
Role of Physiology, Immunity, Microbiota, and Infectious Diseases in the Gut Health of Poultry

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
3. Structure and Physiology
3.1. Chicken Gut and Its Relation to Gut Health
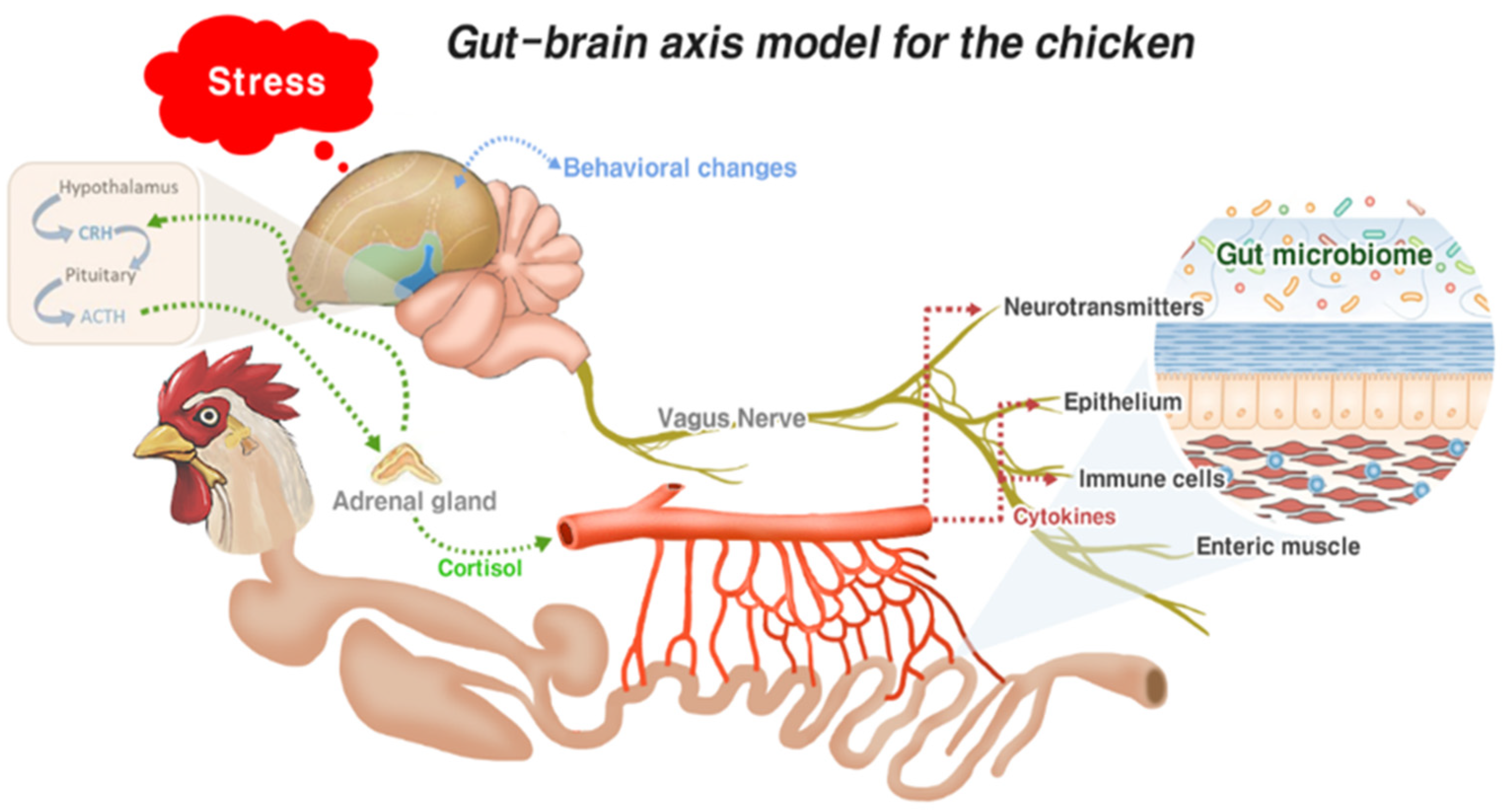
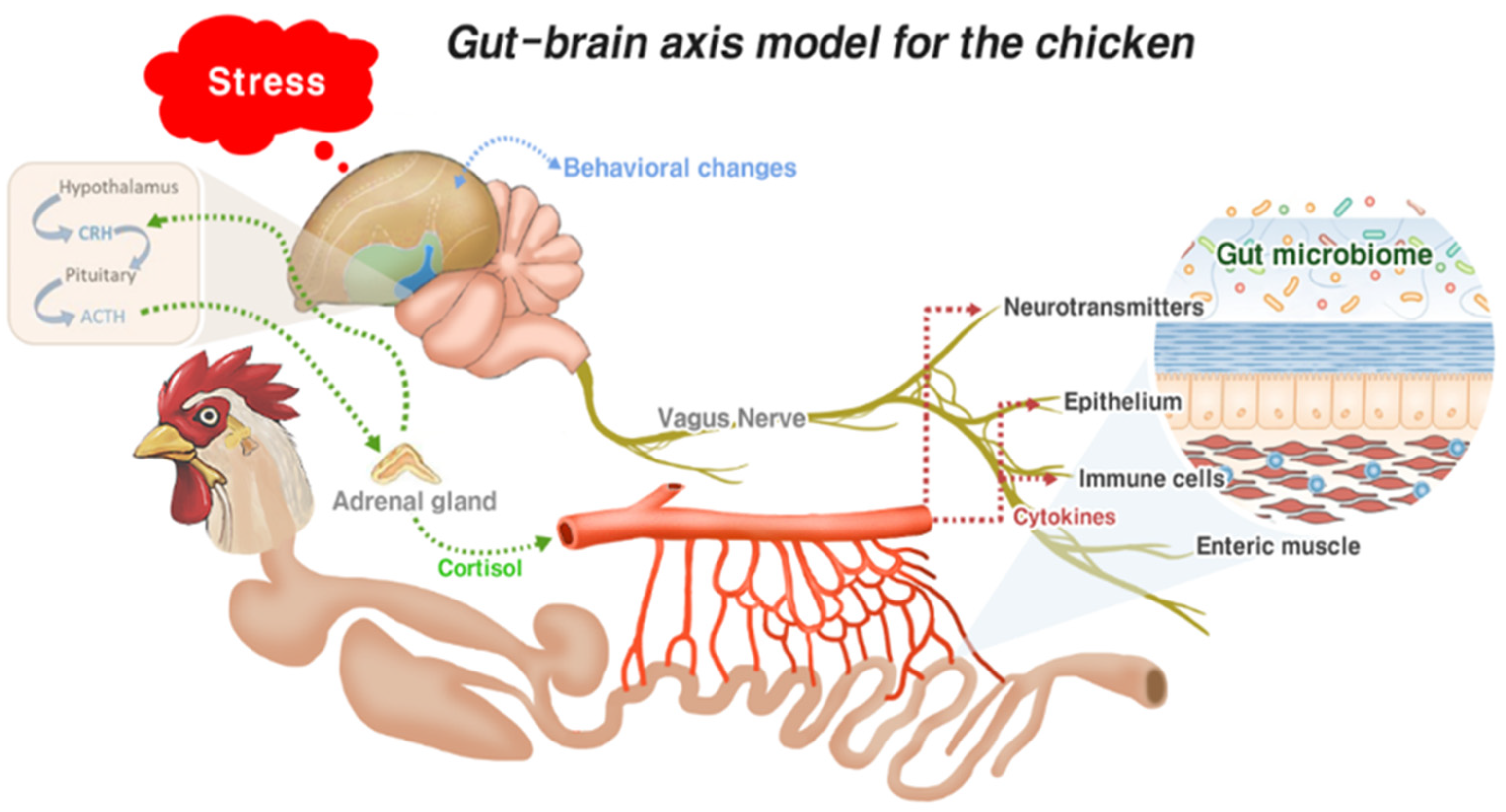
3.2. Gut–Brain Axis and Gut Health
4. Intestinal Immune System Development and Its Role in Gut Health
4.1. Development of the Chicken Gut Immune System
4.2. Development of Avian T and B Cells
4.3. Inflammation in the Gut
4.4. Mechanism of Immune Responses in the Gut
5. Microbiota and Its Role in Chicken Gut Health
The Role of Gut Microbiota in Chicken Gut Health
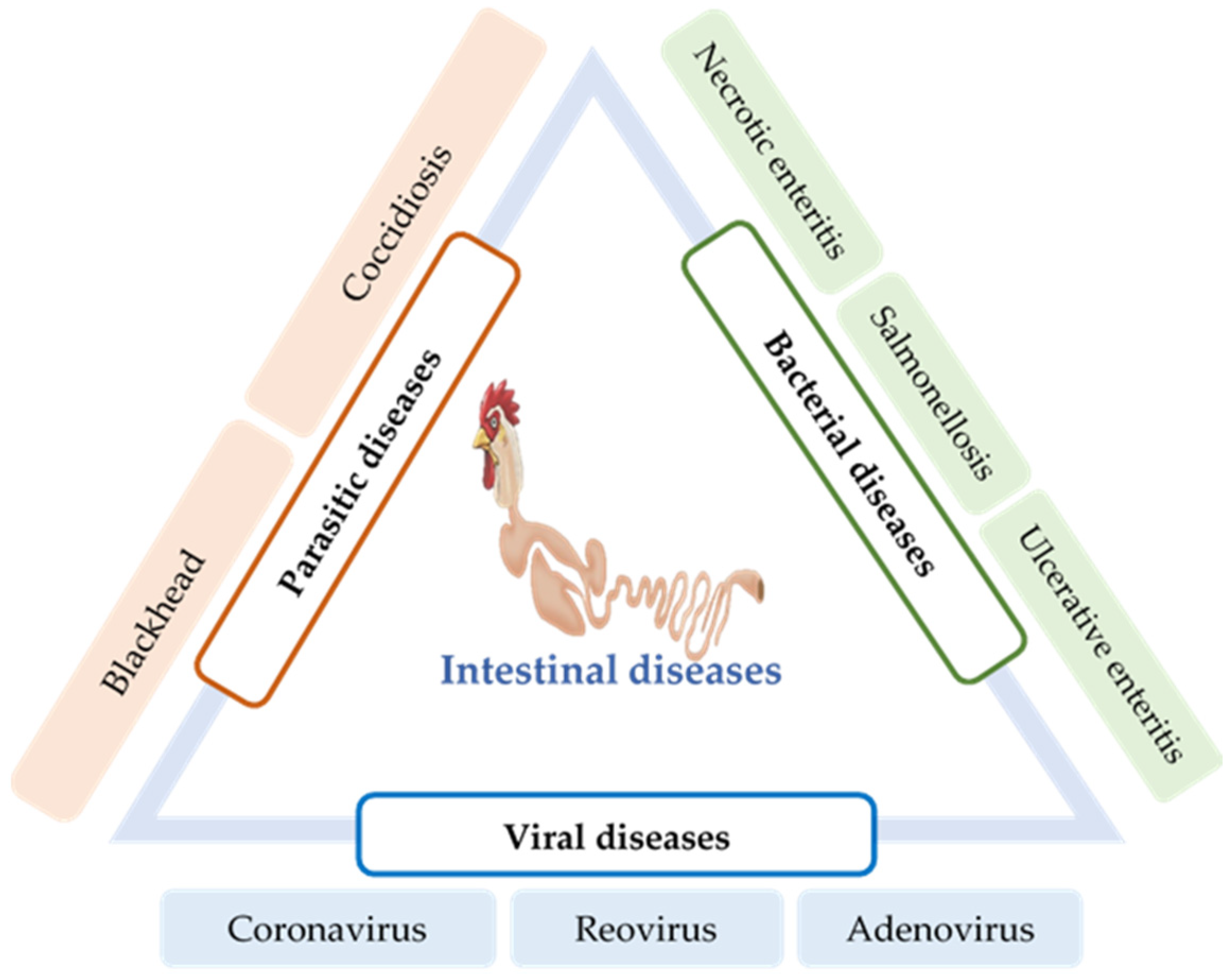
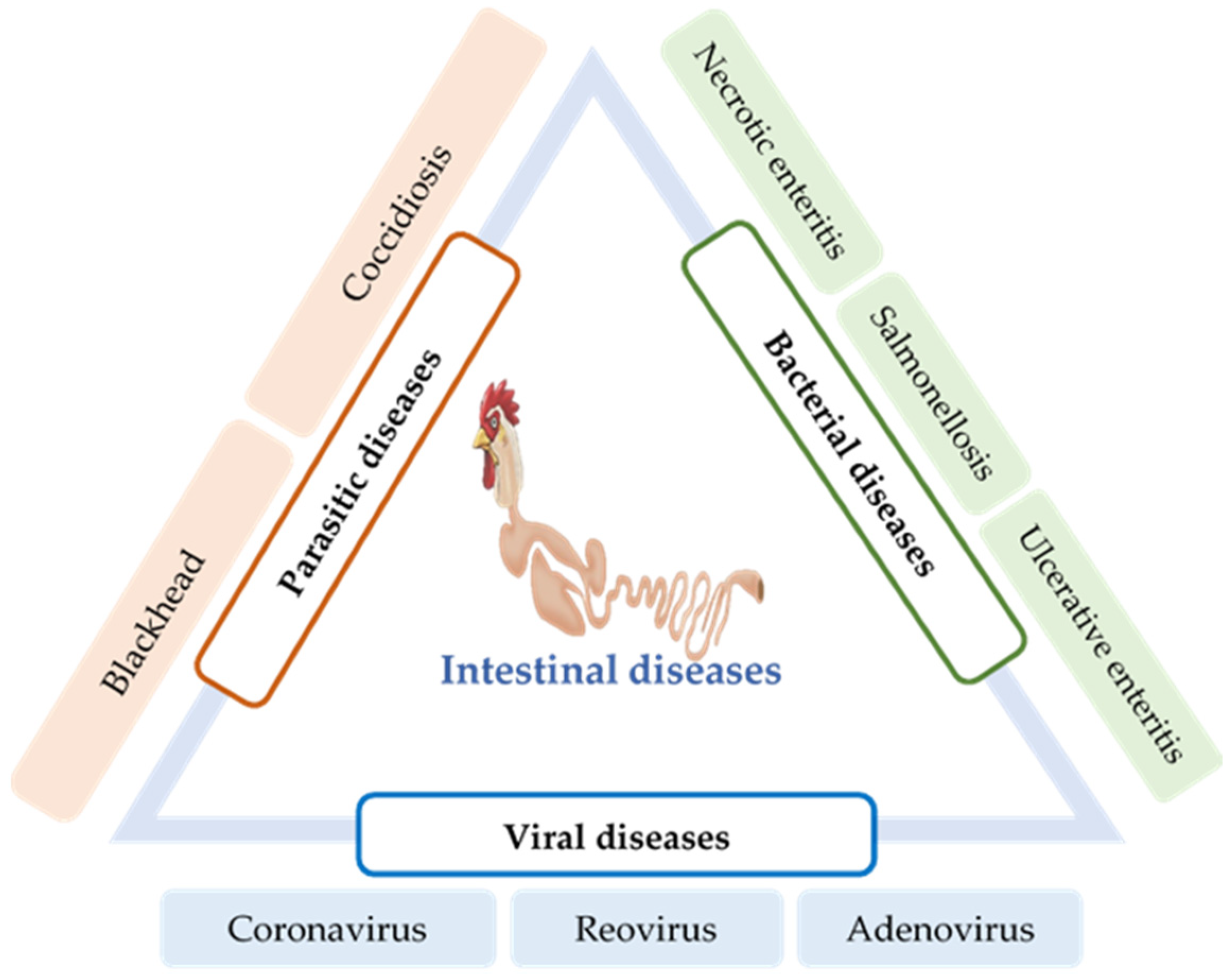
6. Intestinal Infections and Their Impact on Gut Health
6.1. Parasitic Diseases
6.1.1. Coccidiosis
6.1.2. Blackhead
6.2. Bacterial Diseases
6.2.1. Necrotic Enteritis
6.2.2. Ulcerative Enteritis
6.2.3. Salmonellosis
6.3. Viral Diseases
6.3.1. Coronavirus
6.3.2. Reovirus
6.3.3. Adenovirus/Hemorrhagic Enteritis (HE)
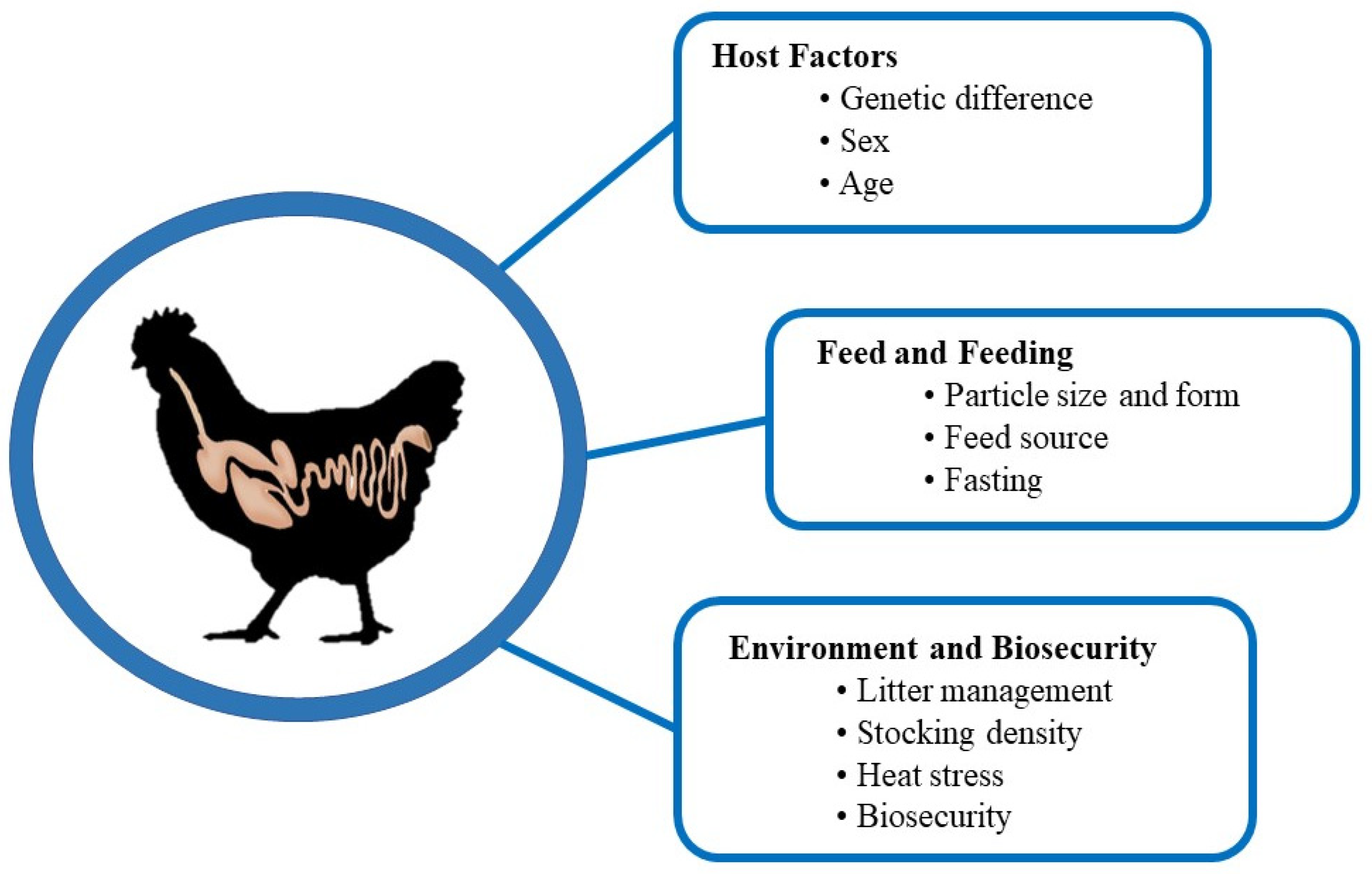
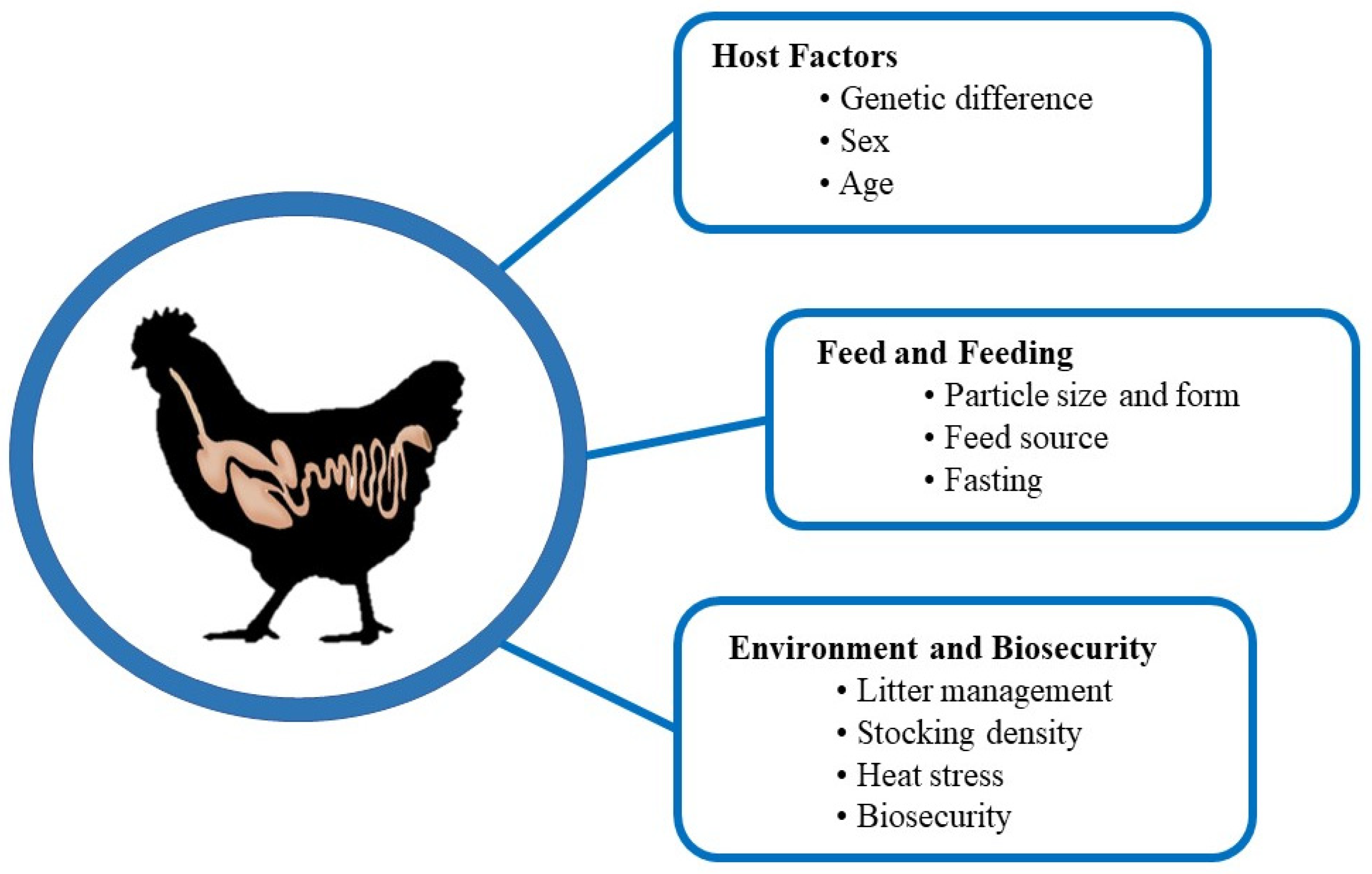
7. Factors Affecting Intestinal Health
7.1. Host Factors
7.1.1. Genetic Difference
7.1.2. Sex
7.1.3. Age
7.2. Feed and Nutrition
7.2.1. Particle Size and Form of Feed
7.2.2. Feeding Stuff
7.2.3. Impact of Fasting
7.3. Environmental Management
7.3.1. Litter Management
7.3.2. Stocking Density
7.3.3. Heat Stress
7.3.4. Biosecurity Measures
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ehrlich, P.R.; Harte, J. Opinion: To feed the world in 2050 will require a global revolution. Proc. Natl. Acad. Sci. USA 2015, 112, 14743–14744. [Google Scholar] [CrossRef] [Green Version]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef]
- Mehdi, Y.; Létourneau-Montminy, M.P.; Gaucher, M.L.; Chorfi, Y.; Suresh, G.; Rouissi, T.; Brar, S.K.; Côté, C.; Ramirez, A.A.; Godbout, S. Use of antibiotics in broiler production: Global impacts and alternatives. Anim. Nutr. 2018, 2, 170–178. [Google Scholar] [CrossRef]
- Lillehoj, H.; Liu, Y.; Calsamiglia, S.; Fernandez-Miyakawa, M.E.; Chi, F.; Cravens, R.L.; Oh, S.; Gay, C.G. Phytochemicals as antibiotic alternatives to promote growth and enhance host health. Vet. Res. 2018, 49, 76. [Google Scholar] [CrossRef] [Green Version]
- Thoo, L.; Noti, M.; Krebs, P. Keep calm: The intestinal barrier at the interface of peace and war. Cell Death Dis. 2019, 10, 849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staudacher, H.M.; Loughman, A. Gut health: Definitions and determinants. Lancet Gastroenterol. Hepatol. 2021, 6, 269. [Google Scholar] [CrossRef]
- Bischoff, S.C. Gut health: A new objective in medicine? BMC Med. 2011, 9, 24. [Google Scholar] [CrossRef]
- Kogut, M.H.; Arsenault, R.J. Editorial: Gut health: The New Paradigm in Food Animal Production. Front. Vet. Sci. 2016, 3, 71. [Google Scholar] [CrossRef] [Green Version]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steiner, R.E.; Kluenter, A.-M.; Verlhac, V. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal nutrition. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Jha, R.; Fouhse, J.M.; Tivari, U.P.; Li, L.; Willing, B.P. Dietary fiber and intestinal health of monogastric animals. Front. Vet. Sci. 2019, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Conway, P.L. Function and regulation of the gastrointestinal microbiota of the pig. In Proceedings of the VIth International Symposium on Digestive Physiology in Pigs; Souffrant, W., Hagemeister, H., Eds.; EAAP Publication: Ban Doberan, Germany, 1994; pp. 231–240. [Google Scholar]
- Calefi, A.S.; da Silva Fonseca, J.G.; Cohn, D.W.H.; Honda, B.T.B.; Costola-de-Souza, C.; Tsugiyama, L.E.; Quinteiro-Filho, W.M.; Piantino Ferreira, A.J.; Palermo-Neto, J. The gut-brain axis interactions during heat stress and avian necrotic enteritis. Poult. Sci. 2016, 95, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Susi, F.R. Keratinization in the mucosa of the ventral surface of the chicken tongue. J. Anat. 1969, 105, 477. [Google Scholar]
- Scanes, C.G.; Pierzchala-Koziec, K. Biology of the gastrointestinal tract in poultry. Avian Biol. Res. 2014, 7, 193–222. [Google Scholar] [CrossRef]
- Alshamy, Z.; Richardson, K.C.; Hünigen, H.; Hafez, H.M.; Plendl, J.; Al Masri, S. Comparison of the gastrointestinal tract of a dual-purpose to a broiler chicken line: A qualitative and quantitative macroscopic and microscopic study. PLoS ONE 2018, 13, e0204921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabata, H.; Yasugi, S. Tissue interaction regulates expression of a spasmolytic polypeptide gene in chicken stomach epithelium. Dev. Growth Differ. 1998, 40, 519–526. [Google Scholar] [CrossRef] [Green Version]
- van Dijk, A.; Veldhuizen, E.J.; Kalkhove, S.I.; Tjeerdsma-van Bokhoven, J.L.; Romijn, R.A.; Haagsman, H.P. The beta-defensin gallinacin-6 is expressed in the chicken digestive tract and has antimicrobial activity against foodborne pathogens. Antimicrob. Agents Chemother. 2007, 51, 912–922. [Google Scholar] [CrossRef] [Green Version]
- Svihus, B. The gizzard: Function, influence of diet structure and effects on nutrient availability. World’s Poult. Sci. J. 2011, 67, 207–224. [Google Scholar] [CrossRef]
- Aguzey, H.A.; Gao, Z.; Haohao, W.; Guilan, C. Influence of Feed Form and Particle Size on Gizzard, Intestinal Morphology and Microbiota Composition of Broiler Chicken. Poult. Fish. Wildl. Sci. 2018, 6, 2. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Lillehoj, E.P. Avian coccidiosis. A review of acquired intestinal immunity and vaccination strategies. Avian Dis. 2000, 44, 408–425. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V. Do symbiotic bacteria subvert host immunity? Nat. Rev. Microbiol. 2009, 7, 367–374. [Google Scholar] [CrossRef]
- Coleman, O.I.; Haller, D. Bacterial signaling at the intestinal epithelial interface in inflammation and cancer. Front. Immunol. 2018, 8, 1927. [Google Scholar] [CrossRef]
- Turk, D.E. The anatomy of the avian digestive tract as related to feed utilization. Poult. Sci. 1982, 61, 1225–1244. [Google Scholar] [CrossRef]
- Hong, Y.H.; Song, W.; Lee, S.H.; Lillehoj, H.S. Differential gene expression profiles of β-defensins in the crop, intestine, and spleen using a necrotic enteritis model in 2 commercial broiler chicken lines. Poult. Sci. 2012, 91, 1081–1088. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Dadak, A.; Hess, M.; Böhm, J. Single and combined effects of deoxynivalenol mycotoxin and a microbial feed additive on lymphocyte DNA damage and oxidative stress in broiler chickens. PLoS ONE 2014, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svihus, B.; Choct, M.; Classen, H.L. Function and nutritional roles of the avian caeca: A review. World’s Poult. Sci. J. 2013, 69, 249–264. [Google Scholar] [CrossRef]
- Dunkley, K.D.; Callaway, T.R.; Chalova, V.I.; McReynolds, J.L.; Hume, M.E.; Dunkley, C.S.; Kubena, L.F.; Nisbet, D.J.; Ricke, S.C. Foodborne Salmonella ecology in the avian gastrointestinal tract. Anaerobe 2009, 15, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Majeed, M.F.; Al-Asadi, F.S.; Nassir, A.A.; Rahi, E.H. The morphological and histological study of the caecum in broiler chicken. Basra J. Vet. Res. 2009, 8, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Grenham, S.; Clarke, G.; Cryan, J.F.; Dinan, T.G. Brain–gut–microbe communication in health and disease. Front. Physiol. 2011, 2, 94. [Google Scholar] [CrossRef] [Green Version]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203. [Google Scholar] [PubMed]
- Villageliũ, D.N.; Lyte, M. Microbial endocrinology: Why the intersection of microbiology and neurobiology matters to poultry health. Poult. Sci. 2017, 96, 2501–2508. [Google Scholar] [CrossRef]
- Kabiersch, A.; del Rey, A.; Honegger, C.G.; Besedovsky, H.O. Interleukin-1 induces changes in norepinephrine metabolism in the rat brain. Brain Behav. Immun. 1988, 2, 267–274. [Google Scholar] [CrossRef]
- Calefi, A.S.; Honda, B.T.B.; Costola-de-Souza, C.; de Siqueira, A.; Namazu, L.B.; Quinteiro-Filho, W.M.; Fonseca, J.G.D.S.; Aloia, T.P.A.; Piantino-Ferreira, A.J.; Palermo-Neto, J. Effects of long-term heat stress in an experimental model of avian necrotic enteritis. Poult. Sci. 2014, 93, 1344–1353. [Google Scholar] [CrossRef]
- Calefi, A.S.; Fonseca, J.G.D.S.; Nunes, C.A.D.Q.; Lima, A.P.N.; Quinteiro-Filho, W.M.; Flório, J.C.; Zager, A.; Ferreira, A.J.P.; Palermo-Neto, J. Heat Stress Modulates Brain Monoamines and Their Metabolites Production in Broiler Chickens Co-Infected with Clostridium perfringens Type A and Eimeria spp. Vet. Sci. 2019, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Borsoi, A.; Quinteiro-Filho, W.M.; Calefi, A.S.; Piantino Ferreira, A.J.; Astolfi-Ferreira, C.S.; Florio, J.C.; Palermo-Neto, J. Effects of cold stress and Salmonella Heidelberg infection on bacterial load and immunity of chickens. Avian Pathol. 2015, 44, 490–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geyra, A.; Uni, Z.; Sklan, D. Enterocyte dynamics and mucosal development in the posthatch chick. Poult. Sci. 2001, 80, 776–782. [Google Scholar] [CrossRef]
- Friedman, A.; Bar-Shira, E.; Sklan, D. Ontogeny of gut associated immune competence in the chick. Worlds. Poult. Sci. J. 2003, 59, 209–219. [Google Scholar] [CrossRef]
- Bar-Shira, E.; Friedman, A. Development and adaptations of innate immunity in the gastrointestinal tract of the newly hatched chick. Dev. Comp. Immunol. 2006, 30, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.S.; Trout, J.M. Avian gut-associated lymphoid tissues and intestinal immune responses to Eimeria parasites. Clin. Microbiol. Rev. 1996, 9, 349–360. [Google Scholar] [CrossRef]
- Lillehoj, H.S. Analysis of Eimeria acervulina-induced changes in the intestinal T lymphocyte subpopulations in two chicken strains showing different levels of susceptibility to coccidiosis. Res. Vet. Sci. 1994, 56, 1–7. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Min, W.; Dalloul, R.A. Recent progress on the cytokine regulation of intestinal immune responses to Eimeria. Poult. Sci. 2004, 83, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Dunon, D.; Allioli, N.; Vainio, O.; Ody, C.; Imhof, B.A. Quantification of T-cell progenitors during ontogeny: Thymus colonization depends on blood delivery of progenitors. Blood. 1999, 93, 2234–2243. [Google Scholar] [CrossRef]
- Coltey, M.; Bucy, R.P.; Chen, C.H.; Cihak, J.; Losch, U.; Char, D.; Le Douarin, N.M.; Cooper, M.D. Analysis of the first two waves of thymus homing stem cells and their T cell progeny in chick–quail chimeras. J. Exp. Med. 1989, 170, 543–557. [Google Scholar] [CrossRef]
- Imhof, B.A.; Dunon, D.; Courtois, D.; Luhtala, M.; Vainio, O. Intestinal CD8 alpha alpha and CD8 alpha beta intraepithelial lymphocytes are thymus derived and exhibit subtle differences in TCR beta repertoires. J. Immunol. 2000, 165, 6716–6722. [Google Scholar] [CrossRef] [Green Version]
- Weill, J.C.; Weller, S.; Reynaud, C.A. A bird’s eye view on human B cells. Semin. Immunol. 2004, 16, 277–281. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, M.J.H. Antibodies, immunoglobulin genes and the bursa of Fabricius in chicken B cell development. Dev. Comp. Immunol. 2006, 30, 101–118. [Google Scholar] [CrossRef]
- Ratcliffe, M.J.; Jacobsen, K.A. Rearrangement of immunoglobulin genes in chicken B cell development. Semin. Immunol. 1994, 6, 175–184. [Google Scholar] [CrossRef]
- Reynaud, C.A.; Imhof, B.A.; Anquez, V.; Weill, J.C. Emergence of committed B lymphoid progenitors in the developing chicken embryo. EMBO J. 1992, 11, 4349–4358. [Google Scholar] [CrossRef] [PubMed]
- Mansikka, A.; Sandberg, M.; Veromaa, T.; Vainio, O.; Granfors, K.; Toivanen, P. B cell maturation in the chicken Harderian gland. J. Immunol. 1989, 142, 1826–1833. [Google Scholar] [PubMed]
- Gadde, U.; Rathinam, T.; Lillehoj, H.S. Passive immunization with hyperimmune egg-yolk IgY as prophylaxis and therapy for poultry diseases—A review. Anim. Health Res. Rev. 2015, 16, 1–14. [Google Scholar] [CrossRef]
- Sahin, O.; Luo, N.; Huang, S.; Zhang, Q. Effect of Campylobacter-specific maternal antibodies on Campylobacter jejuni colonization in young chickens. Appl. Environ. Microbiol. 2003, 69, 5372–5379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.L.; Rose, M.E.; Wakelin, D. The role of natural killer cells in resistance to coccidiosis: Investigations in a murine model. Clin. Exp. Immunol. 1994, 97, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Bitay, Z.; Szell, Z.; Varga, I. Assessment of maternal immunity to Cryptosporidium baileyi in chickens. Vet. Parasitol. 1998, 79, 203–212. [Google Scholar] [CrossRef]
- Broom, L.J.; Kogut, M.H. Inflammation: Friend and foe for animal production? Poult. Sci. 2018, 94, 510–514. [Google Scholar] [CrossRef]
- Kannaki, T.R.; Reddy, M.R.; Shanmugam, M.; Verma, P.C.; Sharma, R.P. Chicken toll-like receptors and their role in immunity. Worlds Poult. Sci. J. 2010, 66, 727–738. [Google Scholar]
- Zhang, J.; Li, C.; Tang, X.; Lu, Q.; Sa, R.; Zhang, H. Proteome changes in the small intestinal mucosa of broilers (Gallus gallus) induced by high concentrations of atmospheric ammonia. Proteome Sci. 2015, 13, 9. [Google Scholar] [CrossRef] [Green Version]
- Deplancke, B.; Gaskins, H.R. Microbial modulation ofinnate defence: Goblet cells and the intestinal mucus layer. Am. J. Clin. Nutr. 2001, 73, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Wlodarska, M.; Willing, B.; Keeney, K.M.; Menendez, A.; Bergstrom, K.S.; Gill, N.; Russell, S.L.; Vallance, B.A.; Finlay, B.B. Antibiotic treatment alters the colonic mucus layer and pre-disposes the host to exacerbated Citrobacter rodentium-induced colitis. Infect. Immun. 2011, 79, 1536–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, K.; Deng, Z.; Hou, Y.; Zhang, G. Regulation of the intestinal barrier function by host defense peptides. Front. Vet. Sci. 2015, 2, 57. [Google Scholar] [CrossRef]
- Hong, Y.H.; Lillehoj, H.S.; Siragusa, G.R.; Bannerman, D.D.; Lillehoj, E.P. Antimicrobial activity of chicken NK-lysin against Eimeria sporozoites. Avian Dis. 2008, 52, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Evaluation of the Immunomodulatory Activity of the Chicken NK-Lysin-Derived Peptide cNK-2. Sci. Rep. 2017, 7, 45099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickramasuriya, S.S.; Park, I.; Lee, Y.; Kim, W.H.; Przybyszewski, C.; Gay, C.G.; van Oosterwijk, J.G.; Lillehoj, H.S. Oral Delivery of Bacillus subtilis Expressing Chicken NK-2 Peptide Protects Against Eimeria acervulina Infection in Broiler Chickens. Front. Vet. Sci. 2021, 8, 684818. [Google Scholar] [CrossRef]
- Wang, G. Human antimicrobial peptides and proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef] [Green Version]
- Mantis, N.J.; Rol, N.; Corthesy, B. Secretory IgA’s com-plex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. 2011, 4, 603–611. [Google Scholar] [CrossRef]
- Honda, K.; Takeda, K. Regulatory mechanisms of immuneresponses to intestinal bacteria. Mucosal Immunol. 2009, 2, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, A.; Perez, E.; Amit-Romach, E.; Sklan, D.; Uni, Z. Mucin dynamics and microbial populations in chicken small intestine are changed by dietary probiotic and antibiotic growth promoter supplementation. J. Nutr. 2005, 135, 187–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisbin, J.T.; Gong, J.; Sharif, S. Interactions between commensal bacteria and the gut-associated immune system of the chicken. Anim. Health Res. Rev. 2008, 9, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Rehman, H.U.; Vahjen, W.; Awad, W.A.; Zentek, J. Indigenous bacteria and bacterial metabolic products in the gastrointestinal tract of broiler chickens. Arch. Anim. Nutrit. 2007, 61, 319–335. [Google Scholar] [CrossRef]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken Gut Microbiota: Importance and Detection Technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef]
- Waite, D.W.; Taylor, M. Exploring the avian gut microbiota: Current trends and future directions. Front. Microbiol. 2015, 6, 673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, J.J.; Klima, C.L.; Sylte, M.J.; Looft, T. The Microbial pecking order: Utilization of intestinal microbiota for poultry health. Microorganisms 2019, 7, 376. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. J. Anim. Sci. Biotechnol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Shuling, N.; Kunyuan, T.; Qiuyang, Z.; Hewen, D.; ChenCheng, G.; Tianhe, Y.; Liancheng, L.; Xin, F. Characteristics of the intestinal flora of specific pathogen free chickens with age. Microb. Pathog. 2019, 132, 325–334. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Yegani, M.; Korver, D.R. Factors Affecting Intestinal Health in Poultry. Poult. Sci. 2008, 87, 2052–2063. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Hase, K. Gut microbiota-generated metabolites in animal health and disease. Nat. Chem. Biol. 2014, 10, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Wilson, I.D. Opinion: Understanding ‘global’ systems biology: Metabonomics and the continuum of metabolism. Nat. Rev. Drug Discov. 2003, 2, 668–676. [Google Scholar] [CrossRef]
- Cully, M. Microbiome therapeutics go small molecule. Nat. Rev. Drug. Discov. 2019, 18, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S.; et al. ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef]
- Kogut, M.H. The effect of microbiome modulation on the intestinal health of poultry. Anim. Feed Sci. Technol. 2019, 250, 32–40. [Google Scholar] [CrossRef]
- Tan, J.; Applegate, T.J.; Liu, S.; Guo, Y.; Eicher, S.D. Supplemental dietary L-Arginine attenuates intestinal mucosal disruption during a coccidial vaccine challenge in broiler chickens. Br. J. Nutr. 2014, 112, 1098–1109. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.A.; Cannons, J.L.; Grainger, J.R.; Dos Santos, L.M.; Hand, T.W.; Naik, S.; Wohlfert, E.A.; Chou, D.B.; Oldenhove, G.; Robinson, M.; et al. Essential role for retinoic acid in the promotion of CD4(+) T cell effector responses via retinoic acid receptor alpha. Immunity 2011, 34, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Park, I.; Oh, S.; Lillehoj, E.P.; Lillehoj, H.S. Dietary Supplementation with Magnolia bark extract alters chicken intestinal metabolite levels. Front. Vet. Sci. 2020, 7, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, I.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.G.; Lillehoj, E.P.; Lillehoj, H.S. Dietary supplementation with Bacillus subtilis direct-fed microbials alters chicken intestinal metabolite levels. Front. Vet. Sci. 2020, 7, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieterich, W.; Schink, M.; Zopf, Y. Microbiota in the Gastrointestinal Tract. Med. Sci. 2018, 6, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, F.; Backhed, F. The gut microbiota—masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef]
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef] [PubMed]
- Dalloul, R.A.; Lillehoj, H.S. Poultry coccidiosis: Recent advancements in control measures and vaccine development. Expert Rev. Vaccines. 2006, 5, 143–163. [Google Scholar] [CrossRef]
- De Gussem, M. Coccidiosis in poultry: Review on diagnosis, control, prevention and interaction with overall gut health. In Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007; pp. 253–261. [Google Scholar]
- Nematollahi, A.; Moghaddam, G.; Pourabad, R.F. Prevalence of Eimeria species among broiler chicks in Tabriz (Northwest of Iran). Mun. Ent. Zool. 2009, 4, 53–58. [Google Scholar]
- Sharman, P.A.; Smith, N.C.; Wallach, M.G.; Katrib, M. Chasing the golden egg: Vaccination against poultry coccidiosis. Parasite Immunol. 2010, 32, 590–598. [Google Scholar] [CrossRef]
- Kim, W.H.; Lillehoj, H.S.; Min, W. Indole treatment alleviates intestinal tissue damage induced by chicken coccidiosis through activation of the aryl hydrocarbon receptor. Front. Immunol. 2019, 10, 560. [Google Scholar] [CrossRef]
- Mitra, T.; Kidane, F.A.; Hess, M.; Liebhart, D. Unravelling the immunity of poultry against the extracellular protozoan parasite Histomonas meleagridis is a cornerstone for vaccine development: A review. Front. Immunol. 2018, 9, 2518. [Google Scholar] [CrossRef]
- Regmi, P.R.; Shaw, A.L.; Hungerford, L.L.; Messenheimer, J.R.; Zhou, T.; Pillai, P.; Omer, A.; Gilbert, J.M. Regulatory Considerations for the Approval of Drugs Against Histomoniasis (Blackhead Disease) in Turkeys, Chickens, and Game Birds in the United States. Avian Dis. 2016, 60, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.L.; Beckstead, R.B.; Hayes, J.R.; Rissi, D.R. Pathologic and molecular characterization of histomoniasis in peafowl (Pavo cristatus). J. Vet. Diagn. 2017, 29, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sentíes-Cué, G.; Chin, R.P.; Shivaprasad, H.L. Systemic histomoniasis associated with high mortality and unusual lesions in the bursa of Fabricius, kidneys, and lungs in commercial turkeys. Avian Dis. 2009, 53, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Sigmon, C.S.; Malheiros, R.D.; Anderson, K.E.; Payne, J.A.; Beckstead, R.B. Blackhead Disease: Recovery of Layer Flock after Disease Challenge. J. Appl. Poult. Res. 2019, 28, 755–760. [Google Scholar] [CrossRef]
- Liebhart, D.; Ganas, P.; Sulejmanovic, T.; Hess, M. Histomonosis in poultry: Previous and current strategies for prevention and therapy. Avian Pathol. 2017, 46, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wade, B.; Keyburn, A. The true cost of necrotic enteritis. Poult. World. 2015, 31, 16–17. [Google Scholar]
- Keyburn, A.L.; Bannam, T.L.; Moore, R.J.; Rood, J.I. NetB, a Pore-forming toxin from necrotic enteritis strains of Clostridium perfringens. Toxins 2010, 2, 1913–1927. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Lillehoj, H.S.; Park, M.S.; Jang, S.I.; Ritter, G.D.; Hong, Y.H.; Jeong, W.; Jeoung, H.Y.; An, D.J.; Lillehoj, E.P. Clostridium perfringens α-toxin and NetB toxin antibodies and their possible role in protection against necrotic enteritis and gangrenous dermatitis in broiler chickens. Avian Dis. 2012, 56, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, W.H.; jin Lee, S.; Lillehoj, H.S. Detection of chicken interleukin-10 production in intestinal epithelial cells and necrotic enteritis induced by Clostridium perfringens using capture ELISA. Vet. Immunol. Immunopathol. 2018, 204, 52–58. [Google Scholar] [CrossRef]
- Kim, D.K.; Lillehoj, H.S.; Jang, S.I.; Lee, S.H.; Hong, Y.H.; Cheng, H.H. Transcriptional profiles of host-pathogen responses to necrotic enteritis and differential regulation of immune genes in two inbreed chicken lines showing disparate disease susceptibility. PLoS ONE 2014, 9, 114960. [Google Scholar] [CrossRef] [PubMed]
- Zaytsoff, S.J.M.; Lyons, S.M.; Garner, A.M.; Uwiera, R.R.E.; Zandberg, W.F.; Abbott, D.W.; Inglis, G.D. Host responses to Clostridium perfringens challenge in a chicken model of chronic stress. Gut Pathog. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Gad, W.; Hauck, R.; Krüger, M.; Hafez, H.M. Prevalence of clostridium perfringens in commercial Turkey and layer flocks. Arch. Geflugelkd. 2011, 75, 74–79. [Google Scholar]
- El-helw, H.; Serum, V.; El-sergany, E.F.; Serum, V. Role of Clostridium perfringens type A as a causative agent of necrotic enteritis in Turkey. Vet. Med. J. Giza. 2014, 60, C1–C22. [Google Scholar]
- Lee, Y.S.; Lee, S.H.; Gadde, U.D.; Oh, S.T.; Lee, S.J.; Lillehoj, H.S. Allium hookeri supplementation improves intestinal immune response against necrotic enteritis in young broiler chickens. Poult. Sci. 2018, 97, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Hermans, D.; Martel, A.; Garmyn, A.; Verlinden, M.; Heyndrickx, M.; Gantois, I.; Haesebrouck, F.; Pasmans, F. Application of medium-chain fatty acids in drinking water increases Campylobacter jejuni colonization threshold in broiler chicks. Poult. Sci. 2012, 91, 1733–1738. [Google Scholar] [CrossRef]
- Li, J.; Miao, Z.; Tian, W.; Yang, Y.; Wang, J.; Yang, Y. Effects of different rearing systems on growth, small intestinal morphology and selected indices of fermentation status in broilers. Anim. Sci. J. 2017, 88, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Y.; Lee, Y.; Lu, H.; Chou, C.H.; Wang, C. Analysis of contributory gut microbiota and lauric acid against necrotic enteritis in Clostridium perfringens and Eimeria side-by-side challenge model. bioRxiv 2018, 434449. [Google Scholar] [CrossRef] [Green Version]
- Pizarro, M.; Höfle, U.; Rodríguez-Bertos, A.; González-Huecas, M.; Castaño, M. Ulcerative Enteritis (Quail Disease) in Lories. Avian Dis. 2005, 49, 606–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, K.K.; Songer, J.G.; Uzal, F.A. Diagnosing clostridial enteric disease in poultry. J. Vet. Diagnostic Investig. 2013, 25, 314–327. [Google Scholar] [CrossRef] [Green Version]
- Wages, D.P. Ulcerative enteritis. In Diseases of Poultry, 12th ed.; Blackwell: Ames, IA, USA, 2008; pp. 867–871. [Google Scholar]
- Bano, L.; Drigo, I.; Macklin, K.S.; Martin, S.W.; Miller, R.S.; Norton, R.A.; Oyarzabal, O.A.; Bilgili, S.F. Development of a polymerase chain reaction assay for specific identification of Clostridium colinum. Avian Pathol. 2008, 37, 179–181. [Google Scholar] [CrossRef] [Green Version]
- Radi, Z.A. An epizootic of combined Clostridium perfringens, Eimeria spp. and Capillaria spp. enteritis and Histomonas spp. hepatitis with Escherichia coli septicemia in bobwhite quails (Colinus virginianus). Int. J. Poult. Sci. 2004, 3, 438–441. [Google Scholar]
- Porter, R.E. Bacterial Enteritides of Poultry. Poult. Sci. 1998, 77, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Barrow, P.A.; Freitas Neto, O.C. Pullorum disease and fowl typhoid-new thoughts on old diseases: A review. Avian Pathol. 2011, 40, 1–13. [Google Scholar] [CrossRef]
- Desin, T.S.; Köster, W.; Potter, A.A. Salmonella vaccines in poultry: Past, present and future. Expert Rev. Vaccines. 2013, 12, 87–96. [Google Scholar] [CrossRef]
- Shivaprasad, H.L. Fowl typhoid and pullorum disease. OIE Rev. Sci. Technol. 2000, 19, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Bande, F.; Arshad, S.S.; Omar, A.R.; Hair-Bejo, M.; Mahmuda, A.; Nair, V. Global distributions and strain diversity of avian infectious bronchitis virus: A review. Anim. Health Res. Rev. 2017, 18, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Britton, P.; Armesto, M.; Cavanagh, D.; Keep, S. Modification of the avian coronavirus infectious bronchitis virus for vaccine development. Bioeng. Bugs. 2012, 3, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, G.D.; Jones, R.C. Infectious bronchitis virus: Immunopathogenesis of infection in the chicken. Avian Pathol. 1997, 26, 677–706. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, X.; Wang, Y.; Li, C.; Han, Z.; Shao, Y.; Li, H.; Kong, X. Molecular characterization and pathogenicity of infectious bronchitis coronaviruses: Complicated evolution and epidemiology in China caused by cocirculation of multiple types of infectious bronchitis coronaviruses. Intervirology 2009, 52, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Davis, P.J.; Mockett, A.P.A. Amino acids within hypervariable region 1 of avian coronavirus IBV (Massachusetts serotype) spike glycoprotein are associated with neutralization epitopes. Virus Res. 1988, 11, 141–150. [Google Scholar] [CrossRef]
- Schat, K.A.; Skinner, M.A. Avian Immunosuppressive Diseases and Immuneevasion. Avian Immunol. 2008, 299–322. [Google Scholar] [CrossRef]
- Montgomery, R.D.; Villegas, P.; Dawe, D.L.; Brown, J. Effect of Avian Reoviruses on Lymphoid Organ Weights and Antibody Response in Chickens. Avian Dis. 1985, 29, 552. [Google Scholar] [CrossRef]
- Guy, J.S. Virus Infections of the Gastrointestinal Tract of Poultry. Poult. Sci. 1998, 77, 1166–1175. [Google Scholar] [CrossRef]
- Shivaprasad, H.L.; Franca, M.; Woolcock, P.R.; Nordhausen, R.; Day, J.M.; Pantin-Jackwood, M. Myocarditis Associated with Reovirus in Turkey Poults. Avian Dis. 2009, 53, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.C. Avian reovirus infections. OIE Rev. Sci. Technol. 2000, 19, 614–625. [Google Scholar] [CrossRef]
- van der Heide, L. The History of Avian Reovirus. Avian Dis. 2000, 44, 638. [Google Scholar] [CrossRef]
- Spackman, E.; Pantin-Jackwood, M.; Michael Day, J.; Sellers, H. The pathogenesis of turkey origin reoviruses in turkeys and chickens. Avian Pathol. 2005, 34, 291–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, Y.; Kemp, M.C. A comparative study of avian Reovirus pathogenicity: Virus spread and replication and induction of lesions. Avian Dis. 1995, 39, 554. [Google Scholar] [CrossRef]
- Wigand, R.; Bartlia, A.; Dreizin, R.S.; Esche, H.; Ginsberg, H.S.; Green, M.; Hierholzer, J.C.; Kalter, S.S.; McFerran, J.B.; Pettersson, U.; et al. Adenoviridae: Second report. Intervirology 1982, 18, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Kouwenhoven, B.; Davelaar, F.G.; Van Walsum, J. Infectious proventriculitis causing runting in broilers. Avian Pathol. 1978, 7, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, M.A. Adenovirus inclusion body ventriculitis in chickens and captive bobwhite quail (Colinus virginianus). Avian Dis. 1993, 37, 568. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, M.A.; Hill, D.L.; Dekich, M.A.; Putnam, M.R. Multisystemic Adenovirus infection in broiler chicks with hypoglycemia and spiking mortality. Avian Dis. 1993, 37, 625. [Google Scholar] [CrossRef]
- Barnes, H.J.; Guy, J.S. Poult enteritis—Mortality syndrome. In Diseases of Poultry, 11th ed.; Saif, Y.M., Barnes, H.J., Glisson, J.R., Fadly, A.M., McDougald, L.R., Eds.; Iowa State University Press: Ames, IA, USA, 2003; pp. 320–326. [Google Scholar]
- Domermuth, C.H.; Harris, J.R.; Gross, W.B.; DuBose, R.T. A naturally occurring infection of chickens with a hemorrhagic enteritis/marble spleen disease type of virus. Avian Dis. 1979, 23, 479. [Google Scholar] [CrossRef]
- Pierson, F.W.; Fitzgerald, S.D. Hemorrhagic enteritis and related infections. In Diseases of Poultry, 13th ed.; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; Iowa State University Press: Ames, IA, USA, 2013; pp. 237–247. [Google Scholar]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and environmental factors affecting the intestinal microbiota in chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.K.; Lillehoj, H.S.; Jang, S.I.; Lee, S.H.; Hong, Y.H.; Lamont, S.J. Genetically disparate fayoumi chicken lines show different response to avian necrotic enteritis. J. Poult. Sci. 2015, 52, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Swaggerty, C.L.; Kogut, M.H.; Chiang, H.I.; Wang, Y.; Genovese, K.J.; He, H.; Zhou, H. Gene expression profiling of the local cecal response of genetic chicken lines that differ in their susceptibility to Campylobacter jejuni colonization. PLoS ONE 2010, 5, e11827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.T.; Lillehoj, H.S. The role of host genetic factors and host immunity in necrotic enteritis. Avian Pathol. 2016, 45, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Willer, T.; Pielsticker, C.; Gerzova, L.; Rychlik, I.; Rautenschlein, S. Differences in host breed and diet influence colonization by Campylobacter jejuni and induction of local immune responses in chickens. Gut Pathog. 2016, 8, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Lillehoj, E.P.; Hong, Y.H.; An, D.J.; Jeoung, H.Y.; Chun, J.E. Relative disease susceptibility and clostridial toxin antibody responses in three commercial broiler lines coinfected with Clostridial perfringens and Eimeria maxima using an experimental model of necrotic enteritis. Avian Dis. 2013, 57, 684–687. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lillehoj, H.S.; Hong, Y.H.; Kim, G.B.; Lee, S.H.; Lillehoj, E.P.; Bravo, D.M. Dietary Capsicum and Curcuma longa oleoresins increase intestinal microbiome and necrotic enteritis in three commercial broiler breeds. Res. Vet. Sci. 2015, 102, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Adeleye, O.O.; Otakoya, I.O.; Fafiolu, A.O.; Alabi, J.O.; Egbeyale, L.T.; Idowu, O.M.O. Serum chemistry and gut morphology of two broiler chickens to varying interval of post hatch feeding. Vet. Anim. Sci. 2017, 5, 20–25. [Google Scholar] [CrossRef]
- Smith, M.W.; Mitchell, M.A.; Peacock, M.A. Effects of genetic selection on growth rate and intestinal structure in the domestic fowl (Gallus domesticus). Comp. Biochem. Physiol. 1990, 97A, 57–63. [Google Scholar] [CrossRef]
- Uni, Z.; Noy, Y.; Sklan, D. Development of the small intestine in heavy and light strain chicks before and after hatching. Brit. Poult. Sci. 1995, 36, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Uni, Z.; Noy, Y.; Sklan, D. Posthatch changes in morphology and function of the small intestine in heavy- and light-strain chicks. Poult. Sci. 1995, 74, 1622–1629. [Google Scholar] [CrossRef]
- Zulkifli, I.; Iman Rahayu, H.S.; Alimon, A.R.; Vidyadaran, M.K.; Babjee, S.A. Gut microflora and intestinal morphology of commercial broiler chickens and red jungle fowl fed diets containing palm kernel meal. Arch. Geflügelkd. 2009, 73, 49–55. [Google Scholar]
- Sun, J.; Wang, Y.; Li, N.; Zhong, H.; Xu, H.; Zhu, Q.; Liu, Y. Comparative analysis of the gut microbial composition and meat flavor of two chicken breeds in different rearing patterns. Biomed. Res. Int. 2018, 4343196. [Google Scholar] [CrossRef] [Green Version]
- Theerawatanasirikul, S.; Koomkrong, N.; Kayan, A.; Boonkaewwan, C. Intestinal barrier and mucosal immunity in broilers, Thai Betong, and native Thai Praduhangdum chickens. Turk. J. Vet. Anim. Sci. 2017, 41, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Lumpkins, B.S.; Batal, A.B.; Lee, M. The effect of gender on the bacterial community in the gastrointestinal tract of broilers. Poult. Sci. 2008, 87, 964–967. [Google Scholar] [CrossRef]
- Torok, V.A.; Dyson, C.; Mckay, A.; Ophel-Keller, K. Quantitative molecular assays for evaluating changes in broiler gut microbiota linked with diet and performance. Anim. Prod. Sci. 2013, 53, 1260–1268. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.C.; Kil, D.Y.; Sul, W.J. Cecal microbiome divergence of broiler chickens by sex and body weight. J. Microbiol. 2017, 55, 939–945. [Google Scholar] [CrossRef]
- Miles, R.D.; Butcher, G.D.; Henry, P.R.; Littell, R.C. Effect of antibiotic growth promoters on broiler performance, intestinal growth parameters, and quantitative morphology. Poult. Sci. 2006, 85, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Rattanawut, J. Effects of dietary bamboo charcoal powder including bamboo vinegar liquid supplementation on growth performance, fecal microflora population and intestinal morphology in Betong chickens. J. Poult. Sci. 2014, 51, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Thanabalan, A.; Leung, H.; Kakhki, R.A.M.; Patterson, R.; Kiarie, E.G. The effects of feeding yeast bioactives to broiler breeders and/or their offspring on growth performance, gut development, and immune function in broiler chickens challenged with Eimeria. Poult. Sci. 2019, 98, 6411–6421. [Google Scholar] [CrossRef]
- Boulton, K.; Nolan, M.J.; Wu, Z.; Psifidi, A.; Riggio, V.; Harman, K.; Bishop, S.C.; Kaiser, P.; Abrahamsen, M.S.; Hawken, R.; et al. Phenotypic and genetic variation in the response of chickens to Eimeria tenella induced coccidiosis. Genet. Sel. Evol. 2018, 50, 63. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Mora, J.R.; Mora, J.R.; et al. Gut immune maturation depends on colonization with a host-specific microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef] [Green Version]
- Volf, J.; Polansky, O.; Sekelova, Z.; Velge, P.; Schouler, C.; Kasper, B.; Rychlik, I. Gene expression in the chicken cecum is dependent on microbiota composition. Vet. Res. 2017, 48, 85. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Dai, R.; Yang, L.; He, C.; Xu, K.; Liu, S.; Zhao, W.; Xiao, L.; Luo, L.; Zhang, Y.; et al. Inheritance and establishment of gut microbiota in chickens. Front. Microbiol. 2017, 8, 1967. [Google Scholar] [CrossRef]
- Roto, S.M.; Kwon, Y.M.; Ricke, S.C. Applications of in ovo technique for the optimal development of the gastrointestinal tract and the potential influence on the establishment of its microbiome in poultry. Front. Vet. Sci. 2016, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, E.E.; Stanley, D.; Hughes, R.J.; Moore, R.J. The time-course of broiler intestinal microbiota development after administration of cecal contents to incubating eggs. Peer J. 2017, 5, 3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurburg, S.D.; Brouwer, M.S.M.; Ceccarelli, D.; van der Goot, J.; Jansman, A.J.M.; Bossers, A. Patterns of community assembly in the developing chicken microbiome reveal rapid primary succession. Microbiol. Open 2019, 8, 821. [Google Scholar] [CrossRef] [PubMed]
- Zaefarian, F.; Abdollahi, M.R.; Ravindran, V. Particle size and feed form in broiler diets: Impact on gastrointestinal tract development and gut health. Worlds Poult. Sci. J. 2016, 72, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Kiarie, E.G.; Mills, A. Role of feed processing on gut health and function in pigs and poultry: Conundrum of optimal particle size and hydrothermal regimens. Front. Vet. Sci. 2019, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Engberg, R.M.; Hedemann, M.S.; Jensen, B.B. The influence of grinding and pelleting of feed on the microbial composition and activity in the digestive tract of broiler chickens. Brit. Poult. Sci. 2002, 43, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Yan, L.; Wang, Z.; An, S.; Wu, M.; Lv, Z. Effects of feed form and feed particle size on growth performance, carcass characteristics and digestive tract development of broilers. Anim. Nutr. 2015, 1, 252–256. [Google Scholar] [CrossRef]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.B. Upregulation of genes encoding digestive enzymes and nutrient transporters in the digestive system of broiler chickens by dietary supplementation of fiber and inclusion of coarse particle size corn. BMC Genom. 2018, 19, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amerah, A.M.; Ravindran, V.; Lentle, R.G.; Thomas, D.G. Infleunce of feed particle size and feed form on the performance, energy utilization, digestive tract development, and digesta parameters of broiler starters. Poult. Sci. 2007, 86, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.S.; Li, D.F.; Xing, J.J.; Ma, Y.X.; Li, Z.J.; Lv, S.Q. Effects of feed particle size and feed form on survival of Salmonella typhimurium in the alimentary tract and cecal S. typhimurium reduction in growing broilers. Poult. Sci. 2006, 85, 831–836. [Google Scholar] [CrossRef]
- Abadi, M.H.M.G.; Moravej, H.; Shivazad, M.; Karimi Torshizi, M.A.; Kim, W.K. Effects of feed form and particle size, and pellet binder on performance, digestive tract parameters, intestinal morphology, and cecal microflora populations in broilers. Poult. Sci. 2019, 98, 1432–1440. [Google Scholar] [CrossRef]
- Truong, H.H.; Moss, A.F.; Liu, S.Y.; Selle, P.H. Pre- and post-pellet whole grain inclusions enhance feed conversion efficiency, energy utilisation and gut integrity in broiler chickens offered wheat-based diets. Anim. Feed. Sci. Technol. 2017, 224, 115–123. [Google Scholar] [CrossRef]
- Lourenco, J.M.; Rothrock, M.J., Jr.; Sanad, Y.M.; Callaway, T.R. The effects of feeding a soybean-based or a soy-free diet on the gut microbiome of pasture-raised chickens throughout their lifecycle. Front. Sustain. Food. Syst. 2019, 3, 36. [Google Scholar] [CrossRef]
- Röhe, I.; Boroojeni, F.G.; Zentek, J. Effect of feeding soybean meal and differently processed peas on intestinal morphology and functional glucose transport in the small intestine of broilers. Poult. Sci. 2017, 96, 4075–4084. [Google Scholar] [CrossRef]
- Röhe, I.; Göbel, T.W.; Boroojeni, F.G.; Zentek, J. Effect of feeding soybean meal and differently processed peas on the gut mucosal immune system of broilers. Poult. Sci. 2017, 96, 2064–2073. [Google Scholar] [CrossRef]
- Lauridsen, C. Effects of dietary fatty acids on gut health and function of pigs pre- and post-weaning. J. Anim. Sci. 2020, 98, 86. [Google Scholar] [CrossRef]
- Kim, Y.B.; Kim, D.H.; Jeong, S.B.; Lee, J.W.; Kim, T.H.; Lee, H.G.; Lee, K.W. Black soldier fly larvae oil as an alternative fat source in broiler nutrition. Poult. Sci. 2020, 99, 3133–3143. [Google Scholar] [CrossRef]
- Knarreborg, A.; Simon, M.A.; Engberg, R.M.; Jensen, B.B.; Tannock, G.W. Effects of dietary fat source and subtherapeutic levels of antibiotic on the bacterial community in the ileum of broiler chickens at various ages. Appl. Environ. Microbiol. 2002, 68, 5918–5924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrer, C.; Pedragosa, E.; Torras-Llort, M.; Parcerisa, X.; Rafecas, M.; Ferrer, R.; Amat, C.; Moretó, M. Dietary lipids modify brush border membrane composition and nutrient transport in chicken small intestine. J. Nutr. 2003, 133, 1147–1153. [Google Scholar] [CrossRef]
- Khatun, J.; Loh, T.C.; Akit, H.; Foo, H.L.; Mohamad, R. Influence of different sources of oil on performance, meat quality, gut morphology, ileal digestibility and serum lipid profile in broilers. J. Appl. Anim. Res. 2018, 46, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Konieczka, P.; Barszcz, M.; Choct, M.; Smulikowska, S. The interactive effect of dietary n-6: N-3 fatty acid ratio and vitamin E level on tissue lipid peroxidation, DNA damage in intestinal epithelial cells, and gut morphology in chickens of different ages. Poult. Sci. 2018, 97, 149–158. [Google Scholar] [CrossRef]
- Dibner, J.J.; Atwell, C.A.; Kitchell, M.L.; Shermer, W.D.; Ivey, F.J. Feeding of oxidized fats to broilers and swine: Effects on enterocyte turnover hepatocyte proliferation and the gut associated lymphoid tissue. Anim. Feed. Sci. Technol. 1996, 62, 1–13. [Google Scholar] [CrossRef]
- Tan, L.; Rong, D.; Yang, Y.; Zhang, B. The effect of oxidized fish oils on growth performance, oxidative status, and intestinal barrier function in broiler chickens. J. Appl. Poult. Sci. 2019, 28, 31–41. [Google Scholar] [CrossRef]
- Adebowale, T.O.; Yao, K.; Oso, A.O. Major cereal carbohydrates in relation to intestinal health of monogastric animals: A review. Anim. Nutr. 2019, 5, 331–339. [Google Scholar] [CrossRef] [PubMed]
- M’Sadeq, S.A.; Wu, S.; Swick, R.A.; Choct, M. Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide. Anim. Nutr. 2015, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Adedokun, S.A.; Olojede, O.C. Optimizing gastrointestinal integrity in poultry: The role of nutrients and feed additives. Front. Vet. Sci. 2019, 5, 348. [Google Scholar] [CrossRef] [Green Version]
- Cardoso Dal Pont, G.; Farnell, M.; Farnell, Y.; Kogut, M.H. Dietary factors as triggers of low-grade chronic intestinal inflammation in poultry. Microorganisms 2020, 8, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaghobfar, A.; Kalantar, M. Effect of non-starch polysaccharide (NSP) of wheat and barley supplemented with exogenous enzyme blend on growth performance, gut microbial, pancreatic enzyme activities, expression of glucose transporter (SGLT1) and mucin producer (MUC2) genes of broiler chickens. Braz. J. Poult. Sci. 2017, 19, 629–638. [Google Scholar]
- Paraskeuas, V.; Mountzouris, K.C. Broiler gut microbiota and expressions of gut barrier genes affected by cereal type and phytogenic inclusion. Anim. Nutr. 2019, 5, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Ghayour-Najafabadi, P.; Khosravinia, H.; Gheisari, A.; Azafar, A.; Khanahmadi, M. Productive performance, nutrient digestibility and intestinal morphometry in broiler chickens fed corn or wheat-based diets supplemented with bacterial- or fungal-originated xylanase. Ital. J. Anim. Sci. 2018, 17, 165–174. [Google Scholar] [CrossRef]
- Regassa, A.; Nyachoti, C.M. Application of resistant starch in swine and poultry diets with particular reference to gut health and function. Anim. Nutr. 2018, 4, 305–310. [Google Scholar] [CrossRef]
- Thompson, K.L.; Applegate, T.J. Feed withdrawal alters small-intestinal morphology and mucus of broilers. Poult. Sci. 2006, 85, 1535–1540. [Google Scholar] [CrossRef]
- Yamauchi, K.; Kamisoyama, H.; Isshiki, Y. Effects of fasting and refeeding on structures of the intestinal villi and epithelial cells in White Leghorn hens. Brit. Poult. Sci. 1996, 37, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.E.P.; Martins, M.R.F.B.; Mendes, A.A.; Almeida, P.A.Z.I.C.L.; Komiyama, C.M.; Milbradt, E.L.; Fernandes, B.C.d.S. Effects of pre-slaughter fasting on broiler welfare, meat quality, and intestinal integrity. Braz. J. Poult. Sci. 2013, 15, 119–122. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Lilburn, M.; Yu, Z. Intestinal microbiota of broiler chickens as affected by litter management regimens. Front. Microbiol. 2016, 7, 593. [Google Scholar] [CrossRef] [Green Version]
- Cressman, M.D.; Yu, Z.; Nelson, M.C.; Moeller, S.J.; Lilburn, M.S.; Zerby, H.N. Interactions between the microbiotas in the litter and in the intestines of commercial broiler chickens. Appl. Environ. Microbiol. 2010, 76, 6572–6582. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.W.; Lillehoj, H.S.; Jeong, W.; Jeoung, H.Y.; An, D.J. Avian necrotic enteritis: Experimental models, host immunity, pathogenesis, risk factors, and vaccine development. Poult. Sci. 2011, 90, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Lillehoj, H.S.; Jang, S.I.; Lee, S.H.; Bautista, D.A.; Siragusa, G.R. Effect of Bacillus subtilis-based direct-fed microbials on immune status in broiler chickens raised on fresh or used litter. Asian-Aust. J. Anim. Sci. 2013, 26, 1592–1597. [Google Scholar] [CrossRef] [Green Version]
- Shanmugasundaram, R.; Lilburn, M.S.; Selvaraj, R.K. Effect of recycled litter on immune cells in the cecal tonsils of chickens. Poult. Sci. 2012, 91, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xiang, H.; Zhang, H.; Zhu, X.; Wang, D.; Wang, J.; Yin, T.; Liu, L.; Kong, M.; Li, H.; et al. Rearing system causes changes of behavior, microbiome, and gene expression of chickens. Poult. Sci. 2019, 98, 3365–3376. [Google Scholar] [CrossRef]
- Goo, D.; Kim, J.H.; Choi, H.S.; Park, G.H.; Han, G.P.; Kil, D.Y. Effect of stocking density and sex on growth performance, meat quality, and intestinal barrier function in broiler chickens. Poult. Sci. 2019, 98, 1153–1160. [Google Scholar] [CrossRef] [PubMed]
- Goo, D.; Kim, J.H.; Park, G.H.; Delos Reyes, J.B.; Kil, D.Y. Effect of stocking density and dietary tryptophan on growth performance and intestinal barrier function in broiler chickens. Poult. Sci. 2019, 98, 4504–4508. [Google Scholar] [CrossRef]
- Guardia, S.; Konsak, B.; Combes, S.; Levenez, F.; Cauquil, L.; Guillot, J.F.; Moreau-Vauzelle, C.; Lessire, M.; Juin, H.; Gabriel, I. Effects of stocking density on the growth performance and digestive microbiota of broiler chickens. Poult. Sci. 2011, 90, 1878–1889. [Google Scholar] [CrossRef] [PubMed]
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. High stocking density as a predisposing factor for necrotic enteritis in broiler chickens. Avian. Pathol. 2015, 44, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Jiao, L.F.; Xiao, K.; Luan, Z.S.; Hu, C.H.; Shi, B.; Zhan, X.A. Cello-oligosaccharide ameliorates heat stress-induced impairment of intestinal microflora, morphology and barrier integrity in broilers. Anim. Feed. Sci. Technol. 2013, 185, 175–181. [Google Scholar] [CrossRef]
- Song, J.; Xiao, K.; Ke, Y.L.; Jiao, L.F.; Hu, C.H.; Diao, Q.Y.; Shi, B.; Zou, X.T. Effect of a probiotic mixture on intestinal microflora, morphology, and barrier integrity of broilers subjected to heat stress. Poult. Sci. 2014, 93, 581–588. [Google Scholar] [CrossRef]
- Santos, R.R.; Awati, A.; Roubos-van den Hil, P.J.; van Keumpen, T.A.T.G.; Tersteeg-Zijderveld, M.H.G.; Koolmeers, P.A.; Smits, C.; Fink-Gremmels, J. Effects of a feed additive blend on broilers challenged with heat stress. Avian. Pathol. 2019, 48, 582–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varasteh, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Differences in susceptibility to heat stress along the chicken intestine and the protective effects of galacto-oligosaccharides. PLoS ONE 2015, 10, e0138975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. Heat stress as a predisposing factor for necrotic enteritis in broiler chickens. Avian Pathol. 2018, 47, 616–624. [Google Scholar] [CrossRef]
- Georgiev, M.; Beauvais, W.; Guitian, J. Effect of enhanced biosecurity and selected on-farm factors on Campylobacter colonization of chicken broilers. Epidemiol. Infect. 2017, 145, 553–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothrock, M.J., Jr.; Locatelli, A. Importance of farm environment to shape poultry-related microbiomes throughout the farm-to-fork continuum of pasture-raised broiler flocks. Front. Sustain. Food. Syst. 2019, 3, 48. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wickramasuriya, S.S.; Park, I.; Lee, K.; Lee, Y.; Kim, W.H.; Nam, H.; Lillehoj, H.S. Role of Physiology, Immunity, Microbiota, and Infectious Diseases in the Gut Health of Poultry. Vaccines 2022, 10, 172. https://doi.org/10.3390/vaccines10020172
Wickramasuriya SS, Park I, Lee K, Lee Y, Kim WH, Nam H, Lillehoj HS. Role of Physiology, Immunity, Microbiota, and Infectious Diseases in the Gut Health of Poultry. Vaccines. 2022; 10(2):172. https://doi.org/10.3390/vaccines10020172
Chicago/Turabian StyleWickramasuriya, Samiru S., Inkyung Park, Kyungwoo Lee, Youngsub Lee, Woo H. Kim, Hyoyoun Nam, and Hyun S. Lillehoj. 2022. "Role of Physiology, Immunity, Microbiota, and Infectious Diseases in the Gut Health of Poultry" Vaccines 10, no. 2: 172. https://doi.org/10.3390/vaccines10020172
APA StyleWickramasuriya, S. S., Park, I., Lee, K., Lee, Y., Kim, W. H., Nam, H., & Lillehoj, H. S. (2022). Role of Physiology, Immunity, Microbiota, and Infectious Diseases in the Gut Health of Poultry. Vaccines, 10(2), 172. https://doi.org/10.3390/vaccines10020172