
Unbiased Identification of Dengue Virus Non-Structural Protein 1 Peptides for Use in Vaccine Design


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
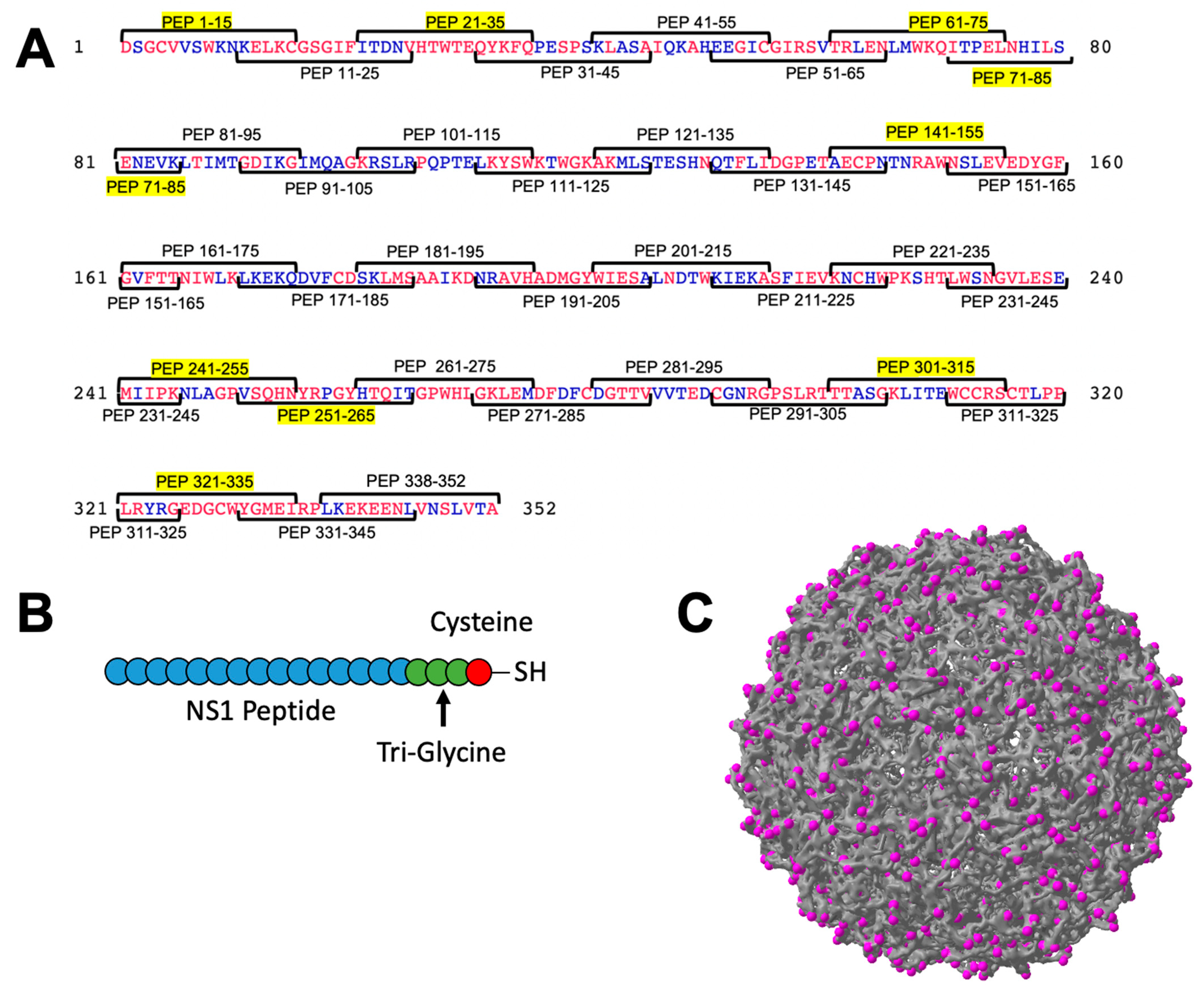
2.1. Synthesis of NS1 Peptides
2.2. Qβ VLP Vaccine Production
2.3. Enzyme-Linked Immunosorbent Assays (ELISAs)
2.4. Animals
2.5. Cells and Viral Stocks
2.6. Anti-NS1 Antiserum Binding to DENV-Infected HEK-293 Cells
3. Results
3.1. Strategy for Unbiased Identification of NS1 Peptide Eptiopes of Interest
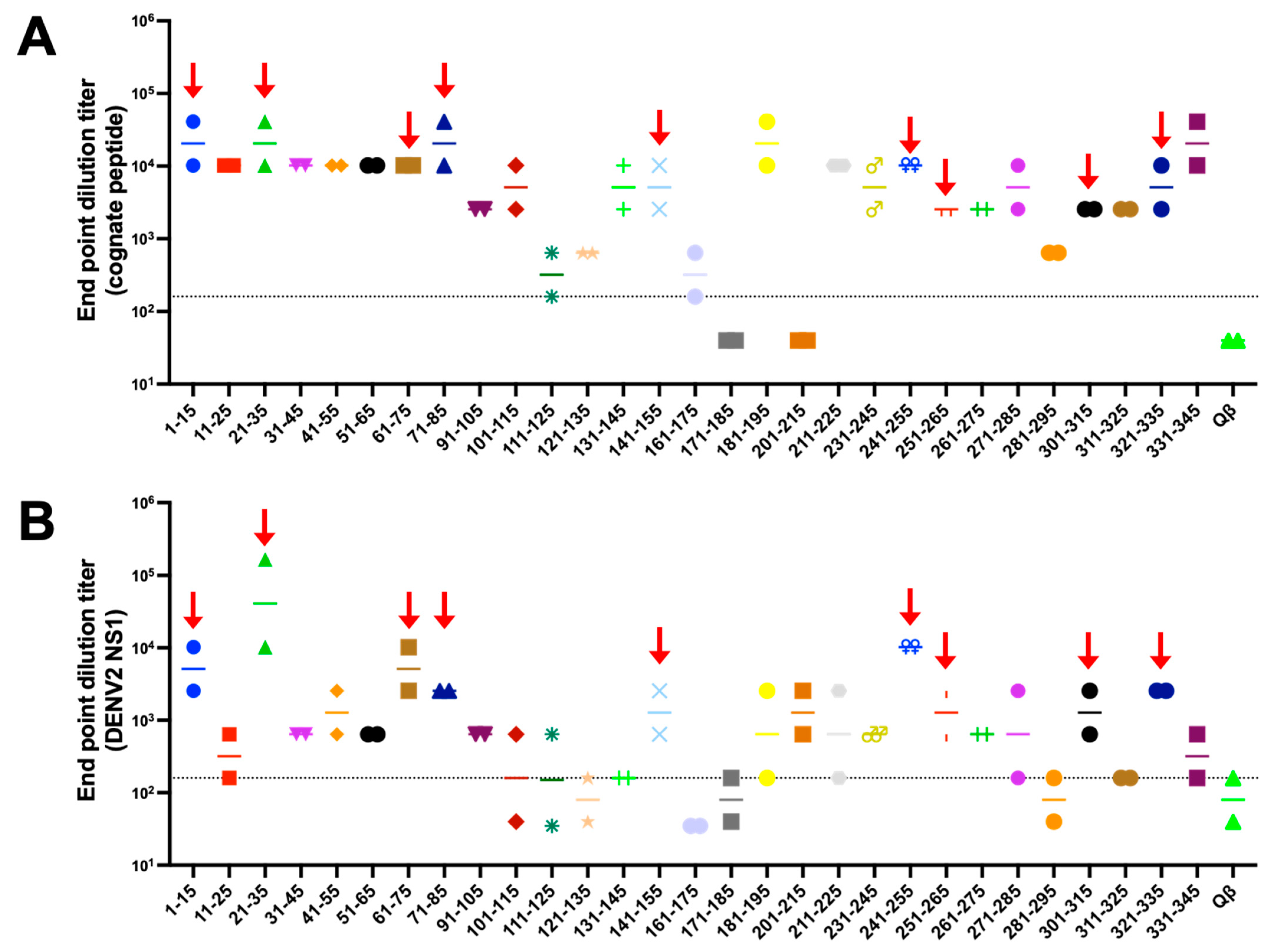
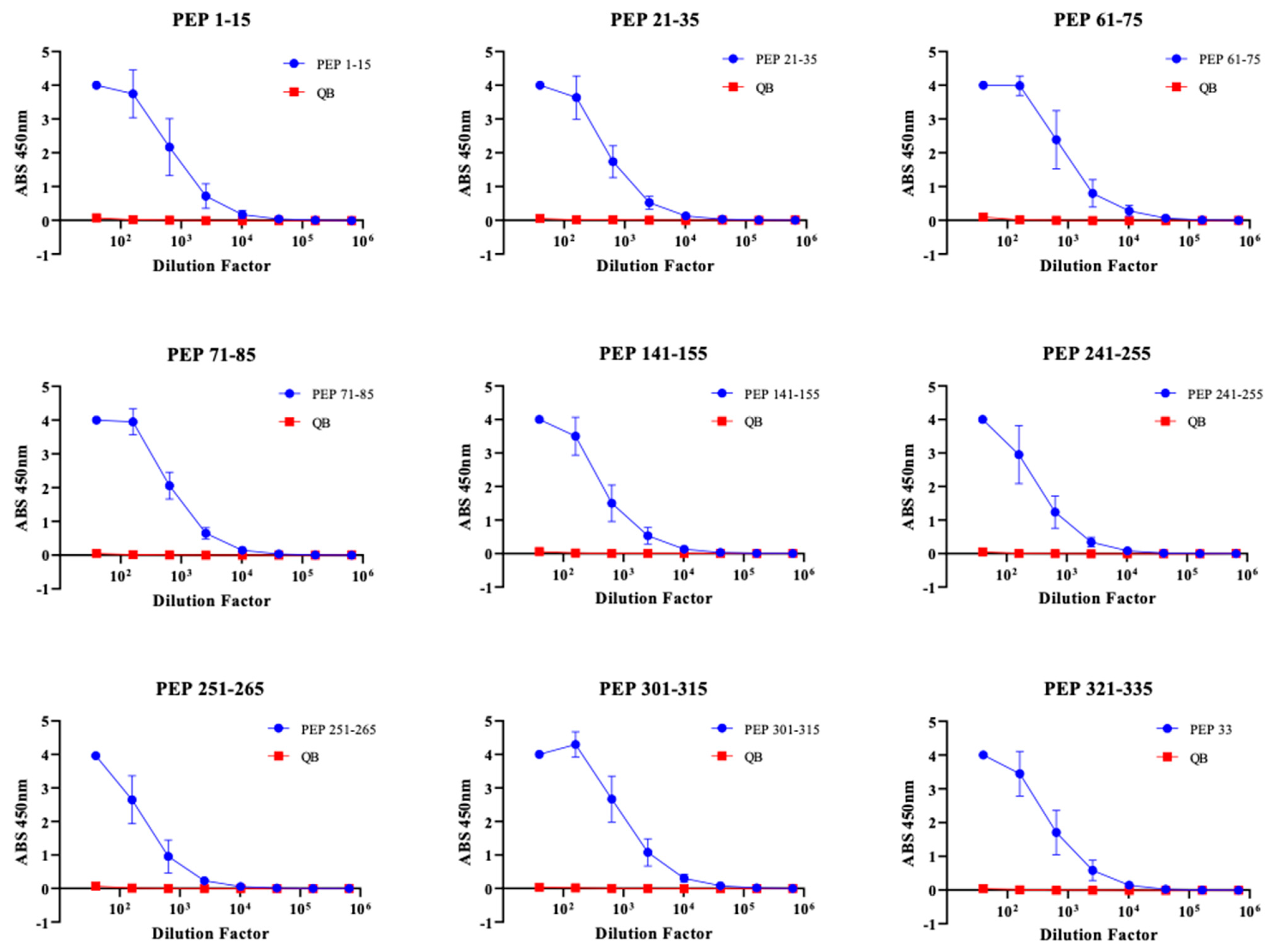
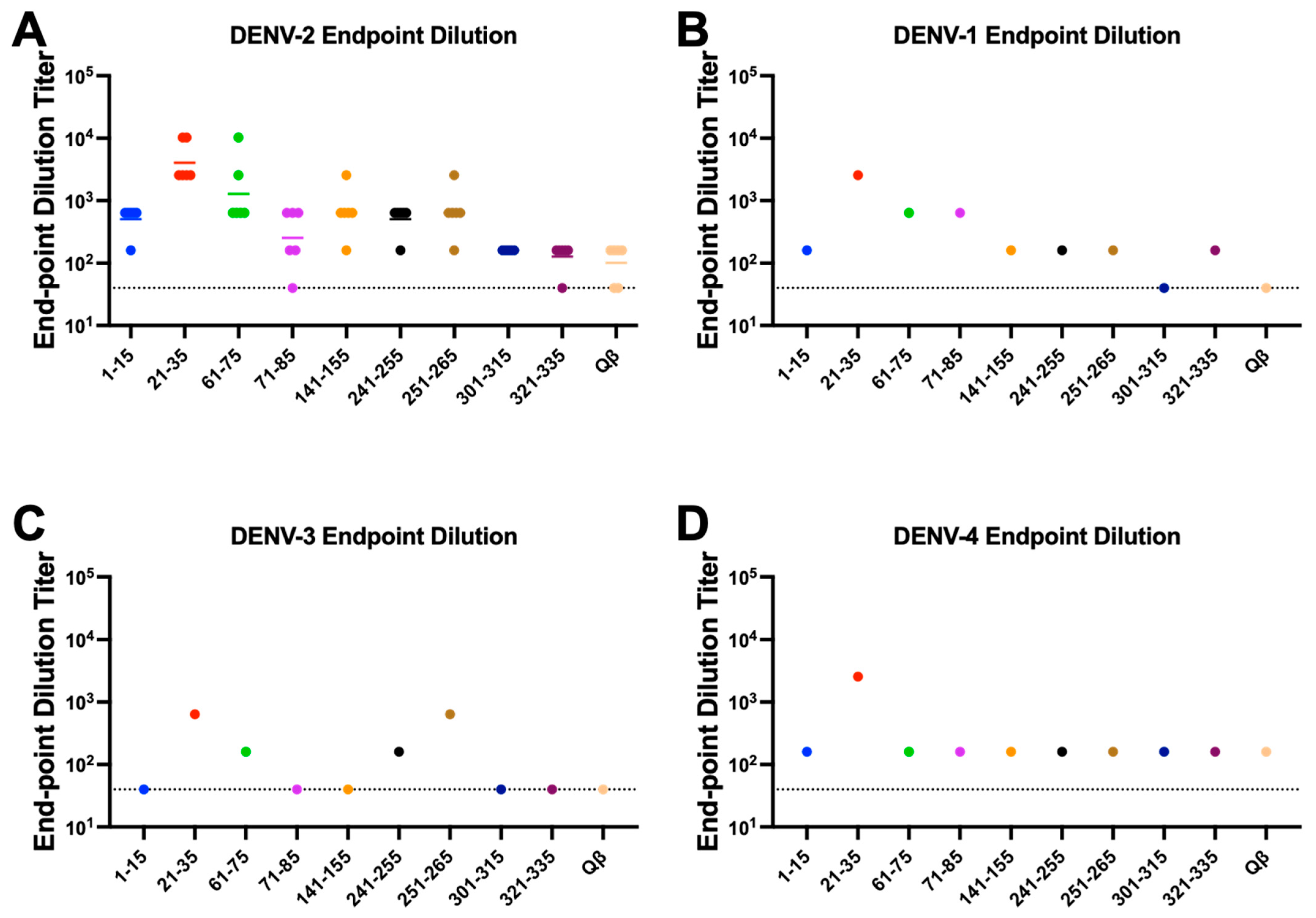
3.2. Immunogenicity of Qβ VLPs Displaying Unbiased NS1 Peptide Epitopes
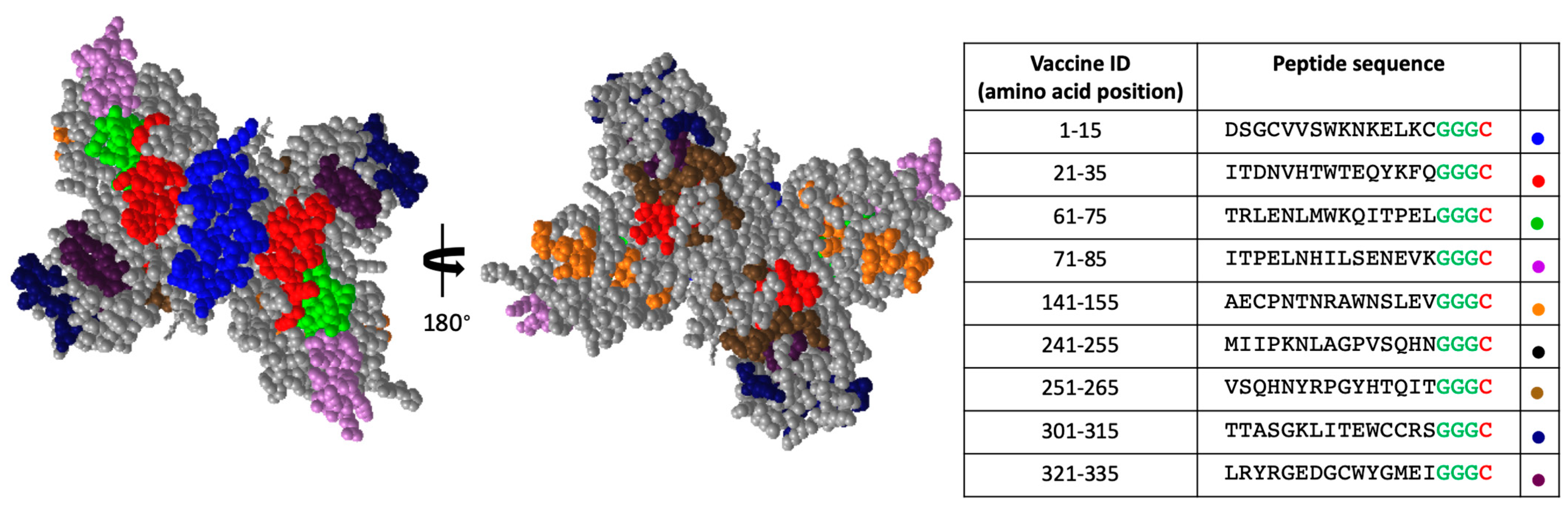
3.3. Structural Locations of Down-Selected NS1 Peptide Epitopes
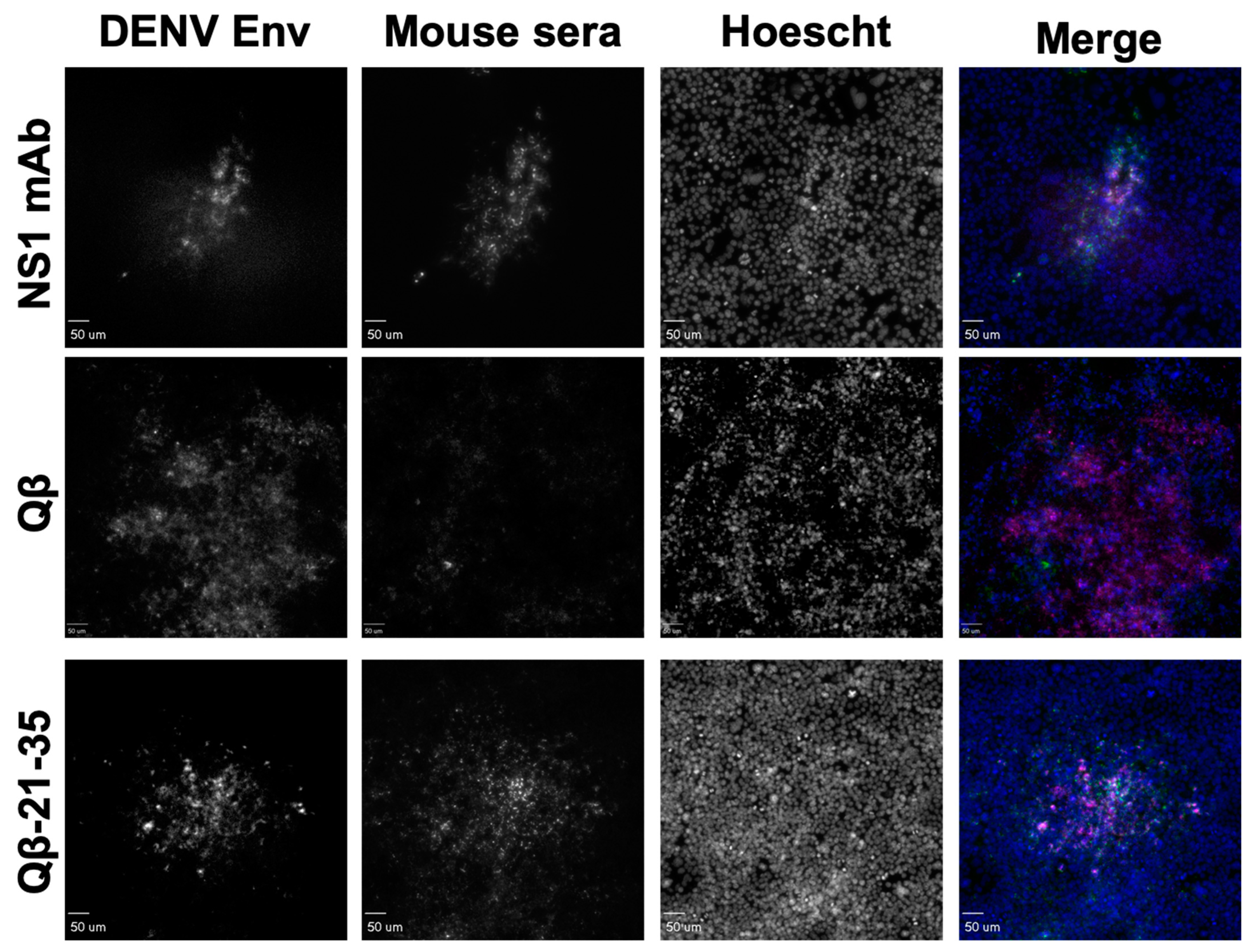
3.4. Binding Characteristics of IgG Elicited by Qβ VLPs Displaying NS1 Peptides Epitopes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Halstead, S.B. Dengue Antibody-Dependent Enhancement: Knowns and Unknowns. Microbiol. Spectr. 2014, 2, AID-0022-2014. [Google Scholar] [CrossRef] [PubMed]
- Malavige, G.N.; Ogg, G.S. Pathogenesis of Vascular Leak in Dengue Virus Infection. Immunology 2017, 151, 261–269. [Google Scholar] [CrossRef]
- Chen, H.R.; Lai, Y.C.; Yeh, T.M. Dengue Virus Non-Structural Protein 1: A Pathogenic Factor, Therapeutic Target, and Vaccine Candidate. J. Biomed. Sci. 2018, 25, 58. [Google Scholar] [CrossRef]
- Libraty, D.H.; Young, P.R.; Pickering, D.; Endy, T.P.; Kalayanarooj, S.; Green, S.; Vaughn, D.W.; Nisalak, A.; Ennis, F.A.; Rothman, A.L. High Circulating Levels of the Dengue Virus Nonstructural Protein NS1 Early in Dengue Illness Correlate with the Development of Dengue Hemorrhagic Fever. J. Infect. Dis. 2002, 186, 1165–1168. [Google Scholar] [CrossRef] [PubMed]
- Puerta-Guardo, H.; Glasner, D.R.; Harris, E. Dengue Virus NS1 Disrupts the Endothelial Glycocalyx, Leading to Hyperpermeability. PLoS Pathog. 2016, 12, e1005738. [Google Scholar] [CrossRef] [PubMed]
- Glasner, D.R.; Ratnasiri, K.; Puerta-Guardo, H.; Espinosa, D.A.; Beatty, P.R.; Harris, E. Dengue Virus NS1 Cytokine-Independent Vascular Leak Is Dependent on Endothelial Glycocalyx Components. PLoS Pathog. 2017, 13, e1006673. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.R.; Chao, C.H.; Liu, C.C.; Ho, T.S.; Tsai, H.P.; Perng, G.C.; Lin, Y.S.; Wang, J.R.; Yeh, T.M. Macrophage Migration Inhibitory Factor Is Critical for Dengue NS1-Induced Endothelial Glycocalyx Degradation and Hyperpermeability. PLoS Pathog. 2018, 14, e1007033. [Google Scholar] [CrossRef]
- Modhiran, N.; Watterson, D.; Muller, D.A.; Panetta, A.K.; Sester, D.P.; Liu, L.; Hume, D.A.; Stacey, K.J.; Young, P.R. Dengue Virus NS1 Protein Activates Cells via Toll-like Receptor 4 and Disrupts Endothelial Cell Monolayer Integrity. Sci. Transl. Med. 2015, 7, 304ra142. [Google Scholar] [CrossRef]
- Modhiran, N.; Watterson, D.; Blumenthal, A.; Baxter, A.G.; Young, P.R.; Stacey, K.J. Dengue Virus NS1 Protein Activates Immune Cells via TLR4 but Not TLR2 or TLR6. Immunol. Cell Biol. 2017, 95, 491–495. [Google Scholar] [CrossRef]
- Chen, J.; Ng, M.M.; Chu, J.J. Activation of TLR2 and TLR6 by Dengue NS1 Protein and Its Implications in the Immunopathogenesis of Dengue Virus Infection. PLoS Pathog. 2015, 11, e1005053. [Google Scholar] [CrossRef]
- Beatty, P.R.; Puerta-Guardo, H.; Killingbeck, S.S.; Glasner, D.R.; Hopkins, K.; Harris, E. Dengue Virus NS1 Triggers Endothelial Permeability and Vascular Leak That Is Prevented by NS1 Vaccination. Sci. Transl. Med. 2015, 7, 304ra141. [Google Scholar] [CrossRef]
- Adikari, T.N.; Gomes, L.; Wickramasinghe, N.; Salimi, M.; Wijesiriwardana, N.; Kamaladasa, A.; Shyamali, N.L.; Ogg, G.S.; Malavige, G.N. Dengue NS1 Antigen Contributes to Disease Severity by Inducing Interleukin (IL)-10 by Monocytes. Clin. Exp. Immunol. 2016, 184, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, J.J.; Brandriss, M.W.; Walsh, E.E. Protection of Mice against Dengue 2 Virus Encephalitis by Immunization with the Dengue 2 Virus Non-Structural Glycoprotein NS1. J. Gen. Virol. 1987, 68 Pt 3, 853–857. [Google Scholar] [CrossRef]
- Henchal, E.A.; Henchal, L.S.; Schlesinger, J.J. Synergistic Interactions of Anti-NS1 Monoclonal Antibodies Protect Passively Immunized Mice from Lethal Challenge with Dengue 2 Virus. J. Gen. Virol. 1988, 69 Pt 8, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.C.; Chuang, Y.C.; Liu, C.C.; Ho, T.S.; Lin, Y.S.; Anderson, R.; Yeh, T.M. Antibodies Against Modified NS1 Wing Domain Peptide Protect Against Dengue Virus Infection. Sci. Rep. 2017, 7, 6975. [Google Scholar] [CrossRef]
- Wan, S.W.; Lu, Y.T.; Huang, C.H.; Lin, C.F.; Anderson, R.; Liu, H.S.; Yeh, T.M.; Yen, Y.T.; Wu-Hsieh, B.A.; Lin, Y.S. Protection against Dengue Virus Infection in Mice by Administration of Antibodies against Modified Nonstructural Protein 1. PLoS ONE 2014, 9, e92495. [Google Scholar] [CrossRef]
- Falconar, A.K. Antibody Responses Are Generated to Immunodominant ELK/KLE-Type Motifs on the Nonstructural-1 Glycoprotein during Live Dengue Virus Infections in Mice and Humans: Implications for Diagnosis, Pathogenesis, and Vaccine Design. Clin. Vaccine Immunol. 2007, 14, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Jayathilaka, D.; Gomes, L.; Jeewandara, C.; Jayarathna, G.S.B.; Herath, D.; Perera, P.A.; Fernando, S.; Wijewickrama, A.; Hardman, C.S.; Ogg, G.S.; et al. Role of NS1 Antibodies in the Pathogenesis of Acute Secondary Dengue Infection. Nat. Commun. 2018, 9, 5242. [Google Scholar] [CrossRef] [PubMed]
- Lebeau, G.; Lagrave, A.; Ogire, E.; Grondin, L.; Seriacaroupin, S.; Moutoussamy, C.; Mavingui, P.; Hoarau, J.-J.; Roche, M.; Krejbich-Trotot, P.; et al. Viral Toxin NS1 Implication in Dengue Pathogenesis Making It a Pivotal Target in Development of Efficient Vaccine. Vaccines 2021, 9, 946. [Google Scholar] [CrossRef]
- Falconar, A.K. The Dengue Virus Nonstructural-1 Protein (NS1) Generates Antibodies to Common Epitopes on Human Blood Clotting, Integrin/Adhesin Proteins and Binds to Human Endothelial Cells: Potential Implications in Haemorrhagic Fever Pathogenesis. Arch. Virol. 1997, 142, 897–916. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Yeh, T.M.; Lin, C.F.; Wan, S.W.; Chuang, Y.C.; Hsu, T.K.; Liu, H.S.; Liu, C.C.; Anderson, R.; Lei, H.Y. Molecular Mimicry between Virus and Host and Its Implications for Dengue Disease Pathogenesis. Exp. Biol. Med. 2011, 236, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.S.; King, C.C.; Huang, H.S.; Shih, Y.L.; Lee, C.C.; Tsai, W.J.; Yu, C.C.; Chang, H.H. Antiplatelet Autoantibodies Elicited by Dengue Virus Non-Structural Protein 1 Cause Thrombocytopenia and Mortality in Mice. J. Thromb. Haemost. 2007, 5, 2291–2299. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.J.; Lei, H.Y.; Lin, C.F.; Luo, Y.H.; Wan, S.W.; Liu, H.S.; Yeh, T.M.; Lin, Y.S. Anti-Dengue Virus Nonstructural Protein 1 Antibodies Recognize Protein Disulfide Isomerase on Platelets and Inhibit Platelet Aggregation. Mol. Immunol. 2009, 47, 398–406. [Google Scholar] [CrossRef]
- Chen, M.C.; Lin, C.F.; Lei, H.Y.; Lin, S.C.; Liu, H.S.; Yeh, T.M.; Anderson, R.; Lin, Y.S. Deletion of the C-Terminal Region of Dengue Virus Nonstructural Protein 1 (NS1) Abolishes Anti-NS1-Mediated Platelet Dysfunction and Bleeding Tendency. J. Immunol. 2009, 183, 1797–1803. [Google Scholar] [CrossRef]
- Liu, I.J.; Chiu, C.Y.; Chen, Y.C.; Wu, H.C. Molecular Mimicry of Human Endothelial Cell Antigen by Autoantibodies to Nonstructural Protein 1 of Dengue Virus. J. Biol. Chem. 2011, 286, 9726–9736. [Google Scholar] [CrossRef]
- Cheng, H.J.; Lin, C.F.; Lei, H.Y.; Liu, H.S.; Yeh, T.M.; Luo, Y.H.; Lin, Y.S. Proteomic Analysis of Endothelial Cell Autoantigens Recognized by Anti-Dengue Virus Nonstructural Protein 1 Antibodies. Exp. Biol. Med. 2009, 234, 63–73. [Google Scholar] [CrossRef]
- Akey, D.L.; Brown, W.C.; Dutta, S.; Konwerski, J.; Jose, J.; Jurkiw, T.J.; DelProposto, J.; Ogata, C.M.; Skiniotis, G.; Kuhn, R.J.; et al. Flavivirus NS1 Structures Reveal Surfaces for Associations with Membranes and the Immune System. Science 2014, 343, 881–885. [Google Scholar] [CrossRef]
- Hertz, T.; Beatty, P.R.; MacMillen, Z.; Killingbeck, S.S.; Wang, C.; Harris, E. Antibody Epitopes Identified in Critical Regions of Dengue Virus Nonstructural 1 Protein in Mouse Vaccination and Natural Human Infections. J. Immunol. 2017, 198, 4025–4035. [Google Scholar] [CrossRef]
- Warner, N.L.; Frietze, K.M. Development of Bacteriophage Virus-Like Particle Vaccines Displaying Conserved Epitopes of Dengue Virus Non-Structural Protein 1. Vaccines 2021, 9, 726. [Google Scholar] [CrossRef]
- Glasner, D.R.; Puerta-Guardo, H.; Beatty, P.R.; Harris, E. The Good, the Bad, and the Shocking: The Multiple Roles of Dengue Virus Nonstructural Protein 1 in Protection and Pathogenesis. Annu. Rev. Virol. 2018, 5, 227–253. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.A.; Landsberg, M.J.; Bletchly, C.; Rothnagel, R.; Waddington, L.; Hankamer, B.; Young, P.R. Structure of the Dengue Virus Glycoprotein Non-Structural Protein 1 by Electron Microscopy and Single-Particle Analysis. J. Gen. Virol. 2012, 93, 771–779. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warner, N.L.; Core, S.B.; Frietze, K.M. Unbiased Identification of Dengue Virus Non-Structural Protein 1 Peptides for Use in Vaccine Design. Vaccines 2022, 10, 2028. https://doi.org/10.3390/vaccines10122028
Warner NL, Core SB, Frietze KM. Unbiased Identification of Dengue Virus Non-Structural Protein 1 Peptides for Use in Vaccine Design. Vaccines. 2022; 10(12):2028. https://doi.org/10.3390/vaccines10122028
Chicago/Turabian StyleWarner, Nikole L., Susan B. Core, and Kathryn M. Frietze. 2022. "Unbiased Identification of Dengue Virus Non-Structural Protein 1 Peptides for Use in Vaccine Design" Vaccines 10, no. 12: 2028. https://doi.org/10.3390/vaccines10122028
APA StyleWarner, N. L., Core, S. B., & Frietze, K. M. (2022). Unbiased Identification of Dengue Virus Non-Structural Protein 1 Peptides for Use in Vaccine Design. Vaccines, 10(12), 2028. https://doi.org/10.3390/vaccines10122028