Fermented Soy-Derived Bioactive Peptides Selected by a Molecular Docking Approach Show Antioxidant Properties Involving the Keap1/Nrf2 Pathway

 ,
,  , ,
, ,  , ,
, , 
Abstract
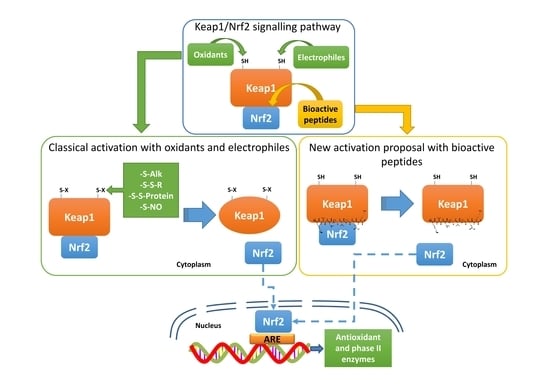
1. Introduction
2. Materials and Methods
2.1. Chemicals and Consumables
2.2. Preparation of Soy-Fermented Product
2.3. Simulated Gastrointestinal Digestion of Fermented Soy
2.4. Purification of Digested Fraction
2.5. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) Analysis
2.6. Database Search and Peptide Identification
2.7. Molecular Docking Analysis
2.8. Peptide Synthesis
2.9. Antioxidant Activities In Vitro with ABTS Tests
2.10. Cellular Studies
2.10.1. Caco-2 Cell Model
2.10.2. MTT Assay
2.10.3. Estimation of ROS Level
2.10.4. Caco-2 Cell Lysates
2.10.5. TrxR Activity
2.10.6. Nuclear Fraction and Nrf2 Detection by Western Blot
2.10.7. Quantification of Phase II Antioxidant Enzymes
2.10.8. Lipid Peroxidation Assay
2.10.9. Simulation of Intestinal Absorption
2.10.10. Reversed-Phase High-Performance Liquid Chromatography (RP-HPLC) Analysis
2.10.11. MS Analysis
2.10.12. Statistical Analysis
3. Results
3.1. Purification of Enriched Peptide Fractions
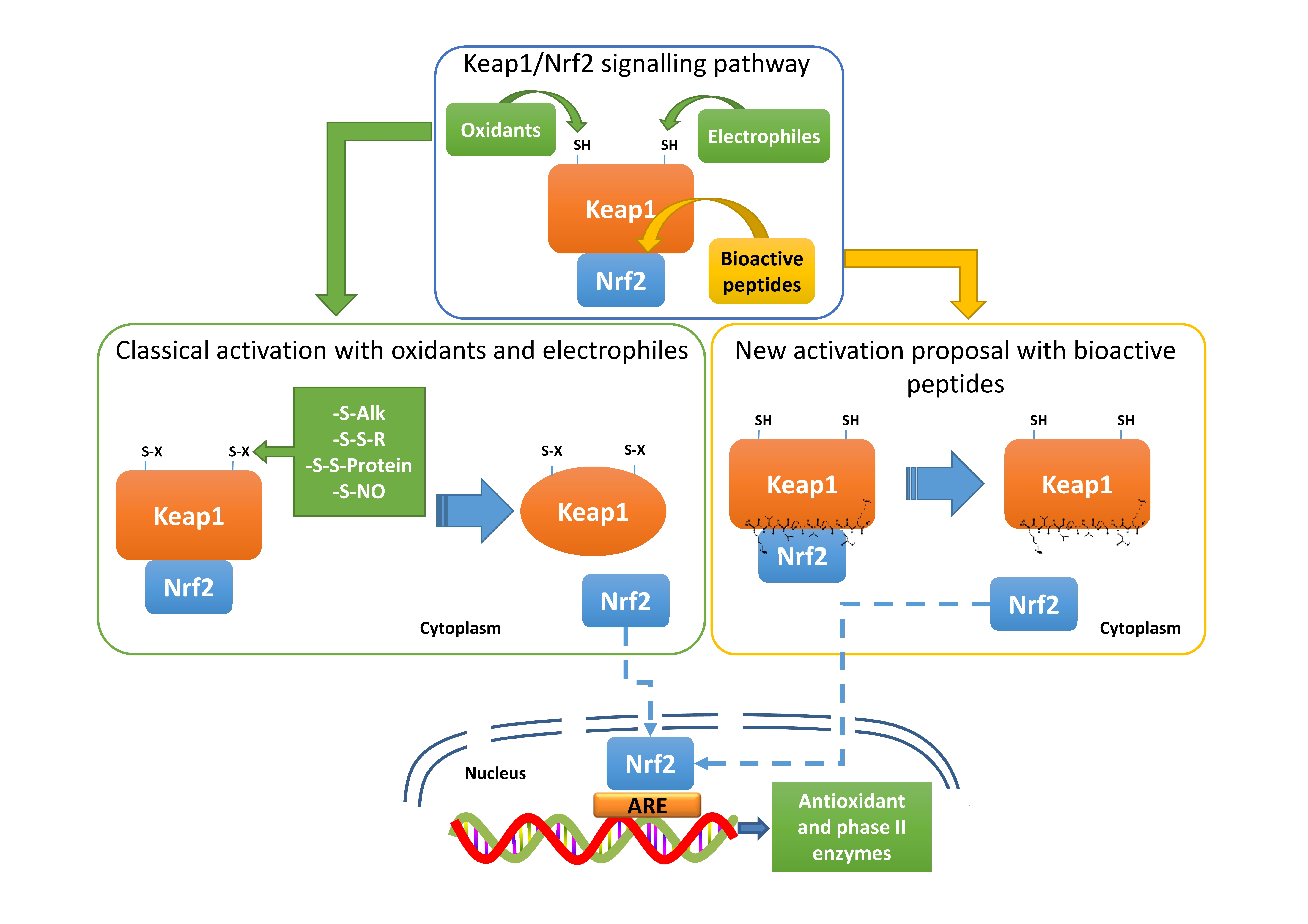
3.1.1. Antioxidant Activity of Peptide-Enriched Fractions In Vitro and in Caco-2 Cells
3.1.2. Simulation of Intestinal Absorption of Peptides
3.2. Identification of Bioactive Peptides in the Purified Fraction
3.3. Molecular Docking Analysis
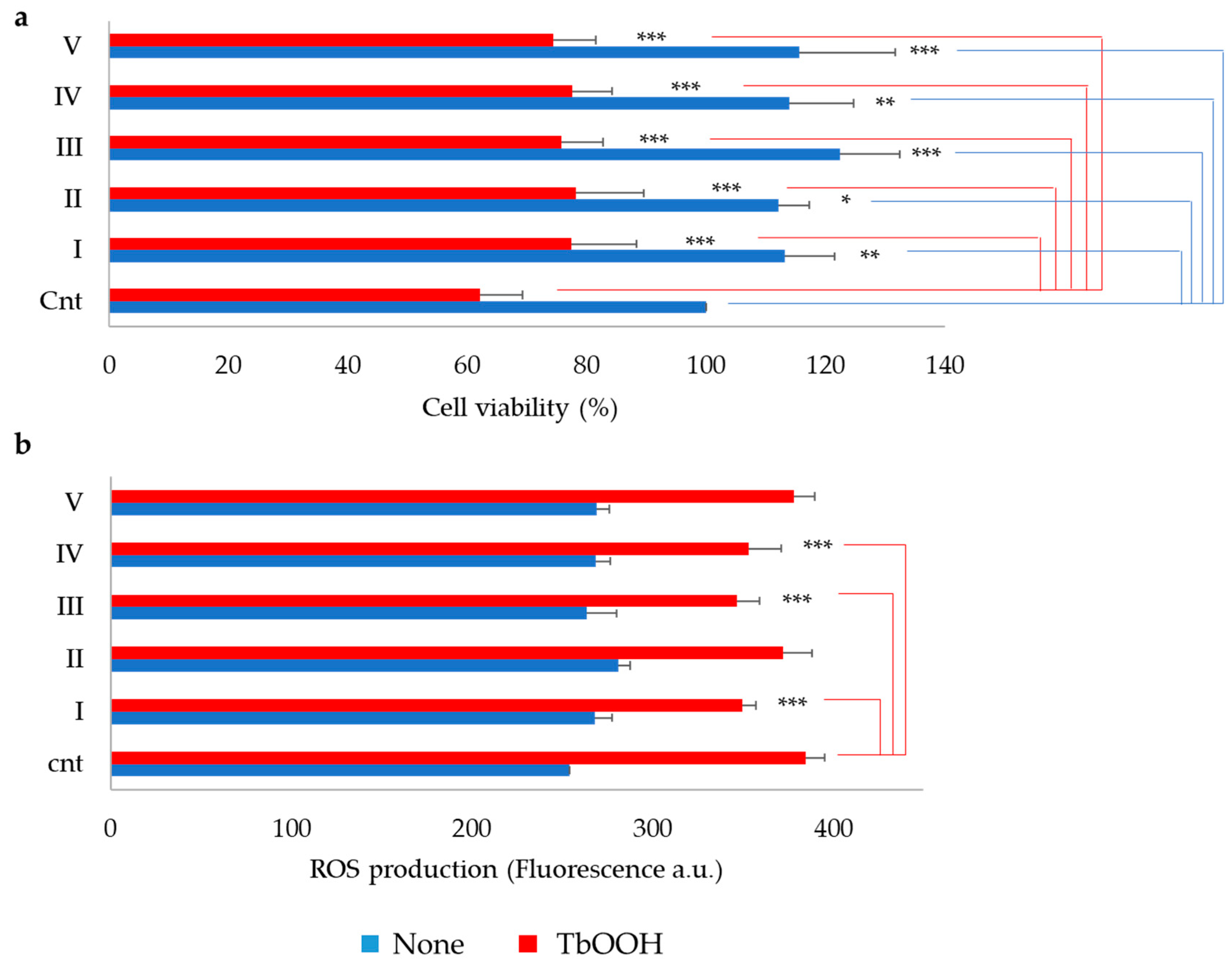
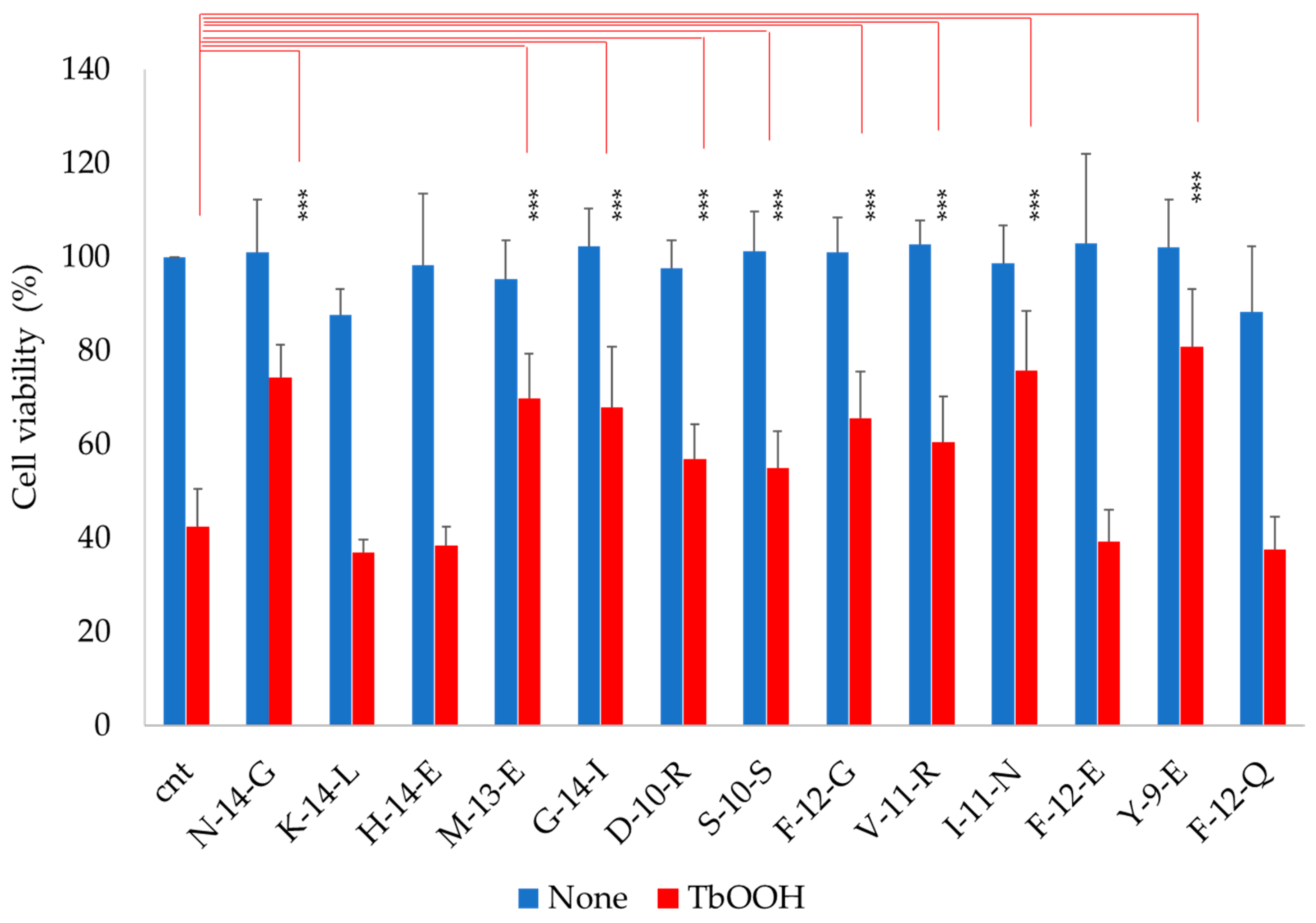
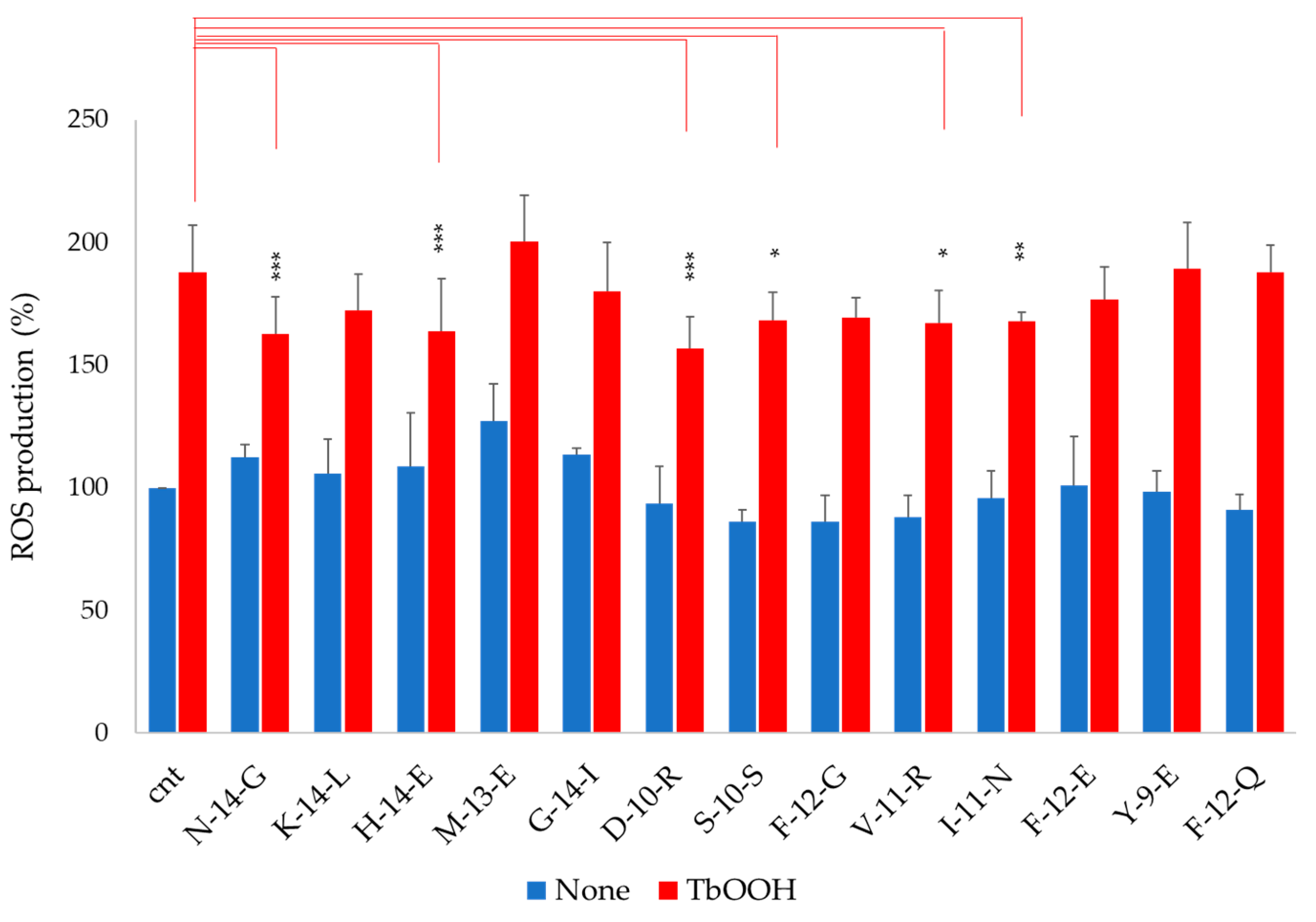
3.4. Antioxidant Capacity of Soy Peptides In Vitro and in Caco-2 Cell
3.5. Mechanism of Action of Antioxidant Peptides
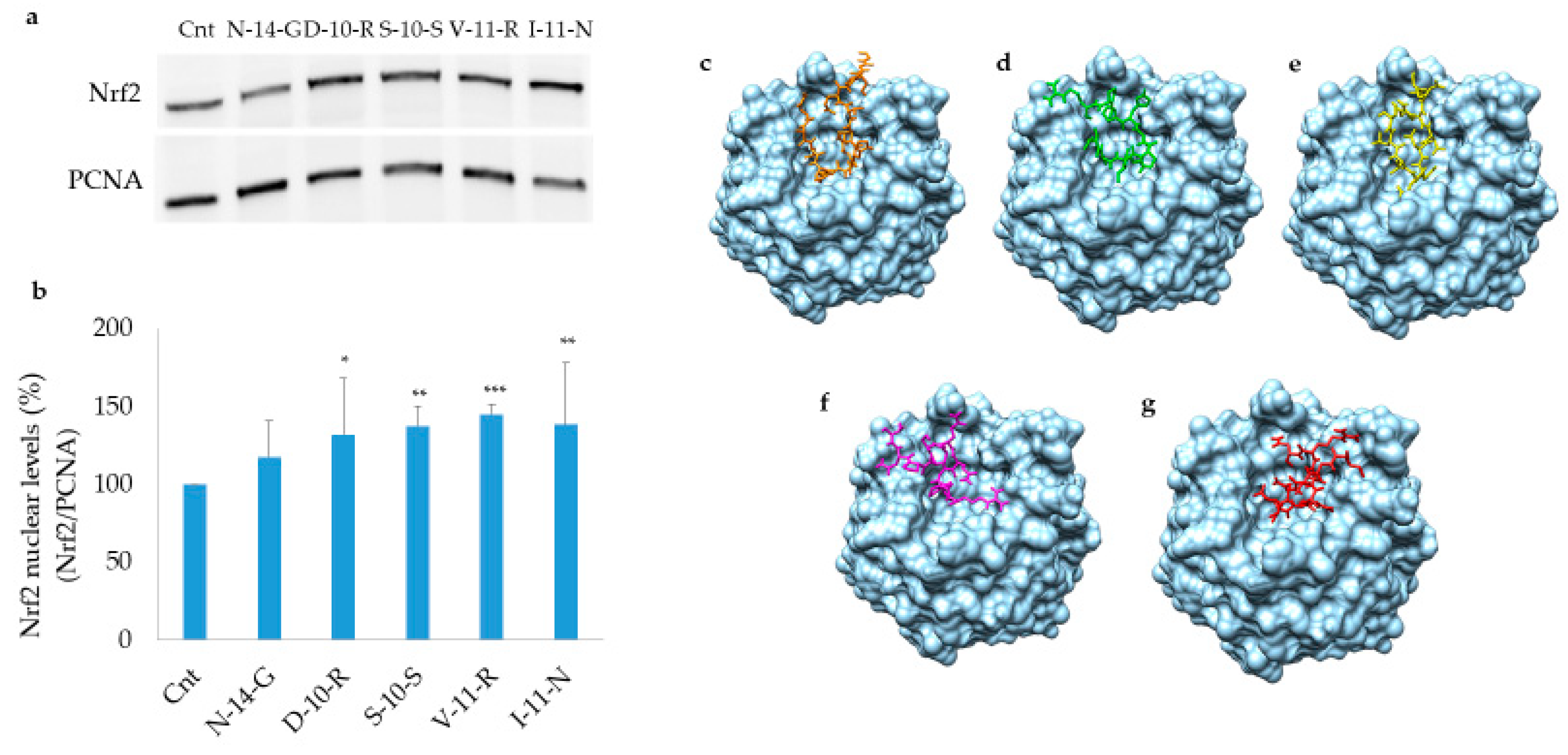
3.5.1. Keap1/Nrf2 Pathway Activation
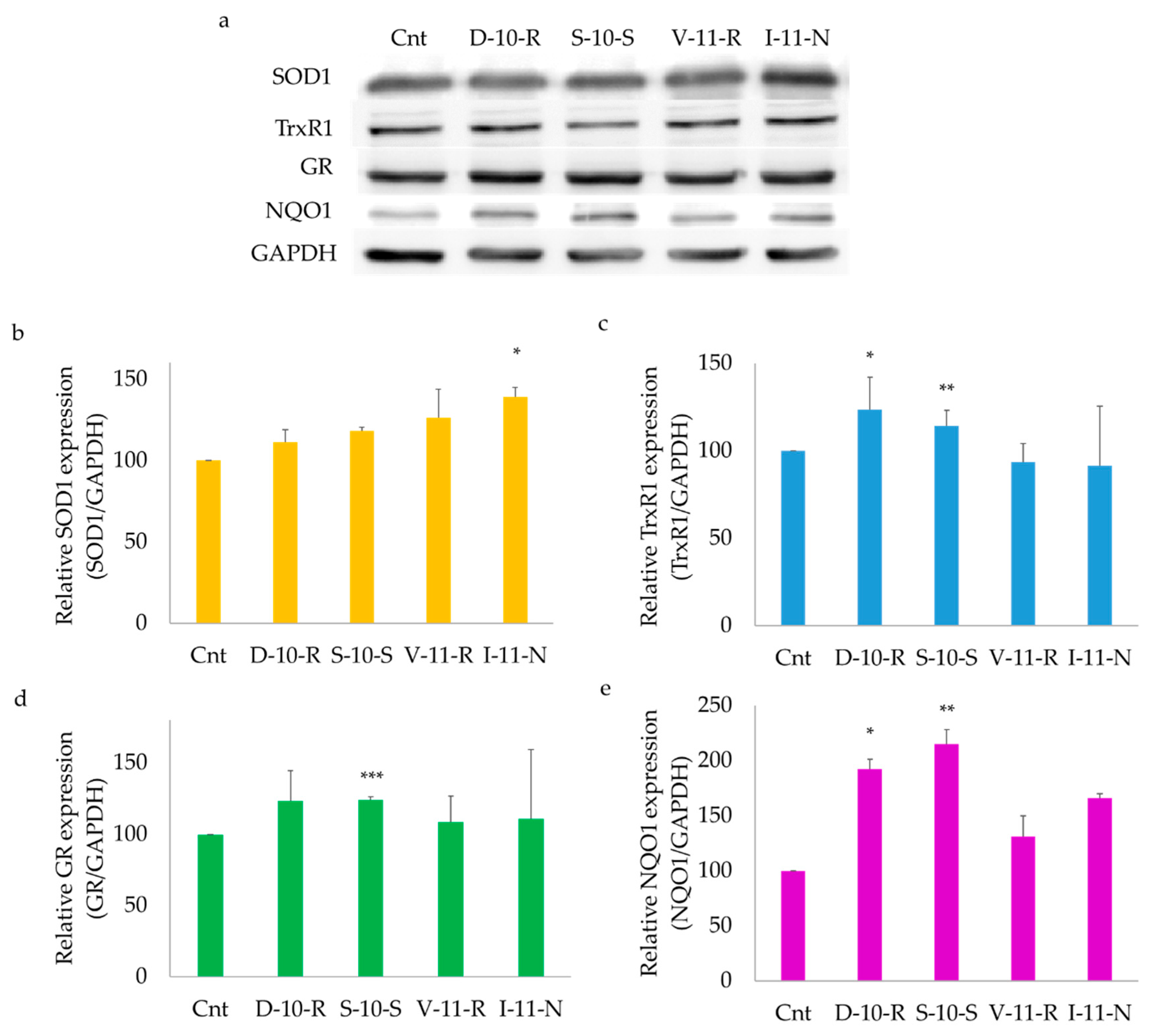
3.5.2. Overexpression of ARE-Regulated Enzymes
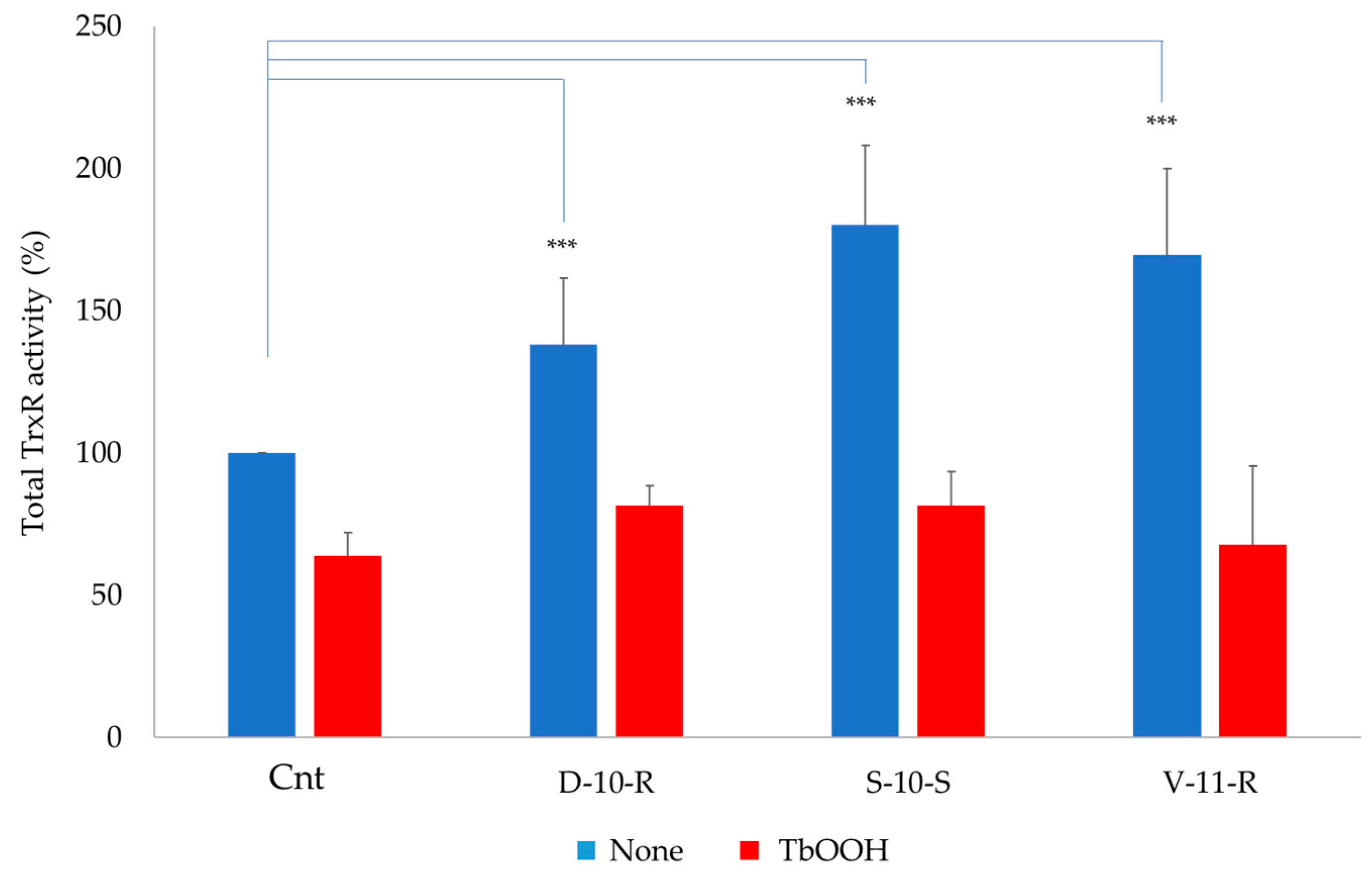
3.5.3. Total TrxR Enzymatic Activity
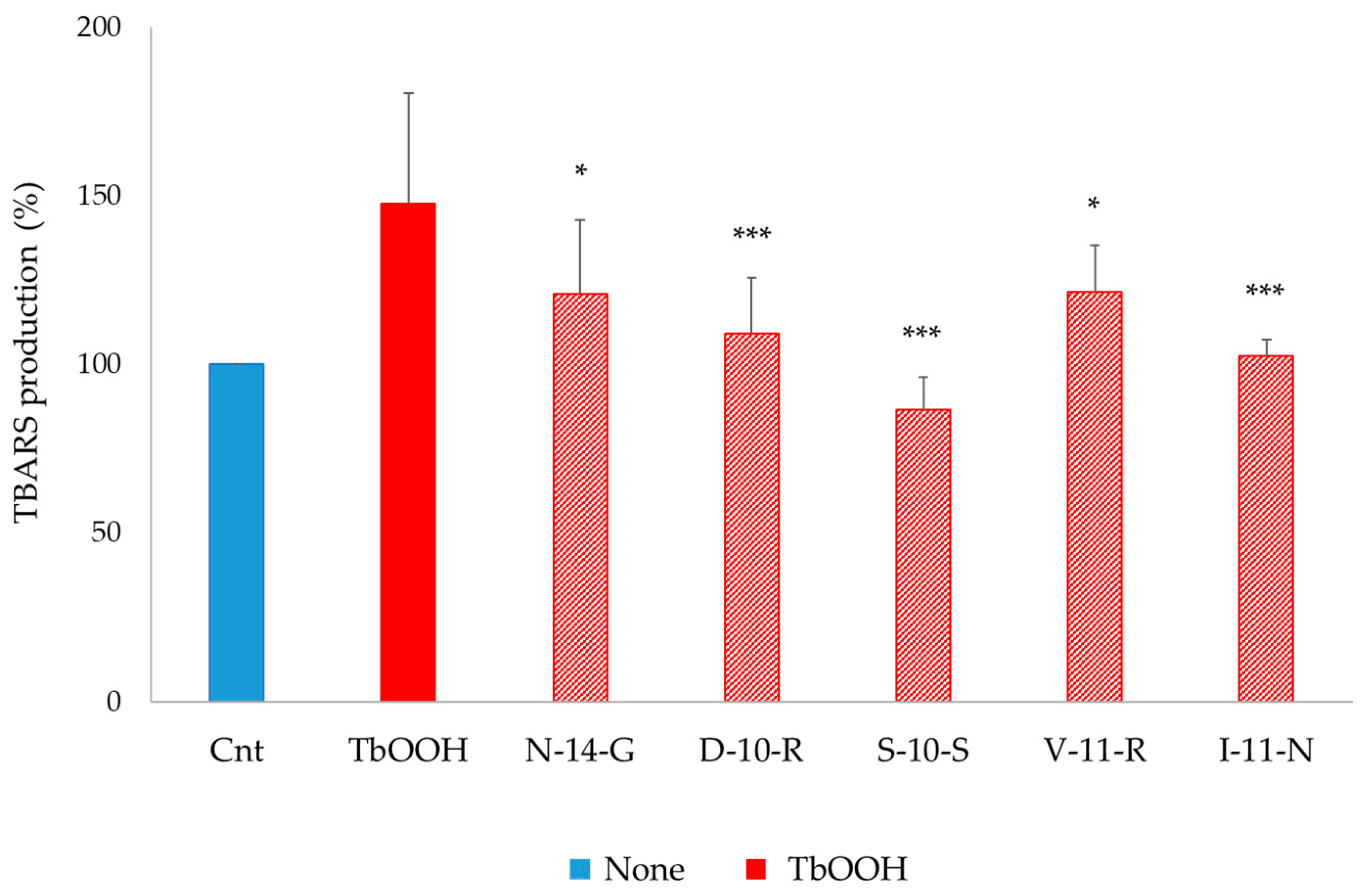
3.5.4. Lipid Peroxidation Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sethi, S.; Tyagi, S.K.; Anurag, R.K. Plant-based milk alternatives an emerging segment of functional beverages: A review. J. Food Sci. Technol. 2016, 53, 3408–3423. [Google Scholar] [CrossRef] [PubMed]
- Vanga, S.K.; Raghavan, V. How well do plant based alternatives fare nutritionally compared to cow’s milk? J. Food Sci. Technol. 2018, 55, 10–20. [Google Scholar] [CrossRef] [PubMed]
- El-Shemy, H. Soybean and Health; INTECH Open Access Publisher: Rijeka, Croatia, 2011; ISBN 978-953-307-535-8. [Google Scholar] [CrossRef]
- Hsieh, C.-C.; Fernández-Tomé, S.; Hernández-Ledesma, B. Chapter 27—Functionality of Soybean Compounds in the Oxidative Stress-Related Disorders. In Gastrointestinal Tissue; Gracia-Sancho, J., Salvadó, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 339–353. ISBN 978-0-12-805377-5. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Oh, D.H.; Lee, B.H. Bioactive Peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef]
- Singh, B.P.; Vij, S.; Hati, S. Functional significance of bioactive peptides derived from soybean. Peptides 2014, 54, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Battistini, C.; Gullón, B.; Ichimura, E.S.; Gomes, A.M.P.; Ribeiro, E.P.; Kunigk, L.; Moreira, J.U.V.; Jurkiewicz, C. Development and characterization of an innovative synbiotic fermented beverage based on vegetable soybean. Braz. J. Microbiol. 2018, 49, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Singh, B.P.; Vij, S. In Vitro stability of bioactive peptides derived from fermented soy milk against heat treatment, pH and gastrointestinal enzymes. LWT 2018, 91, 303–307. [Google Scholar] [CrossRef]
- Tonolo, F.; Moretto, L.; Folda, A.; Scalcon, V.; Bindoli, A.; Bellamio, M.; Feller, E.; Rigobello, M.P. Antioxidant Properties of Fermented Soy during Shelf Life. Plant Foods Hum. Nutr. 2019, 74, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Baird, L.; Dinkova-Kostova, A.T. The cytoprotective role of the Keap1–Nrf2 pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef] [PubMed]
- Tonolo, F.; Folda, A.; Cesaro, L.; Scalcon, V.; Marin, O.; Ferro, S.; Bindoli, A.; Rigobello, M.P. Milk-derived bioactive peptides exhibit antioxidant activity through the Keap1-Nrf2 signaling pathway. J. Funct. Food 2019. [Google Scholar] [CrossRef]
- Tonolo, F.; Fiorese, F.; Moretto, L.; Folda, A.; Scalcon, V.; Grinzato, A.; Ferro, S.; Arrigoni, G.; Bindoli, A.; Feller, E.; et al. Identification of New Peptides from Fermented Milk Showing Antioxidant Properties: Mechanism of Action. Antioxidants 2020, 9, 117. [Google Scholar] [CrossRef]
- Capriotti, A.L.; Caruso, G.; Cavaliere, C.; Samperi, R.; Ventura, S.; Chiozzi, R.Z.; Laganà, A. Identification of potential bioactive peptides generated by simulated gastrointestinal digestion of soybean seeds and soy milk proteins. J. Food Compos. Anal. 2015, 44, 205–213. [Google Scholar] [CrossRef]
- Kurcinski, M.; Jamroz, M.; Blaszczyk, M.; Kolinski, A.; Kmiecik, S. CABS-dock web server for the flexible docking of peptides to proteins without prior knowledge of the binding site. Nucleic Acids Res. 2015, 43, W419–W424. [Google Scholar] [CrossRef]
- Lee, H.; Heo, L.; Lee, M.S.; Seok, C. GalaxyPepDock: A protein–Peptide docking tool based on interaction similarity and energy optimization. Nucleic Acids Res. 2015, 43, W431–W435. [Google Scholar] [CrossRef]
- Dominguez, C.; Boelens, R.; Bonvin, A.M.J.J. HADDOCK: A Protein−Protein Docking Approach Based on Biochemical or Biophysical Information. J. Am. Chem. Soc. 2003, 125, 1731–1737. [Google Scholar] [CrossRef]
- van Zundert, G.C.P.; Rodrigues, J.P.G.L.M.; Trellet, M.; Schmitz, C.; Kastritis, P.L.; Karaca, E.; Melquiond, A.S.J.; van Dijk, M.; de Vries, S.J.; Bonvin, A.M.J.J. The HADDOCK2.2 Web Server: User-Friendly Integrative Modeling of Biomolecular Complexes. J. Mol. Biol. 2016, 428, 720–725. [Google Scholar] [CrossRef]
- Krissinel, E.; Henrick, K. Inference of Macromolecular Assemblies from Crystalline State. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Fields, G.B.; Noble, R.L. Solid phase peptide synthesis utilizing 9-fluorenylmethoxycarbonyl amino acids. Int. J. Pept. Protein Res. 1990, 35, 161–214. [Google Scholar] [CrossRef] [PubMed]
- Carpino, L.A.; Imazumi, H.; El-Faham, A.; Ferrer, F.J.; Zhang, C.; Lee, Y.; Foxman, B.M.; Henklein, P.; Hanay, C.; Mügge, C.; et al. The Uronium/Guanidinium Peptide Coupling Reagents: Finally the True Uronium Salts. Angew. Chem. Int. Ed. 2002, 41, 441–445. [Google Scholar] [CrossRef]
- Tonolo, F.; Moretto, L.; Ferro, S.; Folda, A.; Scalcon, V.; Sandre, M.; Fiorese, F.; Marin, O.; Bindoli, A.; Rigobello, M.P. Insight into antioxidant properties of milk-derived bioactive peptides in vitro and in a cellular model. J. Pept. Sci. 2019, 25, e3162. [Google Scholar] [CrossRef] [PubMed]
- Tonolo, F.; Sandre, M.; Ferro, S.; Folda, A.; Scalcon, V.; Scutari, G.; Feller, E.; Marin, O.; Bindoli, A.; Rigobello, M.P. Milk-derived bioactive peptides protect against oxidative stress in a Caco-2 cell model. Food Funct. 2018, 9, 1245–1253. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER measurement techniques for in vitro barrier model systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Peptide Name | Native Protein | MW (Da) Monoisotopic | Number aa | #PSM | Ion Score |
|---|---|---|---|---|---|---|
| YLAGNQEQE | Y-9-E | Glycinin G1–G2 | 1050.45 | 9 | 17 | 54 |
| IKPPTDEQQQRPQE | I-14-E | Glycinin G1 | 1692.83 | 14 | 2 | 41 |
| GKHQQEEENEGGSI | G-14-I | Glycinin G1 | 1540.66 | 14 | 1 | 23 |
| RQNIGQNSSPD | R-11-D | Glycinin G2 | 1214.55 | 11 | 2 | 41 |
| YQEPQESQQRG | Y-11-G | Glycinin G2 | 1348.59 | 11 | 1 | 55 |
| RNLQGENEEEDSGA | R-14-A | Glycinin G2 | 1546.63 | 14 | 1 | 52 |
| MRKPQQEEDDDDE | M-13-E | Glycinin G2 | 1633.64 | 13 | 1 | 41 |
| NNQLDQTPR | N-9-R | Glycinin G4 | 1084.51 | 9 | 2 | 50 |
| DEQIPSHPPR | D-10-R | Glycinin G4 | 1174.56 | 10 | 1 | 37 |
| VNPESQQGSPR | V-11-R | Glycinin G4 | 1197.56 | 11 | 3 | 79 |
| WQEQQDEDEDE | W-11-E | Glycinin G4 | 1449.50 | 11 | 1 | 44 |
| NALEPDHRVESEGG | N-14-G | Glycinin G4 | 1508.67 | 14 | 1 | 49 |
| KQGQHQQEEEEEGGSV | K-16-V | Glycinin G4 | 1797.76 | 16 | 2 | 51 |
| SLVNNDDRDS | S-10-S | β-conglycinin α subunit 2 | 1133.48 | 10 | 3 | 48 |
| VGLKEQQQEQ | V-10-Q | β-conglycinin α subunit 2 | 1185.58 | 10 | 1 | 44 |
| IGINAENNQRN | I-11-N | β-conglycinin α subunit 2 | 1241.60 | 11 | 1 | 67 |
| VGLKEQQQEQQ | V-11-Q | β-conglycinin α subunit 2 | 1313.64 | 11 | 1 | 37 |
| FSREEGQQQGEQ | F-12-Q | β-conglycinin α subunit 2 | 1421.60 | 12 | 2 | 59 |
| KEQQQEQQQEEQPL | K-14-L | β-conglycinin α subunit 2 | 1768.81 | 14 | 1 | 58 |
| QREEQEWPRKEEK | Q-13-K | β-conglycinin α subunit 2 | 1770.85 | 13 | 1 | 25 |
| TLVNNDDRDS | T-10-S | β-conglycinin α’ subunit 2/β subunit 2 | 1147.49 | 10 | 2 | 45 |
| FVDAQPQQKEEG | F-12-G | β-conglycinin α’ subunit 2/β subunit 2 | 1374.63 | 12 | 1 | 48 |
| FGREEGQQQGEE | F-12-E | β-conglycinin α ’subunit | 1392.58 | 12 | 1 | 60 |
| HEQKEEHEWHRKEE | H-14-E | β-conglycinin α ’subunit | 1929.86 | 14 | 1 | 12 |
| QRIPAGTT | Q-8-T | β-conglycinin β subunit 2 | 842.45 | 8 | 1 | 31 |
| LKVREDENNP | L-10-P | β-conglycinin β subunit 2 | 1212.59 | 10 | 1 | 30 |
| NKVDENGTPKPS | N-12-S | Lectin | 1284.62 | 12 | 4 | 57 |
| FNENESGDQV | F-10-V | Lectin | 1137.44 | 10 | 1 | 61 |
| Sequence | Peptide | MW (Da) | Class of Amino Acids (%) | pI (Theoretical) | ||||
|---|---|---|---|---|---|---|---|---|
| Basic | Acid | Neutral | Hydrophobic | |||||
| DEQIPSHPPR | D-10-R | 1174.56 | 20 | 20 | 50 | 10 | 5.32 | |
| NALEPDHRVESEGG | N-14-G | 1508.67 | 14.29 | 28.57 | 35.71 | 21.43 | 4.4 | |
| SLVNNDDRDS | S-10-S | 1133.48 | 10 | 30 | 40 | 20 | 3.93 | |
| KEQQQEQQQEEQPL | K-14-L | 1768.81 | 7.14 | 28.57 | 57.14 | 7.14 | 4.09 | |
| FVDAQPQQKEEG | F-12-G | 1374.63 | 8.33 | 25 | 41.67 | 25 | 4.14 | |
| VNPESQQGSPR | V-11-R | 1197.56 | 9.09 | 9.09 | 72.73 | 9.09 | 5.97 | |
| IGINAENNQRN | I-11-N | 1241.60 | 9.09 | 9.09 | 54.55 | 27.27 | 6 | |
| FGREEGQQQGEE | F-12-E | 1392.58 | 8.33 | 33.33 | 50 | 8.33 | 4.09 | |
| HEQKEEHEWHRKEE | H-14-E | 1929.86 | 42.86 | 42.86 | 7.14 | 7.14 | 5.39 | |
| MRKPQQEEDDDDE | M-13-E | 1633.64 | 15.38 | 53.85 | 23.08 | 7.69 | 3.9 | |
| GKHQQEEENEGGSI | G-14-I | 1540.66 | 14.29 | 28.57 | 50 | 7.14 | 4.48 | |
| YLAGNQEQE | Y-9-E | 1050.45 | 0 | 22.22 | 44.44 | 33.33 | 3.79 | |
| YQEPQESQQRG | Y-11-G | 1348.59 | 9.09 | 18.18 | 63.64 | 9.09 | 4.53 | |
| RQNIGQNSSPD | R-11-D | 1214.55 | 9.09 | 9.09 | 72.73 | 9.09 | 5.84 | |
| NNQLDQTPR | N-9-R | 1084.51 | 11.11 | 11.11 | 66.67 | 11.11 | 5.84 | |
| KQGQHQQEEEEEGGSV | K-16-V | 1797.76 | 12.5 | 31.25 | 50 | 6.25 | 4.32 | |
| VGLKEQQQEQ | V-10-Q | 1185.58 | 10 | 20 | 50 | 20 | 4.53 | |
| VGLKEQQQEQQ | V-11-Q | 1313.64 | 9.09 | 18.18 | 54.55 | 18.18 | 4.53 | |
| QRIPAGTT | Q-8-T | 842.45 | 12.5 | 0 | 62.5 | 25 | 9.75 | |
| LKVREDENNP | L-10-P | 1212.59 | 20 | 30 | 30 | 20 | 4.68 | |
| NKVDENGTPKPS | N-12-S | 1284.62 | 16.67 | 16.67 | 58.33 | 8.33 | 6.07 | |
| FNENESGDQV | F-10-V | 1137.44 | 0 | 30 | 50 | 20 | 3.57 | |
| RNLQGENEEEDSGA | R-14-A | 1546.63 | 7.14 | 35.71 | 42.86 | 14.29 | 3.91 | |
| IKPPTDEQQQRPQE | I-14-E | 1692.83 | 14.29 | 21.43 | 57.14 | 7.14 | 4.68 | |
| FSREEGQQQGEQ | F-12-Q | 1421.60 | 8.33 | 25 | 58.33 | 8.33 | 4.25 | |
| WQEQQDEDEDE | W-11-E | 1449.50 | 0 | 63.64 | 27.27 | 9.09 | 3.26 | |
| TLVNNDDRDS | T-10-S | 1147.49 | 10 | 30 | 40 | 20 | 3.93 | |
| QREEQEWPRKEEK | Q-13-K | 1770.85 | 30.77 | 38.46 | 23.08 | 7.69 | 5.07 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonolo, F.; Moretto, L.; Grinzato, A.; Fiorese, F.; Folda, A.; Scalcon, V.; Ferro, S.; Arrigoni, G.; Bellamio, M.; Feller, E.; et al. Fermented Soy-Derived Bioactive Peptides Selected by a Molecular Docking Approach Show Antioxidant Properties Involving the Keap1/Nrf2 Pathway. Antioxidants 2020, 9, 1306. https://doi.org/10.3390/antiox9121306
Tonolo F, Moretto L, Grinzato A, Fiorese F, Folda A, Scalcon V, Ferro S, Arrigoni G, Bellamio M, Feller E, et al. Fermented Soy-Derived Bioactive Peptides Selected by a Molecular Docking Approach Show Antioxidant Properties Involving the Keap1/Nrf2 Pathway. Antioxidants. 2020; 9(12):1306. https://doi.org/10.3390/antiox9121306
Chicago/Turabian StyleTonolo, Federica, Laura Moretto, Alessandro Grinzato, Federico Fiorese, Alessandra Folda, Valeria Scalcon, Stefania Ferro, Giorgio Arrigoni, Marco Bellamio, Emiliano Feller, and et al. 2020. "Fermented Soy-Derived Bioactive Peptides Selected by a Molecular Docking Approach Show Antioxidant Properties Involving the Keap1/Nrf2 Pathway" Antioxidants 9, no. 12: 1306. https://doi.org/10.3390/antiox9121306
APA StyleTonolo, F., Moretto, L., Grinzato, A., Fiorese, F., Folda, A., Scalcon, V., Ferro, S., Arrigoni, G., Bellamio, M., Feller, E., Bindoli, A., Marin, O., & Rigobello, M. P. (2020). Fermented Soy-Derived Bioactive Peptides Selected by a Molecular Docking Approach Show Antioxidant Properties Involving the Keap1/Nrf2 Pathway. Antioxidants, 9(12), 1306. https://doi.org/10.3390/antiox9121306