Prevention of Oxidative Stress-Induced Pancreatic Beta Cell Damage by Broussonetia kazinoki Siebold Fruit Extract via the ERK-Nox4 Pathway

 and
and 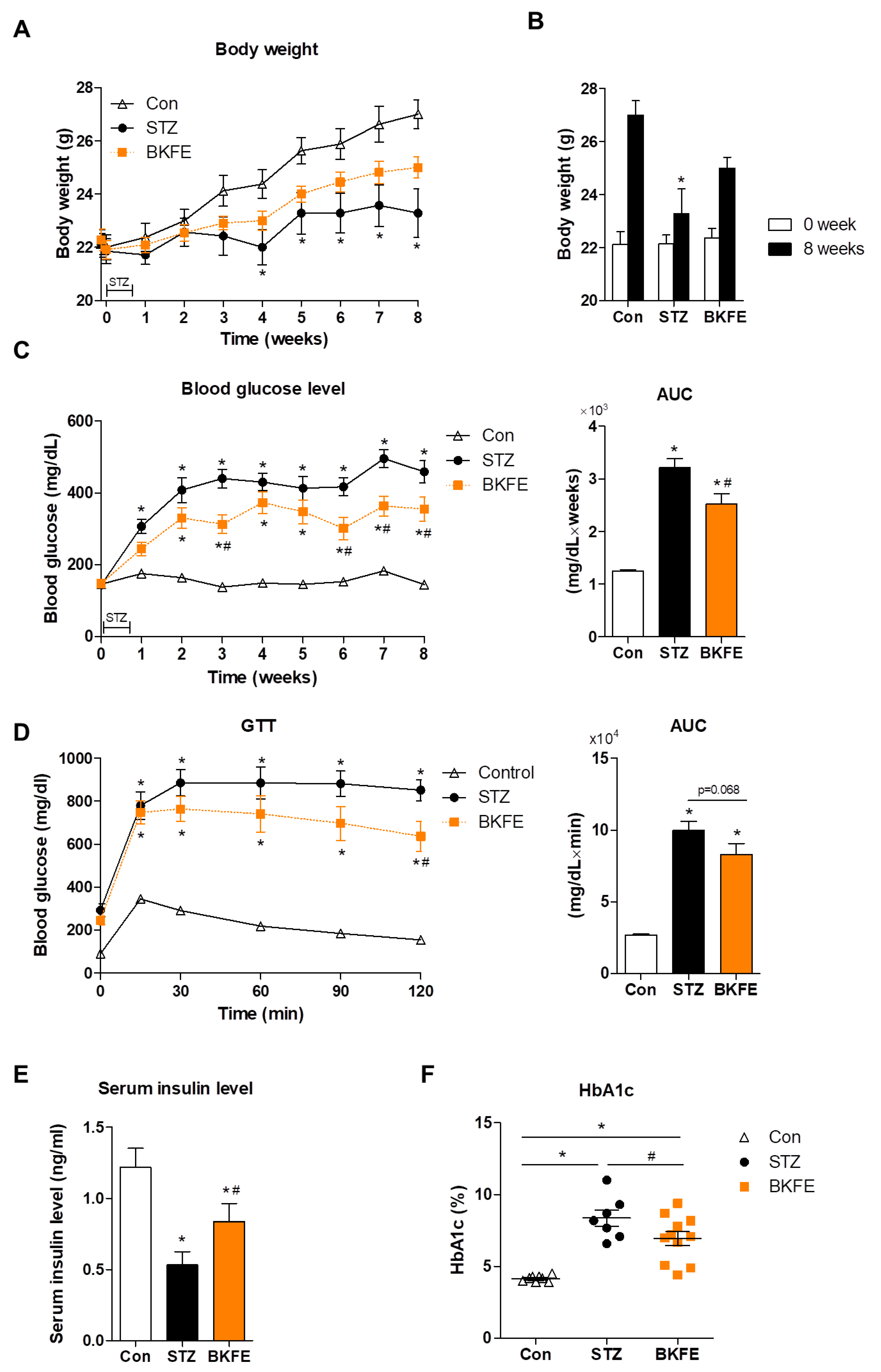{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of BKFE
2.2. Animals
2.3. Glucose Tolerance Test (GTT)
2.4. Measurement of Serum Insulin Levels
2.5. Measurement of Hemoglobin A1c (HbA1c) Levels
2.6. Immunostaining
2.7. Cell Culture
2.8. Cell Viability Assay
2.9. Annexin V and Propidium Iodide Staining
2.10. Measurement of ROS Levels
2.11. Western Blotting
2.12. Quantitative Real Time (RT)-Polymerase Chain Reaction (PCR)
2.13. Transfection
2.14. Statistical Analysis
3. Results
3.1. BKFE Ameliorates STZ-Induced Diabetes
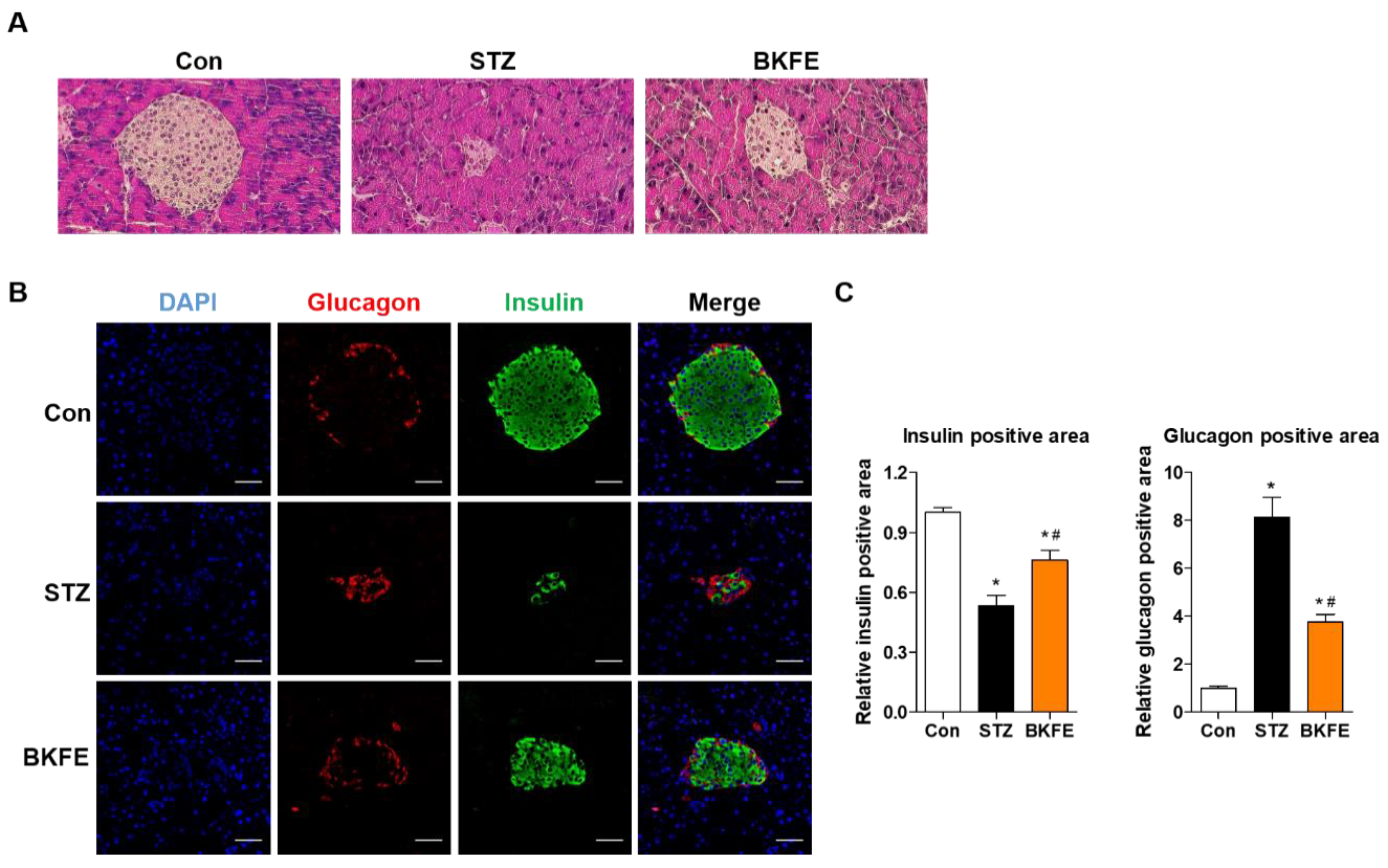
3.2. BKFE Reduces Beta Cell Damage in STZ-Induced Diabetic Mice
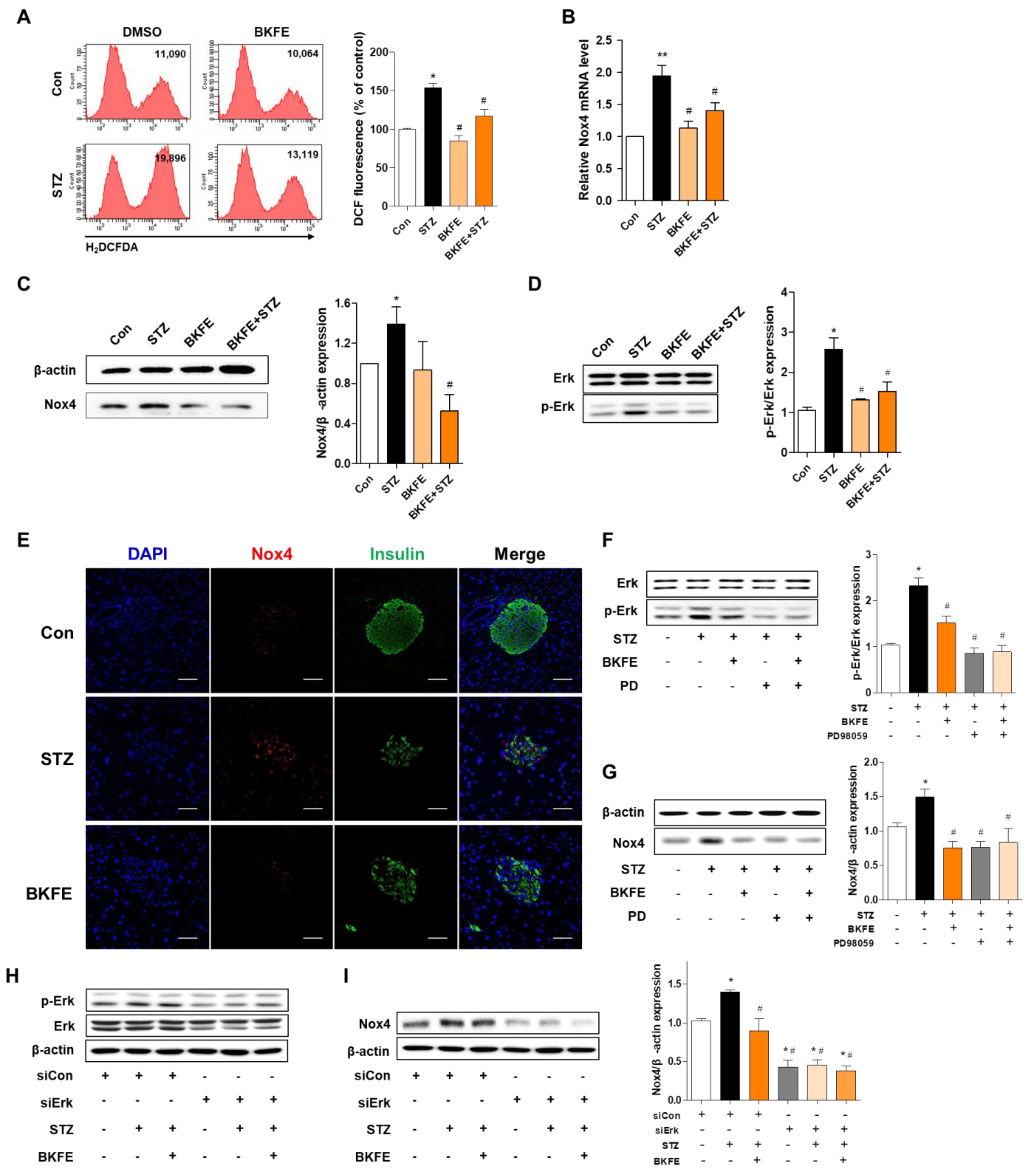
3.3. BKFE Reduces Oxidative Stress in Pancreatic Islets of STZ-Induced Diabetic Mice
3.4. BKFE Protects MIN6 Cells From STZ-Induced Toxicity
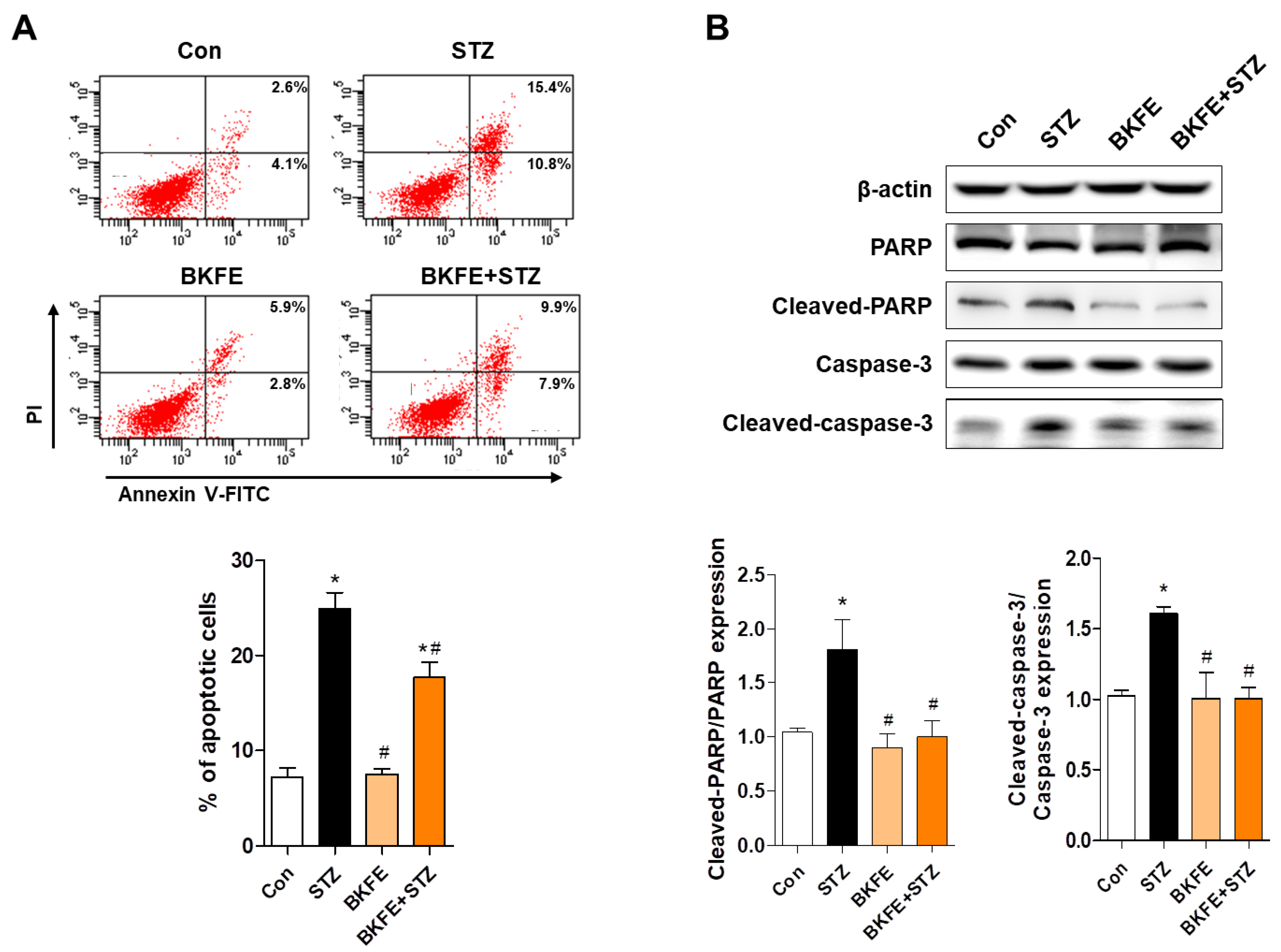
3.5. BKFE Inhibits STZ-Induced Apoptosis in MIN6 Cells
3.6. BKFE Inhibits STZ-Induced Production of ROS by Downregulation of P-Erk/Nox4 Pathway in MIN6 Cells and Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chiang, J.L.; Kirkman, M.S.; Laffel, L.M.; Peters, A.L. Type 1 diabetes through the life span: A position statement of the American Diabetes Association. Diabetes Care 2014, 37, 2034–2054. [Google Scholar] [CrossRef] [PubMed]
- Cnop, M.; Welsh, N.; Jonas, J.C.; Jorns, A.; Lenzen, S.; Eizirik, D.L. Mechanisms of pancreatic beta-cell death in type 1 and type 2 diabetes: Many differences, few similarities. Diabetes 2005, 54 (Suppl. 2). S97–S107. [Google Scholar] [CrossRef]
- Hameed, I.; Masoodi, S.R.; Mir, S.A.; Nabi, M.; Ghazanfar, K.; Ganai, B.A. Type 2 diabetes mellitus: From a metabolic disorder to an inflammatory condition. World J. Diabetes 2015, 6, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.J. Beta cell mass in diabetes: A realistic therapeutic target? Diabetologia 2008, 51, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Kohnert, K.D.; Freyse, E.J.; Salzsieder, E. Glycaemic variability and pancreatic beta-cell dysfunction. Curr. Diabetes Rev. 2012, 8, 345–354. [Google Scholar] [CrossRef]
- Ahmed Alfar, E.; Kirova, D.; Konantz, J.; Birke, S.; Mansfeld, J.; Ninov, N. Distinct Levels of Reactive Oxygen Species Coordinate Metabolic Activity with Beta-cell Mass Plasticity. Sci. Rep. 2017, 7, 3994. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Drews, G.; Krippeit-Drews, P.; Dufer, M. Oxidative stress and beta-cell dysfunction. Pflug. Arch. Eur. J. Physiol. 2010, 460, 703–718. [Google Scholar] [CrossRef]
- Cernea, S.; Dobreanu, M. Diabetes and beta cell function: From mechanisms to evaluation and clinical implications. Biochem. Med. 2013, 23, 266–280. [Google Scholar] [CrossRef]
- Marrif, H.I.; Al-Sunousi, S.I. Pancreatic beta Cell Mass Death. Front. Pharmacol. 2016, 7, 83. [Google Scholar] [CrossRef]
- Thomas, H.E.; McKenzie, M.D.; Angstetra, E.; Campbell, P.D.; Kay, T.W. Beta cell apoptosis in diabetes. Apoptosis Int. J. Program. Cell Death 2009, 14, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Gori, M.; Campbell, R.K. Natural products and diabetes treatment. Diabetes Educ. 1998, 24, 201–202, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Kooti, W.; Farokhipour, M.; Asadzadeh, Z.; Ashtary-Larky, D.; Asadi-Samani, M. The role of medicinal plants in the treatment of diabetes: A systematic review. Electron. Physician 2016, 8, 1832–1842. [Google Scholar] [CrossRef]
- Shukia, R.; Sharma, S.B.; Puri, D.; Prabhu, K.M.; Murthy, P.S. Medicinal plants for treatment of diabetes mellitus. Indian J. Clin. Biochem. 2000, 15, 169–177. [Google Scholar] [CrossRef]
- Veeresham, C. Natural products derived from plants as a source of drugs. J. Adv. Pharm. Technol. Res. 2012, 3, 200–201. [Google Scholar] [CrossRef]
- Gomes, R.M.; de Paulo, L.F.; Bonato Panizzon, C.; Neves, C.Q.; Cordeiro, B.C.; Zanoni, J.N.; Francisco, F.A.; Piovan, S.; de Freitas Mathias, P.C.; Longhini, R.; et al. Anti-Diabetic Effects of the Ethyl-Acetate Fraction of Trichilia catigua in Streptozo-tocin-Induced Type 1 Diabetic Rats. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 42, 1087–1097. [Google Scholar] [CrossRef]
- Jeon, Y.D.; Kang, S.H.; Moon, K.H.; Lee, J.H.; Kim, D.G.; Kim, W.; Kim, J.S.; Ahn, B.Y.; Jin, J.S. The Effect of Aronia Berry on Type 1 Diabetes in Vivo and In Vitro. J. Med. Food 2018, 21, 244–253. [Google Scholar] [CrossRef]
- Roh, S.S.; Kwon, O.J.; Yang, J.H.; Kim, Y.S.; Lee, S.H.; Jin, J.S.; Jeon, Y.D.; Yokozawa, T.; Kim, H.J. Allium hookeri root protects oxidative stress-induced inflammatory responses and beta-cell damage in pancreas of streptozotocin-induced diabetic rats. BMC Complementary Altern. Med. 2016, 16, 63. [Google Scholar] [CrossRef]
- Schumacher, N.S.; Colomeu, T.C.; de Figueiredo, D.; Carvalho Vde, C.; Cazarin, C.B.; Prado, M.A.; Meletti, L.M.; Zollner Rde, L. Identification and Antioxidant Activity of the Extracts of Eugenia uniflora Leaves. Characterization of the Anti-Inflammatory Properties of Aqueous Extract on Diabetes Expression in an Experimental Model of Spontaneous Type 1 Diabetes (NOD Mice). Antioxidants 2015, 4, 662–680. [Google Scholar] [CrossRef]
- Xu, J.; Liu, T.; Li, Y.; Yuan, C.; Ma, H.; Seeram, N.P.; Liu, F.; Mu, Y.; Huang, X.; Li, L. Hypoglycemic and hypolipidemic effects of triterpenoid-enriched Jamun (Eugenia jambolana Lam.) fruit extract in streptozotocin-induced type 1 diabetic mice. Food Funct. 2018, 9, 3330–3337. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.; Oh, P.; Jang, M. The properties of proteolytic enzymes from the fruit of Broussonetia kazinoki Siebold. J. Korean Soc. Food Nutr. (Korea) 1993, 22, 803–806. [Google Scholar]
- Cha, J.Y.; Kim, Y.T.; Kim, H.S.; Cho, Y.S. Antihyperglycemic effect of stem bark powder from paper mulberry (Broussonetia kazinoki Sieb.) in type 2 diabetic Otsuka Long-Evans Tokushima fatty rats. J. Med. Food 2008, 11, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.Y.; Ha, H.; Lee, H.; Lee, J.A.; Jeong, S.I.; Choi, Y.J.; Shin, H.K. Broussonetia kazinoki Siebold stimulates immune response in ovalbumin-immunized mice. J. Korean Orient. Med. 2011, 32, 8. [Google Scholar]
- Lee, H.; Ha, H.; Lee, J.K.; Park, S.J.; Jeong, S.I.; Shin, H.K. The Leaves of Broussonetia kazinoki Siebold Inhibit Atopic Dermatitis-Like Response on Mite Allergen-Treated Nc/Nga Mice. Biomol. Ther. 2014, 22, 438–444. [Google Scholar] [CrossRef]
- Wei, B.L.; Chen, Y.C.; Hsu, H.Y. Kazinol Q from Broussonetia kazinoki enhances cell death induced by Cu(II) through increased reactive oxygen species. Molecules 2011, 16, 3212–3221. [Google Scholar] [CrossRef]
- Kim, D.; Kim, H.J.; Cha, S.H.; Jun, H.S. Protective Effects of Broussonetia kazinoki Siebold Fruit Extract against Palmitate-Induced Lipotoxicity in Mesangial Cells. Evid. Based Complementary Altern. Med. 2019, 2019, 4509403. [Google Scholar] [CrossRef]
- Bae, U.J.; Jang, H.Y.; Lim, J.M.; Hua, L.; Ryu, J.H.; Park, B.H. Polyphenols isolated from Broussonetia kazinoki prevent cytokine-induced beta-cell damage and the development of type 1 diabetes. Exp. Mol. Med. 2015, 47, e160. [Google Scholar] [CrossRef]
- Bae, U.J.; Lee, D.Y.; Song, M.Y.; Lee, S.M.; Park, J.W.; Ryu, J.H.; Park, B.H. A prenylated flavan from Broussonetia kazinoki prevents cytokine-induced beta-cell death through suppression of nuclear factor-kappaB activity. Biol. Pharm. Bull. 2011, 34, 1026–1031. [Google Scholar] [CrossRef][Green Version]
- Rohlfing, C.L.; Little, R.R.; Wiedmeyer, H.M.; England, J.D.; Madsen, R.; Harris, M.I.; Flegal, K.M.; Eberhardt, M.S.; Goldstein, D.E. Use of GHb (HbA1c) in screening for undiagnosed diabetes in the U.S. population. Diabetes Care 2000, 23, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Szkudelski, T. The mechanism of alloxan and streptozotocin action in B cells of the rat pancreas. Physiol. Res. 2001, 50, 537–546. [Google Scholar] [PubMed]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Minami, K.; Kudo, M.; Iemoto, K.; Takahashi, H.; Seino, S. Liraglutide improves pancreatic Beta cell mass and function in alloxan-induced diabetic mice. PLoS ONE 2015, 10, e0126003. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.F.; Qiao, M.; Schroder, K.; Zhao, Q.; Asmis, R. Nox4 is a novel inducible source of reactive oxygen species in monocytes and macrophages and mediates oxidized low density lipoprotein-induced macrophage death. Circ. Res. 2010, 106, 1489–1497. [Google Scholar] [CrossRef]
- Donath, M.Y.; Halban, P.A. Decreased beta-cell mass in diabetes: Significance, mechanisms and therapeutic implications. Diabetologia 2004, 47, 581–589. [Google Scholar] [CrossRef]
- Rojas, J.; Bermudez, V.; Palmar, J.; Martinez, M.S.; Olivar, L.C.; Nava, M.; Tomey, D.; Rojas, M.; Salazar, J.; Garicano, C.; et al. Pancreatic Beta Cell Death: Novel Potential Mechanisms in Diabetes Therapy. J. Diabetes Res. 2018, 2018, 9601801. [Google Scholar] [CrossRef]
- Wu, J.; Yan, L.J. Streptozotocin-induced type 1 diabetes in rodents as a model for studying mitochondrial mechanisms of diabetic beta cell glucotoxicity. Diabetes Metab. Syndr. Obes. Targets Ther. 2015, 8, 181–188. [Google Scholar] [CrossRef]
- Yin, D.; Tao, J.; Lee, D.D.; Shen, J.; Hara, M.; Lopez, J.; Kuznetsov, A.; Philipson, L.H.; Chong, A.S. Recovery of islet beta-cell function in streptozotocin- induced diabetic mice: An indirect role for the spleen. Diabetes 2006, 55, 3256–3263. [Google Scholar] [CrossRef]
- Lei, H.; Han, J.; Wang, Q.; Guo, S.; Sun, H.; Zhang, X. Effects of sesamin on streptozotocin (STZ)-induced NIT-1 pancreatic beta-cell damage. Int. J. Mol. Sci. 2012, 13, 16961–16970. [Google Scholar] [CrossRef]
- Giardino, G.; Cicalese, M.P.; Delmonte, O.; Migliavacca, M.; Palterer, B.; Loffredo, L.; Cirillo, E.; Gallo, V.; Violi, F.; Pignata, C. NADPH Oxidase Deficiency: A Multisystem Approach. Oxidative Med. Cell. Longev. 2017, 2017, 4590127. [Google Scholar] [CrossRef]
- Teshima, Y.; Takahashi, N.; Nishio, S.; Saito, S.; Kondo, H.; Fukui, A.; Aoki, K.; Yufu, K.; Nakagawa, M.; Saikawa, T. Production of reactive oxygen species in the diabetic heart. Roles of mitochondria and NADPH oxidase. Circ. J. Off. J. Jpn. Circ. Soc. 2014, 78, 300–306. [Google Scholar] [CrossRef]
- Augsburger, F.; Filippova, A.; Rasti, D.; Seredenina, T.; Lam, M.; Maghzal, G.; Mahiout, Z.; Jansen-Durr, P.; Knaus, U.G.; Doroshow, J.; et al. Pharmacological characterization of the seven human NOX isoforms and their inhibitors. Redox Biol. 2019, 26, 101272. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Morgan, D.; Rebelato, E.; Oliveira-Emilio, H.C.; Procopio, J.; Curi, R.; Carpinelli, A. Insights into the critical role of NADPH oxidase(s) in the normal and dysregulated pancreatic beta cell. Diabetologia 2009, 52, 2489–2498. [Google Scholar] [CrossRef] [PubMed]
- Etoh, T.; Inoguchi, T.; Kakimoto, M.; Sonoda, N.; Kobayashi, K.; Kuroda, J.; Sumimoto, H.; Nawata, H. Increased expression of NAD(P)H oxidase subunits, NOX4 and p22phox, in the kidney of streptozotocin-induced diabetic rats and its reversibity by interventive insulin treatment. Diabetologia 2003, 46, 1428–1437. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Inoguchi, T.; Maeda, Y.; Sasaki, S.; Sawada, F.; Saito, R.; Kobayashi, K.; Sumimoto, H.; Takayanagi, R. Pitavastatin ameliorates albuminuria and renal mesangial expansion by downregulating NOX4 in db/db mice. Kidney Int. 2007, 72, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Elksnis, A.; Wikstrom, P.; Walum, E.; Welsh, N.; Carlsson, P.O. The novel NADPH oxidase 4 selective inhibitor GLX7013114 counteracts human islet cell death in vitro. PLoS ONE 2018, 13, e0204271. [Google Scholar] [CrossRef]
- Gorin, Y.; Wauquier, F. Upstream regulators and downstream effectors of NADPH oxidases as novel therapeutic targets for diabetic kidney disease. Mol. Cells 2015, 38, 285–296. [Google Scholar] [CrossRef]
- Peng, H.; Li, W.; Seth, D.M.; Nair, A.R.; Francis, J.; Feng, Y. (Pro)renin receptor mediates both angiotensin II-dependent and -independent oxidative stress in neuronal cells. PLoS ONE 2013, 8, e58339. [Google Scholar] [CrossRef]
- Yan, J.; Wang, C.; Jin, Y.; Meng, Q.; Liu, Q.; Liu, Z.; Liu, K.; Sun, H. Catalpol ameliorates hepatic insulin resistance in type 2 diabetes through acting on AMPK/NOX4/PI3K/AKT pathway. Pharmacol. Res. 2018, 130, 466–480. [Google Scholar] [CrossRef]
- Zhou, B.; Mu, J.; Gong, Y.; Lu, C.; Zhao, Y.; He, T.; Qin, Z. Brd4 inhibition attenuates unilateral ureteral obstruction-induced fibrosis by blocking TGF-beta-mediated Nox4 expression. Redox Biol. 2017, 11, 390–402. [Google Scholar] [CrossRef]
- Horiuchi, Y.; Nakatsu, D.; Kano, F.; Murata, M. Pyruvate kinase M1 interacts with A-Raf and inhibits endoplasmic reticulum stress-induced apoptosis by activating MEK1/ERK pathway in mouse insulinoma cells. Cell. Signal. 2017, 38, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Chang, H.; La, X.; Li, J.A.; Ma, L. Wushenziye Formula Inhibits Pancreatic beta Cell Apoptosis in Type 2 Diabetes Mellitus via MEK-ERK-Caspase-3 Signaling Pathway. Evid. Based Complementary Altern. Med. 2018, 2018, 4084259. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.I.; Awazu, M.; Tamiya, M.; Iwasaki, Y.; Harada, A.; Kugisaki, S.; Tanimura, S.; Kohno, M. Targeting the ERK signaling pathway as a potential treatment for insulin resistance and type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E643–e651. [Google Scholar] [CrossRef] [PubMed]
- Che, G.; Gao, H.; Hu, Q.; Xie, H.; Zhang, Y. Angiotensin II promotes podocyte injury by activating Arf6-Erk1/2-Nox4 signaling pathway. PLoS ONE 2020, 15, e0229747. [Google Scholar] [CrossRef] [PubMed]
- Gorin, Y.; Ricono, J.M.; Wagner, B.; Kim, N.H.; Bhandari, B.; Choudhury, G.G.; Abboud, H.E. Angiotensin II-induced ERK1/ERK2 activation and protein synthesis are redox-dependent in glomerular mesangial cells. Biochem. J. 2004, 381, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Kim, B.; Shin, H.S.; Kwon, H.J.; Park, E.S. The protective effect of hispidin against hydrogen peroxide-induced apoptosis in H9c2 cardiomyoblast cells through Akt/GSK-3beta and ERK1/2 signaling pathway. Exp. Cell Res. 2014, 327, 264–275. [Google Scholar] [CrossRef]
- Ryu, J.H.; Ahn, H.; Jin Lee, H. Inhibition of nitric oxide production on LPS-activated macrophages by kazinol B from Broussonetia kazinoki. Fitoterapia 2003, 74, 350–354. [Google Scholar] [CrossRef]
- Tsukamoto, D.; Shibano, M.; Okamoto, R.; Kusano, G. Studies on the constituents of Broussonetia species VIII. Four new pyrrolidine alkaloids, broussonetines R, S, T, and V and a new pyrroline alkaloid, broussonetine U, from Broussonetia kazinoki Sieb. Chem. Pharm. Bull. 2001, 49, 492–496. [Google Scholar] [CrossRef]
- Zhang, P.C.; Wang, S.; Wu, Y.; Chen, R.Y.; Yu, D.Q. Five new diprenylated flavonols from the leaves of Broussonetia kazinoki. J. Nat. Prod. 2001, 64, 1206–1209. [Google Scholar] [CrossRef]
- Lee, H.; Li, H.; Jeong, J.H.; Noh, M.; Ryu, J.H. Kazinol B from Broussonetia kazinoki improves insulin sensitivity via Akt and AMPK activation in 3T3-L1 adipocytes. Fitoterapia 2016, 112, 90–96. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Kim, D.; Yoon, H.; Choi, C.S.; Oh, Y.S.; Jun, H.-S. Prevention of Oxidative Stress-Induced Pancreatic Beta Cell Damage by Broussonetia kazinoki Siebold Fruit Extract via the ERK-Nox4 Pathway. Antioxidants 2020, 9, 406. https://doi.org/10.3390/antiox9050406
Kim H-J, Kim D, Yoon H, Choi CS, Oh YS, Jun H-S. Prevention of Oxidative Stress-Induced Pancreatic Beta Cell Damage by Broussonetia kazinoki Siebold Fruit Extract via the ERK-Nox4 Pathway. Antioxidants. 2020; 9(5):406. https://doi.org/10.3390/antiox9050406
Chicago/Turabian StyleKim, Hyo-Jin, Donghee Kim, Haelim Yoon, Cheol Soo Choi, Yoon Sin Oh, and Hee-Sook Jun. 2020. "Prevention of Oxidative Stress-Induced Pancreatic Beta Cell Damage by Broussonetia kazinoki Siebold Fruit Extract via the ERK-Nox4 Pathway" Antioxidants 9, no. 5: 406. https://doi.org/10.3390/antiox9050406
APA StyleKim, H.-J., Kim, D., Yoon, H., Choi, C. S., Oh, Y. S., & Jun, H.-S. (2020). Prevention of Oxidative Stress-Induced Pancreatic Beta Cell Damage by Broussonetia kazinoki Siebold Fruit Extract via the ERK-Nox4 Pathway. Antioxidants, 9(5), 406. https://doi.org/10.3390/antiox9050406