer, R.I. Gallinacin-3, an inducible epithelial β-defensin in the chicken. Infect. Immun. 2001, 69, 2684–2691. [Google Scholar] [CrossRef] [PubMed]
- Sunkara, L.T.; Achanta, M.; Schreiber, N.B.; Bommineni, Y.R.; Dai, G.; Jiang, W.; Lamont, S.; Lillehoj, H.S.; Beker, A.; Teeter, R.G.; et al. Butyrate enhances disease resistance of chickens by inducing antimicrobial host defense peptide gene expression. PLoS ONE 2011, 6, e27225. [Google Scholar] [CrossRef] [PubMed]
- Erf, G.F. Cell-mediated immunity in poultry. Poult. Sci. 2004, 83, 580–590. [Google Scholar] [CrossRef]
- Kim, W.H.; Chaudhari, A.A.; Lillehoj, H.S. Involvement of T cell immunity in avian coccidiosis. Front. Immunol. 2019, 10, 2732. [Google Scholar] [CrossRef] [PubMed]
- Arstila, T.P.; Vainio, O.; Lassila, O. Central role of CD4+ T cells in avian immune response. Poult. Sci. 1994, 73, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, M.; Abualrous, E.T.; Sticht, J.; Alvaro-Benito, M.; Stolzenberg, S.; Noe, F.; Freund, C. Major histocompatibility complex (MHC) class I and MHC class II proteins: Conformational plasticity in antigen presentation. Front. Immunol. 2017, 8, 292. [Google Scholar] [CrossRef]
- Dong, C.; Juedes, A.E.; Temann, U.A.; Shresta, S.; Allison, J.P.; Ruddle, N.H.; Flavell, R.A. ICOS co-stimulatory receptor is essential for T-cell activation and function. Nature 2001, 409, 97–101. [Google Scholar] [CrossRef]
- Min, W.; Kim, W.H.; Lillehoj, E.P.; Lillehoj, H.S. Recent progress in host immunity to avian coccidiosis: IL-17 family cytokines as sentinels of the intestinal mucosa. Dev. Comp. Immunol. 2013, 41, 418–428. [Google Scholar] [CrossRef]
- Breed, D.G.J.; Dorrestein, J.; Vermeulen, A.N. Immunity to Eimeria tenella in chickens: Phenotypical and functional changes in peripheral blood T-cell subsets. Avian Dis. 1996, 40, 37–48. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Bacon, L.D. Increase of intestinal intraepithelial lymphocytes expressing CD8 antigen following challenge infection with Eimeria acervulina. Avian Dis. 1991, 35, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Rose, M.E.; Hesketh, P.; Wakelin, D. Immune control of murine coccidiosis: CD4+ and CD8+ T lymphocytes contribute differentially in resistance to primary and secondary infections. Parasitology 1992, 105, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.A.A.; Song, X.; Xu, L.; Yan, R.; Song, H.; Ruirui, Z.; Chengyu, L.; Li, X. The DNA-induced protective immunity with chicken interferon gamma against poultry coccidiosis. Parasitol. Res. 2010, 107, 747–750. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.S.; Choi, K.D. Recombinant chicken interferon-gamma-mediated inhibition of Eimeria tenella development in vitro and reduction of oocyst production and body weight loss following Eimeria acervulina challenge infection. Avian Dis. 1998, 42, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.S. Lymphocytes involved in cell-mediated immune responses and methods to assess cell-mediated immunity. Poult. Sci. 1991, 70, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Beura, L.K.; Mitchell, J.S.; Thompson, E.A.; Schenkel, J.M.; Mohammed, J.; Wijeyesinghe, S.; Fonseca, R.; Burbach, B.J.; Hickman, H.D.; Vezys, V.; et al. Intravital mucosal imaging of CD8+ resident memory T cells shows tissue-autonomous recall responses that amplify secondary memory article. Nat. Immunol. 2018, 19, 173–182. [Google Scholar] [CrossRef]
- Wang, S.; Suo, X. Still naïve or primed: Anticoccidial vaccines call for memory. Exp. Parasitol. 2020, 216, 107945. [Google Scholar] [CrossRef]
- Smith, A.L.; Hayday, A.C. Genetic dissection of primary and secondary responses to a widespread natural pathogen of the gut, Eimeria vermiformis. Infect. Immun. 2000, 68, 6273–6280. [Google Scholar] [CrossRef]
- Sheridan, B.S.; Romagnoli, P.A.; Pham, Q.M.; Fu, H.H.; Alonzo, F.; Schubert, W.D.; Freitag, N.E.; Lefrançois, L. γδ T Cells exhibit multifunctional and protective memory in intestinal tissues. Immunity 2013, 39, 184–195. [Google Scholar] [CrossRef]
- Maes, M. Depression is an inflammatory disease, but cell-mediated immune activation is the key component of depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 664–675. [Google Scholar] [CrossRef]
- Chai, J.Y.; Lillehoj, H.S. Isolation and functional characterization of chicken intestinal intra-epithelial lymphocytes showing natural killer cell activity against tumour target cells. Immunology 1988, 63, 111–117. [Google Scholar] [PubMed]
- Lam, K.M.; Linna, T.J. Transfer of natural resistance to Marek’s disease (JMV) with non-immune spleen cells. I. Studies of cell population transferring resistance. Int. J. Cancer 1979, 24, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Leibold, W.; Janotte, G.; Peter, H.H. Spontaneous Cell-mediated Cytotoxicity (SCMC) in Various Mammalian Species and Chickens: Selective Reaction Pattern and Different Mechanisms. Scand. J. Immunol. 1980, 11, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Boo, S.Y.; Tan, S.W.; Alitheen, N.B.; Ho, C.L.; Omar, A.R.; Yeap, S.K. Identification of reference genes in chicken intraepithelial lymphocyte natural killer cells infected with very-virulent infectious bursal disease virus. Sci. Rep. 2020, 10, 8561. [Google Scholar] [CrossRef] [PubMed]
- Gobel, T.W.F.; Kaspers, B.; Stangassinger, M. NK and T cells constitute two major, functionally distinct intestinal epithelial lymphocyte subsets in the chicken. Int. Immunol. 2001, 13, 757–762. [Google Scholar] [CrossRef]
- Lillehoj, H.S. Intestinal intraepithelial and splenic natural killer cell responses to Eimeria infections in inbred chickens. Infect. Immun. 1989, 57, 1879–1884. [Google Scholar] [CrossRef]
- Cornelissen, J.B.W.J.; Swinkels, W.J.C.; Boersma, W.A.; Rebel, J.M.J. Host response to simultaneous infections with Eimeria acervulina, maxima and tenella: A cumulation of single responses. Vet. Parasitol. 2009, 162, 58–66. [Google Scholar] [CrossRef]
- Shoaib, M.; Xiaokai, S.; UL-hasan, M.; Zafar, A.; Riaz, A.; Umar, S.; Ali shah, M.; Xiangrui, L. Role of dendritic cells in immunity against avian coccidiosis. Worlds Poult. Sci. J. 2017, 73, 737–746. [Google Scholar] [CrossRef]
- Zmrhal, V.; Slama, P. Immunomodulation of avian dendritic cells under the induction of prebiotics. Animals 2020, 10, 698. [Google Scholar] [CrossRef]
- Steinman, R.M.; Hemmi, H. Dendritic cells: Translating innate to adaptive immunity. Curr. Top. Microbiol. Immunol. 2006, 311, 17–58. [Google Scholar] [CrossRef]
- Liu, K. Dendritic Cells. Encycl. Cell Biol. 2016, 3, 741–749. [Google Scholar] [CrossRef]
- Delcayre, A.; Le Pecq, J.B. Exosomes as novel therapeutic nanodevices. Curr. Opin. Mol. Ther. 2006, 8, 31–38. [Google Scholar] [PubMed]
- del Cacho, E.; Gallego, M.; Lee, S.H.; Lillehoj, H.S.; Quilez, J.; Lillehoj, E.P.; Sánchez-Acedo, C. Induction of protective immunity against Eimeria tenella, Eimeria maxima, and Eimeria acervulina infections using dendritic cell-derived exosomes. Infect. Immun. 2012, 80, 1909–1916. [Google Scholar] [CrossRef] [PubMed]
- Colino, J.; Snapper, C.M. Exosomes from bone marrow dendritic cells pulsed with diphtheria toxoid preferentially induce type 1 antigen-specific IgG responses in naive recipients in the absence of free antigen. J. Immunol. 2006, 177, 3757–3762. [Google Scholar] [CrossRef]
- Schnitzer, J.K.; Berzel, S.; Fajardo-Moser, M.; Remer, K.A.; Moll, H. Fragments of antigen-loaded dendritic cells (DC) and DC-derived exosomes induce protective immunity against Leishmania major. Vaccine 2010, 28, 5785–5793. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lillehoj, H.S.; Lillehoj, E.P.; Lee, S.H. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following Eimeria maxima infection of chickens. Vet. Immunopathol. 2006, 114, 259–272. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lillehoj, H.S.; Lee, S.H.; Dalloul, R.A.; Lillehoj, E.P. Analysis of chicken cytokine and chemokine gene expression following Eimeria acervulina and Eimeria tenella infections. Vet. Immunol. Immunopathol. 2006, 114, 209–223. [Google Scholar] [CrossRef]
- Degen, W.G.J.; van Daal, N.; van Zuilekom, H.I.; Burnside, J.; Schijns, V.E.J.C. Identification and molecular cloning of functional chicken IL-12. J. Immunol. 2004, 172, 4371–4380. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lillehoj, H.S.; Lee, S.H.; Park, D.W.; Lillehoj, E.P. Molecular cloning and characterization of chicken lipopolysaccharide-induced TNF-α factor (LITAF). Dev. Comp. Immunol. 2006, 30, 919–929. [Google Scholar] [CrossRef]
- Jakowlew, S.B.; Dillard, P.J.; Winokur, T.S.; Flanders, K.C.; Sporn, M.B.; Roberts, A.B. Expression of transforming growth factor-βs 1–4 in chicken embryo chondrocytes and myocytes. Dev. Biol. 1991, 143, 135–148. [Google Scholar] [CrossRef]
- Jeong, J.; Kim, W.H.; Yoo, J.; Lee, C.; Kim, S.; Cho, J.H.; Jang, H.K.; Kim, D.W.; Lillehoj, H.S.; Min, W. Identification and comparative expression analysis of interleukin 2/15 receptor β chain in chickens infected with E. tenella. PLoS ONE 2012, 7, e37704. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, L.; Young, J.R.; Zoorob, R.; Whittaker, C.A.; Hesketh, P.; Archer, A.; Smith, A.L.; Kaiser, P. Cloning and Characterization of Chicken IL-10 and Its Role in the Immune Response to Eimeria maxima. J. Immunol. 2004, 173, 2675–2682. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Puehler, F.; Baeuerle, D.; Elvers, S.; Staeheli, P.; Kaspers, B.; Weining, K.C. cDNA cloning of biologically active chicken interleukin-18. J. Interf. Cytokine Res. 2000, 20, 879–883. [Google Scholar] [CrossRef]
- Yoo, J.; Jang, S.I.; Kim, S.; Cho, J.H.; Lee, H.J.; Rhee, M.H.; Lillehoj, H.S.; Min, W. Molecular characterization of duck interleukin-17. Vet. Immunol. Immunopathol. 2009, 132, 318–322. [Google Scholar] [CrossRef]
- Suo, X.; Lillehoj, H.S.; Tuo, W.; Wu, Z. Immunoparasitology: A Unique Interplay Between Host and Pathogen. Poult. Sci. 1991, 1365–3024. [Google Scholar]
- Zhang, L.; Liu, R.; Song, M.; Hu, Y.; Pan, B.; Cai, J.; Wang, M. Eimeria tenella: Interleukin 17 contributes to host immunopathology in the gut during experimental infection. Exp. Parasitol. 2013, 133, 121–130. [Google Scholar] [CrossRef]
- Bremner, A.; Kim, S.; Morris, K.M.; Nolan, M.J.; Borowska, D.; Wu, Z.; Tomley, F.; Blake, D.P.; Hawken, R.; Kaiser, P.; et al. Kinetics of the cellular and transcriptomic response to eimeria maxima in relatively resistant and susceptible chicken lines. Front. Immunol. 2021, 12, 882. [Google Scholar] [CrossRef]
- Rose, M.E.; Wakelin, D.; Hesketh, P. Interferon-gamma-mediated effects upon immunity to coccidial infections in the mouse. Parasite Immunol. 1991, 13, 63–74. [Google Scholar] [CrossRef]
- Smith, A.L.; Hayday, A.C. Genetic analysis of the essential components of the immunoprotective response to infection with Eimeria vermiformis. Int. J. Parasitol. 1998, 28, 1061–1069. [Google Scholar] [CrossRef]
- Choi, K.D.; Lillehoj, H.S.; Zalenga, D.S. Changes in local IFN-γ and TGF-β4 mRNA expression and intraepithelial lymphocytes following Eimeria acervulina infection. Vet. Immunol. Immunopathol. 1999, 71, 263–275. [Google Scholar] [CrossRef]
- Breed, D.G.J.; Dorrestein, J.; Schetters, T.P.M.; Waart, L.V.D.; Rijke, E.; Vermeulen, A.N. Peripheral blood lymphocytes from Eimeria tenella infected chickens produce gamma-interferon after stimulation in vitro. Parasite Immunol. 1997, 19, 127–135. [Google Scholar] [CrossRef]
- Breed, D.G.J.; Schetters, T.P.M.; Verhoeven, N.A.P.; Vermeulen, A.N. Characterization of phenotype related responsiveness of peripheral blood lymphocytes from Eimeria tenella infected chickens. Parasite Immunol. 1997, 19, 563–569. [Google Scholar] [CrossRef]
- Lowenthal, J.W.; York, J.J.; O’Neil, T.E.; Rhodes, S.; Prowse, S.J.; Strom, A.D.G.; Digby, M.R. In vivo effects of chicken interferon-γ during infection with Eimeria. J. Interf. Cytokine Res. 1997, 17, 551–558. [Google Scholar] [CrossRef]
- Dimier, I.H.; Quéré, P.; Naciri, M.; Bout, D.T. Inhibition of Eimeria tenella development in vitro mediated by chicken macrophages and fibroblasts treated with chicken cell supernatants with IFN-γ activity. Avian Dis. 1998, 42, 239–247. [Google Scholar] [CrossRef]
- Yamada, T.; Fujieda, S.; Yanagi, S.; Yamamura, H.; Inatome, R.; Yamamoto, H.; Igawa, H.; Saito, H. IL-1 induced chemokine production through the association of syk with TNF receptor-associated factor-6 in nasal fibroblast lines. J. Immunol. 2001, 167, 283–288. [Google Scholar] [CrossRef]
- Gracie, J.A. Interleukin-18 as a potential target in inflammatory arthritis. Clin. Exp. Immunol. 2004, 136, 402–404. [Google Scholar] [CrossRef]
- Laurent, F.; Mancassola, R.; Lacroix, S.; Menezes, R.; Naciri, M. Analysis of chicken mucosal immune response to Eimeria tenella and Eimeria maxima infection by quantitative reverse transcription-PCR. Infect. Immun. 2001, 69, 2527–2534. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Bliss, T.W.; Hong, Y.H.; Ben-Chouikha, I.; Park, D.W.; Keeler, C.L.; Lillehoj, H.S. Unique responses of the avian macrophage to different species of Eimeria. Mol. Immunol. 2007, 44, 558–566. [Google Scholar] [CrossRef]
- Sundick, R.S.; Gill-Dixon, C. A cloned chicken lymphokine homologous to both mammalian IL-2 and IL-15. J. Immunol. 1997, 159, 720–725. [Google Scholar]
- Lillehoj, H.S.; Min, W.; Choi, K.D.; Babu, U.S.; Burnside, J.; Miyamoto, T.; Rosenthal, B.M.; Lillehoj, E.P. Molecular, cellular, and functional characterization of chicken cytokines homologous to mammalian IL-15 and IL-2. Vet. Immunol. Immunopathol. 2001, 82, 229–244. [Google Scholar] [CrossRef]
- Ding, X.; Lillehoj, H.S.; Quiroz, M.A.; Bevensee, E.; Lillehoj, E.P. Protective immunity against Eimeria acervulina following in ovo immunization with a recombinant subunit vaccine and cytokine genes. Infect. Immun. 2004, 72, 6939–6944. [Google Scholar] [CrossRef]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 6th ed.; Saunders Elsevier Inc.: Philadelphia, PA, USA, 2010. [Google Scholar]
- Lynagh, G.R.; Bailey, M.; Kaiser, P. Interleukin-6 is produced during both murine and avian Eimeria infections. Vet. Immunol. Immunopathol. 2000, 76, 89–102. [Google Scholar] [CrossRef]
- Morita, Y.; Yamamura, M.; Kawashima, M.; Aita, T.; Harada, S.; Okamoto, H.; Inoue, H.; Makino, H. Differential in vitro effects of IL-4, IL-10, and IL-13 on proinflammatory cytokine production and fibroblast proliferation in rheumatoid synovium. Rheumatol. Int. 2001, 20, 49–54. [Google Scholar] [CrossRef]
- Gazzinelli, R.T.; Wysocka, M.; Hieny, S.; Scharton-Kersten, T.; Cheever, A.; Kühn, R.; Müller, W.; Trinchieri, G.; Sher, A. In the absence of endogenous IL-10, mice acutely infected with Toxoplasma gondii succumb to a lethal immune response dependent on CD4+ T cells and accompanied by overproduction of IL-12, IFN-gamma and TNF-alpha. J. Immunol. 1996, 157, 798–805. [Google Scholar]
- Morris, A.; Shanmugasundaram, R.; McDonald, J.; Selvaraj, R.K. Effect of in vitro and in vivo 25-hydroxyvitamin D treatment on macrophages, T cells, and layer chickens during a coccidia challenge. J. Anim. Sci. 2015, 93, 2894–2903. [Google Scholar] [CrossRef]
- Zhang, S.; Lillehoj, H.S.; Ruff, M.D. Chicken tumor necrosis-like factor. I. In vitro production by macrophages stimulated with Eimeria tenella or bacterial lipopolysaccharide. Poult. Sci. 1995, 74, 1304–1310. [Google Scholar] [CrossRef]
- Zhang, S.; Lillehoj, H.S.; Ruff, M.D. In vivo role of tumor necrosis-like factor in Eimeria tenella infection. Avian Dis. 1995, 39, 859–866. [Google Scholar] [CrossRef]
- Ebnet, K.; Vestweber, D. Molecular Mechanisms that Control Leukocyte Extravasation: The Selectins and the Chemokines. Histochem. Cell Biol. 1999, 112, 1–23. [Google Scholar] [CrossRef]
- Oppenheim, J.J.; Zachariae, C.O.C.; Mukaida, N.; Matsushima, K. Properties of the novel proinflammatory supergene “intercrine” cytokine family. Annu. Rev. Immunol. 1991, 9, 617–648. [Google Scholar] [CrossRef]
- Kaiser, P.; Poh, T.Y.; Rothwell, L.; Avery, S.; Balu, S.; Pathania, U.S.; Hughes, S.; Goodchild, M.; Morrell, S.; Watson, M.; et al. A genomic analysis of chicken cytokines and chemokines. J. Interf. Cytokine Res. 2005, 25, 467–484. [Google Scholar] [CrossRef]
- Siveke, J.T.; Hamann, A. T helper 1 and T helper 2 cells respond differentially to chemokines. J. Immunol. 1998, 160, 550–554. [Google Scholar]
- Zhao, Y.; Bao, Y.; Zhang, L.; Chang, L.; Jiang, L.; Liu, Y.; Zhang, L.; Qin, J. Biosafety of the plasmid pcDNA3-1E of Eimeria acervulina in chicken. Exp. Parasitol. 2013, 133, 231–236. [Google Scholar] [CrossRef]
- Carrasco, J.M.D.; Casanova, N.A.; Miyakawa, M.E.F. Microbiota, gut health and chicken productivity: What is the connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef]
- Stanley, D.; Denman, S.E.; Hughes, R.J.; Geier, M.S.; Crowley, T.M.; Chen, H.; Haring, V.R.; Moore, R.J. Intestinal microbiota associated with differential feed conversion efficiency in chickens. Appl. Microbiol. Biotechnol. 2012, 96, 1361–1369. [Google Scholar] [CrossRef]
- Stanley, D.; Geier, M.S.; Denman, S.E.; Haring, V.R.; Crowley, T.M.; Hughes, R.J.; Moore, R.J. Identification of chicken intestinal microbiota correlated with the efficiency of energy extraction from feed. Vet. Microbiol. 2013, 164, 85–92. [Google Scholar] [CrossRef]
- Yan, W.; Sun, C.; Yuan, J.; Yang, N. Gut metagenomic analysis reveals prominent roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Sci. Rep. 2017, 7, 45308. [Google Scholar] [CrossRef]
- Benson, A.; Pifer, R.; Behrendt, C.L.; Hooper, L.V.; Yarovinsky, F. Gut commensal bacteria direct a protective immune response against toxoplasma gondii. Cell Host Microbe 2009, 6, 187–196. [Google Scholar] [CrossRef]
- Lazar, V.; Ditu, L.M.; Pircalabioru, G.G.; Gheorghe, I.; Curutiu, C.; Holban, A.M.; Picu, A.; Petcu, L.; Chifiriuc, M.C. Aspects of gut microbiota and immune system interactions in infectious diseases, immunopathology, and cancer. Front. Immunol. 2018, 9, 1830. [Google Scholar] [CrossRef]
- Huang, G.; Tang, X.; Bi, F.; Hao, Z.; Han, Z.; Suo, J.; Zhang, S.; Wang, S.; Duan, C.; Yu, Z.; et al. Eimeria tenella infection perturbs the chicken gut microbiota from the onset of oocyst shedding. Vet. Parasitol. 2018, 258, 30–37. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Nolan, M.J.; Harman, K.; Boulton, K.; Hume, D.A.; Tomley, F.M.; Stabler, R.A.; Blake, D.P. Effects of Eimeria tenella infection on chicken caecal microbiome diversity, exploring variation associated with severity of pathology. PLoS ONE 2017, 12, e0184890. [Google Scholar] [CrossRef]
- Borda-Molina, D.; Seifert, J.; Camarinha-Silva, A. Current Perspectives of the Chicken Gastrointestinal Tract and Its Microbiome. Comput. Struct. Biotechnol. J. 2018, 16, 131–139, 101016/jcsbj201803002. [Google Scholar] [CrossRef]
- Chen, H.L.; Zhao, X.Y.; Zhao, G.X.; Huang, H.B.; Li, H.R.; Shi, C.W.; Yang, W.T.; Jiang, Y.L.; Wang, J.Z.; Ye, L.P.; et al. Dissection of the cecal microbial community in chickens after Eimeria tenella infection. Parasites Vectors 2020, 13, 56. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Scapini, L.B.; Ribeiro, M.V.; Pivetta, M.R.; Buzim, R.; Fernandes, J.I.M. Effects of β-mannanase supplementation on the intestinal microbiota composition of broiler chickens challenged with a coccidiosis vaccine. Livest. Sci. 2019, 228, 187–194. [Google Scholar] [CrossRef]
- Greppi, A.; Asare, P.T.; Schwab, C.; Zemp, N.; Stephan, R.; Lacroix, C. Isolation and comparative genomic analysis of eeuterin-producing Lactobacillus reuteri from the chicken gastrointestinal tract. Front. Microbiol. 2020, 11, 1166. [Google Scholar] [CrossRef]
- Tierney, J.; Gowing, H.; Van Sinderen, D.; Flynn, S.; Stanley, L.; McHardy, N.; Hallahan, S.; Mulcahy, G. In vitro inhibition of Eimeria tenella invasion by indigenous chicken Lactobacillus species. Vet. Parasitol. 2004, 122, 171–182. [Google Scholar] [CrossRef]
- Cui, N.; Wang, X.; Wang, Q.; Li, H.; Wang, F.; Zhao, X. Effect of dual infection with Eimeria tenella and subgroup J Avian leukosis virus on the cecal microbiome in specific-pathogen-free chicks. Front. Vet. Sci. 2017, 4, 177. [Google Scholar] [CrossRef]
- Biggs, P.; Parsons, C.M. The effects of several organic acids on growth performance, nutrient digestibilities, and cecal microbial populations in young chicks. Poult. Sci. 2008, 87, 2581–2589. [Google Scholar] [CrossRef]
- Hessenberger, S.; Schatzmayr, G.; Teichmann, K. In vitro inhibition of Eimeria tenella sporozoite invasion into host cells by probiotics. Vet. Parasitol. 2016, 229, 93–98. [Google Scholar] [CrossRef]
- Chapman, H.D. Biochemical, genetic and applied aspects of drug resistance in Eimeria parasites of the fowl. Avian Pathol. 1997, 26, 221–244. [Google Scholar] [CrossRef]
- Chapman, H.D. Anticoccidial drugs and their effects upon the development of immunity to Eimeria infections in poultry. Avian Pathol. 1999, 28, 521–535. [Google Scholar] [CrossRef]
- Pastor-Fernández, I.; Kim, S.; Marugán-Hernández, V.; Soutter, F.; Tomley, F.M.; Blake, D.P. Vaccination with transgenic Eimeria tenella expressing Eimeria maxima AMA1 and IMP1 confers partial protection against high-level E. maxima challenge in a broiler model of coccidiosis. Parasites Vectors 2020, 13, 343. [Google Scholar] [CrossRef]
- Blake, D.P.; Pastor-Fernández, I.; Nolan, M.J.; Tomley, F.M. Recombinant anticoccidial vaccines—A cup half full? Infect. Genet. Evol. 2017, 55, 358–365. [Google Scholar] [CrossRef]
- Liu, L.; Huang, X.; Liu, J.; Li, W.; Ji, Y.; Tian, D.; Tian, L.; Yang, X.; Xu, L.; Yan, R.; et al. Identification of common immunodominant antigens of Eimeria tenella, Eimeria acervulina and Eimeria maxima by immunoproteomic analysis. Oncotarget 2017, 8, 34935–34945. [Google Scholar] [CrossRef]
- Lin, R.Q.; Lillehoj, H.S.; Lee, S.K.; Oh, S.; Panebra, A.; Lillehoj, E.P. Vaccination with Eimeria tenella elongation factor-1α recombinant protein induces protective immunity against E. tenella and E. maxima infections. Vet. Parasitol. 2017, 243, 79–84. [Google Scholar] [CrossRef]
- Tian, L.; Li, W.; Huang, X.; Tian, D.; Liu, J.; Yang, X.; Liu, L.; Yan, R.; Xu, L.; Li, X.; et al. Protective efficacy of coccidial common antigen glyceraldehyde 3-phosphate dehydrogenase (GAPDH) against challenge with three Eimeria species. Front. Microbiol. 2017, 8, 1245. [Google Scholar] [CrossRef]
- Liu, T.; Huang, J.; Ehsan, M.; Wang, S.; Fei, H.; Zhou, Z.; Song, X.; Yan, R.; Xu, L.; Li, X. Protective immunity against Eimeria maxima induced by vaccines of Em14-3-3 antigen. Vet. Parasitol. 2018, 253, 79–86. [Google Scholar] [CrossRef]
- Klotz, C.; Gehre, F.; Lucius, R.; Pogonka, T. Identification of Eimeria tenella genes encoding for secretory proteins and evaluation of candidates by DNA immunisation studies in chickens. Vaccine 2007, 25, 6625–6634. [Google Scholar] [CrossRef]
- Rafiqi, S.I.; Garg, R.; Reena, K.K.; Ram, H.; Singh, M.; Banerjee, P.S. Immune response and protective efficacy of Eimeria tenella recombinant refractile body protein, EtSO7, in chickens. Vet. Parasitol. 2018, 258, 108–113. [Google Scholar] [CrossRef]
- Huang, J.; Liu, T.; Li, K.; Song, X.; Yan, R.; Xu, L.; Li, X. Proteomic analysis of protein interactions between Eimeria maxima sporozoites and chicken jejunal epithelial cells by shotgun LC-MS/MS. Parasites Vectors 2018, 11, 226. [Google Scholar] [CrossRef]
- Wallach, M. The importance of transmission-blocking immunity in the control of infections by apicomplexan parasites. Int. J. Parasitol. 1997, 27, 1159–1167. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Park, M.S.; Cha, S.R.; Lillehoj, E.P.; Subramanian, B.M.; Sriraman, R.; Srinivasan, V.A. Eimeria maxima recombinant Gam82 gametocyte antigen vaccine protects against coccidiosis and augments humoral and cell-mediated immunity. Vaccine 2010, 28, 2980–2985. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Tao, J. Efficacy of a DNA vaccine carrying Eimeria maxima Gam56 antigen gene against coccidiosis in chickens. Korean J. Parasitol. 2013, 51, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Kota, S.; Subramanian, M.; Shanmugaraj, B.M.; Challa, H.; NM, P.; VA, S.; Sriraman, R. Subunit vaccine based on plant expressed recombinant Eimeria Gametocyte antigen Gam82 elicit protective immune response against chicken coccidiosis. J. Vaccines Vaccin. 2017, 8, 1000374. [Google Scholar] [CrossRef]
- Liu, D.; Cao, L.; Zhu, Y.; Deng, C.; Su, S.; Xu, J.; Jin, W.; Li, J.; Wu, L.; Tao, J. Cloning and characterization of an Eimeria necatrix gene encoding a gametocyte protein and associated with oocyst wall formation. Parasites Vectors 2014, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Rafiqi, S.I.; Garg, R.; Ram, H.; Reena, K.K.; Asari, M.; Kumari, P.; Kundave, V.R.; Singh, M.; Banerjee, P.S. Immunoprophylactic evaluation of recombinant gametocyte 22 antigen of Eimeria tenella in broiler chickens. Parasitol. Res. 2019, 118, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Li, Y.; Zhou, Y.; Zhao, J.; Fang, R. In vivo immunoprotective comparison between recombinant protein and DNA vaccine of Eimeria tenella surface antigen 4. Vet. Parasitol. 2020, 278, 109032. [Google Scholar] [CrossRef]
- Min, W.; Lillehoj, H.S.; Burnside, J.; Weining, K.C.; Staeheli, P.; Zhu, J.J. Adjuvant effects of IL-1β, IL-2, IL-8, IL-15, IFN-α, IFN-γ TGF-β4 and lymphotactin on DNA vaccination against Eimeria acervulina. Vaccine 2001, 20, 267–274. [Google Scholar] [CrossRef]
- Venkatas, J.; Adeleke, M.A. A review of Eimeria antigen identification for the development of novel anticoccidial vaccines. Parasitol. Res. 2019, 118, 1701–1710. [Google Scholar] [CrossRef]
- Tang, X.; Liu, X.; Yin, G.; Suo, J.; Tao, G.; Zhang, S.; Suo, X. A novel vaccine delivery model of the apicomplexan Eimeria tenella expressing Eimeria maxima antigen protects chickens against infection of the two parasites. Front. Immunol. 2018, 8, 1982. [Google Scholar] [CrossRef]
- Jang, S.I.; Kim, D.K.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Bertrand, F.; Dupuis, L.; Deville, S.; Ben Arous, J.; Lillehoj, E.P. Evaluation of MontanideTM ISA 71 VG Adjuvant during profilin vaccination against experimental Coccidiosis. PLoS ONE 2013, 8, e59786. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Jang, S.I.; Panebra, A.; Lillehoj, E.P.; Dupuis, L.; Ben Arous, J.; Lee, S.K.; Oh, S.T. In ovo vaccination using Eimeria profilin and Clostridium perfringens NetB proteins in Montanide IMS adjuvant increases protective immunity against experimentally-induced necrotic enteritis. Asian-Australas. J. Anim. Sci. 2017, 30, 1478–1485. [Google Scholar] [CrossRef]
- Ding, J.; Bao, W.; Liu, Q.; Yu, Q.; Abdille, M.H.; Wei, Z. Immunoprotection of chickens against Eimeria acervulina by recombinant α-tubulin protein. Parasitol. Res. 2008, 103, 1133–1140. [Google Scholar] [CrossRef]
- Li, W.C.; Zhang, X.K.; Du, L.; Pan, L.; Gong, P.T.; Li, J.H.; Yang, J.; Li, H.; Zhang, X.C. Eimeria maxima: Efficacy of recombinant Mycobacterium bovis BCG expressing apical membrane antigen1 against homologous infection. Parasitol. Res. 2013, 112, 3825–3833. [Google Scholar] [CrossRef]
- Jenkins, M.C.; Stevens, L.; O’Brien, C.; Parker, C.; Miska, K.; Konjufca, V. Incorporation of a recombinant Eimeria maxima IMP1 antigen into nanoparticles confers protective immunity against E. maxima challenge infection. Vaccine 2018, 36, 1126–1131. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Choi, K.D.; Jenkins, M.C.; Vakharia, V.N.; Song, K.D.; Han, J.Y.; Lillehoj, E.P. A recombinant Eimeria protein inducing interferon-γ production: Comparison of different gene expression systems and immunization strategies for vaccination against coccidiosis. Avian Dis. 2000, 44, 379–389. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Ding, X.; Dalloul, R.A.; Sato, T.; Yasuda, A.; Lillehoj, E.P. Embryo vaccination against Eimeria tenella and E. acervulina infections using recombinant proteins and cytokine adjuvants. J. Parasitol. 2005, 91, 666–673. [Google Scholar] [CrossRef]
- Wu, S.Q.; Wang, M.; Liu, Q.; Zhu, Y.J.; Suo, X.; Jiang, J.S. Construction of DNA vaccines and their induced protective immunity against experimental Eimeria tenella infection. Parasitol. Res. 2004, 94, 332–336. [Google Scholar] [CrossRef]
- Mathew, D.E.; Larsen, K.; Janeczek, P.; Lewohl, J.M. Expression of 14-3-3 transcript isoforms in response to ethanol exposure and their regulation by miRNAs. Mol. Cell. Neurosci. 2016, 75, 44–49. [Google Scholar] [CrossRef]
- Zhang, B.; Yuan, C.; Song, X.; Xu, L.; Yan, R.; Shah, M.A.A.; Guo, C.; Zhu, S.; Li, X. Optimization of Immunization Procedure for Eimeria tenella DNA Vaccine pVAX1-pEtK2-IL-2 and Its Stability. Acta Parasitol. 2019, 64, 745–752. [Google Scholar] [CrossRef]
- Lee, S.H.; Lillehoj, H.S.; Jang, S.I.; Hong, Y.H.; Min, W.; Lillehoj, E.P.; Yancey, R.J.; Dominowski, P. Embryo vaccination of chickens using a novel adjuvant formulation stimulates protective immunity against Eimeria maxima infection. Vaccine 2010, 28, 7774–7778. [Google Scholar] [CrossRef]
- Zhang, D.F.; Xu, H.; Sun, B.B.; Li, J.Q.; Zhou, Q.J.; Zhang, H.L.; Du, A.F. Adjuvant effect of ginsenoside-based nanoparticles (ginsomes) on the recombinant vaccine against Eimeria tenella in chickens. Parasitol. Res. 2012, 110, 2445–2453. [Google Scholar] [CrossRef] [PubMed]
- Trovato, M.; De Berardinis, P. Novel antigen delivery systems. World J. Virol. 2015, 4, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Wang, L.; Wang, T.; Zhang, J.; Liu, Q.; Chen, P.; Chen, Z.; Wang, F.; Li, H.; Xiao, Y.; et al. Display of Eimeria tenella EtMic2 protein on the surface of Saccharomyces cerevisiae as a potential oral vaccine against chicken coccidiosis. Vaccine 2014, 32, 1869–1876. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, L.; Li, J.; Zheng, J.; Cai, N.; Gong, P.; Li, S.; Li, H.; Zhang, X. A novel recombinant BCG vaccine encoding Eimeria tenella rhomboid and chicken IL-2 induces protective immunity against coccidiosis. Korean J. Parasitol. 2014, 52, 251–256. [Google Scholar] [CrossRef]
- Galen, J.E.; Pasetti, M.F.; Tennant, S.; Ruiz-Olvera, P.; Sztein, M.B.; Levine, M.M. Salmonella enterica serovar Typhi live vector vaccines finally come of age. Immunol. Cell Biol. 2009, 87, 400–412. [Google Scholar] [CrossRef]
- Shivaramaiah, S.; Barta, J.R.; Layton, S.L.; Lester, C.; Kwon, Y.M.; Berghman, L.R.; Hargis, B.M.; Tellez, G. Development and evaluation of an δaroA I δhtrA salmonella enteritidis vector expressing eimeria maxima trap family protein emtfp250 with CD 154 (CD 40L) as candidate vaccines against coccidiosis in broilers. Int. J. Poult. Sci. 2010, 9, 1031–1037. [Google Scholar] [CrossRef][Green Version]
- Clark, J.D.; Oakes, R.D.; Redhead, K.; Crouch, C.F.; Francis, M.J.; Tomley, F.M.; Blake, D.P. Eimeria species parasites as novel vaccine delivery vectors: Anti-Campylobacter jejuni protective immunity induced by Eimeria tenella-delivered CjaA. Vaccine 2012, 30, 2683–2688. [Google Scholar] [CrossRef]
- Vrba, V.; Pakandl, M. Host specificity of turkey and chicken Eimeria: Controlled cross-transmission studies and a phylogenetic view. Vet. Parasitol. 2015, 208, 118–124. [Google Scholar] [CrossRef]
- Shirley, M.W.; Ivens, A.; Gruber, A.; Madeira, A.M.B.N.; Wan, K.L.; Dear, P.H.; Tomley, F.M. The Eimeria genome projects: A sequence of events. Trends Parasitol. 2004, 20, 199–201. [Google Scholar] [CrossRef]
- Tang, X.; Suo, J.; Li, C.; Du, M.; Wang, C.; Hu, D.; Duan, C.; Lyu, Y.; Liu, X.; Suo, X. Transgenic Eimeria tenella expressing profilin of Eimeria maxima elicits enhanced protective immunity and alters gut microbiome of chickens. Infect. Immun. 2018, 86, e00888-17. [Google Scholar] [CrossRef]
- Marugan-Hernandez, V.; Cockle, C.; Macdonald, S.; Pegg, E.; Crouch, C.; Blake, D.P.; Tomley, F.M. Viral proteins expressed in the protozoan parasite Eimeria tenella are detected by the chicken immune system. Parasites Vectors 2016, 9, 463. [Google Scholar] [CrossRef] [PubMed]
- Shanmugaraj, B.M.; Ramalingam, S. Plant expression platform for the production of recombinant pharmaceutical proteins. Austin. J. Biotechnol. Bioeng. 2014, 1, 4–7. [Google Scholar] [CrossRef]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.; Liu, Y.; Calsamiglia, S.; Fernandez-Miyakawa, M.E.; Chi, F.; Cravens, R.L.; Oh, S.; Gay, C.G. Phytochemicals as antibiotic alternatives to promote growth and enhance host health. Vet. Res. 2018, 49, 76. [Google Scholar] [CrossRef]
- De La Mora, Z.V.; Nuño, K.; Vázquez-Paulino, O.; Avalos, H.; Castro-Rosas, J.; Gómez-Aldapa, C.; Angulo, C.; Ascencio, F.; Villarruel-López, A. Effect of a synbiotic mix on intestinal structural changes, and salmonella typhimurium and clostridium perfringens colonization in broiler chickens. Animals 2019, 9, 777. [Google Scholar] [CrossRef]
- Ren, H.; Vahjen, W.; Dadi, T.; Saliu, E.M.; Boroojeni, F.G.; Zentek, J. Synergistic effects of probiotics and phytobiotics on the intestinal microbiota in young broiler chicken. Microorganisms 2019, 7, 684. [Google Scholar] [CrossRef]
- Gadde, U.; Rathinam, T.; Lillehoj, H.S. Passive immunization with hyperimmune egg-yolk IgY as prophylaxis and therapy for poultry diseases-A review. Anim. Health Res. Rev. 2015, 16, 163–176. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.; Li, J.; Li, Y.; Wang, L.; Wang, Q.; Fang, L.; Ding, X.; Huang, P.; Yin, J.; et al. Protective effect of chicken egg yolk immunoglobulins (IgY) against enterotoxigenic Escherichia coli K88 adhesion in weaned piglets. BMC Vet. Res. 2019, 15, 234. [Google Scholar] [CrossRef]
- Vega, C.G.; Bok, M.; Ebinger, M.; Rocha, L.A.; Rivolta, A.A.; Thomas, V.G.; Muntadas, P.; D’Aloia, R.; Pinto, V.; Parreño, V.; et al. A new passive immune strategy based on IgY antibodies as a key element to control neonatal calf diarrhea in dairy farms. BMC Vet. Res. 2020, 16, 264. [Google Scholar] [CrossRef]
- Khalf, N.; El-Sawy, H.; Hanna, T.N.; El-Meneisy, A.A.; Khodeir, M.H. Efficacy of IgY immunoglobulin prepared in chicken egg yolk for the protection of chicken against necrotic enteritis. Benha Vet. Med. J. 2016, 31, 101–105. [Google Scholar] [CrossRef]
- Vandeputte, J.; Martel, A.; Canessa, S.; Van Rysselberghe, N.; De Zutter, L.; Heyndrickx, M.; Haesebrouck, F.; Pasmans, F.; Garmyn, A. Reducing Campylobacter jejuni colonization in broiler chickens by in-feed supplementation with hyperimmune egg yolk antibodies. Sci. Rep. 2019, 9, 8931. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.E. Triennial Growth Symposium: A review of science leading to host-targeted antibody strategies for preventing growth depression due to microbial colonization. J. Anim. Sci. 2011, 89, 1981–1990. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kovacs-Nolan, J.; Mine, Y. Egg yolk antibodies for passive immunity. Annu. Rev. Food Sci. Technol. 2012, 3, 163–182. [Google Scholar] [CrossRef]
- Hussein, M.A.; Rehan, I.F.; Rehan, A.F.; Eleiwa, N.Z.; Abdel-Rahman, M.A.M.; Fahmy, S.G.; Ahmed, A.S.; Youssef, M.; Diab, H.M.; Batiha, G.E.; et al. Egg Yolk IgY: A novel trend of feed additives to limit drugs and to improve poultry meat quality. Front. Vet. Sci. 2020, 7, 1–10. [Google Scholar] [CrossRef]
- Lee, S.H.; Lillehoj, H.S.; Park, D.W.; Jang, S.I.; Morales, A.; García, D.; Lucio, E.; Larios, R.; Victoria, G.; Marrufo, D.; et al. Induction of passive immunity in broiler chickens against Eimeria acervulina by hyperimmune egg yolk immunoglobulin Y. Poult. Sci. 2009, 88, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lillehoj, H.S.; Park, D.W.; Jang, S.I.; Morales, A.; García, D.; Lucio, E.; Larios, R.; Victoria, G.; Marrufo, D.; et al. Protective effect of hyperimmune egg yolk IgY antibodies against Eimeria tenella and Eimeria maxima infections. Vet. Parasitol. 2009, 163, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.J.; Ren, C.Z.; Wang, S.S.; Liu, D.D.; Cao, L.Q.; Tao, J.P. Protection efficacy of multivalent egg yolk immunoglobulin against Eimeria tenella infection in Chickens. Iran. J. Parasitol. 2013, 8, 449–458. [Google Scholar] [PubMed]
- Park, I.; Goo, D.; Nam, H.; Wickramasuriya, S.S.; Lee, K.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.G.; Lillehoj, H.S. Effects of dietary maltol on innate immunity, gut health, and growth performance of broiler chickens challenged with Eimeria maxima. Front. Vet. Sci. 2021, 8, 508. [Google Scholar] [CrossRef]
- Lee, S.; Lillehoj, H.S.; Park, D.W.; Hong, Y.H.; Lin, J.J. Effects of Pediococcus- and Saccharomyces-based probiotic (MitoMax®) on coccidiosis in broiler chickens. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 261–268. [Google Scholar] [CrossRef]
- Lee, K.W.; Lee, S.H.; Lillehoj, H.S.; Li, G.X.; Jang, S.I.; Babu, U.S.; Park, M.S.; Kim, D.K.; Lillehoj, E.P.; Neumann, A.P.; et al. Effects of direct-fed microbials on growth performance, gut morphometry, and immune characteristics in broiler chickens. Poult. Sci. 2010, 89, 203–216. [Google Scholar] [CrossRef]
- Talebi, A.; Amirzadeh, B.; Mokhtari, B.; Gahri, H. Effects of a multi-strain probiotic (PrimaLac) on performance and antibody responses to Newcastle disease virus and infectious bursal disease virus vaccination in broiler chickens. Avian Pathol. 2008, 37, 509–512. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Lillehoj, H.S.; Shellem, T.A.; Doerr, J.A. Enhanced mucosal immunity against Eimeria acervulina in broilers fed a Lactobacillus-based probiotic. Poult. Sci. 2003, 82, 62–66. [Google Scholar] [CrossRef]
- Dalloul, R.A.; Lillehoj, H.S.; Tamim, N.M.; Shellem, T.A.; Doerr, J.A. Induction of local protective immunity to Eimeria acervulina by a Lactobacillus-based probiotic. Comp. Immunol. Microbiol. Infect. Dis. 2005, 28, 351–361. [Google Scholar] [CrossRef]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult. Sci. 2020, 99, 725–733. [Google Scholar] [CrossRef]
- Slawinska, A.; Dunislawska, A.; Plowiec, A.; Radomska, M.; Lachmanska, J.; Siwek, M.; Tavaniello, S.; Maiorano, G. Modulation of microbial communities and mucosal gene expression in chicken intestines after galactooligosaccharides delivery In Ovo. PLoS ONE 2019, 14, e0212318. [Google Scholar] [CrossRef]
- Al-Sheraji, S.H.; Ismail, A.; Manap, M.Y.; Mustafa, S.; Yusof, R.M.; Hassan, F.A. Prebiotics as functional foods: A review. J. Funct. Foods 2013, 5, 1542–1553. [Google Scholar] [CrossRef]
- Muthamilselvan, T.; Kuo, T.F.; Wu, Y.C.; Yang, W.C. Herbal remedies for coccidiosis control: A review of plants, compounds, and anticoccidial actions. Evidence-based Complement. Altern. Med. 2016, 2016, 2657981. [Google Scholar] [CrossRef]
- Ganguly, S. Supplementation of prebiotics, probiotics and acids on immunity in poultry feed: A brief review. Worlds Poult. Sci. J. 2013, 69, 639–648. [Google Scholar] [CrossRef]
- Sugiharto, S. Role of nutraceuticals in gut health and growth performance of poultry. J. Saudi Soc. Agric. Sci. 2016, 15, 99–111. [Google Scholar] [CrossRef]
- Angwech, H.; Tavaniello, S.; Ongwech, A.; Kaaya, A.N.; Maiorano, G. Efficacy of in ovo delivered prebiotics on growth performance, meat quality and gut health of kuroiler chickens in the face of a natural coccidiosis challenge. Animals 2019, 9, 876. [Google Scholar] [CrossRef]
- Bozkurt, M.; Aysul, N.; Küçükyilmaz, K.; Aypak, S.; Ege, G.; Çatli, A.U.; Akşit, H.; Çöven, F.; Seyrek, K.; Çinar, M. Efficacy of in-feed preparations of an anticoccidial, multienzyme, prebiotic, probiotic, and herbal essential oil mixture in healthy and Eimeria spp.-infected broilers. Poult. Sci. 2014, 93, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Elmusharaf, M.A.; Peek, H.W.; Nollet, L.; Beynen, A.C. The effect of an in-feed mannanoligosaccharide preparation (MOS) on a coccidiosis infection in broilers. Anim. Feed Sci. Technol. 2007, 134, 347–354. [Google Scholar] [CrossRef]
- Cuperus, T.; Coorens, M.; van Dijk, A.; Haagsman, H.P. Avian host defense peptides. Dev. Comp. Immunol. 2013, 41, 352–369. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.H.; Lillehoj, H.S.; Gay, C.G. Using genomics to identify novel antimicrobials. OIE Rev. Sci. Tech. 2016, 35, 95–103. [Google Scholar] [CrossRef]
- Yount, N.Y.; Bayer, A.S.; Xiong, Y.Q.; Yeaman, M.R. Advances in antimicrobial peptide immunobiology. Biopolym. Pept. Sci. Sect. 2006, 84, 435–458. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lillehoj, H.S.; Dalloul, R.A.; Min, W.; Miska, K.B.; Tuo, W.; Lee, S.H.; Han, J.Y.; Lillehoj, E.P. Molecular cloning and characterization of chicken NK-lysin. Vet. Immunol. Immunopathol. 2006, 110, 339–347. [Google Scholar] [CrossRef]
- Lee, S.H.; Lillehoj, H.S.; Tuo, W.; Murphy, C.A.; Hong, Y.H.; Lillehoj, E.P. Parasiticidal activity of a novel synthetic peptide from the core α-helical region of NK-lysin. Vet. Parasitol. 2013, 197, 113–121. [Google Scholar] [CrossRef]
- Su, S.; Dwyer, D.M.; Miska, K.B.; Fetterer, R.H.; Jenkins, M.C.; Wong, E.A. Expression of host defense peptides in the intestine of Eimeria-challenged chickens. Poult. Sci. 2017, 96, 2421–2427. [Google Scholar] [CrossRef]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Evaluation of the immunomodulatory activity of the chicken NK-Lysin-derived peptide cNK-2. Sci. Rep. 2017, 7, 45099. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Munawar, S.H.; Manzoor, Z.; Iqbal, Z.; Khan, M.N.; Saleemi, M.K.; Zia, M.A.; Yousaf, A. Anticoccidial effects of acetic acid on performance and pathogenic parameters in broiler chickens challenged with Eimeria tenella. Pesqui. Vet. Bras. 2011, 31, 99–103. [Google Scholar] [CrossRef]
- Ali, A.M.; Seddiek, S.A.; Khater, H.F. Effect of butyrate, clopidol and their combination on the performance of broilers infected with Eimeria maxima. Br. Poult. Sci. 2014, 55, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Dittoe, D.K.; Ricke, S.C.; Kiess, A.S. Organic acids and potential for modifying the avian gastrointestinal tract and reducing pathogens and disease. Front. Vet. Sci. 2018, 5, 216–222, 103389/fvets201800216. [Google Scholar] [CrossRef] [PubMed]
- Vesteggh, H.A.J. Lactic acid has positive effect on broiler performance. World Poult. 1999, 8, 16–17. [Google Scholar]
- Kiarie, E.G.; Leung, H.; Akbari Moghaddam Kakhki, R.; Patterson, R.; Barta, J.R. Utility of feed enzymes and yeast derivatives in ameliorating deleterious effects of coccidiosis on intestinal health and function in broiler chickens. Front. Vet. Sci. 2019, 6, 473. [Google Scholar] [CrossRef]
- Leung, H.; Yitbarek, A.; Snyder, R.; Patterson, R.; Barta, J.R.; Karrow, N.; Kiarie, E. Responses of broiler chickens to Eimeria challenge when fed a nucleotide-rich yeast extract. Poult. Sci. 2019, 98, 1622–1633. [Google Scholar] [CrossRef]
- Su, S.; Miska, K.B.; Fetterer, R.H.; Jenkins, M.C.; Wong, E.A. Expression of digestive enzymes and nutrient transporters in Eimeria acervulina-challenged layers and broilers. Poult. Sci. 2014, 93, 1217–1226. [Google Scholar] [CrossRef]
- Scapini, L.B.; de Cristo, A.B.; Schmidt, J.M.; Buzim, R.; Nogueira, L.K.; Palma, S.C.; Fernandes, J.I.M. Effect of β-mannanase supplementation in conventional diets on the performance, immune competence and intestinal quality of broilers challenged with Eimeria sp. J. Appl. Poult. Res. 2019, 28, 1048–1057. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Gibbs, K.; Awati, A.; Klasing, K.C. The effects of enzymes and direct fed microbial combination on performance and immune response of broilers under a coccidia challenge. J. Appl. Anim. Nutr. 2016, 4, e6. [Google Scholar] [CrossRef]
- Kurt, T.; Wong, N.; Fowler, H.; Gay, C.; Lillehoj, H.; Plummer, P.; Scott, H.M.; Hoelzer, K. Strategic priorities for research on antibiotic alternatives in animal agriculture—Results from an expert workshop. Front. Vet. Sci. 2019, 6, 429. [Google Scholar] [CrossRef]


{kind=link}
| Target Antigens | Source (Eimeria spp.) | Administration Route | Vectors | Immune Response or Effects on Chcikens | References |
|---|---|---|---|---|---|
| EF1-α/EF2 * | E. acervulina, E. maxima, E. tenella | Immunized subcutaneously | pcDNA3.1 (+) | Increased body weight gain, improved immune response, and decreased fecal oocyst shedding | [114,115] |
| SO7 | E. tenella | Immunized intramuscularly | pcDNA3, pVR1012 | Increased body weight gain, reduced oocyst shedding, and cecal lesion score | [118,119] |
| Gam82 | E. maxima | Immunized intramuscularly | pET28a (+), pTRA-ERH | Improved immune responses, increased body weight gain, reduced oocyst shedding and gut pathology | [120,122,124] |
| Gam56 | E. maxima | Immunized intramuscularly | pcDNA3.1(zeo)+ | Improved immune responses, increased body weight gain, and decreased oocyst shedding | [120,123] |
| EtSAG4 | E. tenella | Chest intramuscular injection | pET28a | Improved cell-mediated immunity, increased average body weight, and reduced oocyst output | [127] |
| α-tubulin | E. acervulina | Immunized subcutaneously | pGEM-T | Reduced duodenal lesions | [133] |
| GAPDH * | E. acervulina, E. maxima E. tenella | Immunized intramuscularly | pSDEP2AIMP1S | Improved immune response, reduced gut lesions, increased body weight gain, and decreased oocyst shedding | [114,116] |
| Em14-3-3 * | E. maxima | Immunized subcutaneously, oral immunization | pVAX1 | Improved immune responses, decreased gut lesions, and increased body weight gain | [114,117] |
| IMP1 | E. maxima | Oral immunization | pSDEP2AIMP1S, pGEMT | Increased body weight gain, reduced parasite replication and gut lesions | [112,135] |
| AMA1 | E. maxima | Oral immunization | pSDEP2AIMP1S | Increased body weight gain, reduced Eimeria replication, and reduced gut lesions | [112,134] |
| Profilin (3-1E) | E. acervulina, E. tenella, E. maxima | in ovo immunization, immunized intramuscularly | pcDNA3.1 (+), pET32a (+), pSDEP2ARS, | Enhanced immunogenicity, increased body weight gain, and reduced gut pathology | [81,93,131,132,151] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Lu, M.; Lillehoj, H.S. Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies. Vaccines 2022, 10, 215. https://doi.org/10.3390/vaccines10020215
Lee Y, Lu M, Lillehoj HS. Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies. Vaccines. 2022; 10(2):215. https://doi.org/10.3390/vaccines10020215
Chicago/Turabian StyleLee, Youngsub, Mingmin Lu, and Hyun S. Lillehoj. 2022. "Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies" Vaccines 10, no. 2: 215. https://doi.org/10.3390/vaccines10020215
APA StyleLee, Y., Lu, M., & Lillehoj, H. S. (2022). Coccidiosis: Recent Progress in Host Immunity and Alternatives to Antibiotic Strategies. Vaccines, 10(2), 215. https://doi.org/10.3390/vaccines10020215