Sunflower (Helianthus annuus L.) Plants at Various Growth Stages Subjected to Extraction—Comparison of the Antioxidant Activity and Phenolic Profile

 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Chemicals
2.3. Extraction
2.4. Determination of the Total Phenolic Content
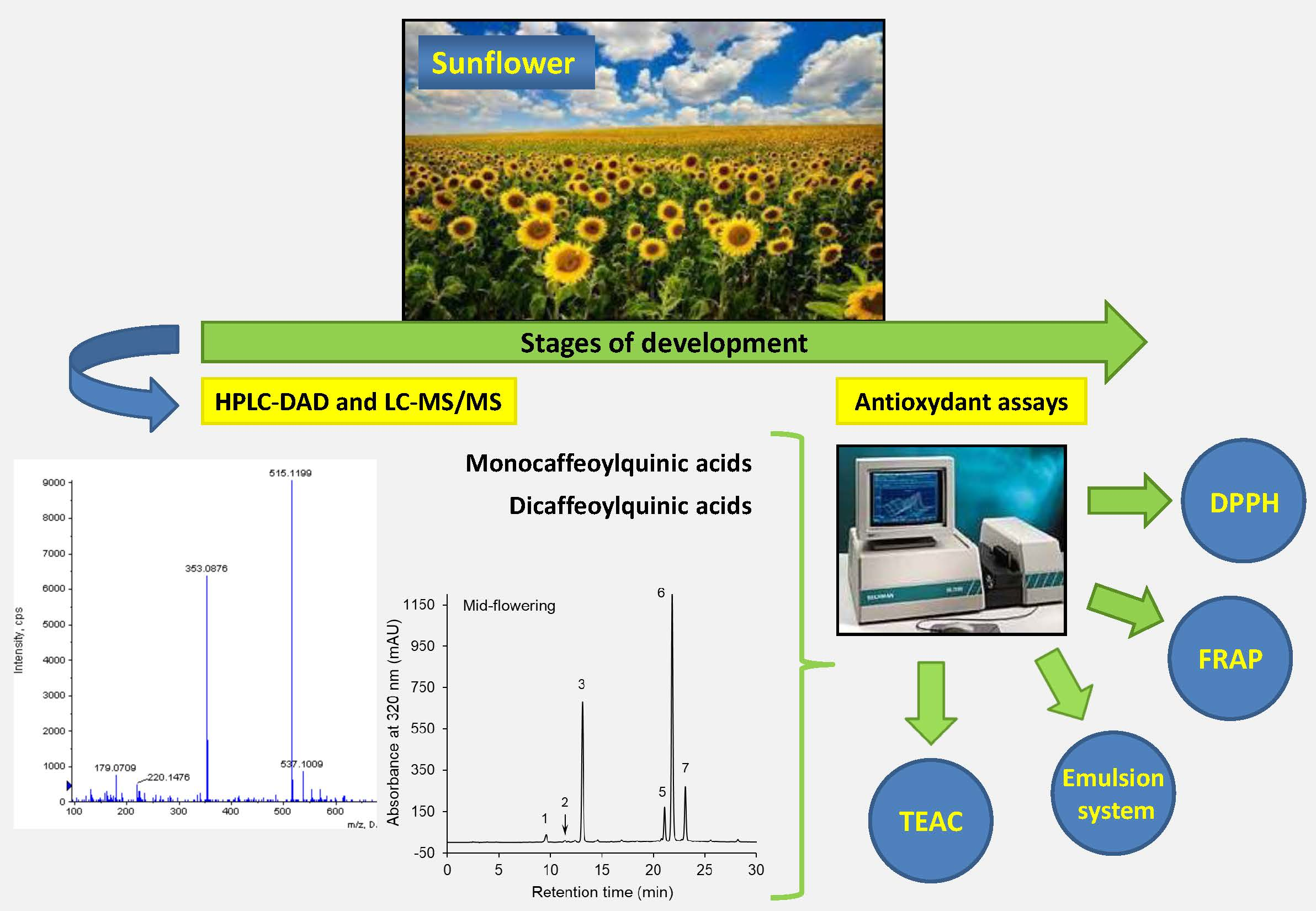
2.5. Identification and Quantification of the Phenolic Compounds
2.6. Determination of the Trolox Equivalent Antioxidant Capacity
2.7. Determination of the Ferric-Reducing Antioxidant Power
2.8. Determination of the DPPH Radical Scavenging Activity
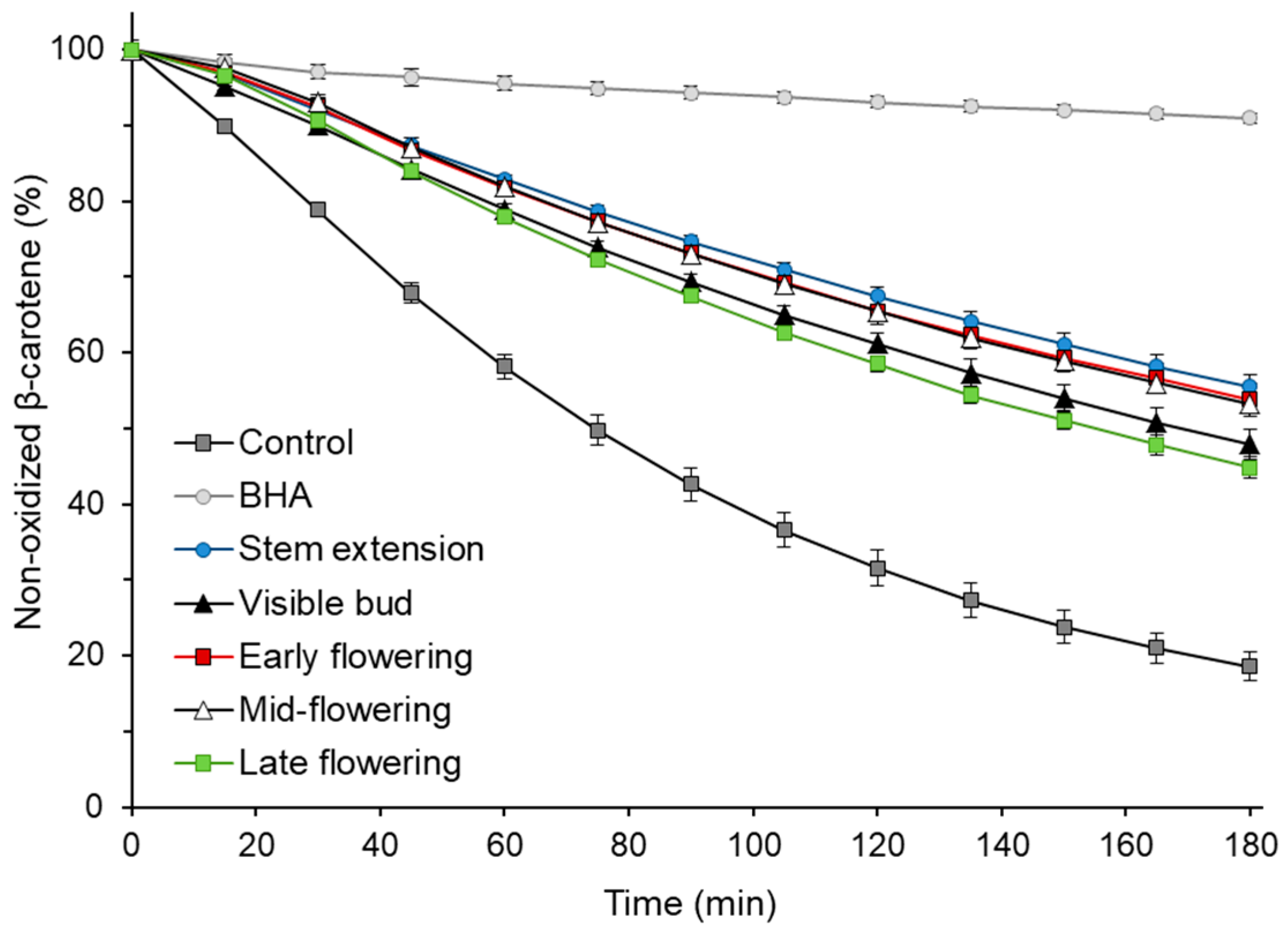
2.9. β-Carotene–Linoleic Acid Emulsion Oxidation
2.10. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Collin, F. Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef]
- Liu, Z.; Zhou, T.; Ziegler, A.C.; Dimitrion, P.; Zuo, L. Oxidative stress in neurodegenerative diseases: From molecular mechanisms to clinical applications. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Maher, P. The potential of flavonoids for the treatment of neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 3056. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Zamora, R.; Hidalgo, F.J. The triple defensive barrier of phenolic compounds against the lipid oxidation-induced damage in food products. Trends Food Sci. Tech. 2016, 54, 165–174. [Google Scholar] [CrossRef]
- Liović, N.; Bošković, P.; Drvenica, I.; Jambrak, A.R.; Dropulić, A.M.; Krešić, G.; Nedović, V.; Bugarski, B.; Zorić, Z.; Pedisić, S.; et al. Phenolic extracts from Vaccinium corymbosum L. loaded in microemulsions and liposomes as enhancers of olive oil oxidative stability. Pol. J. Food Nutr. Sci. 2019, 69, 23–33. [Google Scholar] [CrossRef]
- Wildermuth, S.R.; Young, E.E.; Were, L.M. Chlorogenic acid oxidation and its reaction with sunflower proteins to form green-colored complexes. Compr. Rev. Food Sci. Food Saf. 2016, 15, 829–843. [Google Scholar] [CrossRef]
- National Sunflower Association of Canada. Sunflower Production Guide. Available online: http://www.canadasunflower.com/production/sunflower-production-guide/ (accessed on 11 May 2020).
- Demirel, M.; Bolat, D.; Celik, S.; Bakici, Y.; Eratak, S. Determination of fermentation and digestibility characteristics of corn, sunflower and combination of corn and sunflower silages. J. Anim. Vet. Adv. 2008, 7, 707–711. [Google Scholar]
- Peiretti, P.G.; Meineri, G. Evolution of chemical composition, nutritive value, and fatty acid content of sunflower (Helianthus annuus L.) during the growth cycle. J. Anim. Vet. Adv. 2010, 9, 112–117. [Google Scholar] [CrossRef]
- Konca, Y.; Beyzi, S.B.; Ayasan, T.; Kaliber, M.; Kiraz, A.B. The effects of freezing and supplementation of molasses and inoculants on chemical and nutritional composition of sunflower silage. Asian Australas. J. Anim. Sci. 2016, 29, 965–970. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Duke, J.A.; Bogenschutz-Godwin, M.J.; duCellier, J.; Duke, P.A.K. Handbook of Medicinal Herbs, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2002; p. 707. [Google Scholar]
- De Leonardis, A.; Macciola, V.; Di Domenico, N. A first pilot study to produce a food antioxidant from sunflower seed shells (Helianthus annuus). Eur. J. Lipid Sci. Tech. 2005, 107, 220–227. [Google Scholar] [CrossRef]
- Giada, M.D.L.R.; Mancini-Filho, J. Antioxidant capacity of the striped sunflower (Helianthus annuus L.) seed extracts evaluated by three in vitro methods. Int. J. Food Sci. Nutr. 2009, 60, 395–401. [Google Scholar] [CrossRef]
- Zoumpoulakis, P.; Sinanoglou, V.J.; Siapi, E.; Heropoulos, G.; Proestos, C. Evaluating modern techniques for the extraction and characterisation of sunflower (Hellianthus annus L.) seeds phenolics. Antioxidants 2017, 6, 46. [Google Scholar] [CrossRef]
- Sarkis, J.R.; Côrrea, A.P.F.; Michel, I.; Brandeli, A.; Tessaro, I.C.; Marczak, L.D.F. Evaluation of the phenolic content and antioxidant activity of different seed and nut cakes from the edible oil industry. J. Am. Oil Chem. Soc. 2014, 91, 1773–1782. [Google Scholar] [CrossRef]
- Karamać, M.; Kosińska, A.; Estrella, I.; Hernández, T.; Duenas, M. Antioxidant activity of phenolic compounds identified in sunflower seeds. Eur. Food Res. Technol. 2012, 235, 221–230. [Google Scholar] [CrossRef]
- Amakura, Y.; Yoshimura, M.; Yamakami, S.; Yoshida, T. Isolation of phenolic constituents and characterization of antioxidant markers from sunflower (Helianthus annuus) seed extract. Phytochem. Lett. 2013, 6, 302–305. [Google Scholar] [CrossRef]
- Weisz, G.M.; Kammerer, D.R.; Carle, R. Identification and quantification of phenolic compounds from sunflower (Helianthus annuus L.) kernels and shells by HPLC-DAD/ESI-MS. Food Chem. 2009, 115, 758–765. [Google Scholar] [CrossRef]
- Pedrosa, M.M.; Muzquiz, M.; Garcìa-Vallejo, C.; Burbano, C.; Cuadrado, C.; Ayet, G.; Robredo, L.M. Determination of caffeic and chlorogenic acids and their derivatives in different sunflower seeds. J. Sci. Food Agric. 2000, 80, 459–464. [Google Scholar] [CrossRef]
- Liang, Q.; Cui, J.; Li, H.; Liu, J.; Zhao, G. Florets of sunflower (Helianthus annuus L.): Potential new sources of dietary fiber and phenolic acids. J. Agric. Food Chem. 2013, 61, 3435–3442. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Liang, Q.; Li, H.; Zhao, G. Solvent effects on phenolic content, composition, and antioxidant activity of extracts from florets of sunflower (Helianthus annuus L.). Ind. Crop. Prod. 2015, 76, 574–581. [Google Scholar] [CrossRef]
- Fernandez, O.; Urrutia, M.; Berton, T.; Bernillon, S.; Deborde, C.; Jacob, D.; Maucourt, M.; Maury, P.; Duruflé, H.; Gibon, Y.; et al. Metabolomic characterization of sunflower leaf allows discriminating genotype groups or stress levels with a minimal set of metabolic markers. Metabolomics 2019, 15, 56. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.M.; Bowsher, A.W.; Crowell, B.L.; Celoy, R.M.; Tsai, C.J.; Donovan, L.A. Macroevolution of leaf defenses and secondary metabolites across the genus Helianthus. New Phytol. 2016, 209, 1720–1733. [Google Scholar] [CrossRef] [PubMed]
- Onoja, S.O.; Nnadi, C.O.; Udem, S.C.; Anaga, A.O. Potential antidiabetic and antioxidant activities of a heliangolide sesquiterpene lactone isolated from Helianthus annuus L. leaves. Acta Pharm. 2020, 70, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Karamać, M.; Gai, F.; Longato, E.; Meineri, G.; Janiak, M.; Amarowicz, R.; Peiretti, P.G. Antioxidant activity and phenolic composition of amaranth (Amaranthus caudatus) during plant growth. Antioxidants 2019, 8, 173. [Google Scholar] [CrossRef]
- Pavlović, J.; Mitić, S.; Mitić, M.; Kocić, G.; Pavlović, A.; Tošić, S. Variation in the phenolic compounds profile and antioxidant activity in different parts of hawthorn (Crataegus pentagyna Willd.) during harvest periods. Pol. J. Food Nutr. Sci. 2019, 69, 367–378. [Google Scholar] [CrossRef]
- Karamać, M. In vitro study on the efficacy of tannin fractions of edible nuts as antioxidants. Eur. J. Lipid Sci. Tech. 2009, 111, 1063–1071. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of ”antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free-radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Orak, H.H.; Karamać, M.; Orak, A.; Amarowicz, R. Antioxidant potential and phenolic compounds of some widely consumed Turkish white bean (Phaseolus vulgaris L.) varieties. Pol. J. Food Nutr. Sci. 2016, 66, 253–260. [Google Scholar] [CrossRef]
- Miller, H.E. A simplified method for the evaluation of antioxidants. J. Am. Oil Chem. Soc. 1971, 48, 91. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Matthäus, B. Antioxidant activity of extracts obtained from residues of different oilseeds. J. Agric. Food Chem. 2002, 50, 3444–3452. [Google Scholar] [CrossRef]
- Mehmood, A.; Zhao, L.; Ishaq, M.; Safdar, B.; Wang, C.; Nadeem, M. Optimization of total phenolic contents, antioxidant, and in-vitro xanthine oxidase inhibitory activity of sunflower head. CyTA-J. Food 2018, 16, 957–964. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Clifford, M.N.; Knight, S.; Kuhnert, N. Discriminating between the six isomers of dicaffeoylquinic acid by LC-MSn. J. Agric. Food Chem. 2005, 53, 3821–3832. [Google Scholar] [CrossRef]
- Gutierrez, M.C.; Parry, A.; Tena, M.; Jorrin, J.; Edwards, R. Abiotic elicitation of coumarin phytoalexins in sunflower. Phytochemistry 1995, 38, 1185–1191. [Google Scholar] [CrossRef]
- Prats, E.; Bazzalo, M.E.; León, A.; Jorrín, J.V. Accumulation of soluble phenolic compounds in sunflower capitula correlates with resistance to Sclerotinia sclerotiorum. Euphytica 2003, 132, 321–329. [Google Scholar] [CrossRef]
- Koeppe, D.E.; Rohrbaugh, L.M.; Rice, E.L.; Wender, S.H. Tissue age and caffeoylquinic acid concentration in sunflower. Phytochemistry 1970, 9, 297–301. [Google Scholar] [CrossRef]
- Kim, J.Y.; Cho, J.Y.; Ma, Y.K.; Park, K.Y.; Lee, S.H.; Ham, K.S.; Lee, H.J.; Park, K.H.; Moon, J.H. Dicaffeoylquinic acid derivatives and flavonoid glucosides from glasswort (Salicornia herbacea L.) and their antioxidative activity. Food Chem. 2011, 125, 55–62. [Google Scholar] [CrossRef]
- Schmidt, S.; Pokorny, J. Potential application of oilseeds as source of antioxidants for food lipids—A review. Czech. J. Food Sci. 2005, 23, 93–102. [Google Scholar] [CrossRef]
- Karamać, M.; Gai, F.; Peiretti, P.G. Effect of the growth stage of false flax (Camelina sativa L.) on the phenolic compound content and antioxidant potential of the aerial part of the plant. Pol. J. Food Nutr. Sci. 2020, 70, 189–198. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Growth Stage | Days after Sowing | Extraction Yield (%) | Total Phenolic Content | |
|---|---|---|---|---|
| mg GAE/g Extract | mg GAE/g FM | |||
| Stem extension | 37 | 31.5 ± 0.9 a | 21.9 ± 2.3 b | 0.61 ± 0.07 b |
| Visible bud | 43 | 32.2 ± 1.7 a | 17.6 ± 2.8 b | 0.54 ± 0.11 b |
| Early flowering | 50 | 31.0 ± 1.4 a | 20.4 ± 6.1b | 0.56 ± 0.24 b |
| Mid-flowering | 57 | 32.2 ± 0.4 a | 29.3 ± 1.8 a | 1.03 ± 0.07 a |
| Late flowering | 63 | 26.2 ± 1.8 b | 21.7 ± 3.1 b | 0.70 ± 0.10 b |
| Compound No 1 | λmax (nm) | [M−H]− (m/z) | MS2 Ions (m/z) | Compound |
|---|---|---|---|---|
| 1 | 297sh, 324 | 353 | 191, 179, 135 | Neochlorogenic acid |
| 2 | 286sh, 325 | 341 | 179 | Caffeic acid hexose |
| 3 | 299sh, 325 | 353 | 191, 179, 135 | Chlorogenic acid |
| 4 | 298sh, 325 | 353 | 191, 179, 173, 135 | Cryptochlorogenic acid |
| 5 | 301sh, 324 | 515 | 353, 335, 191, 179, 173 | 3,4-Di-O-caffeoylquinic acid |
| 6 | 301sh, 327 | 515 | 353, 191, 179 | 3,5-Di-O-caffeoylquinic acid |
| 7 | 301sh, 326 | 515 | 353, 191, 179, 173 | 4,5-Di-O-caffeoylquinic acid |
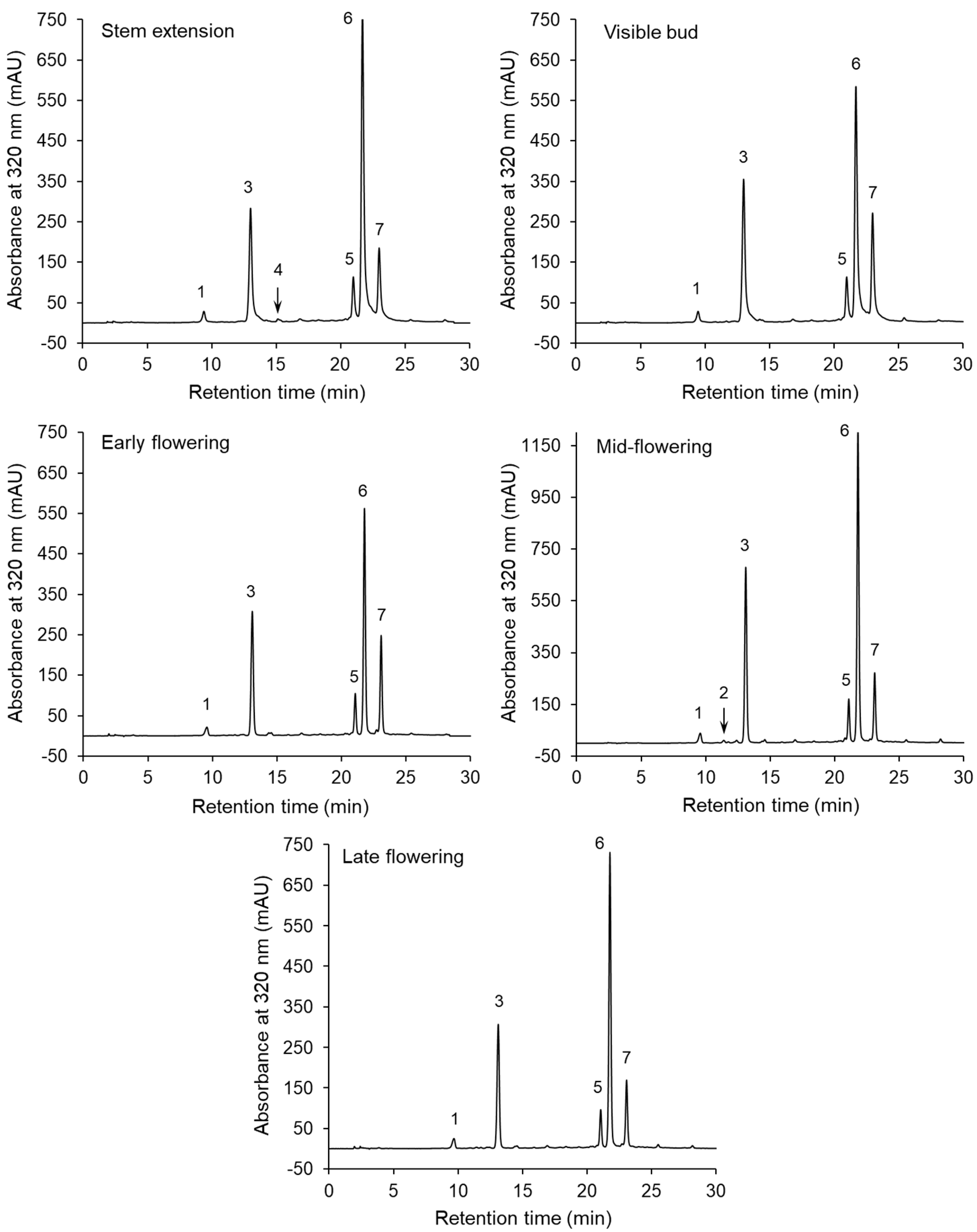
| Compound | Stem Extension | Visible Bud | Early Flowering | Mid-Flowering | Late Flowering |
|---|---|---|---|---|---|
| Neochlorogenic acid | 0.62 ± 0.09 b | 0.57 ± 0.13 b | 0.60 ± 0.05 b | 1.05 ± 0.09 a | 0.92 ± 0.05 a |
| Chlorogenic acid | 6.89 ± 0.12 b | 7.42 ± 1.22 b | 6.42 ± 0.86 b | 12.30 ± 0.52 a | 7.04 ± 0.58 b |
| 3,4-Di-O-caffeoylquinic acid 1 | 2.16 ± 0.46 b | 1.58 ± 0.41 b | 1.70 ± 0.09 b | 2.87 ± 0.12 a | 1.96 ± 0.35 b |
| 3,5-Di-O-caffeoylquinic acid 1 | 12.70 ± 3.69 b | 11.14 ± 2.24 b | 11.30 ± 1.37 b | 20.45 ± 0.82 a | 12.82 ± 1.60 b |
| 4,5-Di-O-caffeoylquinic acid 1 | 4.58 ± 0.71 a,b | 2.78 ± 0.35 c,d | 3.71 ± 0.69 b,c | 4.80 ± 0.39 a | 2.60 ± 0.51 d |
| Sum | 26.94 ± 4.76 b | 23.50 ± 4.27 b | 23.73 ±1.52 b | 41.47 ± 1.59 a | 25.35 ± 2.93 b |
| Compound | Stem Extension | Visible Bud | Early Flowering | Mid-Flowering | Late Flowering |
|---|---|---|---|---|---|
| Neochlorogenic acid | 0.017 ± 0.003 b | 0.017 ± 0.005 b | 0.016 ± 0.002 b | 0.036 ± 0.004 a | 0.031 ± 0.010 a |
| Chlorogenic acid | 0.193 ± 0.008 b | 0.228 ± 0.049 b | 0.175 ± 0.044 b | 0.435 ± 0.032 a | 0.234 ± 0.079 b |
| 3,4-Di-O-caffeoylquinic acid 1 | 0.061 ± 0.014 b | 0.049 ± 0.015 b | 0.046 ± 0.007 b | 0.101 ± 0.007 a | 0.066 ± 0.028 b |
| 3,5-Di-O-caffeoylquinic acid 1 | 0.358 ± 0.109 b | 0.342 ± 0.087 b | 0.306 ± 0.054 b | 0.722 ± 0.053 a | 0.422 ± 0.134 b |
| 4,5-Di-O-caffeoylquinic acid 1 | 0.129 ± 0.023 a,b | 0.086 ± 0.015 b | 0.101 ± 0.025 b | 0.170 ± 0.018 a | 0.088 ± 0.041 b |
| Sum | 0.76 ± 0.15 b | 0.72 ± 0.17 b | 0.64 ± 0.11 b | 1.46 ± 0.11 a | 0.84 ± 0.29 b |
| Growth Stage | TEAC | FRAP | EC50 (mg/mL) | ||
|---|---|---|---|---|---|
| mmol TE/g Extract | µmol TE/g FM | mmol Fe2+/g Extract | µmol Fe2+/g FM | ||
| Stem extension | 0.22 ± 0.02 b | 6.16 ± 0.77 b | 0.33 ± 0.04 a,b | 9.21 ± 1.05 b | 0.18 ± 0.03 a,b |
| Visible bud | 0.20 ± 0.03 b | 6.15 ± 1.15 b | 0.25 ± 0.10 b | 7.79 ± 3.43 b | 0.21 ± 0.03 a |
| Early flowering | 0.22 ± 0.05 b | 6.05 ± 2.09 b | 0.33 ± 0.04 a,b | 9.10 ± 0.02 b | 0.18 ± 0.05 a,b |
| Mid-flowering | 0.28 ± 0.03 a | 10.02 ± 1.22 a | 0.39 ± 0.04 a | 13.90 ± 1.53 a | 0.13 ± 0.01 b |
| Late flowering | 0.24 ± 0.03 a,b | 7.84 ± 1.13 a,b | 0.34 ± 0.08 a,b | 11.63 ± 3.24 a,b | 0.18 ± 0.01 a,b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gai, F.; Karamać, M.; Janiak, M.A.; Amarowicz, R.; Peiretti, P.G. Sunflower (Helianthus annuus L.) Plants at Various Growth Stages Subjected to Extraction—Comparison of the Antioxidant Activity and Phenolic Profile. Antioxidants 2020, 9, 535. https://doi.org/10.3390/antiox9060535
Gai F, Karamać M, Janiak MA, Amarowicz R, Peiretti PG. Sunflower (Helianthus annuus L.) Plants at Various Growth Stages Subjected to Extraction—Comparison of the Antioxidant Activity and Phenolic Profile. Antioxidants. 2020; 9(6):535. https://doi.org/10.3390/antiox9060535
Chicago/Turabian StyleGai, Francesco, Magdalena Karamać, Michał A. Janiak, Ryszard Amarowicz, and Pier Giorgio Peiretti. 2020. "Sunflower (Helianthus annuus L.) Plants at Various Growth Stages Subjected to Extraction—Comparison of the Antioxidant Activity and Phenolic Profile" Antioxidants 9, no. 6: 535. https://doi.org/10.3390/antiox9060535
APA StyleGai, F., Karamać, M., Janiak, M. A., Amarowicz, R., & Peiretti, P. G. (2020). Sunflower (Helianthus annuus L.) Plants at Various Growth Stages Subjected to Extraction—Comparison of the Antioxidant Activity and Phenolic Profile. Antioxidants, 9(6), 535. https://doi.org/10.3390/antiox9060535