Improvement of Health-Promoting Functionality of Rye Bread by Fortification with Free and Microencapsulated Powders from Amelanchier alnifolia Nutt



Abstract
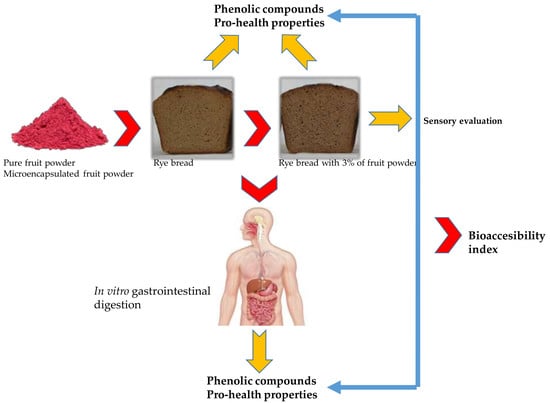
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Materials
2.3. Rye Bread Preparation
2.4. Sensory Attributes and Colour Parameters
2.5. Extraction Procedures
2.5.1. Digestion In Vitro
2.5.2. Chemical Extraction
2.5.3. Buffer Extraction
2.6. Identyfication and Quantyfication of Polyphenolic Compounds
2.7. Health-Promoting Properties
2.7.1. Antiradical Capacity
2.7.2. Reducing Potential
2.7.3. Ability to Inhibit the Activity of COX 1 and COX-2
2.7.4. Ability to Inhibit the Activity of Lipoxygenase (LOX)
2.7.5. Ability for Inhibit Acetylcholinesterase Activity (AChE)
2.7.6. Activity of α-Amylase Inhibitors
2.7.7. Activity of α-Glucoamyalse Inhibitors
2.7.8. Activity of Lipase Inhibitors
2.7.9. Theoretical Approach
2.8. Relative Digestibility (RD)
2.8.1. Relative Digestibility Proteins
2.8.2. Relative Digestibility Starch
2.9. Statistical Analysis
3. Results and Discussion
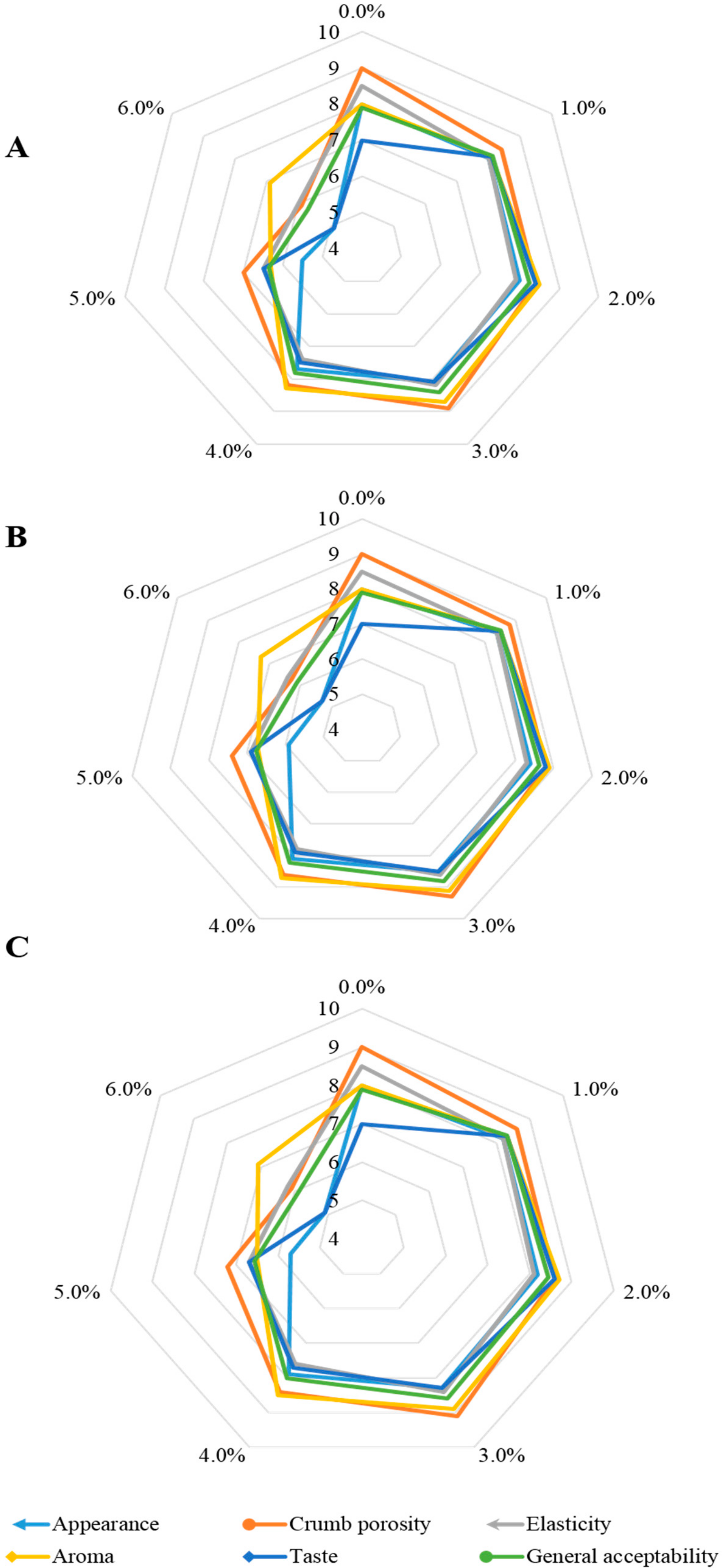
3.1. Sensory Evaluation and Colour Parameters
3.2. Phenolic Compounds and Their Relative Bioaccesibility
3.3. Pro-Healthy Potency and Their Bioaccesibility
3.4. Relative Digestibility of Starches and Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bondia-Pons, I.; Aura, A.-M.; Vuorela, S.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Rye phenolics in nutrition and health. J. Cereal Sci. 2009, 49, 323–336. [Google Scholar] [CrossRef]
- Banu, I.; Vasilean, I.; Aprodu, I. Effect of Lactic Fermentation on Antioxidant Capacity of Rye Sourdough and Bread. Food Sci. Technol. Res. 2010, 16, 571–576. [Google Scholar] [CrossRef]
- Pejcz, E.; Gil, Z.; Wojciechowicz-Budzisz, A.; Półtorak, M.; Romanowska, A. Effect of technological process on the nutritional quality of naked barley enriched rye bread. J. Cereal Sci. 2015, 65, 215–219. [Google Scholar] [CrossRef]
- Bajerska, J.; Chmurzynska, A.; Mildner-Szkudlarz, S.; Drzymała-Czyż, S. Effect of rye bread enriched with tomato pomace on fat absorption and lipid metabolism in rats fed a high-fat diet. J. Sci. Food Agric. 2015, 95, 1918–1924. [Google Scholar] [CrossRef]
- Bajerska, J.; Mildner-Szkudlarz, S.; Walkowiak, J. Effects of Rye Bread Enriched with Green Tea Extract on Weight Maintenance and the Characteristics of Metabolic Syndrome Following Weight Loss: A Pilot Study. J. Med. Food 2014, 18, 698–705. [Google Scholar] [CrossRef]
- Mildner-Szkudlarz, S.; Zawirska-Wojtasiak, R.; Szwengiel, A.; Pacyński, M. Use of grape by-product as a source of dietary fibre and phenolic compounds in sourdough mixed rye bread. Int. J. Food Sci. Technol. 2011, 46, 1485–1493. [Google Scholar] [CrossRef]
- Juríková, T.; Balla, S.; Sochor, J.; Pohanka, M.; Mlcek, J.; Baron, M. Flavonoid profile of saskatoon berries (Amelanchier alnifolia Nutt.) and their health promoting effects. Molecules 2013, 18, 12571–12586. [Google Scholar] [CrossRef]
- Rop, O.; Řezníček, V.; Mlček, J.; Juríková, T.; Sochor, J.; Kizek, R.; Humpolíček, P.; Balík, J. Nutritional values of new Czech cultivars of Saskatoon berries (Amelanchier alnifolia Nutt.). Hortic. Sci. 2012, 39, 123–128. [Google Scholar] [CrossRef]
- Lachowicz, S.; Oszmiański, J.; Seliga, Ł.; Pluta, S. Phytochemical Composition and Antioxidant Capacity of Seven Saskatoon Berry (Amelanchier alnifolia Nutt.) Genotypes Grown in Poland. Molecules 2017, 22, 853. [Google Scholar] [CrossRef]
- Lachowicz, S.; Wiśniewski, R.; Ochmian, I.; Drzymała, K.; Pluta, S. Anti-Microbiological, Anti-Hyperglycemic and Anti-Obesity Potency of Natural Antioxidants in Fruit Fractions of Saskatoon Berry. Antioxidants 2019, 8, 397. [Google Scholar] [CrossRef]
- Tian, Y.; Puganen, A.; Alakomi, H.-L.; Uusitupa, A.; Saarela, M.; Yang, B. Antioxidant and antibacterial activities of aqueous ethanol extracts of berries, leaves, and branches of berry plants. Food Res. Int. 2018, 106, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Khafipour, E.; Sepehri, S.; Huang, F.; Beta, T.; Shen, G.X. Impact of Saskatoon berry powder on insulin resistance and relationship with intestinal microbiota in high fat–high sucrose diet-induced obese mice. J. Nutr. Biochem. 2019, 69, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Ezhilarasi, P.N.; Indrani, D.; Jena, B.S.; Anandharamakrishnan, C. Freeze drying technique for microencapsulation of Garcinia fruit extract and its effect on bread quality. J. Food Eng. 2013, 117, 513–520. [Google Scholar] [CrossRef]
- Vitaglione, P.; Barone Lumaga, R.; Ferracane, R.; Radetsky, I.; Mennella, I.; Schettino, R.; Koder, S.; Shimoni, E.; Fogliano, V. Curcumin Bioavailability from Enriched Bread: The Effect of Microencapsulated Ingredients. J. Agric. Food Chem. 2012, 60, 3357–3366. [Google Scholar] [CrossRef]
- Chávez-Santoscoy, R.A.; Gutiérrez-Uribe, J.A.; Serna-Saldivar, S.O.; Perez-Carrillo, E. Production of maize tortillas and cookies from nixtamalized flour enriched with anthocyanins, flavonoids and saponins extracted from black bean (Phaseolus vulgaris) seed coats. Food Chem. 2016, 192, 90–97. [Google Scholar] [CrossRef]
- Morris, C.; Morris, G.A. The effect of inulin and fructo-oligosaccharide supplementation on the textural, rheological and sensory properties of bread and their role in weight management: A review. Food Chem. 2012, 133, 237–248. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Świeca, M.; Dziki, D.; Sęczyk, Ł.; Złotek, U.; Różyło, R.; Kaszuba, K.; Ryszawy, D.; Czyż, J. Anticancer and Antioxidant Activity of Bread Enriched with Broccoli Sprouts. BioMed Res. Int. 2014, 2014, 608053. [Google Scholar] [CrossRef]
- Lachowicz, S.; Michalska, A.; Lech, K.; Majerska, J.; Oszmiański, J.; Figiel, A. Comparison of the effect of four drying methods on polyphenols in saskatoon berry. LWT 2019, 111, 727–736. [Google Scholar] [CrossRef]
- Lachowicz, S.; Michalska-Ciechanowska, A.; Oszmiański, J. The Impact of Maltodextrin and Inulin on the Protection of Natural Antioxidants in Powders Made of Saskatoon Berry Fruit, Juice, and Pomace as Functional Food Ingredients. Molecules 2020, 25, 1805. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food – an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Oszmiański, J.; Kolniak-Ostek, J.; Lachowicz, S.; Gorzelany, J.; Matłok, N. Effect of dried powder preparation process on polyphenolic content and antioxidant capacity of cranberry (Vaccinium macrocarpon L.). Ind. Crops Prod. 2015, 77, 658–665. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Chen, H.Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Axelrod, B.; Cheesbrough, T.M.; Laakso, S. LOX from soybeans. Methods Enzymol. 1981, 71, 441–451. [Google Scholar]
- Jakubczyk, A.; Karaś, M.; Złotek, U.; Szymanowska, U. Identification of potential inhibitory peptides of enzymes involved in the metabolic syndrome obtained by simulated gastrointestinal digestion of fermented bean (Phaseolus vulgaris L.) seeds. Food Res. Int. 2017, 100, 489–496. [Google Scholar] [CrossRef]
- Sęczyk, Ł.; Świeca, M.; Gawlik-Dziki, U. Effect of carob (Ceratonia siliqua L.) flour on the antioxidant potential, nutritional quality, and sensory characteristics of fortified durum wheat pasta. Food Chem. 2016, 194, 637–642. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Tolić, M.-T.; Krbavčić, I.P.; Vujević, P.; Milinović, B.; Jurčević, I.L.; Vahčić, N. Effects of Weather Conditions on Phenolic Content and Antioxidant Capacity in Juice of Chokeberries (Aronia melanocarpa L.). Pol. J. Food Nutr. Sci. 2017, 67, 67–74. [Google Scholar] [CrossRef]
- Bajerska, J.; Mildner-Szkudlarz, S.; Podgórski, T.; Oszmatek-Pruszyńska, E. Saffron (Crocus sativus L.) Powder as an Ingredient of Rye Bread: An Anti-Diabetic Evaluation. J. Med. Food 2013, 16, 847–856. [Google Scholar] [CrossRef]
- Bakowska-Barczak, A.M.; Kolodziejczyk, P.P. Black currant polyphenols: Their storage stability and microencapsulation. Ind. Crops Prod. 2011, 34, 1301–1309. [Google Scholar] [CrossRef]
- Saénz, C.; Tapia, S.; Chávez, J.; Robert, P. Microencapsulation by spray drying of bioactive compounds from cactus pear (Opuntia ficus-indica). Food Chem. 2009, 114, 616–622. [Google Scholar] [CrossRef]
- Cai, Y.Z.; Corke, H. Production and Properties of Spray-dried Amaranthus Betacyanin Pigments. J. Food Sci. 2000, 65, 1248–1252. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. Soymilk phenolic compounds, isoflavones and antioxidant activity as affected by in vitro gastrointestinal digestion. Food Chem. 2013, 136, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Świeca, M.; Gawlik-Dziki, U.; Dziki, D.; Baraniak, B. Wheat bread enriched with green coffee—In vitro bioaccessibility and bioavailability of phenolics and antioxidant activity. Food Chem. 2017, 221, 1451–1457. [Google Scholar] [CrossRef]
- Manzocco, L.; Calligaris, S.; Mastrocola, D.; Nicoli, M.C.; Lerici, C.R. Review of non-enzymatic browning and antioxidant capacity in processed foods. Trends Food Sci. Technol. 2000, 11, 340–346. [Google Scholar] [CrossRef]
- Davidov-Pardo, G.; Moreno, M.; Arozarena, I.; Marín-Arroyo, M.R.; Bleibaum, R.N.; Bruhn, C.M. Sensory and Consumer Perception of the Addition of Grape Seed Extracts in Cookies. J. Food Sci. 2012, 77, S430–S438. [Google Scholar] [CrossRef]
- Kraska, P.; Andruszczak, S.; Kwiecinska-Poppe, E.; Rozylo, K.; Swieca, M.; Palys, E. Chemical composition of seeds of linseed (Linum usitatissimum L.) cultivars depending on the intensity of agricultural technology. J. Elem. 2016, 21. [Google Scholar] [CrossRef]
- Złotek, U.; Szychowski, K.A.; Świeca, M. Potential in vitro antioxidant, anti-inflammatory, antidiabetic, and anticancer effect of arachidonic acid-elicited basil leaves. J. Funct. Foods 2017, 36, 290–299. [Google Scholar] [CrossRef]
- Yamamoto, S.; Katsukawa, M.; Nakano, A.; Hiraki, E.; Nishimura, K.; Jisaka, M.; Yokota, K.; Ueda, N. Arachidonate 12-lipoxygenases with reference to their selective inhibitors. Biochem. Biophys. Res. Commun. 2005, 338, 122–127. [Google Scholar] [CrossRef]
- Naithani, S.; Saracco, S.A.; Butler, C.A.; Fox, T.D. Interactions among COX1, COX2, andCOX3 mRNA-specific Translational Activator Proteins on the Inner Surface of the Mitochondrial Inner Membrane of Saccharomyces cerevisiae. Mol. Biol. Cell 2002, 14, 324–333. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jaksevicius, A.; Carew, M.; Mistry, C.; Modjtahedi, H.; Opara, E.I. Inhibitory Effects of Culinary Herbs and Spices on the Growth of HCA-7 Colorectal Cancer Cells and Their COX-2 Expression. Nutrients 2017, 9, 1051. [Google Scholar] [CrossRef] [PubMed]
- Moschona, A.; Liakopoulou-Kyriakides, M. Encapsulation of biological active phenolic compounds extracted from wine wastes in alginate-chitosan microbeads. J. Microencapsul. 2018, 35, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Sujith, K.; Ronald, D.C.; Suba, V. Inhibitory effect of Anacycluspyrethrum extract on acetylcholinesterase enzyme by invitro methods. Pharmacogn. J. 2012, 4, 31–34. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Ramdath, D.D.; Tang, Y.; Chen, P.X.; Liu, R.; Liu, Q.; Tsao, R. Phenolic profiles of 20 Canadian lentil cultivars and their contribution to antioxidant activity and inhibitory effects on α-glucosidase and pancreatic lipase. Food Chem. 2015, 172, 862–872. [Google Scholar] [CrossRef]
- Sandberg, J.C.; Björck, I.M.E.; Nilsson, A.C. Effects of whole grain rye, with and without resistant starch type 2 supplementation, on glucose tolerance, gut hormones, inflammation and appetite regulation in an 11–14.5 hour perspective; a randomized controlled study in healthy subjects. Nutr. J. 2017, 16, 25. [Google Scholar] [CrossRef]
- Yousif, A.; Nhepera, D.; Johnson, S. Influence of sorghum flour addition on flat bread in vitro starch digestibility, antioxidant capacity and consumer acceptability. Food Chem. 2012, 134, 880–887. [Google Scholar] [CrossRef]
- Karim, Z.; Holmes, M.; Orfila, C. Inhibitory effect of chlorogenic acid on digestion of potato starch. Food Chem. 2017, 217, 498–504. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Shares [%] | Rye Bread Enriched with FP | Rye Bread Enriched with FPM | Rye Bread Enriched with FPI | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L* | a* | b* | ΔE | L* | a* | b* | ΔE | L* | a* | b* | ΔE | ||
| Bread crust/dried fruit | BC | 52.3 ± 1.0a 1 | 2.8 ± 0.1e | 17.0 ± 0.3a | - | - | - | - | - | - | - | - | - |
| B1 | 49.3 ± 1.0b | 3.9 ± 0.1d | 14.1 ± 0.3b | 4.4 ± 0.1f | 50.2 ± 1.0a | 3.2 ± 0.1e | 14.3 ± 0.3a | 3.5 ± 0.1f | 49.7 ± 1.0a | 3.4 ± 0.1e | 14.5 ± 0.3a | 3.7 ± 0.1e | |
| B2 | 48.1 ± 1.0c | 4.0 ± 0.1d | 12.0 ± 0.2c | 6.6 ± 0.1e | 49.4 ± 1.0b | 3.7 ± 0.1d | 13.5 ± 0.3b | 4.7 ± 0.1e | 49.1 ± 1.0a | 3.8 ± 0.1d | 13.9 ± 0.3b | 4.6 ± 0.1d | |
| B3 | 46.2 ± 0.9d | 4.9 ± 0.1c | 11.2 ± 0.2d | 8.7 ± 0.2d | 47.4 ± 0.9d | 4.5 ± 0.1c | 12.6 ± 0.3c | 6.8 ± 0.1c | 47.1 ± 0.9b | 4.6 ± 0.1b | 13.0 ± 0.3c | 6.8 ± 0.1b | |
| B4 | 44.9 ± 0.9e | 5.8 ± 0.1b | 9.2 ± 0.2e | 11.2 ± 0.2c | 48.4 ± 1.0c | 4.3 ± 0.1c | 13.0 ± 0.3b | 5.8 ± 0.1d | 47.7 ± 1.0b | 4.2 ± 0.1c | 12.9 ± 0.3c | 6.4 ± 0.1c | |
| B5 | 42.8 ± 0.9f | 6.6 ± 0.1b | 8.2 ± 0.2f | 13.5 ± 0.3b | 45.5 ± 0.9e | 4.8 ± 0.1b | 11.4 ± 0.2d | 9.0 ± 0.2b | 47.5 ± 1.0b | 4.7 ± 0.1b | 12.6 ± 0.3c | 6.8 ± 0.1b | |
| B6 | 40.5 ± 0.8g | 7.3 ± 0.1a | 6.2 ± 0.1g | 16.6 ± 0.3a | 43.1 ± 0.9f | 5.2 ± 0.1a | 8.6 ± 0.2e | 12.7 ± 0.3a | 44.9 ± 0.9c | 5.2 ± 0.1a | 9.5 ± 0.2d | 10.8 ± 0.2a | |
| Bread crumb/dried fruit | BCC | 40.5 ± 0.8a | 7.5 ± 0.2a | 9.1 ± 0.2a | - | - | - | - | - | - | - | - | - |
| B1C | 39.9 ± 0.8b | 6.4 ± 0.1b | 7.8 ± 0.2b | 1.8 ± 0.1c | 40.7 ± 0.8a | 5.3 ± 0.1b | 7.9 ± 0.2b | 2.5 ± 0.1e | 40.3 ± 0.8a | 5.7 ± 0.1b | 8.0 ± 0.2b | 2.1 ± 0.1e | |
| B2C | 39.8 ± 0.8b | 6.3 ± 0.1b | 7.8 ± 0.2b | 1.9 ± 0.1c | 40.8 ± 0.8a | 5.8 ± 0.1a | 8.7 ± 0.2a | 1.8 ± 0.1f | 40.6 ± 0.8a | 6.0 ± 0.1a | 9.0 ± 0.2a | 1.6 ± 0.1f | |
| B3C | 38.4 ± 0.8b | 5.7 ± 0.1c | 5.7 ± 0.1c | 4.4 ± 0.1b | 39.3 ± 0.8b | 5.2 ± 0.1b | 6.4 ± 0.1c | 3.7 ± 0.1d | 39.1 ± 0.8b | 5.4 ± 0.1b | 6.6 ± 0.1d | 3.6 ± 0.1d | |
| B4C | 38.7 ± 0.8b | 5.2 ± 0.1c | 5.6 ± 0.1c | 4.6 ± 0.1b | 41.7 ± 0.8a | 3.8 ± 0.1c | 7.9 ± 0.2b | 4.1 ± 0.1c | 41.0 ± 0.8a | 3.7 ± 0.1c | 7.8 ± 0.2c | 4.1 ± 0.1c | |
| B5C | 36.6 ± 0.7c | 3.4 ± 0.1e | 2.8 ± 0.1e | 8.4 ± 0.2a | 39.0 ± 0.8b | 2.5 ± 0.1d | 3.9 ± 0.1e | 7.4 ± 0.1a | 40.6 ± 0.8a | 2.4 ± 0.1e | 4.3 ± 0.1e | 7.0 ± 0.1b | |
| B6C | 35.9 ± 0.7c | 4.8 ± 0.1d | 3.2 ± 0.1d | 7.9 ± 0.2a | 38.2 ± 0.8c | 3.5 ± 0.1c | 4.5 ± 0.1d | 6.5 ± 0.1b | 39.8 ± 0.8b | 3.4 ± 0.1d | 4.9 ± 0.1e | 5.9 ± 0.1a | |
| Compounds | Bread Sample | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| BR1 | BR2 | BR3 | BR4 | BR5 | BR6 | ||||
| Anthocyanins | cyanidin- -3-O-galactoside | ND | FP | 18.7 ± 0.2m 1 | 38.3 ± 0.3l | 88.2 ± 0.8i | 135 ± 1g | 195 ± 1f | 281 ± 2c |
| FPM | 60.8 ± 0.5k | 69.9 ± 0.6j | 107 ± 1h | 130 ± 1g | 230 ± 2d | 322 ± 2a | |||
| FPI | 65.9 ± 0.6j | 75.8 ± 0.7i | 108 ± 1h | 136 ± 1g | 213 ± 1e | 298 ± 2b | |||
| cyanidin- -3-O-glucoside | ND | FP | 2.5 ± 0.1j | 6.8 ± 0.1i | 8.7 ± 0.1g | 18.1 ± 0.2c | 30.0 ± 0.3b | 41.8 ± 0.4a | |
| FPM | 6.2 ± 0.1i | 7.1 ± 0.1h | 10.3 ± 0.1f | 11.0 ± 0.1e | 12.2 ± 0.1e | 14.6 ± 0.1d | |||
| FPI | 6.4 ± 0.1i | 7.4 ± 0.1h | 8.8 ± 0.1g | 10.9 ± 0.1e | 11.7 ± 0.1e | 14.0 ± 0.1d | |||
| cyanidin- -3-O-arabinoside | ND | FP | ND | 2.3 ± 0.1g | 3.6 ± 0.1f | 5.5 ± 0.1c | 7.5 ± 0.1b | 11.1 ± 0.1a | |
| FPM | 3.2 ± 0.1f | 3.6 ± 0.1f | 4.1 ± 0.1e | 4.2 ± 0.1e | 5.0 ± 0.1d | 6.0 ± 0.1c | |||
| FPI | 3.4 ± 0.1f | 3.9 ± 0.1f | 4.2 ± 0.1e | 4.4 ± 0.1e | 4.7 ± 0.1e | 5.6 ± 0.1c | |||
| cyanidin- -3-O-xyloside | ND | FP | ND | ND | 4.1 ± 0.1e | 5.0 ± 0.1c | 7.2 ± 0.1b | 10.1 ± 0.1a | |
| FPM | 3.3 ± 0.1g | 3.8 ± 0.1f | 4.1 ± 0.e1 | 4.6 ± 0.1d | 4.4 ± 0.1d | 5.2 ± 0.1c | |||
| FPI | 3.1 ± 0.1g | 3.6 ± 0.1f | 3.7 ± 0.1f | 3.6 ± 0.1f | 4.5 ± 0.1d | 5.4 ± 0.1c | |||
| SUM | ND | FP | 21.2 ± 0.9n | 47.4 ± 1.8m | 104 ± 4i | 163 ± 6f | 240 ± 9e | 344 ± 13b | |
| FPM | 73.5 ± 2.8l | 84.5 ± 3.3k | 125 ± 5h | 150 ± 6g | 252 ± 11d | 348 ± 15a | |||
| FPI | 78.8 ± 3.1l | 90.6 ± 3.5j | 124 ± 5h | 155 ± 6g | 234 ± 10e | 323 ± 14c | |||
| Flavan-3-ols | B-type procyjanidin dimer | 10.1 ± 0.1o | FP | 11.0 ± 0.2n | 11.5 ± 0.2m | 30.2 ± 0.6g | 37.1 ± 0.7f | 36.3 ± 0.7f | 38.8 ± 0.8e |
| FPM | 20.6 ± 0.4k | 23.7 ± 0.5j | 39.4 ± 0.8e | 49.6 ± 1.0d | 55.5 ± 1.1c | 66.6 ± 1.3a | |||
| FPI | 19.1 ± 0.4l | 22.0 ± 0.4j | 27.7 ± 0.6i | 29.6 ± 0.6g | 49.9 ± 1.0d | 59.9 ± 1.2b | |||
| Epigallocatechin x | 12.3 ± 0.1k | FP | 25.1 ± 0.5j | 32.2 ± 0.6g | 37.6 ± 0.8e | 39.9 ± 0.8d | 43.4 ± 0.9c | 44.9 ± 0.9b | |
| FPM | 23.7 ± 0.5j | 27.2 ± 0.5i | 39.5 ± 0.8d | 46.6 ± 0.9b | 43.8 ± 0.9c | 54.5 ± 1.1a | |||
| FPI | 30.4 ± 0.6h | 36.2 ± 0.7f | 44.6 ± 0.9b | 44.4 ± 0.9b | 45.1 ± 0.9b | 55.8 ± 1.1a | |||
| B-type procyjanidin dimer | 16.0 ± 0.2o | FP | 31.8 ± 0.6g | 30.6 ± 0.6g | 41.7 ± 0.8d | 49.7 ± 1.0c | 55.8 ± 1.1b | 53.8 ± 1.1a | |
| FPM | 14.9 ± 0.3n | 17.1 ± 0.3m | 29.5 ± 0.6h | 29.7 ± 0.6h | 31.4 ± 0.6g | 35.7 ± 0.7e | |||
| FPI | 21.2 ± 0.4l | 23.2 ± 0.5k | 26.5 ± 0.5j | 28.0 ± 0.6i | 29.3 ± 0.6h | 33.6 ± 0.7f | |||
| (+)-Catechin | 4.0 ± 0.1m | FP | 12.0 ± 0.2l | 31.4 ± 0.6h | 36.1 ± 0.7f | 48.5 ± 1.0d | 52.9 ± 1.1c | 59.3 ± 1.2b | |
| FPM | 17.3 ± 0.3k | 19.9 ± 0.4j | 37.1 ± 0.7f | 42.0 ± 0.8e | 48.6 ± 1.0d | 58.3 ± 1.2b | |||
| FPI | 19.8 ± 0.4j | 22.8 ± 0.5i | 34.2 ± 0.7g | 42.7 ± 0.9e | 52.8 ± 1.1c | 63.3 ± 1.3a | |||
| (−)-Epicatechin | 9.4 ± 0.3k | FP | 11.3 ± 0.2j | 14.7 ± 0.3h | 15.3 ± 0.3g | 19.9 ± 0.4d | 21.5 ± 0.4c | 28.0 ± 0.6a | |
| FPM | 13.9 ± 0.3i | 15.9 ± 0.3 | 18.4 ± 0.4e | 18.1 ± 0.4e | 24.8 ± 0.5b | 27.8 ± 0.6a | |||
| FPI | 13.3 ± 0.3i | 15.3 ± 0.3 | 17.7 ± 0.4f | 18.3 ± 0.4e | 16.4 ± 0.3 | 19.7 ± 0.4d | |||
| SUM | 51.8 ± 1.0n | FP | 91.3 ± 9.6m | 120 ± 10k | 161 ± 10i | 195 ± 12f | 210 ± 14d | 225 ± 12c | |
| FPM | 90.3 ± 4.1m | 104 ± 5l | 164 ± 9h | 186 ± 13g | 204 ± 13e | 243 ± 16a | |||
| FPI | 104 ± 6l | 120 ± 8k | 151 ± 10j | 163 ± 11h | 194 ± 15f | 232 ± 19b | |||
| Phenolic acids | protokatechumic acid | 0.9 ± 0.1j | FP | 3.1 ± 0.1h | 4.9 ± 0.1g | 6.2 ± 0.1e | 8.3 ± 0.1c | 9.5 ± 0.1b | 10.5 ± 0.1a |
| FPM | 0.9 ± 0.1j | 1.1 ± 0.1i | 4.8 ± 0.0g | 0.7 ± 0.0j | 1.0 ± 0.1i | 1.2 ± 0.1i | |||
| FPI | 6.9 ± 0.1e | 7.9 ± 0.1c | 5.8 ± 0.1f | 5.5 ± 0.1f | 5.6 ± 0.1f | 6.8 ± 0.1d | |||
| vanilic acid | 2.5 ± 0.1d | FP | 3.5 ± 0.1b | 3.3 ± 0.1b | 3.6 ± 0.1a | 3.8 ± 0.1a | 3.3 ± 0.1b | 3.6 ± 0.1a | |
| FPM | 2.0 ± 0.1d | 2.3 ± 0.1d | 2.1 ± 0.0d | 2.0 ± 0.0d | 2.1 ± 0.1d | 2.5 ± 0.1c | |||
| FPI | 1.9 ± 0.1d | 2.1 ± 0.1d | 2.3 ± 0.1d | 2.3 ± 0.1d | 2.3 ± 0.1d | 2.7 ± 0.1c | |||
| caffeic acid | 0.9 ± 0.1g | FP | 3.1 ± 0.1f | 3.5 ± 0.1f | 4.2 ± 0.1e | 4.6 ± 0.1e | 5.2 ± 0.1d | 5.7 ± 0.1d | |
| FPM | 3.1 ± 0.1f | 3.6 ± 0.1f | 7.3 ± 0.1b | 7.3 ± 0.1b | 6.9 ± 0.1c | 8.3 ± 0.1a | |||
| FPI | 5.1 ± 0.1d | 5.9 ± 0.1d | 7.2 ± 0.1b | 7.3 ± 0.1b | 7.2 ± 0.1b | 8.6 ± 0.1a | |||
| 3-O- -caffequinic acid | 0.2 ± 0.1m | FP | 60.1 ± 1.0g | 72.2 ± 1.0g | 86.6 ± 1.1e | 103 ± 1d | 106 ± 1c | 110 ± 1a | |
| FPM | 72.7 ± 0.7l | 83.6 ± 0.8k | 103 ± 1i | 109 ± 1g | 116 ± 0e | 140 ± 3b | |||
| FPI | 89.1 ± 0.8j | 102 ± 1i | 106 ± 1h | 110 ± 0g | 114 ± 0f | 137 ± 1c | |||
| ferulic acid | 5.0 ± 0.1b | FP | 5.2 ± 0.1b | 5.2 ± 0.1b | 5.7 ± 0.1b | 6.1 ± 0.1a | 5.4 ± 0.1.0b | 5.9 ± 0.1a | |
| FPM | 3.1 ± 0.1f | 3.6 ± 0.1d | 3.6 ± 0.0d | 3.5 ± 0.0e | 3.7 ± 0.1d | 4.4 ± 0.1c | |||
| FPI | 3.1 ± 0.1f | 3.4 ± 0.1e | 3.4 ± 0.1e | 3.5 ± 0.1e | 3.3 ± 0.1e | 3.9 ± 0.1d | |||
| 5-O- -caffequinic acid | 0.4 ± 0.1 | FP | 43.3 ± 0.4k | 49.5 ± 0.4j | 76.6 ± 0.7g | 102 ± 1e | 115 ± 1c | 127 ± 1b | |
| FPM | 58.4 ± 0.5i | 67.2 ± 0.6h | 102 ± 1e | 104 ± 1e | 115 ± 1c | 138 ± 1a | |||
| FPI | 75.2 ± 0.7g | 86.5 ± 0.8f | 104 ± 1e | 107 ± 1d | 113 ± 1c | 136 ± 1a | |||
| 3-p-coumaroylquinic acid | 0.8 ± 0.1h | FP | 3.4 ± 0.1g | 4.0 ± 0.1g | 6.6 ± 0.1g | 17.3 ± 0.2e | 39.2 ± 0.4b | 43.2 ± 0.4a | |
| FPM | 15.4 ± 0.1f | 17.7 ± 0.2e | 23.0 ± 0.2d | 23.2 ± 0.2d | 23.6 ± 0.2d | 28.3 ± 0.3c | |||
| FPI | 17.4 ± 0.2e | 20.0 ± 0.2d | 21.6 ± 0.2d | 23.5 ± 0.2d | 25.2 ± 0.2d | 30.2 ± 0.3c | |||
| 4-O- -caffequinic acid | 0.2 ± 0.1j | FP | 2.2 ± 0.1i | 4.9 ± 0.1g | 7.3 ± 0.1e | 12.1 ± 0.1b | 14.1 ± 0.1a | 15.4 ± 0.1a | |
| FPM | 3.5 ± 0.1i | 4.0 ± 0.1g | 5.8 ± 0.1f | 7.2 ± 0.1e | 8.3 ± 0.1d | 10.0 ± 0.1c | |||
| FPI | 3.3 ± 0.1i | 3.8 ± 0.1h | 7.7 ± 0.1e | 8.1 ± 0.1d | 8.4 ± 0.1d | 10.1 ± 0.1c | |||
| SUM | 10.9 ± 0.2l | FP | 124 ± 2o | 148 ± 3n | 197 ± 4k | 259 ± 5g | 298 ± 6c | 322 ± 6b | |
| FPM | 159 ± 3m | 183 ± 4l | 253 ± 5h | 257 ± 5g | 277 ± 6e | 333 ± 7a | |||
| FPI | 202 ± 4j | 232 ± 5i | 259 ± 5g | 268 ± 5f | 280 ± 6d | 336 ± 7a | |||
| Flavonols | Kaempferol-3-O-galactoside | 3.1 ± 0.1j | FP | 3.3 ± 0.1j | 7.3 ± 0.1h | 12.6 ± 0.3f | 21.5 ± 0.4c | 26.3 ± 0.5b | 31.5 ± 0.6a |
| FPM | 6.5 ± 0.1i | 7.4 ± 0.1h | 10.9 ± 0.2g | 14.0 ± 0.3f | 16.3 ± 0.3e | 19.2 ± 0.4d | |||
| FPI | 7.5 ± 0.1h | 8.4 ± 0.2h | 10.3 ± 0.2g | 13.3 ± 0.3f | 13.9 ± 0.3f | 16.7 ± 0.3e | |||
| Quercetin-3-O-arabinoglucoside | 4.2 ± 0.1j | FP | 1.8 ± 0.1f | 2.3 ± 0.1e | 2.6 ± 0.1d | 2.9 ± 0.1d | 3.6 ± 0.1b | 4.0 ± 0.0a | |
| FPM | 2.5 ± 0.1e | 2.9 ± 0.1d | 3.4 ± 0.1c | 3.4 ± 0.1c | 3.7 ± 0.1b | 4.2 ± 0.1a | |||
| FPI | 2.5 ± 0.1e | 2.9 ± 0.1d | 3.3 ± 0.1c | 3.3 ± 0.1c | 3.7 ± 0.1b | 4.1 ± 0.0a | |||
| Kaempferol-3-O-glucoside | 32.0 ± 0.3g | FP | 34.0 ± 0.7f | 36.0 ± 0.7e | 37.9 ± 0.8d | 42.6 ± 0.9c | 48.1 ± 1.1b | 51.0 ± 1.1b | |
| FPM | 34.3 ± 0.7f | 39.1 ± 0.8d | 40.8 ± 0.8c | 48.1 ± 1.0b | 49.0 ± 1.0b | 53.0 ± 1.1a | |||
| FPI | 33.5 ± 0.7f | 35.3 ± 0.7e | 37.1 ± 0.7d | 41.8 ± 0.8c | 41.9 ± 0.8c | 50.3 ± 1.0b | |||
| Quercetin-3-O-rutinoside | 3.1 ± 0.1h | FP | 3.3 ± 0.1g | 7.3 ± 0.1f | 10.6 ± 0.1d | 11.5 ± 0.1c | 12.3 ± 0.1b | 15.2 ± 0.1a | |
| FPM | 6.0 ± 0.1e | 7.1 ± 0.1d | 9.7 ± 0.1b | 9.4 ± 0.1c | 12.2 ± 0.1a | 13.6 ± 0.1a | |||
| FPI | 6.0 ± 0.1e | 6.9 ± 0.1d | 9.5 ± 0.1c | 11.8 ± 0.1a | 12.0 ± 0.1a | 13.4 ± 0.1a | |||
| Quercetin-3-O-robinobioside | 1.1 ± 0.1j | FP | 2.5 ± 0.1i | 3.5 ± 0.1g | 3.7 ± 0.1g | 8.1 ± 0.1d | 10.0 ± 0.1b | 11.3 ± 0.1a | |
| FPM | 3.1 ± 0.1h | 3.6 ± 0.1g | 6.1 ± 0.1f | 6.1 ± 0.1f | 8.4 ± 0.1d | 9.4 ± 0.1c | |||
| FPI | 3.0 ± 0.1h | 3.5 ± 0.1g | 6.1 ± 0.1f | 5.9 ± 0.1f | 7.4 ± 0.1e | 8.3 ± 0.1d | |||
| Quercetin-3-O-galactoside | 4.2 ± 0.1h | FP | 16.3 ± 0.1i | 23.8 ± 0.2h | 29.1 ± 0.3g | 41.4 ± 0.4e | 50.7 ± 0.5b | 54.6 ± 0.6a | |
| FPM | 29.7 ± 0.3g | 34.1 ± 0.3f | 34.3 ± 0.3f | 41.6 ± 0.4e | 50.8 ± 0.5d | 56.8 ± 0.5c | |||
| FPI | 31.0 ± 0.3f | 35.6 ± 0.3f | 34.4 ± 0.3f | 51.7 ± 0.5d | 59.9 ± 0.5b | 67.1 ± 0.6a | |||
| Quercetin-3-O-glucoside | ND | FP | 4.1 ± 0.1j | 4.7 ± 0.1i | 6.1 ± 0.1g | 7.0 ± 0.1e | 7.8 ± 0.1d | 8.9 ± 0.1b | |
| FPM | 3.6 ± 0.1k | 4.1 ± 0.1j | 5.5 ± 0.1h | 6.7 ± 0.1f | 7.6 ± 0.1d | 8.6 ± 0.1b | |||
| FPI | 4.1 ± 0.1j | 4.7 ± 0.1i | 6.6 ± 0.1f | 7.9 ± 0.1d | 8.1 ± 0.1c | 9.1 ± 0.1a | |||
| Quercetin-3-O-arabinoside | 1.8 ± 0.01g | FP | 1.7 ± 0.01g | 2.2 ± 0.01f | 3.4 ± 0.1d | 3.8 ± 0.1c | 4.3 ± 0.1b | 5.6 ± 0.1a | |
| FPM | 1.9 ± 0.01g | 2.1 ± 0.01f | 2.2 ± 0.01f | 2.6 ± 0.1e | 2.7 ± 0.1e | 2.9 ± 0.1e | |||
| FPI | 1.8 ± 0.01g | 1.9 ± 0.01g | 2.0 ± 0.01f | 2.3 ± 0.1f | 2.3 ± 0.1f | 2.8 ± 0.1e | |||
| Quercetin-3-O-xyloside | ND | FP | 2.4 ± 0.1i | 3.8 ± 0.1h | 4.6 ± 0.1f | 5.5 ± 0.1d | 6.8 ± 0.1b | 7.0 ± 0.a | |
| FPM | 3.9 ± 0.1h | 4.5 ± 0.1g | 4.8 ± 0.1f | 5.2 ± 0.1e | 6.1 ± 0.1c | 6.9 ± 0.1b | |||
| FPI | 4.4 ± 0.1g | 5.1 ± 0.1e | 5.2 ± 0.1e | 5.5 ± 0.1d | 5.9 ± 0.1c | 6.6 ± 0.1b | |||
| Quercetin-deoxyhexo-hexoside | ND | FP | 0.5 ± 0.1c | 0.5 ± 0.1c | 0.6 ± 0.1b | 0.8 ± 0.1a | 0.9 ± 0.1a | 1.0 ± 0.0a | |
| FPM | 0.3 ± 0.1d | 0.3 ± 0.1d | 0.4 ± 0.1c | 0.5 ± 0.1c | 0.8 ± 0.1a | 0.9 ± 0.1a | |||
| FPI | 0.4 ± 0.1d | 0.4 ± 0.1d | 0.5 ± 0.1kc | 0.7 ± 0.1b | 0.7 ± 0.1b | 0.8 ± 0.0a | |||
| SUM | 43.5 ± 1.3o | FP | 69.9 ± 1.4n | 88.2 ± 1.8m | 106 ± 2 | 133 ± 3h | 155 ± 3e | 171 ± 3c | |
| FPM | 91.9 ± 1.8l | 105 ± 2k | 118 ± 2i | 138 ± 3g | 158 ± 3d | 176 ± 4b | |||
| FPI | 94.2 ± 1.9l | 105 ± 2k | 115 ± 2j | 144 ± 3f | 156 ± 3e | 179 ± 4a | |||
| Sum of phenols | 106 ± 3o | FP | 306 ± 14n | 404 ± 18m | 568 ± 25j | 751 ± 34f | 904 ± 43c | 1062 ± 56a | |
| FPM | 415 ± 20m | 477 ± 23l | 661 ±3 2h | 731 ± 35g | 891 ± 49d | 1100 ± 66b | |||
| FPI | 479 ± 23l | 547 ± 27k | 649 ± 32i | 730 ± 36g | 863 ± 47e | 1071 ± 62a | |||
| Compounds | Bread Sample | Relative Accessibility Index | |||||||
|---|---|---|---|---|---|---|---|---|---|
| BRC | BRP3 | BRM3 | BRI3 | BRC | BRP3 | BRM3 | BRI3 | ||
| Anthocyanins | ANT1 | - | 503 ± 2c 1 | 662 ± 2b | 695 ± 2a | - | 5.7 | 6.2 | 6.4 |
| ANT2 | - | 34.8 ± 0.1b | 45.8 ± 0.2a | 48.1 ± 0.2a | - | 4.0 | 4.4 | 5.4 | |
| ANT3 | - | 17.3 ± 0.1b | 22.7 ± 0.1a | 23.8 ± 0.1a | - | 4.8 | 5.6 | 5.7 | |
| ANT4 | - | 23.0 ± 0.1b | 30.3 ± 0.1a | 31.8 ± 0.1a | - | 5.7 | 7.4 | 8.5 | |
| SUM | - | 578 ± 239c | 761 ± 315b | 799 ± 202a | - | 5.5 | 6.1 | 6.4 | |
| Flavan-3-ols | F301 | 16.7 ± 0.3d | 27.2 ± 0.5c | 40.2 ± 0.8a | 36.6 ± 0.7b | 1.7 | 0.9 | 1.0 | 1.3 |
| F302 | 14.2 ± 0.8c | 30.5 ± 0.6b | 42.7 ± 0.8a | 43.3 ± 0.8a | 1.1 | 0.8 | 1.1 | 1.0 | |
| F303 | 13.1 ± 0.6d | 5.4 ± 0.1c | 30.1 ± 0.6a | 23.4 ± 0.7b | 0.1 | 0.1 | 1.0 | 0.9 | |
| F304 | 4.3 ± 0.2d | 14.4 ± 0.2c | 37.8 ± 0.7b | 44.2 ± 0.8a | 1.7 | 0.8 | 1.0 | 1.0 | |
| F305 | 10.2 ± 0.2c | 6.1 ± 0.1d | 18.7 ± 0.3b | 22.8 ± 0.4a | 1.1 | 0.4 | 1.0 | 1.3 | |
| SUM | 58.5 ± 1.1c | 83.7±1.2b | 169 ± 3a | 170 ± 3a | 1.2 | 0.6 | 1.0 | 1.2 | |
| Phenolic acid | PA1 | 2.5 ± 0.1a | 1.6 ± 0.1b | 2.5 ± 0.1a | 2.7 ± 0.1a | 2.8 | 0.3 | 0.5 | 0.5 |
| PA2 | 5.1 ± 0.1a | 0.9 ± 0.1b | 1.5 ± 0.1b | 1.6 ± 0.1b | 2.0 | 0.3 | 0.7 | 0.7 | |
| PA3 | 12.3 ± 0.1a | 5.5 ± 0.1d | 8.8 ± 0.1c | 9.2 ± 0.1b | 15.0 | 0.8 | 0.4 | 0.4 | |
| PA4 | 5.5 ± 0.1a | 2.7 ± 0.1c | 4.3 ± 0.1b | 4.6 ± 0.1b | 5.9 | 0.7 | 0.6 | 0.6 | |
| PA5 | 2.7 ± 0.1c | 121 ± 1b | 221 ± 1a | 219 ± 1a | 17.0 | 1.0 | 2.1 | 2.1 | |
| PA6 | 5.5 ± 0.1a | 2.7 ± 0.1c | 4.3 ± 0.1b | 4.5 ± 0.1b | 1.1 | 0.5 | 1.2 | 1.3 | |
| PA7 | 0.7 ± 0.1c | 63.2 ± 0.3b | 99.8 ± 0.4a | 95.7 ± 0.4a | 1.7 | 0.8 | 1.0 | 0.9 | |
| PA8 | 1.4 ± 0.1c | 6.1 ± 0.1b | 9.7 ± 0.1a | 10.1 ± 0.1a | 7.5 | 0.8 | 1.7 | 1.3 | |
| SUM | 35.8 ± 3.7d | 204 ± 21c | 352 ± 47a | 347 ± 39b | 3.3 | 0.9 | 1.4 | 1.3 | |
| Flavonols | FL1 | 4.1 ± 0.1b | 22.5 ± 0.1a | 24.7 ± 0.1a | 23.6 ± 0.1a | 1.3 | 1.8 | 2.3 | 2.3 |
| FL2 | 5.1 ± 0.1b | 2.3 ± 0.2a | 3.1 ± 0.2a | 3.2 ± 0.2a | 1.2 | 0.9 | 0.9 | 1.0 | |
| FL3 | 28.7 ± 0.1c | 38.8 ± 0.2b | 52.7 ± 0.2a | 40.7 ± 0.2b | 0.9 | 1.0 | 1.3 | 1.1 | |
| FL4 | 7.0 ± 0.1b | 9.6 ± 0.1a | 10.5 ± 0.1a | 10.1 ± 0.1a | 2.3 | 0.9 | 1.1 | 1.1 | |
| FL5 | 5.1 ± 0.1b | 5.2 ± 0.1a | 5.7 ± 0.1a | 5.4 ± 0.1a | 4.4 | 1.4 | 0.9 | 0.9 | |
| FL6 | - | 40.3 ± 0.2a | 44.3 ± 0.2a | 42.3 ± 0.2a | - | 1.4 | 1.3 | 1.2 | |
| FL7 | - | 8.1 ± 0.1a | 8.9 ± 0.1a | 8.5 ± 0.1a | - | 1.3 | 1.6 | 1.3 | |
| FL8 | 4.9 ± 0.1d | 9.2 ± 0.1a | 6.4 ± 0.1b | 5.1 ± 0.1c | 2.7 | 2.7 | 2.9 | 2.6 | |
| FL9 | - | 12.6 ± 0.1a | 13.8 ± 0.1a | 13.2 ± 0.1a | - | 2.7 | 2.9 | 2.6 | |
| FL10 | - | 5.4 ± 0.1a | 5.9 ± 0.1a | 5.6 ± 0.1a | - | 2.1 | 2.5 | 2.3 | |
| SUM | 54.9 ± 9.1d | 153 ± 15c | 176 ± 18a | 157 ± 15b | 1.2 | 1.4 | 1.5 | 1.4 | |
| Sum of polyphenols | 144 ± 25d | 1047 ± 217c | 1492 ± 270b | 1506 ± 292a | 1.4 | 1.6 | 1.9 | 2.1 | |
| Antioxidant Activity [µmol TE/g ds.] | BRC | Form of Supplement | BR1 | BR2 | BR3 | BR4 | BR5 | BR6 |
|---|---|---|---|---|---|---|---|---|
| FRAP | 21.0 ± 0.42g 1 | FP | 22.30 ± 0.45f | 22.60 ± 0.45e | 24.40 ± 0.37d | 26.70 ± 0.53c | 29.50 ± 0.59b | 34.80 ± 0.70a |
| FPM | 22.70 ± 0.45e | 22.90 ± 0.46e | 26.50 ± 0.53d | 30.20 ± 0.60c | 32.30 ± 0.65b | 39.00 ± 0.78a | ||
| FPI | 22.32 ± 0.33f | 23.06 ± 0.40e | 25.48 ± 0.45d | 28.76 ± 0.58c | 32.60 ± 0.65b | 39.22 ± 0.78a | ||
| ABTS | 13.09 ± 0.26g | FP | 13.93 ± 0.27f | 14.14 ± 0.49c | 15.24 ± 0.40e | 16.68 ± 0.43d | 18.43 ± 0.79b | 21.73 ± 0.80a |
| FPM | 14.19 ± 0.46d | 14.32 ± 0.32f | 16.58 ± 0.34e | 18.84 ± 0.68c | 20.27 ± 0.73b | 24.39 ± 0.87a | ||
| FPI | 13.95 ± 0.35d | 14.41 ± 0.35d | 15.93 ± 0.33e | 17.98 ± 0.63c | 20.40 ± 0.73b | 24.50 ± 0.87a | ||
| DPPH | 3.04 ± 0.02k | FP | 3.34 ± 0.12j | 3.58 ± 0.05i | 3.62 ± 0.09i | 4.27 ± 0.17g | 4.50 ± 0.20f | 5.46 ± 0.17d |
| FPM | 3.70 ± 0.09i | 3.66 ± 0.07i | 6.94 ± 0.21b | 4.51 ± 0.12f | 5.14 ± 0.13e | 8.51 ± 0.12a | ||
| FPI | 3.57 ± 0.15i | 3.62 ± 0.17i | 4.04 ± 0.08h | 4.60 ± 0.19f | 5.12 ± 0.09e | 5.98 ± 0.10c |
| In Vitro Potency | Extract | Bread Sample | Relative Accessibility Index | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| BRC | BRP3 | BRM3 | BRI3 | BRC | BRP3 | BRM3 | BRI3 | |||
| Antioxidant activity [µmol TE/g d. s.] | ABTS | EBD | 13.09 ± 0.26c 1 | 15.24 ± 0.40a | 16.58 ± 0.34b | 15.93 ± 0.33b | 2.20 | 2.48 | 3.15 | 2.72 |
| PBE | 28.78 ± 0.24c | 37.44 ± 0.31a | 52.24 ± 0.24b | 43.37 ± 0.25b | ||||||
| DPPH | EBD | 3.04 ± 0.02d | 3.62 ± 0.09c | 6.94 ± 0.21a | 4.04 ± 0.08b | 1.70 | 2.00 | 2.78 | 2.13 | |
| PBE | 5.17 ± 0.39d | 7.23 ± 0.21c | 19.29 ± 0.72a | 8.60 ± 0.28b | ||||||
| FRAP | EBD | 21.00 ± 0.42c | 24.39 ± 0.37d | 26.53 ± 0.53a | 25.48 ± 0.45b | 0.71 | 1.25 | 1.84 | 2.36 | |
| PBE | 15.00 ± 0.71d | 30.48 ± 0.41c | 48.70 ± 0.51b | 52.99 ± 0.39a | ||||||
| Pro-health potency | Inhibition of LOX activity [kIU/g ds] | EBD | 1257 ± 30b | 1549 ± 2a | 121 ± 2c | 1116 ± 2d | 0.56 | 2.04 | 1.25 | 1.37 |
| PBF | 70.00 ± 0.14c | 3168 ± 5d | 147 ± 2b | 1530 ± 3a | ||||||
| Inhibition of COX-1 activity [kIU/g ds] | EBD | 4.22 ± 0.84b | 8.32 ± 0.66a | - | - | 0.37 | - | - | - | |
| PBF | 1.56 ± 0.11a | - | - | - | ||||||
| Inhibition of COX-2 activity [kIU/g ds] | EBD | 7.75 ± 0.55d | 10.71 ± 1.14c | 19.33 ± 0.39a | 15.96 ± 0.32b | 0.91 | 0.36 | 0.43 | 0.54 | |
| PBF | 7.06 ± 1.41c | 3.85 ± 0.77d | 8.29 ± 0.66b | 8.57 ± 0.71a | ||||||
| Inhibition of AChE activity [IU/g ds] | EBD | 23.54 ± 0.47b | 20.73 ± 0.41d | 21.90 ± 0.44c | 24.50 ± 0.49a | - | - | - | - | |
| PBF | - | - | - | - | ||||||
| Sample | Inhibition of α-Amylase Activity [U/g] | Inhibition of α-Glucosidase Activity [U/g] | Inhibition of Pancreatic Lipase Activity [U/g] |
|---|---|---|---|
| BRC | 4.58 ± 0.09d 1 | 0.04 ± 0.01d | 1.20 ± 0.02c |
| BRP3 | 46.62 ± 0.93a | 4.04 ± 0.08a | 1.03 ± 0.02d |
| BRM3 | 41.58 ± 0.83b | 2.32 ± 0.05b | 1.27 ± 0.03b |
| BRI3 | 16.09 ± 0.32c | 2.04 ± 0.04c | 1.43 ± 0.03a |
| Bread | Free Reducing Sugars [mg/g] | Released Sugar [mg/g] | Relative Digestibility of Starch | Free Amino Acids and Peptides [mg/g] | Released Amino Acids and Peptides [mg/g] | Relative Digestibility of Proteins |
|---|---|---|---|---|---|---|
| BRC | 54.7 ± 1.1d 1 | 473 ± 9a | 100 ± 2a | 4.4 ± 0.1c | 82.3 ± 1.6b | 100 ± 2a |
| BRP3 | 248 ± 5a | 354 ± 7d | 75.0 ± 1.5d | 6.9 ± 0.1a | 105 ± 2a | 127 ± 2b |
| BRM3 | 158 ± 3b | 371 ± 7c | 78.4 ± 1.6c | 4.2 ± 0.1d | 68.5 ± 1.4d | 83.3 ± 1.7d |
| BRI3 | 100 ± 2c | 442 ± 8b | 93.4 ± 1.9b | 4.8 ± 0.1b | 70.8 ± 1.4c | 86.0 ± 1.7c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lachowicz, S.; Świeca, M.; Pejcz, E. Improvement of Health-Promoting Functionality of Rye Bread by Fortification with Free and Microencapsulated Powders from Amelanchier alnifolia Nutt. Antioxidants 2020, 9, 614. https://doi.org/10.3390/antiox9070614
Lachowicz S, Świeca M, Pejcz E. Improvement of Health-Promoting Functionality of Rye Bread by Fortification with Free and Microencapsulated Powders from Amelanchier alnifolia Nutt. Antioxidants. 2020; 9(7):614. https://doi.org/10.3390/antiox9070614
Chicago/Turabian StyleLachowicz, Sabina, Michał Świeca, and Ewa Pejcz. 2020. "Improvement of Health-Promoting Functionality of Rye Bread by Fortification with Free and Microencapsulated Powders from Amelanchier alnifolia Nutt" Antioxidants 9, no. 7: 614. https://doi.org/10.3390/antiox9070614
APA StyleLachowicz, S., Świeca, M., & Pejcz, E. (2020). Improvement of Health-Promoting Functionality of Rye Bread by Fortification with Free and Microencapsulated Powders from Amelanchier alnifolia Nutt. Antioxidants, 9(7), 614. https://doi.org/10.3390/antiox9070614