
microRNA, a Subtle Indicator of Human Cytomegalovirus against Host Immune Cells


Abstract
:1. Introduction
2. Host Immune Response Overview to HCMV Infection
2.1. Innate Immune Responses
2.2. Adaptive Immune Response
3. The Regulation Mechanisms of Latent HCMV on Host Immune Cells
3.1. HCMV-Encoded miRNAs Inhibit Viral DNA Replication
3.2. HCMV-Encoded miRNAs Regulate Biological Functions of Host Immune Cells
3.3. HCMV-Encoded miRNAs Impair Host Inflammatory Signal Transduction Pathways
3.4. HCMV-Encoded miRNAs Induce Host Myelosuppression
3.5. Host miRNAs Regulation toward Viral Genes
4. Indispensable Role of HCMV-Encoded miRNAs in HCMV Reactivation
5. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cannon, M.J.; Schmid, D.S.; Hyde, T.B. Review of cytomegalovirus seroprevalence and demographic characteristics associated with infection. Rev. Med. Virol. 2010, 20, 202–213. [Google Scholar] [CrossRef]
- Struble, E.B.; Murata, H.; Komatsu, T.; Scott, D. Immune Prophylaxis and Therapy for Human Cytomegalovirus Infection. Int. J. Mol. Sci. 2021, 22, 8728. [Google Scholar] [CrossRef]
- Ahn, J.H.; Jang, W.J.; Hayward, G.S. The human cytomegalovirus IE2 and UL112-113 proteins accumulate in viral DNA replication compartments that initiate from the periphery of promyelocytic leukemia protein-associated nuclear bodies (PODs or ND10). J. Virol. 1999, 73, 10458–10471. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, A.E.; Mahjoob, M.O.; Abosalif, K.O.A.; Ejaz, H.; Alameen, A.A.M.; Elsaman, T. Human cytomegalovirus-encoded MicroRNAs: A master regulator of latent infection. Infect. Genet. Evol.: J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2020, 78, 104119. [Google Scholar] [CrossRef] [PubMed]
- Odeberg, J.; Plachter, B.; Brandén, L.; Söderberg-Nauclér, C. Human cytomegalovirus protein pp65 mediates accumulation of HLA-DR in lysosomes and destruction of the HLA-DR alpha-chain. Blood 2003, 101, 4870–4877. [Google Scholar] [CrossRef] [PubMed]
- Vanarsdall, A.L.; Howard, P.W.; Wisner, T.W.; Johnson, D.C. Human Cytomegalovirus gH/gL Forms a Stable Complex with the Fusion Protein gB in Virions. PLoS Pathog. 2016, 12, e1005564. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, P.; Reeves, M. Pathogenesis of human cytomegalovirus in the immunocompromised host. Nat. Rev. Microbiol. 2021, 19, 759–773. [Google Scholar] [CrossRef]
- Murray, M.J.; Peters, N.E.; Reeves, M.B. Navigating the Host Cell Response during Entry into Sites of Latent Cytomegalovirus Infection. Pathogens 2018, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elste, J.; Kaltenbach, D.; Patel, V.R.; Nguyen, M.T.; Sharthiya, H.; Tandon, R.; Mehta, S.K.; Volin, M.V.; Fornaro, M.; Tiwari, V.; et al. Inhibition of Human Cytomegalovirus Entry into Host Cells Through a Pleiotropic Small Molecule. Int. J. Mol. Sci. 2020, 21, 1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Rajewsky, N. The evolution of gene regulation by transcription factors and microRNAs. Nat. Rev. Genet. 2007, 8, 93–103. [Google Scholar] [CrossRef]
- Diggins, N.L.; Hancock, M.H. HCMV miRNA Targets Reveal Important Cellular Pathways for Viral Replication, Latency, and Reactivation. Non-Coding RNA 2018, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Pfeffer, S.; Zavolan, M.; Grässer, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of virus-encoded microRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Griffiths-Jones, S. The microRNA Registry. Nucleic Acids Res. 2004, 32, D109–D111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yu, J.; Liu, Z. MicroRNAs expressed by human cytomegalovirus. Virol. J. 2020, 17, 34. [Google Scholar] [CrossRef] [PubMed]
- Dell’Oste, V.; Biolatti, M.; Galitska, G.; Griffante, G.; Gugliesi, F.; Pasquero, S.; Zingoni, A.; Cerboni, C.; De Andrea, M. Tuning the Orchestra: HCMV vs. Innate Immunity. Front. Microbiol. 2020, 11, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira-Nascimento, L.; Massari, P.; Wetzler, L.M. The Role of TLR2 in Infection and Immunity. Front. Immunol. 2012, 3, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biolatti, M.; Gugliesi, F.; Dell’Oste, V.; Landolfo, S. Modulation of the innate immune response by human cytomegalovirus. Infect. Genet. Evol.: J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2018, 64, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Collins-McMillen, D.; Chesnokova, L.; Lee, B.J.; Fulkerson, H.L.; Brooks, R.; Mosher, B.S.; Yurochko, A.D. HCMV Infection and Apoptosis: How Do Monocytes Survive HCMV Infection? Viruses 2018, 10, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.; Pan, C.; Sheng, J.; Liang, H.; Bian, Z.; Liu, Y.; Trang, P.; Wu, J.; Liu, F.; Zhang, C.Y.; et al. Human cytomegalovirus reprogrammes haematopoietic progenitor cells into immunosuppressive monocytes to achieve latency. Nat. Microbiol. 2018, 3, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Kinast, V.; Döring, M.; Lipps, C.; Duran, V.; Spanier, J.; Tegtmeyer, P.K.; Wirth, D.; Cicin-Sain, L.; Alcamí, A.; et al. Human monocyte-derived macrophages inhibit HCMV spread independent of classical antiviral cytokines. Virulence 2018, 9, 1669–1684. [Google Scholar] [CrossRef] [Green Version]
- Paijo, J.; Döring, M.; Spanier, J.; Grabski, E.; Nooruzzaman, M.; Schmidt, T.; Witte, G.; Messerle, M.; Hornung, V.; Kaever, V.; et al. cGAS Senses Human Cytomegalovirus and Induces Type I Interferon Responses in Human Monocyte-Derived Cells. PLoS Pathog. 2016, 12, e1005546. [Google Scholar] [CrossRef] [PubMed]
- Patro, A.R.K. Subversion of Immune Response by Human Cytomegalovirus. Front. Immunol. 2019, 10, 1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, M.B.; Compton, T. Inhibition of inflammatory interleukin-6 activity via extracellular signal-regulated kinase-mitogen-activated protein kinase signaling antagonizes human cytomegalovirus reactivation from dendritic cells. J. Virol. 2011, 85, 12750–12758. [Google Scholar] [CrossRef] [Green Version]
- Dupont, L.; Du, L.; Poulter, M.; Choi, S.; McIntosh, M.; Reeves, M.B. Src family kinase activity drives cytomegalovirus reactivation by recruiting MOZ histone acetyltransferase activity to the viral promoter. J. Biol. Chem. 2019, 294, 12901–12910. [Google Scholar] [CrossRef] [Green Version]
- Kvale, E.; Dalgaard, J.; Lund-Johansen, F.; Rollag, H.; Farkas, L.; Midtvedt, K.; Jahnsen, F.L.; Brinchmann, J.E.; Olweus, J. CD11c+ dendritic cells and plasmacytoid DCs are activated by human cytomegalovirus and retain efficient T cell-stimulatory capability upon infection. Blood 2006, 107, 2022–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lung, T.L.; Saurwein-Teissl, M.; Parson, W.; Schönitzer, D.; Grubeck-Loebenstein, B. Unimpaired dendritic cells can be derived from monocytes in old age and can mobilize residual function in senescent T cells. Vaccine 2000, 18, 1606–1612. [Google Scholar] [CrossRef]
- Deal, E.M.; Lahl, K.; Narváez, C.F.; Butcher, E.C.; Greenberg, H.B. Plasmacytoid dendritic cells promote rotavirus-induced human and murine B cell responses. J. Clin. Investig. 2013, 123, 2464–2474. [Google Scholar] [CrossRef] [Green Version]
- Seidel, E.; Dassa, L.; Schuler, C.; Oiknine-Djian, E.; Wolf, D.G.; Le-Trilling, V.T.K.; Mandelboim, O. The human cytomegalovirus protein UL147A downregulates the most prevalent MICA allele: MICA*008, to evade NK cell-mediated killing. PLoS Pathog. 2021, 17, e1008807. [Google Scholar] [CrossRef]
- Fielding, C.A.; Aicheler, R.; Stanton, R.J.; Wang, E.C.; Han, S.; Seirafian, S.; Davies, J.; McSharry, B.P.; Weekes, M.P.; Antrobus, P.R.; et al. Two novel human cytomegalovirus NK cell evasion functions target MICA for lysosomal degradation. PLoS Pathog. 2014, 10, e1004058. [Google Scholar] [CrossRef] [Green Version]
- Lanier, L.L. NKG2D Receptor and Its Ligands in Host Defense. Cancer Immunol. Res. 2015, 3, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Gerna, G.; Lilleri, D. Human cytomegalovirus (HCMV) infection/re-infection: Development of a protective HCMV vaccine. New Microbiol. 2019, 42, 1–20. [Google Scholar] [PubMed]
- Gyurova, I.E.; Ali, A.; Waggoner, S.N. Natural Killer Cell Regulation of B Cell Responses in the Context of Viral Infection. Viral Immunol. 2020, 33, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Q.; Romagnani, C. About Training and Memory: NK-Cell Adaptation to Viral Infections. Adv. Immunol. 2017, 133, 171–207. [Google Scholar] [CrossRef]
- Lewis, G.K.; Pazgier, M.; Evans, D.T.; Ferrari, G.; Bournazos, S.; Parsons, M.S.; Bernard, N.F.; Finzi, A. Beyond Viral Neutralization. AIDS Res. Hum. Retrovir. 2017, 33, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Della Chiesa, M.; De Maria, A.; Muccio, L.; Bozzano, F.; Sivori, S.; Moretta, L. Human NK Cells and Herpesviruses: Mechanisms of Recognition, Response and Adaptation. Front. Microbiol. 2019, 10, 2297. [Google Scholar] [CrossRef]
- Bottino, C.; Castriconi, R.; Pende, D.; Rivera, P.; Nanni, M.; Carnemolla, B.; Cantoni, C.; Grassi, J.; Marcenaro, S.; Reymond, N.; et al. Identification of PVR (CD155) and Nectin-2 (CD112) as cell surface ligands for the human DNAM-1 (CD226) activating molecule. J. Exp. Med. 2003, 198, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Prod’homme, V.; Sugrue, D.M.; Stanton, R.J.; Nomoto, A.; Davies, J.; Rickards, C.R.; Cochrane, D.; Moore, M.; Wilkinson, G.W.G.; Tomasec, P. Human cytomegalovirus UL141 promotes efficient downregulation of the natural killer cell activating ligand CD112. J. Gen. Virol. 2010, 91, 2034–2039. [Google Scholar] [CrossRef]
- Muntasell, A.; Costa-Garcia, M.; Vera, A.; Marina-Garcia, N.; Kirschning, C.J.; López-Botet, M. Priming of NK cell anti-viral effector mechanisms by direct recognition of human cytomegalovirus. Front. Immunol. 2013, 4, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holder, K.A.; Grant, M.D. Human cytomegalovirus IL-10 augments NK cell cytotoxicity. J. Leukoc. Biol. 2019, 106, 447–454. [Google Scholar] [CrossRef]
- Abassi, L.; Cicin-Sain, L. The avid competitors of memory inflation. Curr. Opin. Virol. 2020, 44, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Parry, H.M.; Dowell, A.C.; Zuo, J.; Verma, K.; Kinsella, F.A.M.; Begum, J.; Croft, W.; Sharma-Oates, A.; Pratt, G.; Moss, P. PD-1 is imprinted on cytomegalovirus-specific CD4+ T cells and attenuates Th1 cytokine production whilst maintaining cytotoxicity. PLoS Pathog. 2021, 17, e1009349. [Google Scholar] [CrossRef] [PubMed]
- Hertoghs, K.M.; Moerland, P.D.; van Stijn, A.; Remmerswaal, E.B.; Yong, S.L.; van de Berg, P.J.; van Ham, S.M.; Baas, F.; ten Berge, I.J.; van Lier, R.A. Molecular profiling of cytomegalovirus-induced human CD8+ T cell differentiation. J. Clin. Investig. 2010, 120, 4077–4090. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.; Perez, L. Virus-Like Particles and Nanoparticles for Vaccine Development against HCMV. Viruses 2019, 12, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira Braga, F.A.; Hertoghs, K.M.; van Lier, R.A.; van Gisbergen, K.P. Molecular characterization of HCMV-specific immune responses: Parallels between CD8(+) T cells, CD4(+) T cells, and NK cells. Eur. J. Immunol. 2015, 45, 2433–2445. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, S.; Shin, J.; Kim, Y.; Evnouchidou, I.; Kim, D.; Kim, Y.K.; Kim, Y.E.; Ahn, J.H.; Riddell, S.R.; et al. Human cytomegalovirus microRNA miR-US4-1 inhibits CD8(+) T cell responses by targeting the aminopeptidase ERAP1. Nat. Immunol. 2011, 12, 984–991. [Google Scholar] [CrossRef]
- Woods, E.; Zaiatz-Bittencourt, V.; Bannan, C.; Bergin, C.; Finlay, D.K.; Hoffmann, M.; Brown, A.; Turner, B.; Makvandi-Nejad, S.; Vassilev, V.; et al. Specific human cytomegalovirus signature detected in NK cell metabolic changes post vaccination. NPJ Vaccines 2021, 6, 117. [Google Scholar] [CrossRef]
- Rölle, A.; Brodin, P. Immune Adaptation to Environmental Influence: The Case of NK Cells and HCMV. Trends Immunol. 2016, 37, 233–243. [Google Scholar] [CrossRef]
- Schlums, H.; Cichocki, F.; Tesi, B.; Theorell, J.; Beziat, V.; Holmes, T.D.; Han, H.; Chiang, S.C.; Foley, B.; Mattsson, K.; et al. Cytomegalovirus infection drives adaptive epigenetic diversification of NK cells with altered signaling and effector function. Immunity 2015, 42, 443–456. [Google Scholar] [CrossRef] [Green Version]
- Inácio, D.P.; Amado, T.; Silva-Santos, B.; Gomes, A.Q. Control of T cell effector functions by miRNAs. Cancer Lett. 2018, 427, 63–73. [Google Scholar] [CrossRef]
- Bendelac, A.; Savage, P.B.; Teyton, L. The biology of NKT cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Xie, S.; Chen, M.; Li, Y.; Yue, J.; Ma, J.; Shu, X.; He, Y.; Xiao, W.; Tian, Z. Advances in NK cell production. Cell. Mol. Immunol. 2022. [Google Scholar] [CrossRef]
- Piedade, D.; Azevedo-Pereira, J.M. The Role of microRNAs in the Pathogenesis of Herpesvirus Infection. Viruses 2016, 8, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schommartz, T.; Tang, J.; Brost, R.; Brune, W. Differential Requirement of Human Cytomegalovirus UL112-113 Protein Isoforms for Viral Replication. J. Virol. 2017, 91, e00254-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern-Ginossar, N.; Saleh, N.; Goldberg, M.D.; Prichard, M.; Wolf, D.G.; Mandelboim, O. Analysis of human cytomegalovirus-encoded microRNA activity during infection. J. Virol. 2009, 83, 10684–10693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.; Zhu, D.; Wang, Y.; Li, L.; Li, D.; Liu, F.; Zhang, C.Y.; Zen, K. Human Cytomegalovirus miR-UL148D Facilitates Latent Viral Infection by Targeting Host Cell Immediate Early Response Gene 5. PLoS Pathog. 2016, 12, e1006007. [Google Scholar] [CrossRef] [PubMed]
- Grey, F.; Tirabassi, R.; Meyers, H.; Wu, G.; McWeeney, S.; Hook, L.; Nelson, J.A. A viral microRNA down-regulates multiple cell cycle genes through mRNA 5’UTRs. PLoS Pathog. 2010, 6, e1000967. [Google Scholar] [CrossRef] [Green Version]
- Qi, M.; Qi, Y.; Ma, Y.; He, R.; Ji, Y.; Sun, Z.; Ruan, Q. Over-expression of human cytomegalovirus miR-US25-2-3p downregulates eIF4A1 and inhibits HCMV replication. FEBS Lett. 2013, 587, 2266–2271. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Qi, Y.; Huang, Y.; Liu, Z.; Ma, Y.; Shao, Y.; Jiang, S.; Sun, Z.; Ruan, Q. Human cytomegalovirus miR-US33-5p inhibits viral DNA synthesis and viral replication by down-regulating expression of the host Syntaxin3. FEBS Lett. 2015, 589, 440–446. [Google Scholar] [CrossRef]
- Kim, S.; Seo, D.; Kim, D.; Hong, Y.; Chang, H.; Baek, D.; Kim, V.N.; Lee, S.; Ahn, K. Temporal Landscape of MicroRNA-Mediated Host-Virus Crosstalk during Productive Human Cytomegalovirus Infection. Cell Host Microbe 2015, 17, 838–851. [Google Scholar] [CrossRef] [Green Version]
- Stern-Ginossar, N.; Elefant, N.; Zimmermann, A.; Wolf, D.G.; Saleh, N.; Biton, M.; Horwitz, E.; Prokocimer, Z.; Prichard, M.; Hahn, G.; et al. Host immune system gene targeting by a viral miRNA. Science 2007, 317, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Fruci, D.; Rota, R.; Gallo, A. The Role of HCMV and HIV-1 MicroRNAs: Processing, and Mechanisms of Action during Viral Infection. Front. Microbiol. 2017, 8, 689. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Lee, S.; Cho, D.Y.; Lee, S.; Kim, H.; Yu, N.; Lee, S.; Ahn, K. Human cytomegalovirus induces and exploits Roquin to counteract the IRF1-mediated antiviral state. Proc. Natl. Acad. Sci. USA 2019, 116, 18619–18628. [Google Scholar] [CrossRef] [Green Version]
- Romania, P.; Cifaldi, L.; Pignoloni, B.; Starc, N.; D’Alicandro, V.; Melaiu, O.; Li Pira, G.; Giorda, E.; Carrozzo, R.; Bergvall, M.; et al. Identification of a Genetic Variation in ERAP1 Aminopeptidase that Prevents Human Cytomegalovirus miR-UL112-5p-Mediated Immunoevasion. Cell Rep. 2017, 20, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Ding, M.; Wang, X.; Wang, C.; Liu, X.; Zen, K.; Wang, W.; Zhang, C.Y.; Zhang, C. Distinct expression profile of HCMV encoded miRNAs in plasma from oral lichen planus patients. J. Transl. Med. 2017, 15, 133. [Google Scholar] [CrossRef] [Green Version]
- Tirabassi, R.; Hook, L.; Landais, I.; Grey, F.; Meyers, H.; Hewitt, H.; Nelson, J. Human cytomegalovirus US7 is regulated synergistically by two virally encoded microRNAs and by two distinct mechanisms. J. Virol. 2011, 85, 11938–11944. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Song, X.; Ma, P.; Lv, L.; Zhang, Y.; Deng, J.; Zhang, Y. Human Cytomegalovirus miR-US33as-5p Targets IFNAR1 to Achieve Immune Evasion During Both Lytic and Latent Infection. Front. Immunol. 2021, 12, 628364. [Google Scholar] [CrossRef]
- Wang, Y.P.; Qi, Y.; Huang, Y.J.; Qi, M.L.; Ma, Y.P.; He, R.; Ji, Y.H.; Sun, Z.R.; Ruan, Q. Identification of immediate early gene X-1 as a cellular target gene of hcmv-mir-UL148D. Int. J. Mol. Med. 2013, 31, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Babu, S.G.; Pandeya, A.; Verma, N.; Shukla, N.; Kumar, R.V.; Saxena, S. Role of HCMV miR-UL70-3p and miR-UL148D in overcoming the cellular apoptosis. Mol. Cell. Biochem. 2014, 393, 89–98. [Google Scholar] [CrossRef]
- Guo, X.; Huang, Y.; Qi, Y.; Liu, Z.; Ma, Y.; Shao, Y.; Jiang, S.; Sun, Z.; Ruan, Q. Human cytomegalovirus miR-UL36-5p inhibits apoptosis via downregulation of adenine nucleotide translocator 3 in cultured cells. Arch. Virol. 2015, 160, 2483–2490. [Google Scholar] [CrossRef]
- Stark, T.J.; Arnold, J.D.; Spector, D.H.; Yeo, G.W. High-resolution profiling and analysis of viral and host small RNAs during human cytomegalovirus infection. J. Virol. 2012, 86, 226–235. [Google Scholar] [CrossRef] [Green Version]
- Hancock, M.H.; Hook, L.M.; Mitchell, J.; Nelson, J.A. Human Cytomegalovirus MicroRNAs miR-US5-1 and miR-UL112-3p Block Proinflammatory Cytokine Production in Response to NF-κB-Activating Factors through Direct Downregulation of IKKα and IKKβ. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Landais, I.; Pelton, C.; Streblow, D.; DeFilippis, V.; McWeeney, S.; Nelson, J.A. Human Cytomegalovirus miR-UL112-3p Targets TLR2 and Modulates the TLR2/IRAK1/NFκB Signaling Pathway. PLoS Pathog. 2015, 11, e1004881. [Google Scholar] [CrossRef] [PubMed]
- Hook, L.M.; Grey, F.; Grabski, R.; Tirabassi, R.; Doyle, T.; Hancock, M.; Landais, I.; Jeng, S.; McWeeney, S.; Britt, W.; et al. Cytomegalovirus miRNAs target secretory pathway genes to facilitate formation of the virion assembly compartment and reduce cytokine secretion. Cell Host Microbe 2014, 15, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Jones, K.L.; Mansell, A.; Patella, S.; Scott, B.J.; Hedger, M.P.; de Kretser, D.M.; Phillips, D.J. Activin A is a critical component of the inflammatory response, and its binding protein, follistatin, reduces mortality in endotoxemia. Proc. Natl. Acad. Sci. USA 2007, 104, 16239–16244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Lee, S.; Kim, S.; Kim, D.; Ahn, J.H.; Ahn, K. Human cytomegalovirus clinical strain-specific microRNA miR-UL148D targets the human chemokine RANTES during infection. PLoS Pathog. 2012, 8, e1002577. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xia, S.; Yang, X.; Chen, H.; Li, F.; Liu, F.; Chen, Z. Human Cytomegalovirus Encoded miR-US25-1-5p Attenuates CD147/EMMPRIN-Mediated Early Antiviral Response. Viruses 2017, 9, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, M.H.; Crawford, L.B.; Pham, A.H.; Mitchell, J.; Struthers, H.M.; Yurochko, A.D.; Caposio, P.; Nelson, J.A. Human Cytomegalovirus miRNAs Regulate TGF-β to Mediate Myelosuppression while Maintaining Viral Latency in CD34(+) Hematopoietic Progenitor Cells. Cell Host Microbe 2020, 27, 104–114. [Google Scholar] [CrossRef]
- Lee, S.H.; Kalejta, R.F.; Kerry, J.; Semmes, O.J.; O’Connor, C.M.; Khan, Z.; Garcia, B.A.; Shenk, T.; Murphy, E. BclAF1 restriction factor is neutralized by proteasomal degradation and microRNA repression during human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2012, 109, 9575–9580. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Qi, Y.; Ma, Y.; He, R.; Ji, Y.; Sun, Z.; Ruan, Q. Down-regulation of human cytomegalovirus UL138, a novel latency-associated determinant, by hcmv-miR-UL36. J. Biosci. 2013, 38, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Huang, Y.; Qi, Y.; He, R.; Liu, Z.; Ma, Y.; Guo, X.; Shao, Y.; Sun, Z.; Ruan, Q. Human cytomegalovirus miR-US5-1 inhibits viral replication by targeting Geminin mRNA. Virol. Sin. 2017, 32, 431–439. [Google Scholar] [CrossRef]
- Mikell, I.; Crawford, L.B.; Hancock, M.H.; Mitchell, J.; Buehler, J.; Goodrum, F.; Nelson, J.A. HCMV miR-US22 down-regulation of EGR-1 regulates CD34+ hematopoietic progenitor cell proliferation and viral reactivation. PLoS Pathog. 2019, 15, e1007854. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Qi, Y.; Huang, Y.; Liu, Z.; Ma, Y.; Guo, X.; Jiang, S.; Sun, Z.; Ruan, Q. Human cytomegalovirus miR-US4-5p promotes apoptosis via downregulation of p21-activated kinase 2 in cultured cells. Mol. Med. Rep. 2017, 16, 4171–4178. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Zhang, W.; Liu, Q. Human cytomegalovirus-encoded miR-US25-1 aggravates the oxidised low density lipoprotein-induced apoptosis of endothelial cells. BioMed Res. Int. 2014, 2014, 531979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisboa, L.F.; Egli, A.; O’Shea, D.; Åsberg, A.; Hartmann, A.; Rollag, H.; Pang, X.L.; Tyrrell, D.L.; Kumar, D.; Humar, A. Hcmv-miR-UL22A-5p: A Biomarker in Transplantation With Broad Impact on Host Gene Expression and Potential Immunological Implications. Am. J. Transplant.: Off. J. Am. Soc. Transplant. Am. Soc. Transpl. Surg. 2015, 15, 1893–1902. [Google Scholar] [CrossRef]
- Shen, Z.Z.; Pan, X.; Miao, L.F.; Ye, H.Q.; Chavanas, S.; Davrinche, C.; McVoy, M.; Luo, M.H. Comprehensive analysis of human cytomegalovirus microRNA expression during lytic and quiescent infection. PLoS ONE 2014, 9, e88531. [Google Scholar] [CrossRef] [Green Version]
- Heider, J.A.; Bresnahan, W.A.; Shenk, T.E. Construction of a rationally designed human cytomegalovirus variant encoding a temperature-sensitive immediate-early 2 protein. Proc. Natl. Acad. Sci. USA 2002, 99, 3141–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møller, R.; Schwarz, T.M.; Noriega, V.M.; Panis, M.; Sachs, D.; Tortorella, D.; tenOever, B.R. miRNA-mediated targeting of human cytomegalovirus reveals biological host and viral targets of IE2. Proc. Natl. Acad. Sci. USA 2018, 115, 1069–1074. [Google Scholar] [CrossRef] [Green Version]
- Shan, L.; Li, S.; Meeldijk, J.; Blijenberg, B.; Hendriks, A.; van Boxtel, K.; van den Berg, S.P.H.; Groves, I.J.; Potts, M.; Svrlanska, A.; et al. Killer cell proteases can target viral immediate-early proteins to control human cytomegalovirus infection in a noncytotoxic manner. PLoS Pathog. 2020, 16, e1008426. [Google Scholar] [CrossRef] [Green Version]
- Lau, B.; Poole, E.; Van Damme, E.; Bunkens, L.; Sowash, M.; King, H.; Murphy, E.; Wills, M.; Van Loock, M.; Sinclair, J. Human cytomegalovirus miR-UL112-1 promotes the down-regulation of viral immediate early-gene expression during latency to prevent T-cell recognition of latently infected cells. J. Gen. Virol. 2016, 97, 2387–2398. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, C.; Ding, M.; Bian, Y.; Zhong, Y.; Shen, H.; Wang, J.; Zhang, C.Y.; Zhang, C. Different expression pattern of human cytomegalovirus-encoded microRNAs in circulation from virus latency to reactivation. J. Transl. Med. 2020, 18, 469. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Qi, Y.; He, R.; Huang, Y.; Liu, Z.; Ma, Y.; Guo, X.; Shao, Y.; Sun, Z.; Ruan, Q. Human cytomegalovirus microRNA miR-US25-1-5p inhibits viral replication by targeting multiple cellular genes during infection. Gene 2015, 570, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Close, W.L.; Glassbrook, J.E.; Gurczynski, S.J.; Pellett, P.E. Infection-Induced Changes Within the Endocytic Recycling Compartment Suggest a Roadmap of Human Cytomegalovirus Egress. Front. Microbiol. 2018, 9, 1888. [Google Scholar] [CrossRef] [Green Version]
- Diggins, N.L.; Skalsky, R.L.; Hancock, M.H. Regulation of Latency and Reactivation by Human Cytomegalovirus miRNAs. Pathogens 2021, 10, 200. [Google Scholar] [CrossRef]
- Krestinina, O.; Baburina, Y.; Krestinin, R. Mitochondrion as a Target of Astaxanthin Therapy in Heart Failure. Int. J. Mol. Sci. 2021, 22, 7964. [Google Scholar] [CrossRef]
- Sha, Z.; Goldberg, A.L. Multiple myeloma cells are exceptionally sensitive to heat shock, which overwhelms their proteostasis network and induces apoptosis. Proc. Natl. Acad. Sci. USA 2020, 117, 21588–21597. [Google Scholar] [CrossRef]
- Jiang, X.; Kim, H.E.; Shu, H.; Zhao, Y.; Zhang, H.; Kofron, J.; Donnelly, J.; Burns, D.; Ng, S.C.; Rosenberg, S.; et al. Distinctive roles of PHAP proteins and prothymosin-alpha in a death regulatory pathway. Science 2003, 299, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Hancock, M.H.; Crawford, L.B.; Perez, W.; Struthers, H.M.; Mitchell, J.; Caposio, P. Human Cytomegalovirus UL7, miR-US5-1, and miR-UL112-3p Inactivation of FOXO3a Protects CD34(+) Hematopoietic Progenitor Cells from Apoptosis. mSphere 2021, 6, e00986-20. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, D.; He, J.; Cai, J.; Shen, K.; Liu, X.; Yang, X.; Xu, L. Hcmv-miR-UL112 attenuates NK cell activity by inhibition type I interferon secretion. Immunol. Lett. 2015, 163, 151–156. [Google Scholar] [CrossRef]
- Huang, Y.; Qi, Y.; Ma, Y.; He, R.; Ji, Y.; Sun, Z.; Ruan, Q. The expression of interleukin-32 is activated by human cytomegalovirus infection and down regulated by hcmv-miR-UL112-1. Virol. J. 2013, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Esteso, G.; Luzón, E.; Sarmiento, E.; Gómez-Caro, R.; Steinle, A.; Murphy, G.; Carbone, J.; Valés-Gómez, M.; Reyburn, H.T. Altered microRNA expression after infection with human cytomegalovirus leads to TIMP3 downregulation and increased shedding of metalloprotease substrates, including MICA. J. Immunol. 2014, 193, 1344–1352. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Lehar, A.; Meir, J.U.; Koch, C.; Morgan, A.; Warren, L.E.; Rydzik, R.; Youngstrom, D.W.; Chandok, H.; George, J.; et al. Targeting myostatin/activin A protects against skeletal muscle and bone loss during spaceflight. Proc. Natl. Acad. Sci. USA 2020, 117, 23942–23951. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.; Poole, E.; Krishna, B.; Sellart, I.; Wills, M.R.; Murphy, E.; Sinclair, J. The Expression of Human Cytomegalovirus MicroRNA MiR-UL148D during Latent Infection in Primary Myeloid Cells Inhibits Activin A-triggered Secretion of IL-6. Sci. Rep. 2016, 6, 31205. [Google Scholar] [CrossRef] [PubMed]
- Vacinova, G.; Vejražkova, D.; Rusina, R.; Holmerová, I.; Vaňková, H.; Jarolímová, E.; Včelák, J.; Bendlová, B.; Vaňková, M. Regulated upon activation, normal T cell expressed and secreted (RANTES) levels in the peripheral blood of patients with Alzheimer’s disease. Neural Regen. Res. 2021, 16, 796–800. [Google Scholar] [CrossRef]
- Admon, A. ERAP1 shapes just part of the immunopeptidome. Hum. Immunol. 2019, 80, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Melaiu, O.; D’Amico, S.; Tempora, P.; Lucarini, V.; Fruci, D. Impact of Natural Occurring ERAP1 Single Nucleotide Polymorphisms within miRNA-Binding Sites on HCMV Infection. Int. J. Mol. Sci. 2020, 21, 5861. [Google Scholar] [CrossRef]
- Compton, T.; Kurt-Jones, E.A.; Boehme, K.W.; Belko, J.; Latz, E.; Golenbock, D.T.; Finberg, R.W. Human cytomegalovirus activates inflammatory cytokine responses via CD14 and Toll-like receptor 2. J. Virol. 2003, 77, 4588–4596. [Google Scholar] [CrossRef] [Green Version]
- Biswas, C.; Zhang, Y.; DeCastro, R.; Guo, H.; Nakamura, T.; Kataoka, H.; Nabeshima, K. The human tumor cell-derived collagenase stimulatory factor (renamed EMMPRIN) is a member of the immunoglobulin superfamily. Cancer Res. 1995, 55, 434–439. [Google Scholar]
- Yurchenko, V.; Zybarth, G.; O’Connor, M.; Dai, W.W.; Franchin, G.; Hao, T.; Guo, H.; Hung, H.C.; Toole, B.; Gallay, P.; et al. Active site residues of cyclophilin A are crucial for its signaling activity via CD147. J. Biol. Chem. 2002, 277, 22959–22965. [Google Scholar] [CrossRef] [Green Version]
- Yurochko, A.D. New Mechanism by Which Human Cytomegalovirus MicroRNAs Negate the Proinflammatory Response to Infection. mBio 2017, 8, e00505-17. [Google Scholar] [CrossRef] [Green Version]
- Torok-Storb, B.; Simmons, P.; Khaira, D.; Stachel, D.; Myerson, D. Cytomegalovirus and marrow function. Ann. Hematol. 1992, 64, A128–A131. [Google Scholar] [CrossRef]
- Hu, M.; Lu, Y.; Wang, S.; Zhang, Z.; Qi, Y.; Chen, N.; Shen, M.; Chen, F.; Chen, M.; Yang, L.; et al. CD63 acts as a functional marker in maintaining hematopoietic stem cell quiescence through supporting TGFβ signaling in mice. Cell Death Differ. 2022, 29, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Challen, G.A.; Boles, N.C.; Chambers, S.M.; Goodell, M.A. Distinct hematopoietic stem cell subtypes are differentially regulated by TGF-beta1. Cell Stem Cell 2010, 6, 265–278. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, C.M.; Vanicek, J.; Murphy, E.A. Host microRNA regulation of human cytomegalovirus immediate early protein translation promotes viral latency. J. Virol. 2014, 88, 5524–5532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.H.; Lim, B.J.; Ferreira, V.H.; Min, S.Y.; Hong, Y.M.; Jo, J.H.; Han, S.H. Expression of human miR-200b-3p and -200c-3p in cytomegalovirus-infected tissues. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Asou, H.; Suzukawa, K.; Kita, K.; Nakase, K.; Ueda, H.; Morishita, K.; Kamada, N. Establishment of an undifferentiated leukemia cell line (Kasumi-3) with t(3;7)(q27;q22) and activation of the EVI1 gene. Jpn. J. Cancer Res. Gann 1996, 87, 269–274. [Google Scholar] [CrossRef]
- Krishna, B.A.; Lau, B.; Jackson, S.E.; Wills, M.R.; Sinclair, J.H.; Poole, E. Transient activation of human cytomegalovirus lytic gene expression during latency allows cytotoxic T cell killing of latently infected cells. Sci. Rep. 2016, 6, 24674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Teng, J.; Cui, C.; Li, D.; Wen, L. MicroRNA-182 inhibits HCMV replication through activation of type I IFN response by targeting FOXO3 in neural cells. Exp. Cell Res. 2018, 369, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Meshesha, M.K.; Bentwich, Z.; Solomon, S.A.; Avni, Y.S. In vivo expression of human cytomegalovirus (HCMV) microRNAs during latency. Gene 2016, 575, 101–107. [Google Scholar] [CrossRef]
- Hancock, M.H.; Tirabassi, R.S.; Nelson, J.A. Rhesus cytomegalovirus encodes seventeen microRNAs that are differentially expressed in vitro and in vivo. Virology 2012, 425, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, C.; Grey, F.; Kreklywich, C.N.; Andoh, T.F.; Tirabassi, R.S.; Orloff, S.L.; Streblow, D.N. Cytomegalovirus microRNA expression is tissue specific and is associated with persistence. J. Virol. 2011, 85, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.S.; Goldman, D.C.; Bailey, A.S.; Pfaffle, D.L.; Kreklywich, C.N.; Spencer, D.B.; Othieno, F.A.; Streblow, D.N.; Garcia, J.V.; Fleming, W.H.; et al. Granulocyte-colony stimulating factor reactivates human cytomegalovirus in a latently infected humanized mouse model. Cell Host Microbe 2010, 8, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Stern, L.; Withers, B.; Avdic, S.; Gottlieb, D.; Abendroth, A.; Blyth, E.; Slobedman, B. Human Cytomegalovirus Latency and Reactivation in Allogeneic Hematopoietic Stem Cell Transplant Recipients. Front. Microbiol. 2019, 10, 1186. [Google Scholar] [CrossRef] [Green Version]
- Reeves, M.B.; Sinclair, J.H. Circulating dendritic cells isolated from healthy seropositive donors are sites of human cytomegalovirus reactivation in vivo. J. Virol. 2013, 87, 10660–10667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, I.M.; Pietramaggiori, G.; Kim, F.S.; Passegué, E.; Stevenson, K.E.; Wagers, A.J. The transcription factor EGR1 controls both the proliferation and localization of hematopoietic stem cells. Cell Stem Cell 2008, 2, 380–391. [Google Scholar] [CrossRef] [Green Version]
- Hancock, M.H.; Mitchell, J.; Goodrum, F.D.; Nelson, J.A. Human Cytomegalovirus miR-US5-2 Downregulation of GAB1 Regulates Cellular Proliferation and UL138 Expression through Modulation of Epidermal Growth Factor Receptor Signaling Pathways. mSphere 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Diggins, N.L.; Crawford, L.B.; Hancock, M.H.; Mitchell, J.; Nelson, J.A. Human Cytomegalovirus miR-US25-1 Targets the GTPase RhoA To Inhibit CD34(+) Hematopoietic Progenitor Cell Proliferation To Maintain the Latent Viral Genome. mBio 2021, 12. [Google Scholar] [CrossRef]
- Cui, X.; Snapper, C.M. Development of novel vaccines against human cytomegalovirus. Hum. Vaccines Immunother. 2019, 15, 2673–2683. [Google Scholar] [CrossRef] [Green Version]
- Keshavarz, M.; Mirzaei, H.; Salemi, M.; Momeni, F.; Mousavi, M.J.; Sadeghalvad, M.; Arjeini, Y.; Solaymani-Mohammadi, F.; Sadri Nahand, J.; Namdari, H.; et al. Influenza vaccine: Where are we and where do we go? Rev. Med. Virol. 2019, 29, e2014. [Google Scholar] [CrossRef]
- Perez, J.T.; Pham, A.M.; Lorini, M.H.; Chua, M.A.; Steel, J.; tenOever, B.R. MicroRNA-mediated species-specific attenuation of influenza A virus. Nat. Biotechnol. 2009, 27, 572–576. [Google Scholar] [CrossRef]
- Fernández-Moreno, R.; Torre-Cisneros, J.; Cantisán, S. Human cytomegalovirus (HCMV)-encoded microRNAs: Potential biomarkers and clinical applications. RNA Biol. 2021, 18, 2194–2202. [Google Scholar] [CrossRef]
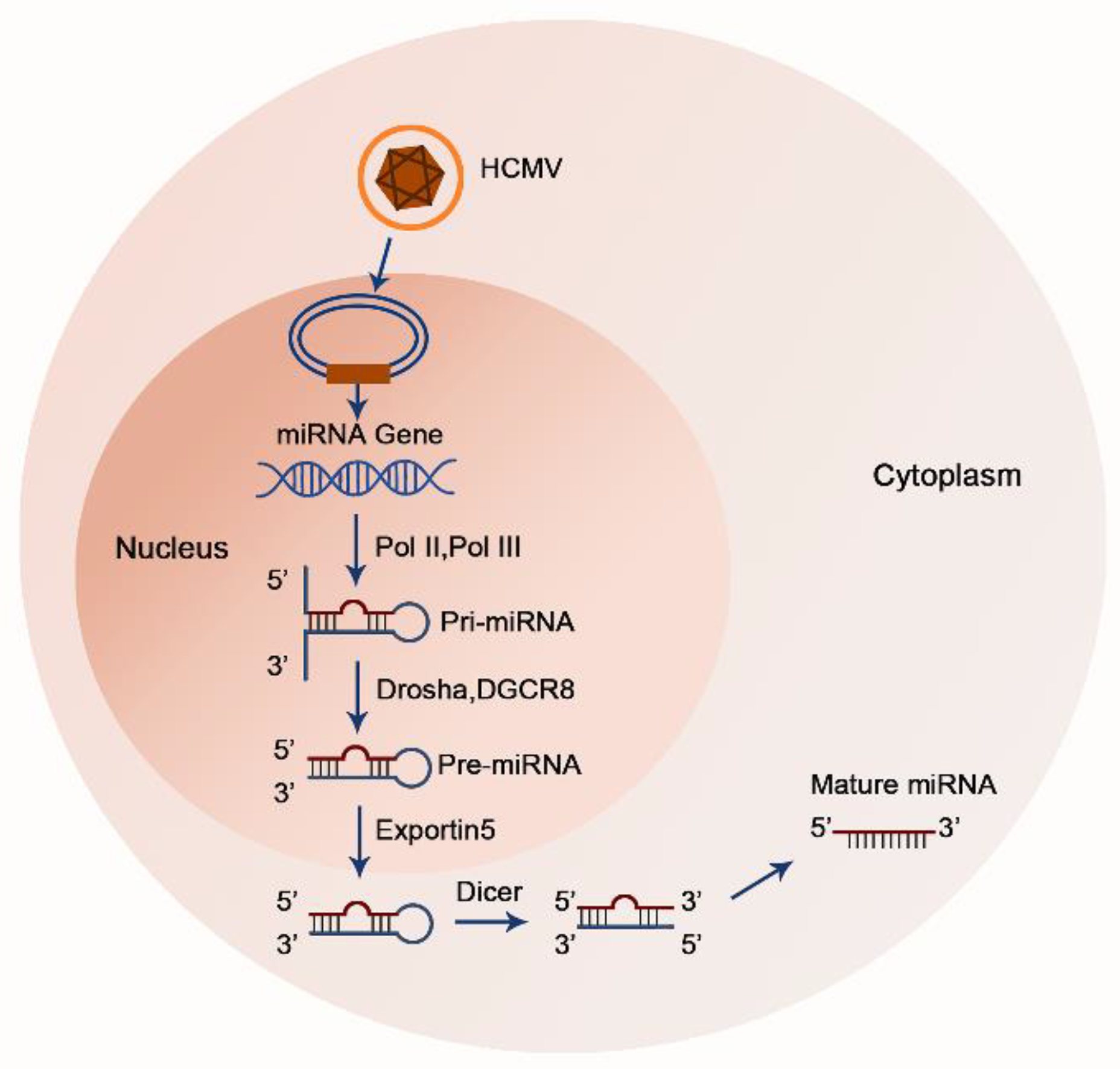
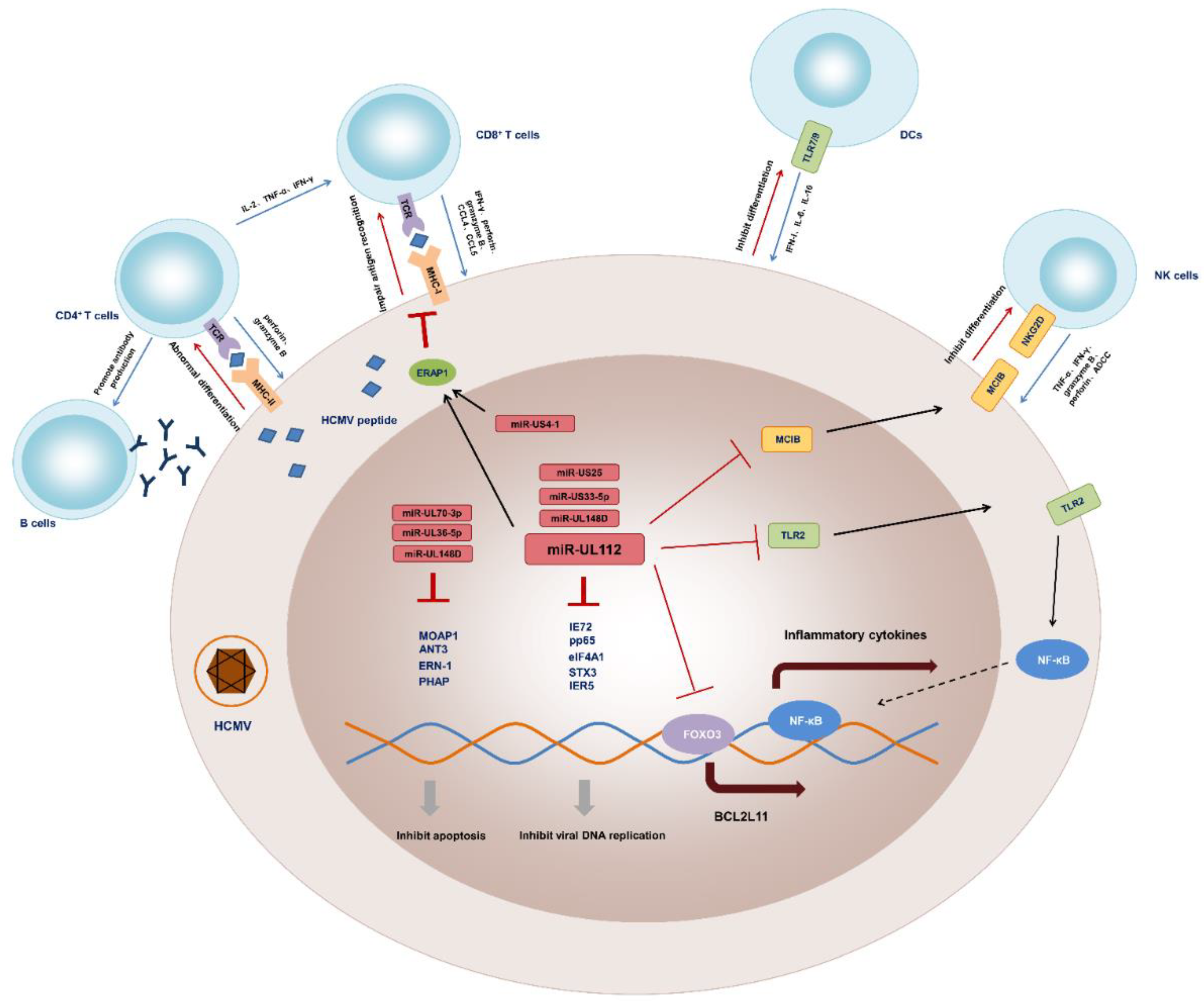

{kind=link}
{kind=link}
| Infection Stage | Function | HCMV-Encoded miRNA | Targets | References |
|---|---|---|---|---|
| Latency | Limit viral gene expression | miR-UL112-3p | HCMV IE72 | [4] |
| HCMV UL112/113 | [55] | |||
| HCMV UL120/121 | ||||
| UL114 | [56] | |||
| miR-UL148D | IER5 | [57] | ||
| miR-US25-1-5p | Cyclin E2 | [58] | ||
| TRIM28 | [58] | |||
| EID1 | [58] | |||
| MAPRE2 | [58] | |||
| miR-US25-2-3p | eIF4A1 | [59] | ||
| miR-US33-5p | CCND1 | [60] | ||
| STX3 | [61] | |||
| Escape immune response | miR-UL112-3p | MICA; NK cells | [62] | |
| MICB; NK cells | [63] | |||
| IRF1; innate immune cells | [64] | |||
| miR-UL112-5p | ERAP1; CD8+ T cells | [65] | ||
| miR-US4-5p | ERAP1; CD8+ T cells | [47] | ||
| miR-UL59 | ULBP1; NK cells | [66] | ||
| miR-US5-1 | HCMV US7; multiple immune cells | [67] | ||
| miR-US5-2-3p | HCMV US7; multiple immune cells | [67] | ||
| miR-US33as-5p | IFNAR1; innate immune cells | [68] | ||
| Inhibit autophagy | miR-UL112-3p | ATG5; HFFs | [61] | |
| miR-US22-5p | ATG5; HFFs | [61] | ||
| miR-US29-5p | ATG5; HFFs | [61] | ||
| Inhibit apoptosis | miR-US4-5p | CASP2; HFFs | [61] | |
| miR-UL112-5p | CASP3; HFFs | [61] | ||
| miR-UL22A-5p | CASP3; HFFs | [61] | ||
| miR-US25-2-3p | CASP3; HFFs | [61] | ||
| miR-UL148D | IEX-1; HEK293 cells | [69] | ||
| PHAP1; HeLa cell S-100 | [70] | |||
| ERN1; HeLa cell S-100 | [70] | |||
| miR-UL22A-3p | CASP7; HFFs | [61] | ||
| miR-UL36-3p | FAS; HFFs | [61] | ||
| miR-US5-1 | FAS; HFFs | [61] | ||
| miR-US5-2-3p | FAS; HFFs | [61] | ||
| miR-UL36-5p | SLC25A6 (ANT3); HEK293 cells, U373 cells and HELF cells | [71] | ||
| miR-UL70-3p | MOAP1; HEK293T cells | [71] | ||
| miR-US4-5p | QARS; CD8+ T cells | [47] | ||
| miR-US22-5p | US22; human fibroblast cells | [72] | ||
| Reduce inflammatory cytokine production | miR-UL112-3p | IKKα/IKKβ; fibroblasts | [73] | |
| miR-US5-1 | IKKα/IKKβ; fibroblasts | [73] | ||
| miR-UL112-3p | IL-32; NK cells | [16] | ||
| TLR2; NK cells | [74] | |||
| miR-UL112-3p | Vamp3; NK cells | [75] | ||
| miR-US5-1 | Vamp3; NK cells | [75] | ||
| miR-US5-2-3p | Vamp3; NK cells | [75] | ||
| miR-UL112-3p | Rab5c; NK cells | [76] | ||
| miR-US5-1 | Rab5c; NK cells | [76] | ||
| miR-US5-2-3p | Rab5c; NK cells | [76] | ||
| miR-UL112-3p | Rab11a; NK cells | [75] | ||
| miR-US5-1 | Rab11a; NK cells | [75] | ||
| miR-US5-2-3p | Rab11a; NK cells | [75] | ||
| miR-UL112-3p | SNAP23; NK cells | [75] | ||
| miR-US5-1 | SNAP23; NK cells | [75] | ||
| miR-US5-2-3p | SNAP23; NK cells | [75] | ||
| miR-UL112-3p | CDC42; NK cells | [75] | ||
| miR-US5-1 | CDC42; NK cells | [75] | ||
| miR-US5-2-3p | CDC42; NK cells | [75] | ||
| miR-UL148D | ACVR1B; NK cells | [76] | ||
| RANTES; NK cells | [77] | |||
| miR-US25-1-5p | CD147; HEK293 cells | [78] | ||
| Suppress cell cycle progression | miR-UL36-3p | CDK6; HFFs | [61] | |
| miR-US5-1 | CDK6; HFFs | [61] | ||
| miR-US5-2-3p | CDK6; HFFs | [61] | ||
| miR-US25-1-3p | CDK6; HFFs | [61] | ||
| miR-US25-2-3p | CDK6; HFFs | [61] | ||
| Induce myelosuppression | miR-UL22A-3p | SMAD3; CD34+ HPCs | [79] | |
| miR-UL22A-5p | SMAD3; CD34+ HPCs | [79] | ||
| miR-US5-2-3p | NAB1; CD34+ HPCs | [79] | ||
| Reactivation | Promote viral gene expression | miR-UL112-3p | BclAF1 | [80] |
| miR-UL36-5p | HCMV UL138 | [81] | ||
| miR-US5-1 | Geminin | [82] | ||
| Induce cell differentiation | miR-US22-5p | EGR1; HEK293 cells, NHDF | [83] | |
| Promote apoptosis | miR-US4-5p | PAK2; HEK293, HELF and THP-1 cells | [84] | |
| miR-US25-1-5p | BRCC3; EAhy926 cells | [85] | ||
| Others | N | miR-UL22A-5p | BMPR2 | [86] |
| miR-US4-3p | CASP7 | [47] | ||
| CDK6 | [61] | |||
| ERAP1 | [47] | |||
| miR-US22-3p | US22 | [72] | ||
| miR-US33-3p | US29 | [87] | ||
| miR-UL69 | N | |||
| miR-UL70-5p | N | |||
| miR-US5-2-5p | N | |||
| miR-US25-2-5p | N | |||
| miR-US29-3p | N |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, M.; Jin, Y.; Zhang, S.; Xu, J.; Zhang, J. microRNA, a Subtle Indicator of Human Cytomegalovirus against Host Immune Cells. Vaccines 2022, 10, 144. https://doi.org/10.3390/vaccines10020144
Yu M, Jin Y, Zhang S, Xu J, Zhang J. microRNA, a Subtle Indicator of Human Cytomegalovirus against Host Immune Cells. Vaccines. 2022; 10(2):144. https://doi.org/10.3390/vaccines10020144
Chicago/Turabian StyleYu, Mengyao, Yuexinzi Jin, Shichang Zhang, Jian Xu, and Jiexin Zhang. 2022. "microRNA, a Subtle Indicator of Human Cytomegalovirus against Host Immune Cells" Vaccines 10, no. 2: 144. https://doi.org/10.3390/vaccines10020144
APA StyleYu, M., Jin, Y., Zhang, S., Xu, J., & Zhang, J. (2022). microRNA, a Subtle Indicator of Human Cytomegalovirus against Host Immune Cells. Vaccines, 10(2), 144. https://doi.org/10.3390/vaccines10020144