Chemical Composition and Biological Activities of the Nord-West Romanian Wild Bilberry (Vaccinium myrtillus L.) and Lingonberry (Vaccinium vitis-idaea L.) Leaves


 ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Samples and Growing Conditions
2.2. Description of Habitats
2.3. Chemicals and Reagents
2.4. Ultrasound-Assisted Extraction Procedure
2.5. Analysis of Phenolic Compounds
2.5.1. HPLC-DAD-ESI-MS Analysis
2.5.2. Total Phenolic Content
2.5.3. Total Flavonoid Content
2.5.4. Total Anthocyanin Content
2.6. DPPH Free-Radical-Scavenging-Assay
2.7. Antimicrobial and Antifungal Capacity
2.7.1. Stains and Cultivation Conditions
2.7.2. Microdilution Technique
2.7.3. Antifungal Assay
2.8. Mutagenic and Antimutagenic Assay
2.9. Statistical Analysis
3. Results and Discussion
3.1. Phenolic Profile of Wild Bilberry and Lingonberry Leaves
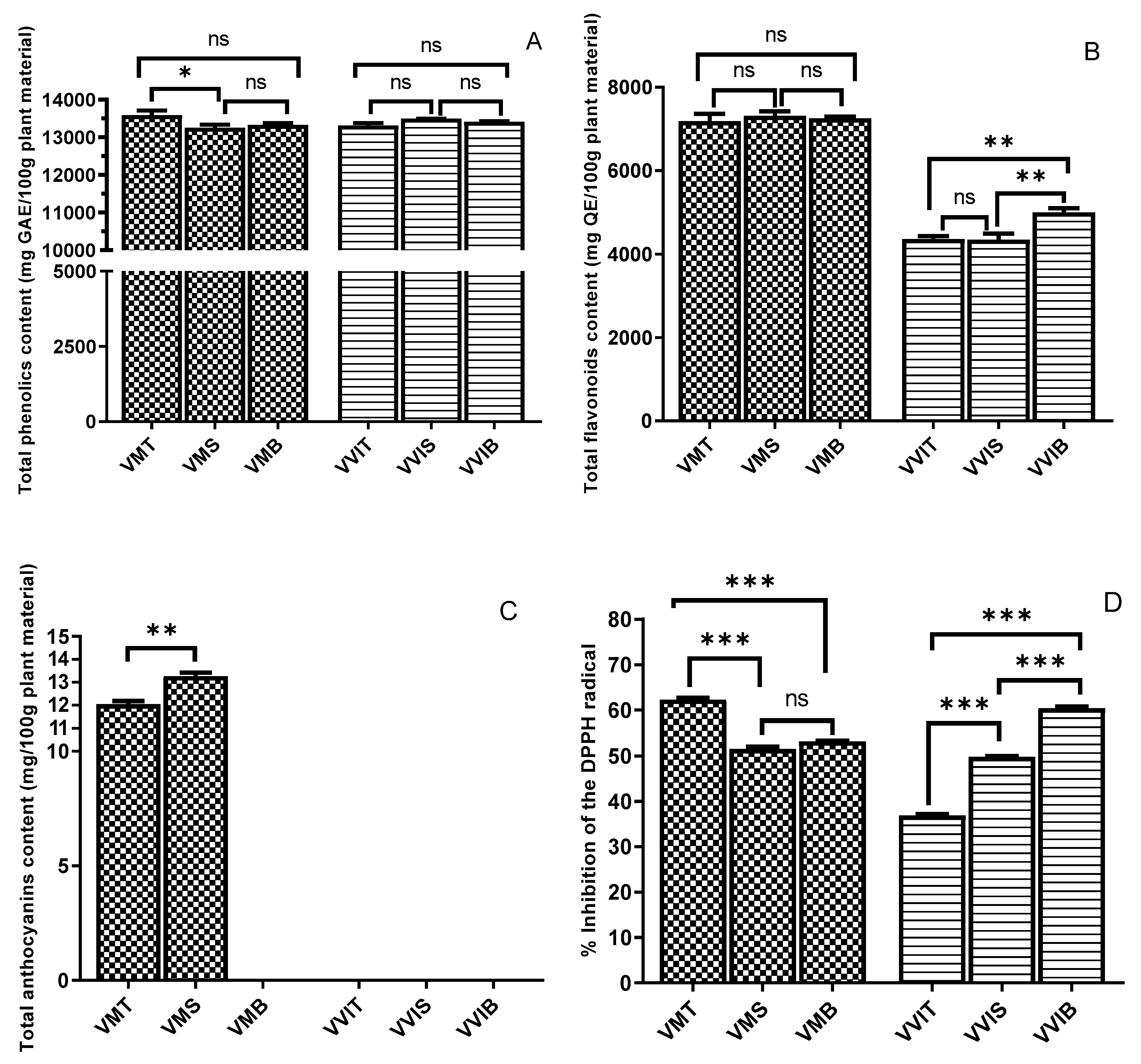
3.2. Total Phenolics and Total Flavonoids
3.3. Total Anthocyanin Content
3.4. DPPH Antioxidant Activity
3.5. Assessment of Antimicrobial Capacity
3.6. Assessment of Antimutagenic Effects of Bilberry and Lingonberry Leaves
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ștefănescu, B.E.; Szabo, K.; Mocan, A.; Crişan, G. Phenolic Compounds from Five Ericaceae Species Leaves and Their Related Bioavailability and Health Benefits. Molecules 2019, 24, 2046. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox Signal. 2012, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J.; Shahidi, F.; Yeo, J. Bioactivities of Phenolics by Focusing on Suppression of Chronic Diseases: A Review. Int. J. Mol. Sci. 2018, 19, 1573. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. 2017, 20, 1689–1699. [Google Scholar] [CrossRef]
- Popa, D.-S.; Bolfa, P.; Kiss, B.; Vlase, L.; Păltinean, R.; Pop, A.; Cătoi, C.; Crişan, G.; Loghin, F. Influence of Genista Tinctoria L or Methylparaben on Subchronic Toxicity of Bisphenol A in Rats. Biomed. Environ. Sci. 2014, 27, 85–96. [Google Scholar] [CrossRef]
- Pop, A.; Berce, C.; Bolfa, P.F.; Cǎtoi, C.; Dumitrescu, I.-B.; Silaghi-Dumitrescu, L.; Loghin, F. Evaluation of the possible endocrine disruptive effect of butylated hydroxyanisole, butylated hydroxytoluene and propyl gallate in immature female rats. Farmacia 2013, 61, 202–211. [Google Scholar]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Mol. Basel Switz. 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef]
- Calinoiu, L.-F.; Mitrea, L.; Precup, G.; Bindea, M.; Rusu, B.; Dulf, F.-V.; Stefanescu, B.-E.; Vodnar, D.-C. Characterization of Grape and Apple Peel Wastes’ Bioactive Compounds and Their Increased Bioavailability After Exposure to Thermal Process. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca-Food Sci. Technol. 2017, 74, 80–89. [Google Scholar] [CrossRef][Green Version]
- Călinoiu, L.F.; Cătoi, A.-F.; Vodnar, D.C. Solid-State Yeast Fermented Wheat and Oat Bran as A Route for Delivery of Antioxidants. Antioxidants 2019, 8, 372. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Barros, L.; Ferreira, I.C.F.R. In vivo antioxidant activity of phenolic compounds: Facts and gaps. Trends Food Sci. Technol. 2016, 48, 1–12. [Google Scholar] [CrossRef]
- Calinoiu, L.F.; Farcas, A.; Socaci, S.; Vodnar, D.C. Innovative Sources; Galanakis, C.M., Ed.; Academic Press Ltd-Elsevier Science Ltd.: London, UK, 2019; pp. 235–265. ISBN 978-0-12-817516-3. [Google Scholar]
- Bujor, O.C.; Ginies, C.; Popa, V.I.; Dufour, C. Phenolic compounds and antioxidant activity of lingonberry (Vaccinium vitis-idaea L.) leaf, stem and fruit at different harvest periods. Food Chem. 2018, 252, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Ancillotti, C.; Ciofi, L.; Rossini, D.; Chiuminatto, U.; Stahl-Zeng, J.; Orlandini, S.; Furlanetto, S.; Del Bubba, M. Liquid chromatographic/electrospray ionization quadrupole/time of flight tandem mass spectrometric study of polyphenolic composition of different Vaccinium berry species and their comparative evaluation. Anal. Bioanal. Chem. 2017, 409, 1347–1368. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Liimatainen, J.; Alanne, A.L.; Lindstedt, A.; Liu, P.; Sinkkonen, J.; Kallio, H.; Yang, B. Phenolic compounds extracted by acidic aqueous ethanol from berries and leaves of different berry plants. Food Chem. 2017, 220, 266–281. [Google Scholar] [CrossRef]
- Grace, M.H.; Esposito, D.; Dunlap, K.L.; Lila, M.A. Comparative Analysis of Phenolic Content and Profile, Antioxidant Capacity, and Anti-inflammatory Bioactivity in Wild Alaskan and Commercial Vaccinium Berries. J. Agric. Food Chem. 2014, 62, 4007–4017. [Google Scholar] [CrossRef]
- Ermis, E.; Hertel, C.; Schneider, C.; Carle, R.; Stintzing, F.; Schmidt, H. Characterization of in vitro antifungal activities of small and American cranberry (Vaccinium oxycoccos L. and V. macrocarpon Aiton) and lingonberry (Vaccinium vitis-idaea L.) concentrates in sugar reduced fruit spreads. Int. J. Food Microbiol. 2015, 204, 111–117. [Google Scholar] [CrossRef]
- Bujor, O.C.; Le Bourvellec, C.; Volf, I.; Popa, V.I.; Dufour, C. Seasonal variations of the phenolic constituents in bilberry (Vaccinium myrtillus L.) leaves, stems and fruits, and their antioxidant activity. Food Chem. 2016, 213, 58–68. [Google Scholar] [CrossRef]
- Bujor, O.-C.; Tanase, C.; Popa, M.E. Phenolic Antioxidants in Aerial Parts of Wild Vaccinium Species: Towards Pharmaceutical and Biological Properties. Antioxid. Basel Switz. 2019, 8, 649. [Google Scholar] [CrossRef]
- Vyas, P.; Kalidindi, S.; Chibrikova, L.; Igamberdiev, A.U.; Weber, J.T. Chemical analysis and effect of blueberry and lingonberry fruits and leaves against glutamate-mediated excitotoxicity. J. Agric. Food Chem. 2013, 61, 7769–7776. [Google Scholar] [CrossRef]
- Teleszko, M.; Wojdyło, A. Comparison of phenolic compounds and antioxidant potential between selected edible fruits and their leaves. J. Funct. Foods 2015, 14, 736–746. [Google Scholar] [CrossRef]
- Hokkanen, J.; Mattila, S.; Jaakola, L.; Pirttila, A.M.; Tolonen, A. Identification of phenolic compounds from lingonberry (Vaccinium vitis-idaea L.), Bilberry (Vaccinium myrtillus L.) and Hybrid Bilberry (Vaccinium x intermedium Ruthe L.) Leaves. J. Agric. Food Chem. 2009, 57, 9437–9447. [Google Scholar] [CrossRef] [PubMed]
- Martz, F.; Jaakola, L.; Julkunen-Tiitto, R.; Stark, S. Phenolic Composition and Antioxidant Capacity of Bilberry (Vaccinium myrtillus) Leaves in Northern Europe Following Foliar Development and Along Environmental Gradients. J. Chem. Ecol. 2010, 36, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Lindstedt, A.; Markkinen, N.; Sinkkonen, J.; Suomela, J.-P.; Yang, B. Characterization of Metabolite Profiles of Leaves of Bilberry (Vaccinium myrtillus L.) and Lingonberry (Vaccinium vitis-idaea L.). J. Agric. Food Chem. 2014, 62, 12015–12026. [Google Scholar] [CrossRef]
- Piterà, F. Compedio di Gemmoterapia Clinica; De Ferrari: Genova, Italy, 1994. [Google Scholar]
- Ieri, F.; Martini, S.; Innocenti, M.; Mulinacci, N. Phenolic distribution in liquid preparations of Vaccinium myrtillus L. and Vaccinium vitis idaea L. Phytochem. Anal. 2013, 24, 467–475. [Google Scholar] [CrossRef]
- Cignarella, A.; Nastasi, M.; Cavalli, E.; Puglisi, L. Novel lipid-lowering properties of Vaccinium myrtillus L. leaves, a traditional antidiabetic treatment, in several models of rat dyslipidaemia: A comparison with ciprofibrate. Thromb. Res. 1996, 84, 311–322. [Google Scholar] [CrossRef]
- Wang, X.; Sun, H.; Fan, Y.; Li, L.; Makino, T.; Kano, Y. Analysis and bioactive evaluation of the compounds absorbed into blood after oral administration of the extracts of Vaccinium vitis-idaea in rat. Biol. Pharm. Bull. 2005, 28, 1106–1108. [Google Scholar] [CrossRef]
- Raudone, L.; Vilkickyte, G.; Pitkauskaite, L.; Raudonis, R.; Vainoriene, R.; Motiekaityte, V. Antioxidant Activities of Vaccinium vitis-idaea L. Leaves within Cultivars and Their Phenolic Compounds. Molecules 2019, 24, 844. [Google Scholar] [CrossRef]
- Alam, Z.; Morales, H.R.; Roncal, J. Environmental conditions affect phenolic content and antioxidant capacity of leaves and fruit in wild partridgeberry (Vaccinium vitis-idaea). Botany 2016, 94, 509–521. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems; with 47 Tables; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003; ISBN 978-3-540-00347-2. [Google Scholar]
- Bilger, W.; Rolland, M.; Nybakken, L. UV screening in higher plants induced by low temperature in the absence of UV-B radiation. Photochem. Photobiol. Sci. 2007, 6, 190–195. [Google Scholar] [CrossRef]
- Markham, K.R.; Tanner, G.J.; Caasi-Lit, M.; Whitecross, M.I.; Nayudu, M.; Mitchell, K.A. Possible protective role for 3′,4′-dihydroxyflavones induced by enhanced UV-B in a UV-tolerant rice cultivar. Phytochemistry 1998, 49, 1913–1919. [Google Scholar] [CrossRef]
- Wellmann, E. UV dose-dependent induction of enzymes related to flavonoid biosynthesis in cell suspension cultures of parsley. FEBS Lett. 1975, 51, 105–107. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä-Riihinen, K.; Kärenlampi, S.; Hohtola, A. Activation of flavonoid biosynthesis by solar radiation in bilberry (Vaccinium myrtillus L.) leaves. Planta 2004, 218, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Uleberg, E.; Rohloff, J.; Jaakola, L.; Trôst, K.; Junttila, O.; Häggman, H.; Martinussen, I. Effects of Temperature and Photoperiod on Yield and Chemical Composition of Northern and Southern Clones of Bilberry (Vaccinium myrtillus L.). J. Agric. Food Chem. 2012, 60, 10406–10414. [Google Scholar] [CrossRef]
- Åkerström, A.; Jaakola, L.; Bång, U.; Jäderlund, A. Effects of Latitude-Related Factors and Geographical Origin on Anthocyanidin Concentrations in Fruits of Vaccinium myrtillus L. (Bilberries). J. Agric. Food Chem. 2010, 58, 11939–11945. [Google Scholar] [CrossRef]
- Jovančević, M.; Balijagić, J.; Menković, N.R.; Scaron, K.; avikin; Zdunić, G.M.; Janković, T.; Dekić-Ivanković, M. Analysis of phenolic compounds in wild populations of bilberry (Vaccinium myrtillus L.) from Montenegro. J. Med. Plant Res. 2011, 5, 910–914. [Google Scholar]
- Mikulic-Petkovsek, M.; Schmitzer, V.; Slatnar, A.; Stampar, F.; Veberic, R. A comparison of fruit quality parameters of wild bilberry (Vaccinium myrtillus L.) growing at different locations. J. Sci. Food Agric. 2015, 95, 776–785. [Google Scholar] [CrossRef]
- Meteoblue. Available online: https://www.meteoblue.com/ro/vreme/historyclimate/weatherarchive/turda_rom%C3%A2nia_664460 (accessed on 4 June 2020).
- Meteoromania. Available online: http://www.meteoromania.ro/servicii/date-meteorologice/arhiva-precipitatii/ (accessed on 4 June 2020).
- Meteoblue. Available online: https://www.meteoblue.com/ro/vreme/historyclimate/weatherarchive/bor%c5%9fa_rom%c3%a2nia_684156 (accessed on 4 June 2020).
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.H.; Toşa, M.I. Total Phenolic Contents, Antioxidant Activities, and Lipid Fractions from Berry Pomaces Obtained by Solid-State Fermentation of Two Sambucus Species with Aspergillus niger. J. Agric. Food Chem. 2015, 63, 3489–3500. [Google Scholar] [CrossRef]
- Dulf, F.V.; Vodnar, D.C.; Socaciu, C. Effects of solid-state fermentation with two filamentous fungi on the total phenolic contents, flavonoids, antioxidant activities and lipid fractions of plum fruit (Prunus domestica L.) by-products. Food Chem. 2016, 209, 27–36. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and Measurement of Anthocyanins by UV-Visible Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Ebrahimabadi, A.H.; Mazoochi, A.; Kashi, F.J.; Djafari-Bidgoli, Z.; Batooli, H. Essential oil composition and antioxidant and antimicrobial properties of the aerial parts of Salvia eremophila Boiss. from Iran. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2010, 48, 1371–1376. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, M.P. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2018; ISBN 978-1-56238-836-2. [Google Scholar]
- Schwalbe, R.; Steele-Moore, L.; Goodwin, A.C. Antimicrobial Susceptibility Testing Protocols; CRC Press: Boca Raton, FL, USA, 2007; ISBN 978-1-4200-1449-5. [Google Scholar]
- Alexander, B.D. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; ISBN 978-1-56238-826-3. [Google Scholar]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. Mutagen. Relat. Subj. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Saraç, N.; Şen, B. Antioxidant, mutagenic, antimutagenic activities, and phenolic compounds of Liquidambar orientalis Mill. var. orientalis. Ind. Crops Prod. 2014, 53, 60–64. [Google Scholar] [CrossRef]
- Ong, T.M.; Whong, W.Z.; Stewart, J.; Brockman, H.E. Chlorophyllin: A potent antimutagen against environmental and dietary complex mixtures. Mutat. Res. Lett. 1986, 173, 111–115. [Google Scholar] [CrossRef]
- Evandri, M.G.; Battinelli, L.; Daniele, C.; Mastrangelo, S.; Bolle, P.; Mazzanti, G. The antimutagenic activity of Lavandula angustifolia (lavender) essential oil in the bacterial reverse mutation assay. Food Chem. Toxicol. 2005, 43, 1381–1387. [Google Scholar] [CrossRef]
- Riihinen, K.; Jaakola, L.; Kärenlampi, S.; Hohtola, A. Organ-specific distribution of phenolic compounds in bilberry (Vaccinium myrtillus) and “northblue” blueberry (Vaccinium corymbosum x V. angustifolium). Food Chem. 2008, 110, 156–160. [Google Scholar] [CrossRef]
- Dabbou, S.; Sifi, S.; Rjiba, I.; Esposto, S.; Taticchi, A.; Servili, M.; Montedoro, G.F.; Hammami, M. Effect of Pedoclimatic Conditions on the Chemical Composition of the Sigoise Olive Cultivar. Chem. Biodivers. 2010, 7, 898–908. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Heinonen, M. Berry Phenolics and Their Antioxidant Activity. J. Agric. Food Chem. 2001, 49, 4076–4082. [Google Scholar] [CrossRef]
- Tian, Y.; Puganen, A.; Alakomi, H.-L.; Uusitupa, A.; Saarela, M.; Yang, B. Antioxidative and antibacterial activities of aqueous ethanol extracts of berries, leaves, and branches of berry plants. Food Res. Int. 2018, 106, 291–303. [Google Scholar] [CrossRef]
- Rieger, G.; Müller, M.; Guttenberger, H.; Bucar, F. Influence of Altitudinal Variation on the Content of Phenolic Compounds in Wild Populations of Calluna vulgaris, Sambucus nigra, and Vaccinium myrtillus. J. Agric. Food Chem. 2008, 56, 9080–9086. [Google Scholar] [CrossRef] [PubMed]
- Bidel, L.P.R.; Meyer, S.; Goulas, Y.; Cadot, Y.; Cerovic, Z.G. Responses of epidermal phenolic compounds to light acclimation: In vivo qualitative and quantitative assessment using chlorophyll fluorescence excitation spectra in leaves of three woody species. J. Photochem. Photobiol. B 2007, 88, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ma, F.; Cheng, L. Primary and secondary metabolism in the sun-exposed peel and the shaded peel of apple fruit. Physiol. Plant. 2013, 148, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Spitaler, R.; Winkler, A.; Lins, I.; Yanar, S.; Stuppner, H.; Zidorn, C. Altitudinal Variation of Phenolic Contents in Flowering Heads of Arnica montana cv. ARBO: A 3-Year Comparison. J. Chem. Ecol. 2008, 34, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Jakopic, J.; Stampar, F.; Veberic, R. The influence of exposure to light on the phenolic content of ‘Fuji’ apple. Sci. Hortic. 2009, 123, 234–239. [Google Scholar] [CrossRef]
- Rosłon, W.; Osińska, E.; Pióro-Jabrucka, E.; Grabowska, A. Morphological and chemical variability of wild populations of bilberry (Vaccinium myrtillus L.). Pol. J. Environ. Stud. 2011, 20, 237–243. [Google Scholar]
- Zheng, W.; Wang, S.Y. Oxygen Radical Absorbing Capacity of Phenolics in Blueberries, Cranberries, Chokeberries, and Lingonberries. J. Agric. Food Chem. 2003, 51, 502–509. [Google Scholar] [CrossRef]
- Jaakola, L.; Määttä, K.; Pirttilä, A.M.; Törrönen, R.; Kärenlampi, S.; Hohtola, A. Expression of Genes Involved in Anthocyanin Biosynthesis in Relation to Anthocyanin, Proanthocyanidin, and Flavonol Levels during Bilberry Fruit Development. Plant Physiol. 2002, 130, 729–739. [Google Scholar] [CrossRef]
- Määttä-Riihinen, K.R.; Kamal-Eldin, A.; Mattila, P.H.; González-Paramás, A.M.; Törrönen, A.R. Distribution and contents of phenolic compounds in eighteen Scandinavian berry species. J. Agric. Food Chem. 2004, 52, 4477–4486. [Google Scholar] [CrossRef]
- Szakiel, A.; Pączkowski, C.; Koivuniemi, H.; Huttunen, S. Comparison of the triterpenoid content of berries and leaves of lingonberry Vaccinium vitis-idaea from Finland and Poland. J. Agric. Food Chem. 2012, 60, 4994–5002. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Goupy, P.; Dufour, C.; Loonis, M.; Dangles, O. Quantitative Kinetic Analysis of Hydrogen Transfer Reactions from Dietary Polyphenols to the DPPH Radical. J. Agric. Food Chem. 2003, 51, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Soobrattee, M.A.; Neergheen, V.S.; Luximon-Ramma, A.; Aruoma, O.I.; Bahorun, T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. 2005, 579, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) Models. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Coppo, E.; Marchese, A. Antibacterial activity of polyphenols. Curr. Pharm. Biotechnol. 2014, 15, 380–390. [Google Scholar] [CrossRef]
- Riihinen, K.R.; Ou, Z.M.; Gödecke, T.; Lankin, D.C.; Pauli, G.F.; Wu, C.D. The antibiofilm activity of lingonberry flavonoids against oral pathogens is a case connected to residual complexity. Fitoterapia 2014, 97, 78–86. [Google Scholar] [CrossRef]
- Vučić, D.M.; Petković, M.R.; Rodić-Grabovac, B.B.; Stefanović, O.D.; Vasić, S.M.; Čomić, L.R. Antibacterial and antioxidant activities of bilberry (Vaccinium myrtillus L.) in vitro. Afr. J. Microbiol. Res. 2013, 7, 5130–5136. [Google Scholar]
- Kylli, P.; Nohynek, L.; Puupponen-Pimiä, R.; Westerlund-Wikström, B.; Leppänen, T.; Welling, J.; Moilanen, E.; Heinonen, M. Lingonberry (Vaccinium vitis-idaea) and European Cranberry (Vaccinium microcarpon) Proanthocyanidins: Isolation, Identification, and Bioactivities. J. Agric. Food Chem. 2011, 59, 3373–3384. [Google Scholar] [CrossRef]
- Puupponen-Pimiä, R.; Nohynek, L.; Alakomi, H.-L.; Oksman-Caldentey, K.-M. Bioactive berry compounds—Novel tools against human pathogens. Appl. Microbiol. Biotechnol. 2005, 67, 8–18. [Google Scholar] [CrossRef]
- Nohynek, L.J.; Alakomi, H.-L.; Kähkönen, M.P.; Heinonen, M.; Helander, I.M.; Oksman-Caldentey, K.-M.; Puupponen-Pimiä, R.H. Berry Phenolics: Antimicrobial Properties and Mechanisms of Action Against Severe Human Pathogens. Nutr. Cancer 2006, 54, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Zahin, M.; Aqil, F.; Ahmad, I. Broad spectrum antimutagenic activity of antioxidant active fraction of punica granatum L. peel extracts. Mutat. Res. 2010, 703, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Brindzová, L.; Zalibera, M.; Jakubík, T.; Mikulášová, M.; Takácsová, M.; Mošovská, S.; Rapta, P. Antimutagenic and Radical Scavenging Activity of Wheat Bran. Cereal Res. Commun. 2009, 37, 45–55. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Thermal Processing for the Release of Phenolic Compounds from Wheat and Oat Bran. Biomolecules 2020, 10, 21. [Google Scholar] [CrossRef]
- Hope Smith, S.; Tate, P.L.; Huang, G.; Magee, J.B.; Meepagala, K.M.; Wedge, D.E.; Larcom, L.L. Antimutagenic activity of berry extracts. J. Med. Food 2004, 7, 450–455. [Google Scholar] [CrossRef]
- Prencipe, F.P.; Bruni, R.; Guerrini, A.; Rossi, D.; Benvenuti, S.; Pellati, F. Metabolite profiling of polyphenols in Vaccinium berries and determination of their chemopreventive properties. J. Pharm. Biomed. Anal. 2014, 89, 257–267. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Côté, J.; Caillet, S.; Doyon, G.; Sylvain, J.-F.; Lacroix, M. Bioactive Compounds in Cranberries and their Biological Properties. Crit. Rev. Food Sci. Nutr. 2010, 50, 666–679. [Google Scholar] [CrossRef]
- Jurikova, T.; Mlcek, J.; Skrovankova, S.; Sumczynski, D.; Sochor, J.; Hlavacova, I.; Snopek, L.; Orsavova, J. Fruits of Black Chokeberry Aronia melanocarpa in the Prevention of Chronic Diseases. Molecules 2017, 22, 944. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Phenolic Compounds | Retention Time Rt (min) | UV λmax (nm) | [M+H]+ (m/z) | VMT | VMS | VMB | VVIT | VVIS | VVIB | |
|---|---|---|---|---|---|---|---|---|---|---|
| Flavanols | Gallocatechin | 2.97 | 279 | 307, 290 | 7.59 ± 0.07 b | 15.37 ± 0.14 a | 4.84 ± 0.05 c | 35.10 ± 0.24 b | 46.81 ± 0.38 a | 31.41 ± 0.21 c |
| Epigallocatechin | 4.24 | 279 | 307, 290 | n.d | 6.56 ± 0.06 | n.d | 25.24 ± 0.72 b | 35.97 ± 0.23 a | 23.35 ± 0.61 c | |
| Catechin | 12.58 | 280 | 291 | 9.87 ± 0.07 a | 4.79 ± 0.07 c | 5.38 ± 0.09 b | 18.51 ± 0.21 b | 21.57 ± 0.2 a | 17.43 ± 0.18 b | |
| Epicatechin | 13.11 | 280 | 291 | 4.31 ± 0.03 b | 9.66 ± 0.08 a | n.d | n.d | n.d | 2.78 ± 0.03 | |
| Procyanidin dimer I | 11.33 | 280 | 579, 291 | n.d | n.d | n.d | 6.38 ± 0.05 b | 8.36 ± 0.06 a | 6.27 ± 0.03 b | |
| Procyanidin dimer II | 19.74 | 280 | 579, 291 | 12.68 ± 0.11 a | 12.13 ± 0.12 a | 8.70 ± 0.07 b | 2.20 ± 0.05 b | 4.23 ± 0.04 a | 4.61 ± 0.03 a | |
| Procyanidin trimer | 13.89 | 280 | 865, 291 | 21.84 ± 0.21 b | 24.30 ± 0.72 a | 10.09 ± 0.12 c | 12.92±0.12 c | 14.21±0.16 b | 18.84±0.22 a | |
| Hydroxycinnamic acids | Chlorogenic acid | 12.01 | 281, 329 | 355, 163 | 3.34 ± 0.03 c | 3.85 ± 0.02 b | 5.94 ± 0.05 a | 0.79 ± 0.01 b | n.d | 1.16 ± 0.01 a |
| Feruloylquinic acid | 14.79 | 283, 330 | 369 | 55.37 ± 0.42 b | 47.66 ± 0.39 c | 59.65 ± 0.44 a | 31.05 ± 0.18 b | 24.61 ± 0.24 c | 33.42 ± 0.37 a | |
| Caffeoylarbutin | 17.20 | 288, 330 | 435 | n.d | n.d | n.d | 6.45 ± 0.04 a | 3.42 ± 0.02 c | 5.14 ± 0.05 b | |
| Dicaffeoylquinic acid | 20.08 | 282, 329 | 517, 163 | 5.01 ± 0.05 a | 4.05 ± 0.04 b | n.d | 1.77 ± 0.02 a | 0.93 ± 0.01 b | 1.74 ± 0.01 a | |
| Flavonols (quercetin derivatives) | Quercetin-rutinoside (Rutin) | 15.35 | 263, 355 | 611, 303 | 44.91 ± 0.21 b | 42.34 ± 0.19 c | 49.83 ± 0.63 a | 18.61 ± 0.19 b | 11.45 ± 0.10 c | 21.88 ± 0.19 a |
| Quercetin-glucoside | 16.20 | 263, 355 | 465, 303 | 1.42 ± 0.01 b | 1.29 ± 0.01 c | 2.37 ± 0.02 a | 3.05 ± 0.03 a | 2.23 ± 0.03 b | 1.91 ± 0.02 c | |
| Quercetin-acetyl-rhamnoside | 17.83 | 263, 356 | 493, 303 | 18.60 ± 0.16 a | 12.67 ± 0.10 c | 15.47 ± 0.14 b | 6.10 ± 0.04 b | 1.71 ± 0.01 c | 8.01 ± 0.07 a | |
| Quercetin-arabinoside | 18.69 | 262, 355 | 435, 303 | 1.55 ± 0.01 a | 1.53 ± 0.01 a | 1.39 ± 0.01 b | 0.41 ± 0.01 b | 0.07 ± 0.01 c | 0.61 ± 0.01 a | |
| Quercetin-xyloside | 18.98 | 262, 355 | 435, 303 | 1.47 ± 0.01 b | 1.30 ± 0.01 c | 1.53 ± 0.01 a | 0.45 ± 0.01 b | 0.05 ± 0.01 c | 0.62 ± 0.01 a | |
| Quercetin-diglucoside | 21.15 | 263, 355 | 628, 303 | 0.91 ± 0.01 c | 1.42 ± 0.01 a | 0.17 ± 0.01 b | 3.11 ± 0.03 b | 3.93 ± 0.05 a | 1.12 ± 0.01 c | |
| Quercetin | 21.88 | 261, 355 | 303 | 3.69 ± 0.03 a | 3.26 ± 0.04 b | 1.16 ± 0.06 c | 4.78 ± 0.04 a | 3.31 ± 0.02 b | 2.61 ± 0.02 c | |
| Anthocyanins | Cyanidin-glucoside | 11.02 | 210, 517 | 449, 287 | 0.28 ± 0.01 a | 0.29 ± 0.01 a | n.d | n.d | n.d | n.d |
| Cyanidin-arabinoside | 11.78 | 214, 517 | 419, 287 | n.d | 0.30 ± 0.01 | n.d | n.d | n.d | n.d | |
| Cyanidin-acetyl-glucoside | 14.28 | 218, 518 | 491, 287 | 0.33 ± 0.01 a | 0.29 ± 0.01 b | n.d | n.d | n.d | n.d | |
| Type of Strains | Gram-Positive | Gram-Negative | Fungi | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample | S. aureus | E. fecalis | R. equi | E. coli enterotoxigen | K. pneumonia | P. aeruginosa | Candida albicans | Candida zeylanoides | Candida parapsilosis |
| mg/mL | |||||||||
| VMT | 0.06 | 0.12 | 0.06 | 0.24 | 0.24 | 0.24 | 125 | 31.25 | 31.25 |
| VMS | 0.12 | 0.24 | 0.06 | 0.48 | 0.12 | 0.24 | 125 | 62.5 | 31.25 |
| VMB | 0.06 | 0.12 | 0.06 | 0.48 | 0.12 | 0.24 | 125 | 31.25 | 31.25 |
| VVIT | 0.12 | 0.12 | 0.06 | 0.48 | 0.12 | 0.96 | 125 | 62.5 | 31.25 |
| VVIS | 0.12 | 0.12 | 0.06 | 0.48 | 0.12 | 0.96 | 125 | 31.25 | 31.25 |
| VVIB | 0.12 | 0.12 | 0.06 | 0.48 | 0.12 | 0.96 | 125 | 62.5 | 31.25 |
| Fluconazole μg/mL | - | - | - | - | 15.62 | 7.81 | 15.62 | ||
| Streptomicyn μg/mL | 0.03 | 0.06 | 0.06 | 0.12 | 0.06 | 0.06 | |||
| Type of Strains | Gram-Positive | Gram-Negative | Fungi | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Sample | S. aureus | E. fecalis | R. equi | E. coli enterotoxigen | K. pneumonia | P. aeruginosa | Candida albicans | Candida zeylanoides | Candida parapsilosis |
| mg/mL | |||||||||
| VMT | 0.12 | 0.24 | 0.12 | 0.48 | 0.48 | 0.48 | 250 | 62.5 | 62.5 |
| VMS | 0.24 | 0.48 | 0.12 | 0.96 | 0.24 | 0.48 | 250 | 125 | 62.5 |
| VMB | 0.12 | 0.24 | 0.12 | 0.96 | 0.24 | 0.48 | 250 | 62.5 | 62.5 |
| VVIT | 0.24 | 0.24 | 0.12 | 0.96 | 0.24 | 1.92 | 250 | 125 | 62.5 |
| VVIS | 0.24 | 0.24 | 0.12 | 0.96 | 0.24 | 1.92 | 250 | 62.5 | 62.5 |
| VVIB | 0.24 | 0.24 | 0.12 | 0.96 | 0.24 | 1.92 | 250 | 125 | 62.5 |
| Fluconazole μg/mL | - | - | - | - | 31.24 | 15.62 | 31.24 | ||
| Streptomicyn μg/mL | 0.06 | 0.12 | 0.12 | 0.24 | 0.12 | 0.12 | |||
| Samples | Number of Revertants | |||
|---|---|---|---|---|
| TA 98 | TA100 | |||
| Mean ± S.D | Inhibition % | Mean ± S.D | Inhibition % | |
| Negative Control | 9.25 ± 3.6 a | 9.25 ± 2.4 a | ||
| VMT | 132 ± 3.2 | 31.95 | 198 ± 4.2 | 43.26 |
| VMS | 133 ± 4.4 | 31.44 | 201 ± 6.3 | 42.4 |
| VMB | 137 ± 3.6 | 29.38 | 202 ± 5.4 | 42.12 |
| VVIT | 144 ± 4.7 | 25.77 | 223 ± 2.6 | 36.1 |
| VVIS | 145 ± 2.1 | 25.25 | 245 ± 4.3 | 29.79 |
| VVIB | 144 ± 5.9 | 25.77 | 234 ± 7.9 | 32.95 |
| 4-NPD b | 194 ± 3.3 | - | - | - |
| NaN3 b | - | - | 349 ± 15.22 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ștefănescu, B.-E.; Călinoiu, L.F.; Ranga, F.; Fetea, F.; Mocan, A.; Vodnar, D.C.; Crișan, G. Chemical Composition and Biological Activities of the Nord-West Romanian Wild Bilberry (Vaccinium myrtillus L.) and Lingonberry (Vaccinium vitis-idaea L.) Leaves. Antioxidants 2020, 9, 495. https://doi.org/10.3390/antiox9060495
Ștefănescu B-E, Călinoiu LF, Ranga F, Fetea F, Mocan A, Vodnar DC, Crișan G. Chemical Composition and Biological Activities of the Nord-West Romanian Wild Bilberry (Vaccinium myrtillus L.) and Lingonberry (Vaccinium vitis-idaea L.) Leaves. Antioxidants. 2020; 9(6):495. https://doi.org/10.3390/antiox9060495
Chicago/Turabian StyleȘtefănescu, Bianca-Eugenia, Lavinia Florina Călinoiu, Floricuța Ranga, Florinela Fetea, Andrei Mocan, Dan Cristian Vodnar, and Gianina Crișan. 2020. "Chemical Composition and Biological Activities of the Nord-West Romanian Wild Bilberry (Vaccinium myrtillus L.) and Lingonberry (Vaccinium vitis-idaea L.) Leaves" Antioxidants 9, no. 6: 495. https://doi.org/10.3390/antiox9060495
APA StyleȘtefănescu, B.-E., Călinoiu, L. F., Ranga, F., Fetea, F., Mocan, A., Vodnar, D. C., & Crișan, G. (2020). Chemical Composition and Biological Activities of the Nord-West Romanian Wild Bilberry (Vaccinium myrtillus L.) and Lingonberry (Vaccinium vitis-idaea L.) Leaves. Antioxidants, 9(6), 495. https://doi.org/10.3390/antiox9060495