
Subtype H3N2 Influenza A Viruses: An Unmet Challenge in the Western Pacific


Abstract
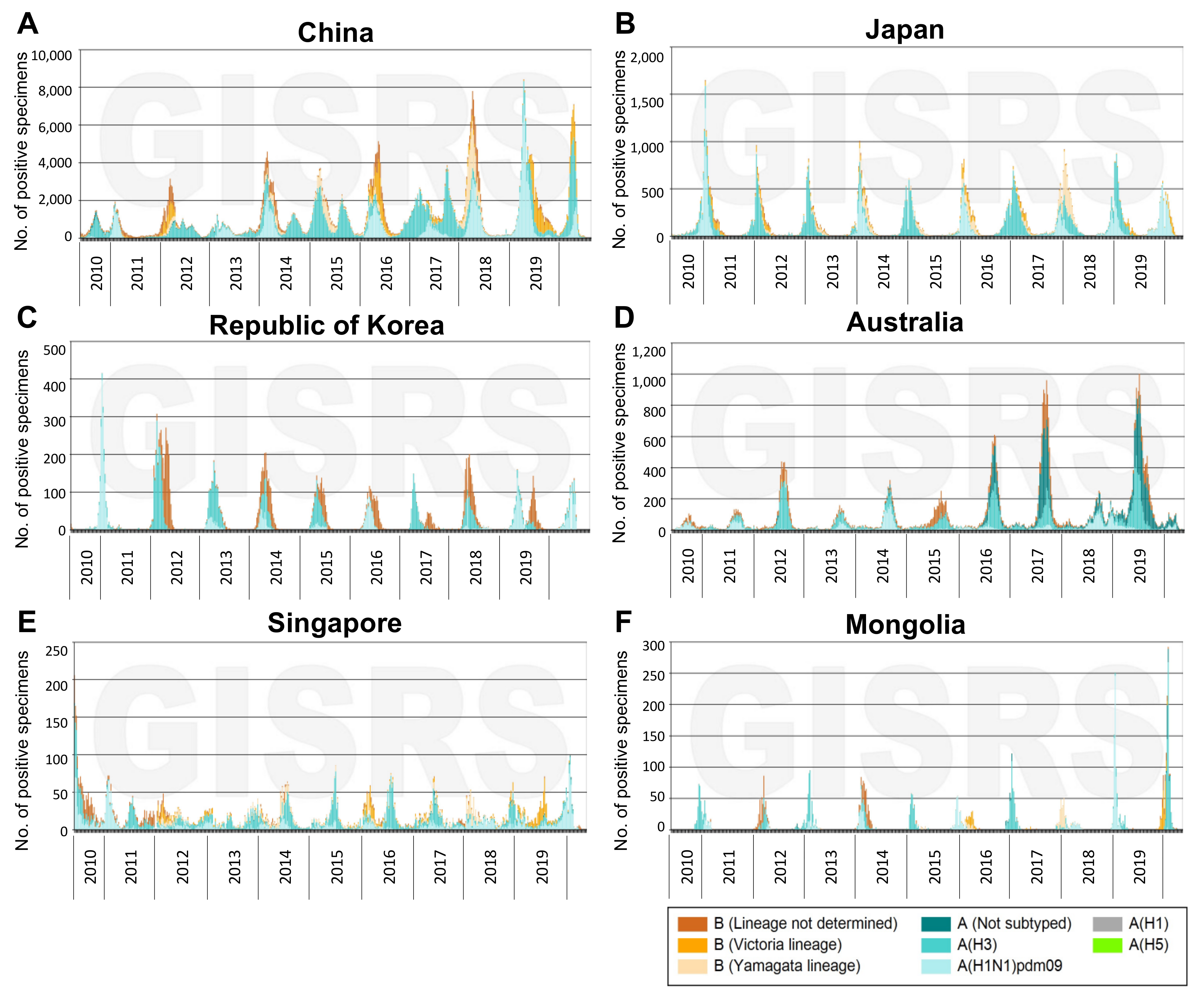
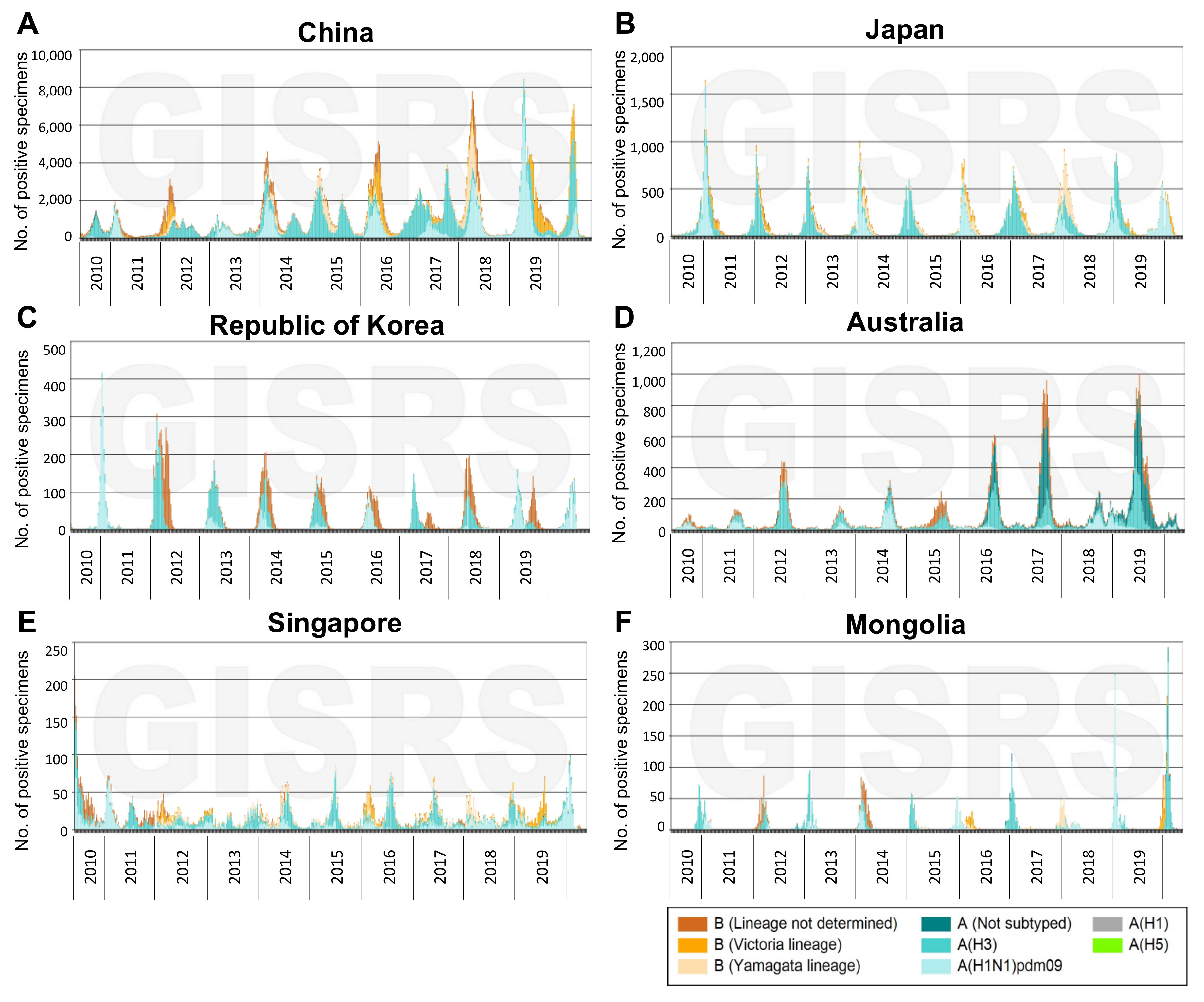
:1. Disease Burden of Influenza A(H3N2) Viruses in the Western Pacific
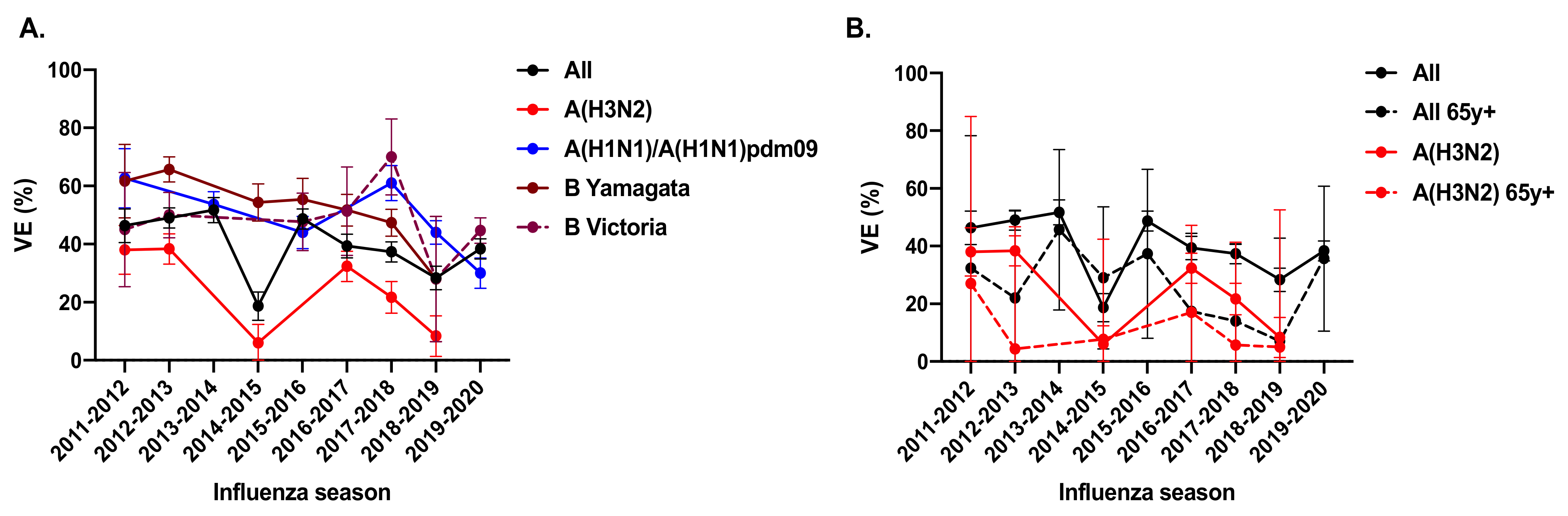
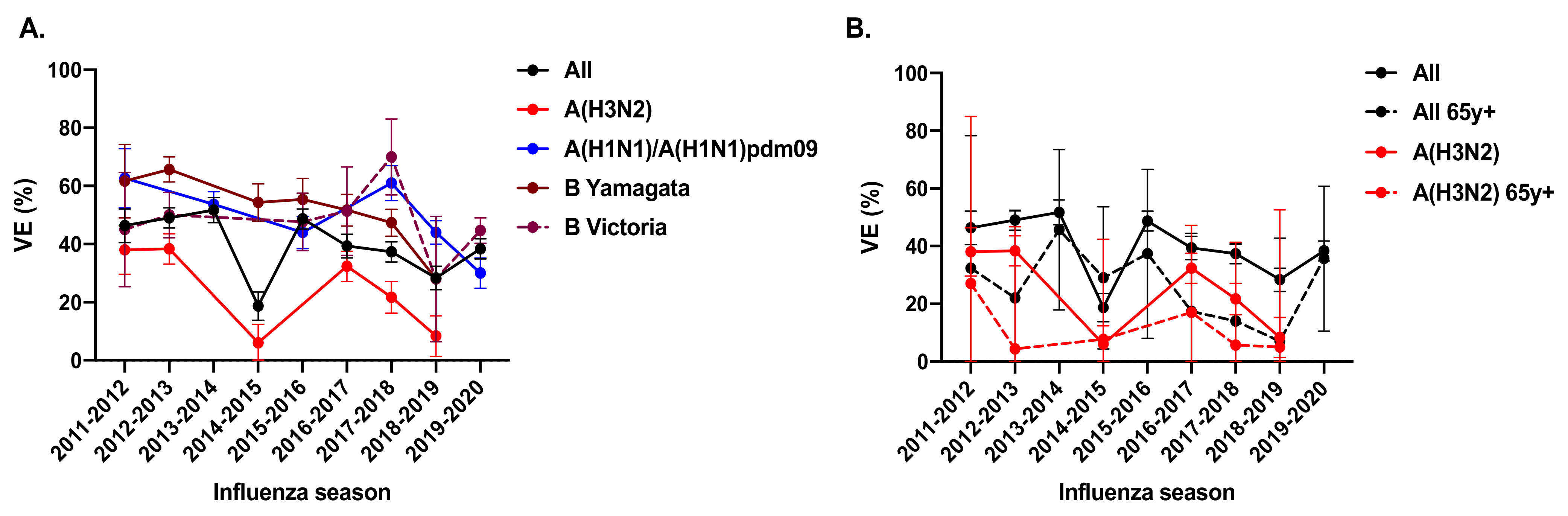
2. Unsatisfactory Vaccine Effectiveness of A(H3N2) Influenza Viruses
3. Reasons for Low VE against A(H3N2) Influenza
3.1. Antigenic Drift of A(H3N2) Viruses
3.2. Egg-Adaptation of A(H3N2) Viruses
3.2.1. Molecular Basis of A(H3N2) Egg Adaptation
3.2.2. Consequence of Egg Adaptation on Immunogenicity and VE
3.2.3. Influence of Pre-Existing Immunity to Influenza Vaccine Responses
4. Vaccines beyond Egg-Based Formula for Better Protection against Influenza
4.1. Cell-Based Vaccine
4.2. Recombinant HA (rHA) Vaccine
4.3. Other Technologies for Non-Egg-Based Influenza Vaccines
5. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Paget, J.; Spreeuwenberg, P.; Charu, V.; Taylor, R.J.; Iuliano, A.D.; Bresee, J.; Simonsen, L.; Viboud, C.; Global Seasonal Influenza-Associated Mortality Collaborator Network and GLaMOR Collaborating Teams. Global mortality associated with seasonal influenza epidemics: New burden estimates and predictors from the GLaMOR Project. J. Glob. Health 2019, 9, 020421. [Google Scholar] [CrossRef] [PubMed]
- Young, B.E.; Chen, M. Influenza in temperate and tropical Asia: A review of epidemiology and vaccinology. Hum. Vaccines Immunother. 2020, 16, 1659–1667. [Google Scholar] [CrossRef] [Green Version]
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects 2019, Online Edition. Rev. 1. 2019. Available online: https://population.un.org/wpp/Download/Standard/Population/ (accessed on 12 November 2021).
- Vestergaard, L.S.; Nielsen, J.; Krause, T.G.; Espenhain, L.; Tersago, K.; Sierra, N.B.; Denissov, G.; Innos, K.; Virtanen, M.J.; Fouillet, A.; et al. Excess all-cause and influenza-attributable mortality in Europe, December 2016 to February 2017. Eurosurveillance 2017, 22, 30506. [Google Scholar] [CrossRef]
- Feng, L.; Feng, S.; Chen, T.; Yang, J.; Lau, Y.C.; Peng, Z.; Li, L.; Wang, X.; Wong, J.Y.T.; Qin, Y.; et al. Burden of influenza-associated outpatient influenza-like illness consultations in China, 2006–2015: A population-based study. Influenza Other Respir. Viruses 2020, 14, 162–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Huang, J.; Huai, Y.; Guan, X.; Klena, J.; Liu, S.; Peng, Y.; Yang, H.; Luo, J.; Zheng, J.; et al. The substantial hospitalization burden of influenza in central China: Surveillance for severe, acute respiratory infection, and influenza viruses, 2010–2012. Influenza Other Respir. Viruses 2014, 8, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Kwok, K.O.; Riley, S.; Perera, R.A.P.M.; Wei, V.W.I.; Wu, P.; Wei, L.; Chu, D.K.W.; Barr, I.G.; Peiris, J.S.M.; Cowling, B.J. Relative incidence and individual-level severity of seasonal influenza A H3N2 compared with 2009 pandemic H1N1. BMC Infect. Dis. 2017, 17, 337. [Google Scholar] [CrossRef]
- Wu, P.; Goldstein, E.; Ho, L.M.; Yang, L.; Nishiura, H.; Wu, J.T.K.; Ip, D.K.M.; Chuang, S.-K.; Tsang, T.; Cowling, B.J. Excess mortality associated with influenza A and B virus in Hong Kong, 1998–2009. J. Infect. Dis. 2012, 206, 1862–1871. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.Y.; Goldstein, E.; Fang, V.J.; Cowling, B.J.; Wu, P. Real-time estimation of the influenza-associated excess mortality in Hong Kong. Epidemiol. Infect. 2019, 147, e217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Liu, Y.; Wu, P.; Peng, Z.; Wang, X.; Chen, T.; Wong, J.Y.T.; Yang, J.; Bond, H.S.; Wang, L.; et al. Influenza-associated excess respiratory mortality in China, 2010–2015: A population-based study. Lancet Public Health 2019, 4, e473–e481. [Google Scholar] [CrossRef] [Green Version]
- Chow, A.; Ma, S.; Ling, A.E.; Chew, S.K. Influenza-associated deaths in tropical Singapore. Emerg. Infect. Dis. 2006, 12, 114–121. [Google Scholar] [CrossRef]
- Rajaram, S.; Wojcik, R.; Moore, C.; de Lejarazu, R.O.; de Lusignan, S.; Montomoli, E.; Rossi, A.; Perez-Rubio, A.; Trilla, A.; Baldo, V.; et al. The impact of candidate influenza virus and egg-based manufacture on vaccine effectiveness: Literature review and expert consensus. Vaccine 2020, 38, 6047–6056. [Google Scholar] [CrossRef]
- Belongia, E.A.; Simpson, M.D.; King, J.P.; Sundaram, M.E.; Kelley, N.S.; Osterholm, M.T.; McLean, H.Q. Variable influenza vaccine effectiveness by subtype: A systematic review and meta-analysis of test-negative design studies. Lancet Infect. Dis. 2016, 16, 942–951. [Google Scholar] [CrossRef]
- Flannery, B.; Chung, J.R.; Monto, A.S.; Martin, E.T.; Belongia, E.A.; McLean, H.Q.; Gaglani, M.; Murthy, K.; Zimmerman, R.; Nowalk, M.P.; et al. Influenza vaccine effectiveness in the United States during the 2016–2017 season. Clin. Infect. Dis. 2019, 68, 1798–1806. [Google Scholar] [CrossRef]
- Rolfes, M.A.; Flannery, B.; Chung, J.R.; O’Halloran, A.; Garg, S.; Belongia, E.A.; Gaglani, M.; Zimmerman, R.K.; Jackson, M.L.; Monto, A.S.; et al. Effects of influenza vaccination in the United States during the 2017–2018 influenza season. Clin. Infect. Dis. 2019, 69, 1845–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, J.R.; Rolfes, M.A.; Flannery, B.; Prasad, P.; O’Halloran, A.; Garg, S.; Fry, A.M.; Singleton, J.A.; Patel, M.; Reed, C.; et al. Effects of influenza vaccination in the United States during the 2018–2019 influenza season. Clin. Infect. Dis. 2020, 71, e368–e376. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, Y.; Hackert, V.; Van Der Hoek, W.; Meijer, A.; Krafft, T.; Yang, P.; Wang, Q. The 2015–2016 influenza epidemic in Beijing, China: Unlike elsewhere, circulation of influenza A(H3N2) with moderate vaccine effectiveness. Vaccine 2018, 36, 4993–5001. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, D.M.; Sabaiduc, S.; Leir, S.; Rose, C.; Zou, M.; Murti, M.; Dickinson, J.A.; Olsha, R.; Gubbay, J.B.; Crozen, M.A.; et al. Paradoxical clade- and age-specific vaccine effectiveness during the 2018/19 influenza A(H3N2) epidemic in Canada: Potential imprint-regulated effect of vaccine (I-REV). Euro Surveill. 2019, 24, 1900585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.; Thompson, M.G.; Ma, C.; Shi, W.; Wu, S.; Zhang, D.; Wang, Q. Influenza vaccine effectiveness against medically-attended influenza illness during the 2012–2013 season in Beijing, China. Vaccine 2014, 32, 5285–5289. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, Y.; Ma, C.; Duan, W.; Sun, Y.; Wu, S.; Zhang, M.; Tian, Y.; Zheng, Y.; Yang, P.; et al. Moderate influenza vaccine effectiveness against influenza A(H1N1)pdm09 virus and low effectiveness against A(H3N2) virus among older adults during 2013-2014 influenza season in Beijing, China. Hum. Vaccines Immunother. 2018, 14, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zhang, Y.; Wu, P.; Feng, S.; Zheng, J.; Yang, P.; Pan, Y.; Wang, Q.; Feng, L.; Pang, X.; et al. Influenza vaccine effectiveness in preventing hospitalization among Beijing residents in China, 2013–2015. Vaccine 2016, 34, 2329–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Pan, Y.; Zhang, L.; Zhang, Y.; Wu, S.; Sun, Y.; Duan, W.; Zhang, M.; Wang, Q.; Yang, P. Influenza vaccine effectiveness against medically attended influenza illness in Beijing, China, 2014/15 season. Hum. Vaccines Immunother. 2017, 13, 2379–2384. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, P.; Feng, L.; Yang, P.; Feng, S.; Qin, Y.; Zheng, J.; Puig-Barbera, J.; Muscatello, D.; MacIntyre, R.; et al. Influenza vaccine effectiveness against influenza-associated hospitalization in 2015/16 season, Beijing, China. Vaccine 2017, 35, 3129–3134. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, L.; Yu, J.; Pang, Y.; Zhang, J.; Zhang, T.; Zhao, G. The effectiveness of influenza vaccination among nursery school children in China during the 2016/17 influenza season. Vaccine 2018, 36, 2456–2461. [Google Scholar] [CrossRef]
- Wu, S.; Pan, Y.; Zhang, X.; Zhang, L.; Duan, W.; Ma, C.; Zhang, Y.; Zhang, M.; Sun, Y.; Yang, P.; et al. Influenza vaccine effectiveness in preventing laboratory-confirmed influenza in outpatient settings: A test-negative case-control study in Beijing, China, 2016/17 season. Vaccine 2018, 36, 5774–5780. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; van der Hoek, W.; Krafft, T.; Pilot, E.; van Asten, L.; Lin, G.; Wu, S.; Duan, W.; Yang, P.; Wang, Q. Influenza vaccine effectiveness estimates against influenza A(H3N2) and A(H1N1) pdm09 among children during school-based outbreaks in the 2016–2017 season in Beijing, China. Hum. Vaccines Immunother. 2020, 16, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, Y.; Wang, Q.; Lock, J.; Pan, Y.; Cui, S.; Yang, P.; Hu, Y. The effectiveness of influenza vaccination in preventing hospitalizations in elderly in Beijing, 2016–2018. Vaccine 2019, 37, 1853–1858. [Google Scholar] [CrossRef]
- Cowling, B.J.; Chan, K.-H.; Feng, S.; Chan, E.L.; Lo, J.Y.; Peiris, J.M.; Chiu, S.S. The effectiveness of influenza vaccination in preventing hospitalizations in children in Hong Kong, 2009–2013. Vaccine 2014, 32, 5278–5284. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.-W.D.; Wong, M.-L.; Au, K.-W.; Chuang, S.-K. Seasonal influenza vaccine effectiveness at primary care level, Hong Kong SAR, 2017/2018 winter. Hum. Vaccines Immunother. 2019, 15, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Chiu, S.S.; Chua, H.; Kwan, M.Y.; Chan, E.L.; Wong, J.S.; Peiris, J.M.; Cowling, B.J. Influenza vaccination effectiveness in preventing influenza hospitalization in children, Hong Kong, winter 2019/20. Vaccine 2020, 38, 8078–8081. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Ikeda, S.; Hagiwara, Y.; Tsuzuki, D.; Klai, M.; Sakai, Y.; Crawford, B.; Nealon, J. Epidemiology and burden of illness of seasonal influenza among the elderly in Japan: A systematic literature review and vaccine effectiveness meta-analysis. Influenza Other Respir. Viruses 2021, 15, 293–314. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.Y.; Lim, S.; Song, J.Y.; Choi, W.S.; Jeong, H.W.; Heo, J.Y.; Lee, J.; Bin Seo, Y.; Lee, J.-S.; Wie, S.H.; et al. Interim estimates of the effectiveness of the influenza vaccine against A(H3N2) influenza in adults in South Korea, 2016–2017 season. PLoS ONE 2017, 12, e0178010. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Sullivan, S.G.; Tempone, S.S.; Wong, K.; Regan, A.; Dowse, G.K.; Effler, P.V.; Smith, D.W. Influenza vaccine effectiveness estimates for Western Australia during a period of vaccine and virus strain stability, 2010 to 2012. Vaccine 2014, 32, 6312–6318. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.G.; Carville, K.S.; Chilver, M.; Fielding, J.E.; Grant, K.A.; Kelly, H.; Levy, A.; Stocks, N.P.; Tempone, S.S.; Regan, A.K. Pooled influenza vaccine effectiveness estimates for Australia, 2012–2014. Epidemiol. Infect. 2016, 144, 2317–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fielding, J.E.; Levy, A.; Chilver, M.; Deng, Y.-M.; Regan, A.; Grant, K.A.; Stocks, N.; Sullivan, S. Effectiveness of seasonal influenza vaccine in Australia, 2015: An epidemiological, antigenic and phylogenetic assessment. Vaccine 2016, 34, 4905–4912. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, S.G.; Chilver, M.; Carville, K.S.; Deng, Y.-M.; Grant, K.A.; Higgins, G.; Komadina, N.; Leung, V.K.; Minney-Smith, C.A.; Teng, D.; et al. Low interim influenza vaccine effectiveness, Australia, 1 May to 24 September 2017. Eurosurveillance 2017, 22, 17–00707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, H.P.; Zhao, X.; Pang, J.; Chen, M.I.; Lee, V.J.; Ang, L.W.; Lin, R.V.; Gao, C.Q.; Hsu, L.Y.; Cook, A.R. Effectiveness of seasonal influenza vaccinations against laboratory-confirmed influenza-associated infections among Singapore military personnel in 2010–2013. Influenza Other Respir. Viruses 2014, 8, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Ng, Y.; Nandar, K.; Chua, L.A.V.; Mak, T.M.; Foo, K.; Muhammad, I.R.; Low, C.K.K.; Ma, S.; Ooi, S.P.; Lin, R.T.P.; et al. Evaluating the effectiveness of the influenza vaccine during respiratory outbreaks in Singapore’s long term care facilities, 2017. Vaccine 2019, 37, 3925–3931. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.G.; Pierse, N.; Huang, Q.S.; Prasad, N.; Duque, J.; Newbern, E.C.; Baker, M.G.; Turner, N.; McArthur, C. Influenza vaccine effectiveness in preventing influenza-associated intensive care admissions and attenuating severe disease among adults in New Zealand 2012–2015. Vaccine 2018, 36, 5916–5925. [Google Scholar] [CrossRef]
- Wong, S.-S.; Webby, R.J. Traditional and new influenza vaccines. Clin. Microbiol. Rev. 2013, 26, 476–492. [Google Scholar] [CrossRef] [Green Version]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Primers 2018, 4, 3. [Google Scholar] [CrossRef]
- Neher, R.; Bedford, T.; Daniels, R.S.; Russell, C.A.; Shraiman, B.I. Prediction, dynamics, and visualization of antigenic phenotypes of seasonal influenza viruses. Proc. Natl. Acad. Sci. USA 2016, 113, E1701–E1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, D.D.; Stewart, S.M.; Lee, J.; Ferdman, J.; Bajic, G.; Do, K.T.; Ernandes, M.J.; Suphaphiphat, P.; Settembre, E.C.; Dormitzer, P.R.; et al. Influenza immunization elicits antibodies specific for an egg-adapted vaccine strain. Nat. Med. 2016, 22, 1465–1469. [Google Scholar] [CrossRef] [PubMed]
- Parker, L.; Wharton, S.A.; Martin, S.R.; Cross, K.; Lin, Y.; Liu, Y.; Feizi, T.; Daniels, R.S.; McCauley, J.W. Effects of egg-adaptation on receptor-binding and antigenic properties of recent influenza A (H3N2) vaccine viruses. J. Gen. Virol. 2016, 97, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Harding, A.T.; Heaton, N.S. Efforts to improve the seasonal influenza vaccine. Vaccines 2018, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.C.; Zost, S.J.; Thompson, A.J.; Oyen, D.; Nycholat, C.M.; McBride, R.; Paulson, J.C.; Hensley, S.E.; Wilson, I.A. A structural explanation for the low effectiveness of the seasonal influenza H3N2 vaccine. PLOS Pathog. 2017, 13, e1006682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronski, D.M.; Janjua, N.; De Serres, G.; Sabaiduc, S.; Eshaghi, A.; Dickinson, J.; Fonseca, K.; Winter, A.-L.; Gubbay, J.B.; Krajden, M.; et al. Low 2012–13 influenza vaccine effectiveness associated with mutation in the egg-adapted H3N2 vaccine strain not antigenic drift in circulating viruses. PLoS ONE 2014, 9, e92153. [Google Scholar]
- Gubareva, L.V.; Wood, J.M.; Meyer, W.J.; Katz, J.M.; Robertson, J.S.; Major, D.; Webster, R.G. Codominant mixtures of viruses in reference strains of influenza virus due to host cell variation. Virology 1994, 199, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.W.; Kim, Y.H.; Jung, H.U.; Jeong, O.S.; Hong, E.J.; Kim, H.; Lee, J.I. Comparison of antigenic mutation during egg and cell passage cultivation of H3N2 influenza virus. Clin. Exp. Vaccine Res. 2020, 9, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Zost, S.J.; Parkhouse, K.; Gumina, M.E.; Kim, K.; Perez, S.D.; Wilson, P.C.; Treanor, J.J.; Sant, A.J.; Cobey, S.; Hensley, S.E. Contemporary H3N2 influenza viruses have a glycosylation site that alters binding of antibodies elicited by egg-adapted vaccine strains. Proc. Natl. Acad. Sci. USA 2017, 114, 12578–12583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishida, N.; Fujisaki, S.; Yokoyama, M.; Sato, H.; Saito, R.; Ikematsu, H.; Xu, H.; Takashita, E.; Tashiro, M.; Takao, S.; et al. Evaluation of influenza virus A/H3N2 and B vaccines on the basis of cross-reactivity of postvaccination human serum antibodies against influenza viruses A/H3N2 and B isolated in MDCK cells and embryonated hen eggs. Clin. Vaccine Immunol. 2012, 19, 897–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barr, I.G.; Russell, C.; Besselaar, T.G.; Cox, N.J.; Daniels, R.S.; Donis, R.; Engelhardt, O.G.; Grohmann, G.; Itamura, S.; Kelso, A.; et al. WHO recommendations for the viruses used in the 2013-2014 Northern Hemisphere influenza vaccine: Epidemiology, antigenic and genetic characteristics of influenza A(H1N1)pdm09, A(H3N2) and B influenza viruses collected from October 2012 to January 2013. Vaccine 2014, 32, 4713–4725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Alvarez, J.J.S.; Ng, S.H.; Nielsen, R.; Zhai, W. Passage adaptation correlates with the reduced efficacy of the influenza vaccine. Clin. Infect. Dis. 2019, 69, 1198–1204. [Google Scholar] [CrossRef]
- McLean, H.Q.; Caspard, H.; Griffin, M.R.; Gaglani, M.; Peters, T.R.; Poehling, K.A.; Ambrose, C.S.; Belongia, E.A. Association of prior vaccination with influenza vaccine effectiveness in children receiving live attenuated or inactivated vaccine. JAMA Netw. Open 2018, 1, e183742. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, D.M.; Chambers, C.; Sabaiduc, S.; De Serres, G.; Winter, A.-L.; Dickinson, J.; Krajden, M.; Gubbay, J.B.; Drews, S.J.; Martineau, C.; et al. A perfect storm: Impact of genomic variation and serial vaccination on low influenza vaccine effectiveness during the 2014–2015 season. Clin. Infect. Dis. 2016, 63, 21–32. [Google Scholar] [CrossRef]
- Liu, F.; Gross, F.L.; Jefferson, S.N.; Holiday, C.; Bai, Y.; Wang, L.; Zhou, B.; Levine, M.Z. Age-specific effects of vaccine egg-adaptation and immune priming on A(H3N2) antibody responses following influenza vaccination. J. Clin. Investig. 2021, 131, e146138. [Google Scholar] [CrossRef] [PubMed]
- McLean, H.Q.; Thompson, M.G.; Sundaram, M.E.; Meece, J.K.; McClure, D.L.; Friedrich, T.C.; Belongia, E.A. Impact of repeated vaccination on vaccine effectiveness against influenza A(H3N2) and B during 8 seasons. Clin. Infect. Dis. 2014, 59, 1375–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, W.E.; de Bruijn, I.A.; Palache, A.M.; Westendorp, R.G.; Osterhaus, A.D. Protection against influenza after annually repeated vaccination: A meta-analysis of serologic and field studies. Arch. Intern. Med. 1999, 159, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casado, I.; Domínguez, A.; Toledo, D.; Chamorro, J.; Astray, J.; Egurrola, M.; Fernández-Sierra, M.A.; Martín, V.; Morales-Suárez-Varela, M.; Godoy, P.; et al. Repeated influenza vaccination for preventing severe and fatal influenza infection in older adults: A multicentre case–control study. Can. Med. Assoc. J. 2018, 190, E3–E12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, E.; Smith, K.J.; Nowalk, M.P.; Raviotta, J.M.; Brown, S.T.; DePasse, J.; Zimmerman, R.K. Impact of seasonal influenza vaccination in the presence of vaccine interference. Vaccine 2018, 36, 853–858. [Google Scholar] [CrossRef]
- Nichols, M.K.; Andrew, M.K.; Ye, L.; Hatchette, T.F.; Ambrose, A.; Boivin, G.; Bowie, W.; Dos Santos, G.; Elsherif, M.; Green, K.; et al. The impact of prior season vaccination on subsequent influenza vaccine effectiveness to prevent influenza-related hospitalizations over 4 influenza seasons in Canada. Clin. Infect. Dis. 2019, 69, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Perez Rubio, A.; Eiros, J.M. Cell culture-derived flu vaccine: Present and future. Hum. Vaccines Immunother. 2018, 14, 1874–1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, S.; Vesikari, T.; Szymczakiewicz-Multanowska, A.; Lattanzi, M.; Izu, A.; Groth, N.; Holmes, S. Clinical efficacy of cell culture–derived and egg-derived inactivated subunit influenza vaccines in healthy adults. Clin. Infect. Dis. 2010, 51, 997–1004. [Google Scholar] [CrossRef]
- Izurieta, H.S.; Chillarige, Y.; Kelman, J.; Wei, Y.; Lu, Y.; Xu, W.; Lu, M.; Pratt, D.; Chu, S.; Wernecke, M.; et al. Relative effectiveness of cell-cultured and egg-based influenza vaccines among elderly persons in the United States, 2017–2018. J. Infect. Dis. 2019, 220, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.P.; Xu, X.; Hall, H.E.; Allen, J.R.; Regnery, H.L.; Cox, N.J. Comparison of 10 influenza A (H1N1 and H3N2) haemagglutinin sequences obtained directly from clinical specimens to those of MDCK cell- and egg-grown viruses. J. Gen. Virol. 1993, 74 Pt 11, 2513–2518. [Google Scholar] [CrossRef]
- Lin, Y.; Wharton, S.A.; Whittaker, L.; Dai, M.; Ermetal, B.; Lo, J.; Pontoriero, A.; Baumeister, E.; Daniels, R.S.; McCauley, J.W. The characteristics and antigenic properties of recently emerged subclade 3C.3a and 3C.2a human influenza A(H3N2) viruses passaged in MDCK cells. Influenza Other Respir. Viruses 2017, 11, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Gouma, S.; Zost, S.J.; Parkhouse, K.; Branche, A.; Topham, D.J.; Cobey, S.; Hensley, S.E. Comparison of human H3N2 antibody responses elicited by egg-based, cell-based, and recombinant protein-based influenza vaccines during the 2017–2018 season. Clin. Infect. Dis. 2020, 71, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Alvarado-Facundo, E.; Vassell, R.; Collins, L.; Colombo, R.E.; Ganesan, A.; Geaney, C.; Hrncir, D.; Lalani, T.; Markelz, A.E.; et al. Comparison of A(H3N2) neutralizing antibody responses elicited by 2018-2019 season quadrivalent influenza vaccines derived from eggs, cells, and recombinant hemagglutinin. Clin. Infect. Dis. 2021, 73, e4312–e4320. [Google Scholar] [CrossRef] [PubMed]
- Izurieta, H.S.; Chillarige, Y.; Kelman, J.; Wei, Y.; Lu, Y.; Xu, W.; Lu, M.; Pratt, D.; Wernecke, M.; MaCurdy, T.; et al. Relative effectiveness of influenza vaccines among the United States elderly, 2018–2019. J. Infect. Dis. 2020, 222, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Izurieta, H.S.; Lu, M.; Kelman, J.; Lu, Y.; Lindaas, A.; Loc, J.; Pratt, D.; Wei, Y.; Chillarige, Y.; Wernecke, M.; et al. Comparative effectiveness of influenza vaccines among US medicare beneficiaries ages 65 years and older during the 2019–2020 season. Clin. Infect. Dis. 2020, 73, e4251–e4259. [Google Scholar] [CrossRef]
- Milián, E.; Kamen, A.A. Current and emerging cell culture manufacturing technologies for influenza vaccines. BioMed Res. Int. 2015, 2015, 504831. [Google Scholar] [CrossRef] [Green Version]
- Cox, M.M.J.; Izikson, R.; Post, P.; Dunkle, L.M. Safety, efficacy, and immunogenicity of Flublok in the prevention of seasonal influenza in adults. Ther. Adv. Vaccines 2015, 3, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Rockman, S.; Laurie, K.; Parkes, S.; Wheatley, A.; Barr, I. New technologies for influenza vaccines. Microorganisms 2020, 8, 1745. [Google Scholar] [CrossRef]
- Treanor, J.J.; El Sahly, H.; King, J.; Graham, I.; Izikson, R.; Kohberger, R.; Patriarca, P.; Cox, M. Protective efficacy of a trivalent recombinant hemagglutinin protein vaccine (FluBlok(R)) against influenza in healthy adults: A randomized, placebo-controlled trial. Vaccine 2011, 29, 7733–7739. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Systematic Review of the Efficacy, Effectiveness and Safety of Newer and Enhanced Seasonal Influenza Vaccines; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2020. [Google Scholar]
- Dunkle, L.M.; Izikson, R.; Patriarca, P.; Goldenthal, K.L.; Muse, D.; Callahan, J.; Cox, M.M. Efficacy of recombinant influenza vaccine in adults 50 years of age or older. N. Engl. J. Med. 2017, 376, 2427–2436. [Google Scholar] [CrossRef] [PubMed]
- Baxter, R.; Patriarca, P.A.; Ensor, K.; Izikson, R.; Goldenthal, K.L.; Cox, M.M. Evaluation of the safety, reactogenicity and immunogenicity of FluBlok(R) trivalent recombinant baculovirus-expressed hemagglutinin influenza vaccine administered intramuscularly to healthy adults 50–64 years of age. Vaccine 2011, 29, 2272–2278. [Google Scholar] [CrossRef] [PubMed]
- Belongia, E.A.; Levine, M.Z.; Olaiya, O.; Gross, F.L.; King, J.P.; Flannery, B.; McLean, H.Q. Clinical trial to assess immunogenicity of high-dose, adjuvanted, and recombinant influenza vaccines against cell-grown A(H3N2) viruses in adults 65 to 74 years, 2017–2018. Vaccine 2020, 38, 3121–3128. [Google Scholar] [CrossRef]
- Cowling, B.J.; Perera, R.A.P.M.; Valkenburg, S.A.; Leung, N.H.L.; Iuliano, A.D.; Tam, Y.H.; Wong, J.H.F.; Fang, V.J.; Li, A.P.Y.; So, H.C.; et al. Comparative immunogenicity of several enhanced influenza vaccine options for older adults: A randomized, controlled trial. Clin. Infect. Dis. 2020, 71, 1704–1714. [Google Scholar] [CrossRef] [PubMed]
- Shinde, V.; Cho, I.; Plested, J.S.; Agrawal, S.; Fiske, J.; Cai, R.; Zhou, H.; Pham, X.; Zhu, M.; Cloney-Clark, S.; et al. Comparison of the safety and immunogenicity of a novel Matrix-M-adjuvanted nanoparticle influenza vaccine with a quadrivalent seasonal influenza vaccine in older adults: A phase 3 randomised controlled trial. Lancet Infect. Dis. 2021, 22, 73–84. [Google Scholar] [CrossRef]
- Novavax’ NanoFlu Achieves All Primary Endpoints in Phase 3 Clinical Trial. 2020. Available online: https://ir.novavax.com/2020-03-24-Novavax-NanoFlu-Achieves-All-Primary-Endpoints-In-Phase-3-Clinical-Trial (accessed on 4 November 2021).
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Petsch, B.; Schnee, M.; Vogel, A.B.; Lange, E.; Hoffmann, B.; Voss, D.; Schlake, T.; Thess, A.; Kallen, K.-J.; Stitz, L.; et al. Protective efficacy of in vitro synthesized, specific mRNA vaccines against influenza A virus infection. Nat. Biotechnol. 2012, 30, 1210–1216. [Google Scholar] [CrossRef]
- Sanofi and Translate Bio Initiate Phase 1 Clinical Trial of mRNA Influenza Vaccine. Available online: https://www.sanofi.com/en/media-room/press-releases/2021/2021-06-22-07-00-00-2250633 (accessed on 4 November 2021).
- Feldman, R.A.; Fuhr, R.; Smolenov, I.; Ribeiro, A.; Panther, L.; Watson, M.; Senn, J.J.; Smith, M.; Almarsson, Ӧ.; Pujar, H.S.; et al. mRNA vaccines against H10N8 and H7N9 influenza viruses of pandemic potential are immunogenic and well tolerated in healthy adults in phase 1 randomized clinical trials. Vaccine 2019, 37, 3326–3334. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Parkhouse, K.; Kirkpatrick, E.; McMahon, M.; Zost, S.; Mui, B.L.; Tam, Y.K.; Karikó, K.; Barbosa, C.J.; Madden, T.D.; et al. Nucleoside-modified mRNA immunization elicits influenza virus hemagglutinin stalk-specific antibodies. Nat. Commun. 2018, 9, 3361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scorza, F.B.; Pardi, N. New kids on the block: RNA-based influenza virus vaccines. Vaccines 2018, 6, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, X.; Qi, Y.; Wang, M.; Yu, N.; Nan, F.; Zhang, H.; Tian, M.; Li, C.; Lu, H.; Jin, N. mRNA vaccines encoding the HA protein of influenza a H1N1 virus delivered by cationic lipid nanoparticles induce protective immune responses in mice. Vaccines 2020, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sautto, G.A.; Kirchenbaum, G.A.; Ross, T.M. Towards a universal influenza vaccine: Different approaches for one goal. Virol. J. 2018, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.J.; Crank, M.C.; Shiver, J.; Graham, B.S.; Mascola, J.R.; Nabel, G.J. Next-generation influenza vaccines: Opportunities and challenges. Nat. Rev. Drug. Discov. 2020, 19, 239–252. [Google Scholar] [CrossRef]
- Liu, F.; Tzeng, W.-P.; Horner, L.; Kamal, R.P.; Tatum, H.R.; Blanchard, E.G.; Xu, X.; York, I.; Tumpey, T.M.; Katz, J.M.; et al. Influence of immune priming and egg adaptation in the vaccine on antibody responses to circulating A(H1N1)pdm09 viruses after influenza vaccination in adults. J. Infect. Dis. 2018, 218, 1571–1581. [Google Scholar] [CrossRef]
- Robertson, J.S.; Bootman, J.S.; Newman, R.; Oxford, J.S.; Daniels, R.S.; Webster, R.G.; Schild, G.C. Structural changes in the haemagglutinin which accompany egg adaptation of an influenza A(H1N1) virus. Virology 1987, 160, 31–37. [Google Scholar] [CrossRef]
- Ortiz de Lejarazu-Leonardo, R.; Montomoli, E.; Wojcik, R.; Christopher, S.; Mosnier, A.; Pariani, E.; Trilla Garcia, A.; Fickenscher, H.; Gartner, B.C.; Jandhyala, R.; et al. Estimation of reduction in influenza vaccine effectiveness due to egg-adaptation changes-systematic literature review and expert consensus. Vaccines 2021, 9, 1255. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, S.; Suphaphiphat, P.; Van Boxmeer, J.; Haag, M.; Leav, B.; Iheanacho, I.; Kistler, K.; De Lejarazu, R.O. Retrospective assessment of the antigenic similarity of egg-propagated and cell culture-propagated reference influenza viruses as compared with circulating viruses across influenza seasons 2002–2003 to 2017–2018. Int. J. Environ. Res. Public Health 2020, 17, 5423. [Google Scholar] [CrossRef] [PubMed]
- Mott, J.A.; Fry, A.M.; Kondor, R.; Wentworth, D.E.; Olsen, S.J. Re-emergence of influenza virus circulation during 2020 in parts of tropical Asia: Implications for other countries. Influenza Other Respir. Viruses 2021, 15, 415–418. [Google Scholar] [CrossRef]
- Wen, F.; Bedford, T.; Cobey, S. Explaining the geographical origins of seasonal influenza A (H3N2). Proc. Biol. Sci. 2016, 283. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.A.; Jones, T.C.; Barr, I.G.; Cox, N.J.; Garten, R.J.; Gregory, V.; Gust, I.D.; Hampson, A.W.; Hay, A.J.; Hurt, A.C.; et al. The global circulation of seasonal influenza a (H3N2) viruses. Science 2008, 320, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Bahl, J.; Nelson, M.I.; Chan, K.H.; Chen, R.; Vijaykrishna, D.; Halpin, R.A.; Stockwell, T.; Lin, X.; Wentworth, D.; Ghedin, E.; et al. Temporally structured metapopulation dynamics and persistence of influenza A H3N2 virus in humans. Proc. Natl. Acad. Sci. USA 2011, 108, 19359–19364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedford, T.; Cobey, S.; Beerli, P.; Pascual, M. Global migration dynamics underlie evolution and persistence of human influenza A (H3N2). PLOS Pathog. 2010, 6, e1000918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedford, T.; Riley, S.; Barr, I.G.; Broor, S.; Chadha, M.; Cox, N.J.; Daniels, R.S.; Gunasekaran, C.P.; Hurt, A.C.; Kelso, A.; et al. Global circulation patterns of seasonal influenza viruses vary with antigenic drift. Nature 2015, 523, 217–220. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Percentage of Cases by Influenza Virus Type/Subtype | |||||
|---|---|---|---|---|---|
| Country a | 10-Year Total Number of Influenza Positive Reported | A(H3) | A(H1, H1N1pdm09) | A(Unsubtyped) | B Total |
| China | 679,983 | 40.8% | 27.0% | 0.9% | 31.3% |
| Japan | 82,710 | 45.4% | 29.9% | 0.1% | 24.6% |
| Australia | 53,577 | 34.1% | 15.2% | 29.6% | 21.2% |
| Republic of Korea | 18,444 | 39.6% | 25.9% | 0.0% | 34.6% |
| New Zealand | 17,235 | 25.9% | 15.1% | 25.9% | 33.1% |
| Singapore | 10,985 | 38.4% | 29.8% | 0.8% | 31.0% |
| Vietnam | 6955 | 32.7% | 32.8% | 0.0% | 34.5% |
| Philippines | 6105 | 24.9% | 26.1% | 6.2% | 42.8% |
| Mongolia | 5677 | 48.1% | 25.1% | 2.5% | 24.3% |
| Cambodia | 4603 | 31.4% | 31.2% | 0.0% | 37.4% |
| Laos | 4512 | 33.5% | 27.8% | 0.1% | 38.6% |
| Malaysia | 3322 | 17.8% | 31.1% | 14.2% | 36.9% |
| Papua New Guinea | 261 | 19.5% | 46.4% | 5.0% | 29.1% |
| Total | 894,369 | ||||
| Mean, % (95% CI) | 33.2% (27.6–38.9) | 27.9% (23.2–32.7) | 6.6% (0.4–12.7) | 32.2% (28.5–36.0) | |
| Category | Years | A(H3N2) *,** | A(H1N1) * | B-Lineage * | Region | Ref. |
|---|---|---|---|---|---|---|
| Incidence -Influenza-like illness consultations per 1000 person-year | 2010–2015 | 0.7 (0.4–0.9) | 0.5 (0.3–0.7) | 0.3 (0.0–0.5) | China | [6] |
| Hospitalization -Rates per 100,000 person-years | 2010–2011 | 55 | 33 | 26 | China | [7] |
| -Risk ratio to A(H1N1) | 2009–2011 | 1.8 (1.3–2.6) | 1 | Hong Kong | [8] | |
| Death -Excess mortality rates per 100,000 person-seasons | 2010–2015 | 2.6 (2.4–2.8) | 1.6 (1.5–1.7) | 2.3 (2.1–2.5) | China | [11] |
| -Excess mortality rates per 100,000 person-seasons | 1998–2009 | 6.88 (4.26–9.37) | 1.6 (−0.34–3.34) | 2.5 (−0.51–5.33) | Hong Kong | [9] |
| -All-cause death risk ratio | 2009–2011 | 2.6 (1.8–3.7) | 1 | Hong Kong | [8] | |
| -Respiratory death risk ratio | 2009–2011 | 1.5 (1.0–2.1) | 1 | Hong Kong | [8] | |
| -All-cause death risk ratio | 1996–2003 | 1.04 (1.02–1.05) | 1 | 1.01 (1.00–1.02) | Singapore | [12] |
| -Respiratory death risk ratio | 1996–2003 | 1.08 (1.04–1.12) | 1 | 1.00 (0.97–1.03) | Singapore | [12] |
| -Excess mortality rates per 100,000 person-seasons | 2009–2016 | 8.66 (5.88–11.35) | 5.99 (3.41–8.46) | 4.77 (1.04–8.24) | Hong Kong | [10] |
| Season | Region/City | Overall VE % (95% CI a) | VE % (95% CI) against | Circulating Subtype b | No. Positives | Study Population | Outcome c | Ref. | ||
|---|---|---|---|---|---|---|---|---|---|---|
| A(H3N2) | A(H1N1) | B | ||||||||
| 2012–13 | Beijing, China | 52 (12, 74) | 43 (−30, 75) | 59 (8, 82) | Not provided | A(H1N1) | 695 | All ages | Medically attended ILI | [20] |
| 2013–14 | Beijing, China | 32 (−48, 69) | 22 (−253, 83) | 59 (−79, 90) | −20 (−239, 58) | A(H1N1) | 133 | 60 years and older | Medically attended ILI | [21] |
| 2013–14 | Beijing, China | 47 (−20, 77) | 60 (−110, 92) | Not provided | 42 (−60, 79) | A(H1N1) | 353 | All ages | Influenza-associated hospitalization | [22] |
| 2014–15 | 5 (−53, 41) | 28 (−42, 63) | Not provided | −32 (−154, 32) | A(H3N2) | |||||
| 2014–15 | Beijing, China | −18 (−49, 6) | −25 (−70, 8) | Not provided | −8 (−50, 23) | A(H3N2) | 3434 | All ages | Medically attended ILI | [23] |
| 2015–16 | Beijing, China | 8 (−16, 27) | 54 (16, 74) | 18 (−38, 52) | −7 (−38, 18) | A(H1N1) and B | 2969 | All ages | Medically attended ILI | [18] |
| 2015–16 | Beijing, China | −38 (−103, 6) | −5 (−108, 47) | −62 (−212, 16) | −45 (−153, 16) | A(H1N1) and B | 356 | All ages | Influenza-associated hospitalization | [24] |
| 2016–17 | Suzhou, China | 21 (−42, 56) | 1 (−86, 47) | Not provided | 63 (−65, 92) | A(H3N2) | 70 | Children 36–72 months old | Influenza infection | [25] |
| 2016–17 | Beijing, China | 25 (0, 43) | 2 (−35, 29) | 54 (22–73) | Not provided | A(H3N2) | 2626 | All ages | Influenza-associated outpatient visits | [26] |
| 2016–17 | Beijing, China | 69 (51, 81) | 73 (53, 85) | 60 (−15, 86) | Not provided | A(H3N2) | 176 | School Children 6–19 years old | Influenza infection | [27] |
| 2016–17 | Beijing, China | 33 (−22, 63) | 30 (−30, 62) | Not provided | Not provided | A(H3N2) | 145 | 60 year and older | Influenza-associated hospitalization | [28] |
| 2017–18 | 5 (−72, 47) | −38 (–294, 52) | 29 (−93, 74) | 4 (−114, 56) | A(H1N1) and B | 149 | ||||
| 2009–13 | Hong Kong, China | 62 (43, 74) | 37 (−26, 68) | 72 (39, 87) | 69 (42, 83) | N/A | 451 | Children 6m–17 years old | Influenza-associated hospitalization | [29] |
| 2017–18 | Hong Kong, China | 59 (41, 72) | 41 (−60, 82) | 86 (66, 95) | 54 (35, 75) | N/A | 467 | All ages | Primary care visits | [30] |
| 2019–20 | Hong Kong, China | 65 (46, 78) | 12 (−80, 57) | 74 (54, 85) | 85 (30, 97) | N/A | 198 | Children 6m–17 years old | Influenza-associated hospitalization | [31] |
| 1997–2018 | Japan | 19 (2, 33) | 19 (−13, 43) | 22 (−26,52) | 15 (−14, 36) | N/A | (Meta-analysis from 143 studies) | [32] | ||
| 2016–17 | Korea | −36 (−115, 14) | −52 (−147, 6) | Not provided | Not provided | A(H3N2) | 216 | Adults | Influenza infection | [33] |
| 2010 | Western Australia | 68 (35, 85) | 3 (−495, 84) | 80 (41, 93) | 66 (1,89) | A(H1N1) | 448 | All ages | ILI GP visits | [34] |
| 2011 | 52 (1, 77) | −55 (−386, 5) | 71 (15, 90) | 85 −30, 98) | A(H1N1) | 351 | ||||
| 2012 | 49 (30, 63) | 46 (21, 63) | 8 (−868, 91) | 54 (26, 71) | A(H3N2) | 1161 | ||||
| 2012 | Australia | 38 (24, 49) | 30 (14, 44) | 54 (−28, 83) | 56 (37, 70) | A(H3N2) | 1462 | All ages | ILI GP visits | [35] |
| 2013 | 60 (45, 70) | 67 (39, 82) | 59 (33, 74) | 57 (30, 73) | A(H1N1) and A(H3N2) | 441 | ||||
| 2014 | 44 (31,55) | 26 (1, 45) | 55 (39, 67) | 54 (21, 73) | A(H1N1) | 891 | ||||
| 2015 | Australia | 54 (42, 63) | 44 (21, 60) | 79 (33, 93) | 58 (45, 68) | A(H3N2) and B | 857 | All ages | Medically attended ILI | [36] |
| 2017 | Australia | 33 (17, 46) | 10 (−16, 31) | 50 (8, 74) | 57 (41, 69) | A(H3N2) | 1060 | All ages | Medically attended ILI | [37] |
| 2010–13 | Singapore | not provided | 33 (−4, 57) | 84 (78, 88) | 84 (79, 86) | N/A | 1198 | Military Adults | Influenza infection | [38] |
| 2017 | Singapore | 40 (−12, 68) | 57 (6, 80) | −43 (−312, 50) | Not provided | A(H3N2) | 118 | Seniors in Long-term care facilities | Influenza infection | [39] |
| 2012–15 | Auckland, NZ | 37 (23, 48) | 26 (5, 42) | 42 (14, 61) | 49 (30, 63) | N/A | 842 | Adults | ICU admission and Severe Disease | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, M.; Zanin, M.; Wong, S.-S. Subtype H3N2 Influenza A Viruses: An Unmet Challenge in the Western Pacific. Vaccines 2022, 10, 112. https://doi.org/10.3390/vaccines10010112
Kang M, Zanin M, Wong S-S. Subtype H3N2 Influenza A Viruses: An Unmet Challenge in the Western Pacific. Vaccines. 2022; 10(1):112. https://doi.org/10.3390/vaccines10010112
Chicago/Turabian StyleKang, Min, Mark Zanin, and Sook-San Wong. 2022. "Subtype H3N2 Influenza A Viruses: An Unmet Challenge in the Western Pacific" Vaccines 10, no. 1: 112. https://doi.org/10.3390/vaccines10010112
APA StyleKang, M., Zanin, M., & Wong, S.-S. (2022). Subtype H3N2 Influenza A Viruses: An Unmet Challenge in the Western Pacific. Vaccines, 10(1), 112. https://doi.org/10.3390/vaccines10010112