Abstract
Cancer remains an elusive, highly complex disease and a global burden. Constant change by acquired mutations and metabolic reprogramming contribute to the high inter- and intratumor heterogeneity of malignant cells, their selective growth advantage, and their resistance to anticancer therapies. In the modern era of integrative biomedicine, realizing that a personalized approach could benefit therapy treatments and patients’ prognosis, we should focus on cancer-driving advantageous modifications. Namely, reactive oxygen species (ROS), known to act as regulators of cellular metabolism and growth, exhibit both negative and positive activities, as do antioxidants with potential anticancer effects. Such complexity of oxidative homeostasis is sometimes overseen in the case of studies evaluating the effects of potential anticancer antioxidants. While cancer cells often produce more ROS due to their increased growth-favoring demands, numerous conventional anticancer therapies exploit this feature to ensure selective cancer cell death triggered by excessive ROS levels, also causing serious side effects. The activation of the cellular NRF2 (nuclear factor erythroid 2 like 2) pathway and induction of cytoprotective genes accompanies an increase in ROS levels. A plethora of specific targets, including those involved in thioredoxin (TRX) and glutathione (GSH) systems, are activated by NRF2. In this paper, we briefly review preclinical research findings on the interrelated roles of the NRF2 pathway and TRX and GSH systems, with focus given to clinical findings and their relevance in carcinogenesis and anticancer treatments.
1. Introduction
According to the World Health Organization, cancer is the second leading cause of death, accounting for 9.56 million deaths and an incidence of 18.1 million new cases in 2018, with the GLOBOCAN estimation of 29.5 million new cases and 16.4 million deaths by 2040 [1]. Consequently, cancer remains a global burden and an elusive, ever-changing disease with an extremely complex biology. The high inter- and intratumor heterogeneity is governed by cancer’s need to grow and spread in the organism while surviving therapy-induced unfavorable conditions. Genetic alterations inducing survival pathways combined with metabolic reprogramming ensure the action of diverse players contributing to tumor development and progression.
Reactive oxygen species (ROS) are recognized as essential players contributing to tumorigenesis or suppressing it, with an important role in anticancer therapy as well. Although previously perceived as exogenously and endogenously derived byproducts of cellular metabolism with signaling abilities that affect cellular functions in a concentration-dependent manner, the perception of ROS has been upgraded since. Nowadays, they are considered as cellular tools that are, upon certain stimuli, purposely produced by the cell in an amount that will elicit a specific feedback reaction within the cell or in neighboring ones [2]. The tuning of ROS levels is intertwined with metabolism and antioxidative machinery, as in normal cells and cancer cells, too. However, cancer cells often have more ROS than normal ones, a feature that is exploited by most conventional chemo- and radiotherapy.
The nuclear factor, erythroid 2 like 2 (NRF2) pathway is the main pathway activated upon ROS production. Its activation induces the expression of over 250 genes, regulating diverse processes from redox homeostasis to detoxification, carbohydrate and lipid metabolism, autophagy, apoptosis, DNA repair, etc. [3]. The antioxidant mechanisms involved in ROS balancing, such as thioredoxin (TRX) and glutathione (GSH) systems, not only protect from carcinogenesis but can support it as well. A recent clinical study on colorectal cancer patients demonstrated that redox biomarkers could have a role in the infiltration of inflammatory cells and tumor budding [4]. The same study also demonstrated that the levels of oxidative stress parameters vary based on the stage of cancer. In addition, GSH and TRX are both depicted as important players in carcinogenesis. While GSH is required for cancer initiation, TRX is a predominant mechanism governing cancer progression by abolishing detrimental ROS levels in already established neoplasm [5]. Noteworthy, the NRF2 pathway is often activated in various types of cancer, emphasizing its dual role in tumorigenesis.
Only the understanding of all intertwined cancer-related components, particularly the contributors and/or suppressors of cancer growth, will bring us closer to better-targeted therapy. This review will explore the importance of the NRF2 pathway and TRX and GSH systems in carcinogenesis in relation to anticancer treatment strategies.
2. Reactive Oxygen Species—Friend or Foe?
ROS are continuously formed in aerobic organisms as byproducts of normal intra/intercellular metabolism or in response to adequate stimuli, as is the case of inflammation. Depending on the ROS type, reactivity, and their diffusion distance, their effect on macromolecules varies [6]. Thus, superoxide anion (O2●−) ROS are generated by the action of complex I and complex III via an electron transport chain in mitochondria [7], while membrane-bound nicotinamide adenine dinucleotide phosphate (NADPH) oxidase and lipoxygenases also contribute to O2●−. Additionally, huge amounts of ROS, in particular O2●−, are formed during phagocytosis [6]. The dismutation of O2●− occurs spontaneously or enzymatically, with superoxide dismutase (SOD) generating hydrogen peroxide (H2O2), which is a more stable non-radical form of ROS with a greater diffusion distance that may therefore affect targets distant from the initial oxidative insult. At low concentrations, ROS have important signaling roles both intra- and intercellularly; however, at higher concentrations they have toxic effects. Likewise, H2O2 is an important metabolic signal for insulin secretion in β-cells [8], however excessive H2O2 affects mitochondria and causes β-cell dysfunction [9,10]. Similarly, ROS generated during the respiratory burst of granulocytes can have dual roles in carcinogenesis [6]. Although an important anticancer effect of granulocytes was demonstrated in vitro [11,12], the in vivo results can be contradictory. In experimental animal tumor models, it was shown that ROS can reverse the process of tumor development, leading to spontaneous tumor regression [13,14], and can decelerate tumor growth, extending the survival of animals [15]. Opposite to that, it is known that ROS can also promote tumor development [13,14], so one also needs to consider the importance of intercellular redox signaling in tumor development. Hence, it was recently shown that also hypochlorous acid intercellular redox signaling is involved in granulocyte-mediated tumor inhibition [16].
One of the key mechanisms by which ROS achieve their biological effects is the well-documented induction of either reversible or irreversible modifications of proteins, altering their structure and cellular functions. For example, the oxidative modification of erythrocyte spectrin alpha chain, band 3 protein, and glyceraldehyde-3-phosphate dehydrogenase can affect the erythrocyte structure and energy metabolism [17]. Similarly, the oxidation of mitochondrial adenosine triphosphate synthase beta subunit can affect the enzyme catalytic activities and potentially have a detrimental role for neurons [18].
Both O2●− and H2O2 can serve as precursors for highly reactive hydroxyl radicals (●OH) that can induce direct oxidative damage to macromolecules. Transition metals, such as iron, can promote ROS and support tumorigenesis [19,20]. O2●− and H2O2 are accessible to iron via the Fenton reaction, converting them to ●OH, and can contribute to the initiation of ferroptosis [21], which is also considered in anticancer approaches as a potential target for oncotherapy [22].
On the other hand, ROS can also damage DNA, causing strand breaks or oxidative DNA lesions. As an example, the ●OH-induced nucleic acid oxidative damage can yield a variety of base lesions, among which 8-oxo-2′deoxyguanosine (8-OHdG) and 8-oxo-guanosine have received the most attention [23,24]. The accumulation of unrepaired or defectively repaired DNA damage could eventually promote various disorders, among which cancer is the prominent disease.
Moreover, the bis-allylic site of polyunsaturated fatty acids is highly susceptible to ROS induced damage. The peroxidation of lipids is profoundly triggered by ●OH and hydroperoxyl radical, yielding as final products reactive aldehydes, among which is the most potent “second messenger of ROS”, 4-hydroxynonenal (4-HNE). 4-HNE was first discovered fifty years ago and was thought to be a toxic product of lipid peroxidation [25]. Since then, a huge amount of effort has been put into investigating the roles of 4-HNE, and today we know that, depending on its location and concentration, it can act as a double-edged sword, exhibiting pathological or desirable physiological effects. The high reactivity of 4-HNE is due to its three functional groups: hydroxyl group, carbonyl group, and C=C double bond [26]. 4-HNE can act either directly or by forming adducts with macromolecules, such as proteins [27] or nucleic acids [28,29]. Consequently, 4-HNE can modulate various cellular functions of non-malignant as well as cancer cells [30,31,32,33] and can play a role in tumorigenesis [34,35,36].
3. Redox Homeostasis
Excessive ROS generation alters the cellular redox homeostasis built by antioxidant defense systems to cope with the negative effects of ROS. On the other side, endogenous antioxidant defense systems are affected by the supplementation of exogenous antioxidants, which is common in both healthy and ill people.
The major endogenous antioxidants responsible for cellular ROS detoxification are the TRX system, the GSH system, peroxiredoxin, SOD, and catalase (CAT). The removal of O2●− by SOD elevates cellular H2O2 that can act as a molecular mediator of various signaling pathways, such as insulin, NRF2, and the c-Jun N-terminal kinase (JNK) signaling pathway [10,37]. Whether H2O2 will promote cell survival or induce apoptosis depends on its concentration. Thus, cells have several mechanisms for H2O2 detoxification. Catalase, peroxiredoxin, and the TRX and GSH systems all promote the conversion of H2O2 to H2O and molecular oxygen. Although these might be considered as major cellular antioxidant defenses, one should bear in mind that other antioxidants—for example, protein deglycase DJ-1—can also determine the fate of the tumor, interact with NRF2, regulate GSH levels, and promote TRX (reviewed in [38]).
Therefore, alterations in redox homeostasis are of high importance for cancer development. Although ROS have an important role in the initiation and promotion of tumor development, they are also crucial factors that can inhibit tumor viability and growth, so tumor cells modify their own antioxidant network to enable them to escape the anticancer effects of ROS. Among endogenous antioxidants in the current review, special attention is given to the NRF2, TRX, and GSH antioxidant defense mechanisms in tumorigenesis.
3.1. The NRF2 Pathway in Tumorigenesis
The NRF2 pathway is the main pathway activated as a response to oxidative stress. In homeostasis, transcription factor NRF2, encoded by the NFE2L2 gene (nuclear factor, erythroid 2 like 2) is mainly under the control of its repressor, Kelch-like ECH-associated protein 1 (KEAP1), and subjected to ubiquitination and proteasomal degradation [39]. Additionally, NRF2 can also be repressed by β-transducin repeat-containing protein (β-TRCP) in a glycogen synthase kinase-3 (GSK-3) -dependent and -independent manner [40] or in an HRD1 (E3 ubiquitin-protein ligase synoviolin, SYVN1) -dependent manner upon endoplasmic reticulum (ER) stress [41] (Figure 1). Aside from the mentioned mechanisms of NRF2 regulation, numerous proteins and molecules interact with either KEAP1 or NRF2 or even compete with NRF2, such as BTB and CNC homolog 1 (BACH1) [42], contributing to the highly complex regulation/activation of NRF2, the feature particularly accentuated in cancer (reviewed in [43,44,45,46,47]).
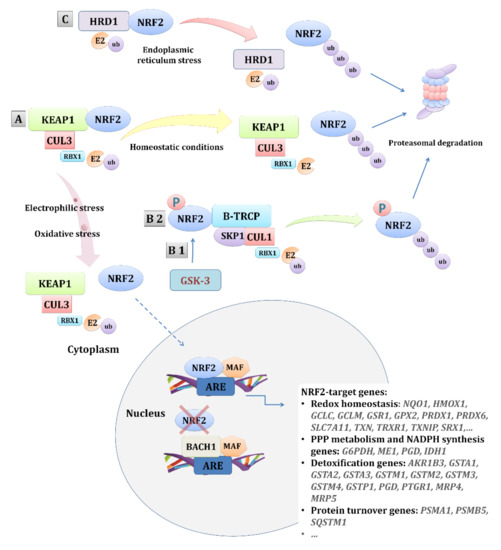
Figure 1.
Regulation of the NRF2 pathway. NRF2 is regulated by three E3 ubiquitin ligases. The main regulation is by the complex KEAP1-CUL3-RBX1 (A). Additionally, NRF2 can be regulated by the β-TRCP-SKP1-CUL1-RBX1 complex in a GSK-3-dependant (B1) or -independent (B2) manner. Upon endoplasmic reticulum stress, NRF2 can also be regulated by HRD1 (C). The main regulation (A) involves the repression of NRF2 by KEAP1 and its proteasomal degradation in basal conditions, while, under oxidative stress conditions, the NRF2-KEAP1 binding is disrupted, leading to NRF2 nuclear translocation and the activation of its target genes. Other mechanisms of NRF2 repression involve the β-TRCP-SKP1-CUL1-RBX1 complex (B 1/2) that, through binding with the Neh6 domain of NRF2, particularly enhanced by the GSK-3 phosphorylation of NRF2, or, by the binding of endoplasmic reticulum stress-induced HRD1 (C) with the Neh4-5 domains of NRF2, mediate its degradation. In addition, other proteins, such as BACH1, can negatively regulate the transcriptional activation of the NRF2-target genes by competing with NRF2. Abbreviations: AKR1B3—aldo-keto reductase family 1 member B; ARE—antioxidant response element; BACH1—BTB and CNC homolog 1; β-TRCP—β-transducin repeat-containing protein; CUL1—cullin 1; CUL3—cullin 3; E2—ubiquitin-conjugating enzyme 2; G6PDH—glucose-6-phosphate dehydrogenase; GCLC—glutamate-cysteine ligase, catalytic subunit; GCLM—glutamate-cysteine ligase, modifier subunit; GPX2—glutathione peroxidase 2; GSR1—glutathione reductase 1; GSTA—glutathione S-transferase alpha; GSTM—glutathione S-transferase mu; GSTP—glutathione S-transferase pi; HMOX-1—heme oxygenase 1; HRD1—E3 ubiquitin-protein ligase synoviolin, SYVN1; IDH1—isocitrate dehydrogenase 1; KEAP1—Kelch-like ECH-associated protein 1; MAF—musculoaponeurotic fibrosarcoma; ME1—malic enzyme 1; MRP—multidrug resistance-associated proteins; NQO1—NAD(P)H quinone dehydrogenase 1; NRF2—nuclear factor, erythroid 2 like 2; PGD—phosphogluconate dehydrogenase; PRDX—peroxiredoxin; PSMA1—proteasome 20S subunit alpha 1; PSMB5—proteasome 20S subunit beta 5; PTGR1—prostaglandin reductase 1; RBX1—ring-box 1; SKP1—S-phase kinase-associated protein 1; SLC7A11—solute carrier family 7 member 11; SQSTM1—sequestosome 1.; SRX1—sulfiredoxin 1; TRXR1—thioredoxin reductase 1; TXN—thioredoxin; TXNIP—thioredoxin interacting protein; ub—ubiquitin.
Current research recognizes the dual role of NRF2 in tumorigenesis. NRF2 was first perceived as a tumor suppressor due to its role in reducing damaging ROS and environmental carcinogens, thus protecting cells from potential neoplastic transformation. Mice Nrf2-knockout models have pointed out the importance of the NRF2 pathway activation in the suppression of carcinogen-induced cancer initiation (e.g., in the skin [48], breast [49], and forestomach [50]). In humans, an NRF2 single nucleotide polymorphism (SNP) homozygous allele (-617A7A) exhibits a decreased expression of NRF2 and its target cytoprotective products, and is consequently associated with an increased risk of lung cancer in smokers [51]. In addition, the genetic variants in NFE2L2, NAD(P)H quinone dehydrogenase 1 (NQO1), nitric oxide synthase 3 (NOS3), and heme oxygenase 1 (HMOX-1), exhibit lower ROS detoxification capabilities—although they are not associated with a breast cancer risk individually—in postmenopausal women with three or more of such modifications, increasing the risk of breast cancer, especially when combined with a high iron intake [52].
Conversely, the discovery of the hyperactivation of NRF2 in quite a number of tumors, conferring an advantage to tumor cells and resulting in growth promotion and therapy resistance, revealed its new role as an oncogene. In this context, the constitutive activation of NRF2 contributes not only to the progression and chemoresistance in the already-established tumor cells but also to the tumor development itself. Indeed, the constitutive activation of NRF2, associated with an increase in cellular antioxidant enzymes and a diminution in the ROS levels, contributes to arsenite-induced human bronchial epithelial cell transformation [53].
Genetic alterations (e.g., NFE2L2 gain-of-function mutations, KEAP1 loss-of-function mutations, and co-occurring driver mutations), interactions with different proteins interfering with the NRF2-KEAP1 bond, and epigenetic and posttranslational modifications can all induce NRF2 activation. Emerging data imply the varying involvement of the NRF2 pathway among tumors (some are listed in Table 1). Cancers with NRF2 hyperactivating mutations share a set of upregulated target genes, including key regulators involved in the thioredoxin and glutathione system, metabolic enzymes, transporters, and others, enabling a cancer growth advantage and consequently a worse overall survival. The upregulation of these genes by NRF2 highly depends upon a more assessable chromatin environment of antioxidant responsive elements (AREs) [54].

Table 1.
The involvement of the NRF2 pathway in diverse types of tumors.
In lung cancer, the co-occurring mutations in KEAP1, NFE2L2, or cullin 3 (CUL3) observed in 7% of non-small cell lung cancer (NSCLC) patients with EGFR (Epidermal Growth Factor Receptor) mutations indicate the activation of the NRF2 pathway as an acquired mechanism, leading to the resistance to usually beneficial EGFR-tyrosine kinase inhibitor therapy [55]. Another protective mechanism observed in lung cancers is metabolic reprogramming to glutaminolysis, mostly observed in KRAS (Kirsten rat sarcoma viral oncogene homolog)-mutant lung adenocarcinoma, which exerts a loss of STK11/LKB1 (serine/threonine kinase 11, liver kinase B1) and co-occurring KEAP1 mutation [56]. In addition, the NRF2 molecular signature, including genes important in activating TRX and GSH systems such as glutathione peroxidase (GPX) 3 and thioredoxin-like 1, is suggested as an excellent predictor of cancer remission and overall survival in patients with lung cancer [57]. The growth advantage and resistance to therapy in NSCLC patients with a worse prognosis are associated with the activation of the NRF2 pathway, contributing to a higher expression of multidrug-resistant protein-3 [58].
The other mechanisms of NRF2 activation include diverse proteins that compete with NRF2 in binding with KEAP1. Examples of these are proteins with an ETGE motif, such as dipeptidyl peptidase 3 protein [59], cell cycle-related kinase (CDK20), as well as others. The overexpression of CDK20 in lung cancer leads to tumor progression and resistance to radio and chemotherapies [60]. The p62 is another NRF2-binding competitor of KEAP1. Its upregulation was suggested as a protection mechanism of hepatocellular carcinoma (HCC)-initiating cells in the oxidative stress hostile environment that promotes HCC carcinogenesis [61], as well as anticancer drug tolerance in tumor regions positive for the hepatitis C virus [62]. The alkylation of KEAP1 by succinylacetone [63] or the hypermethylation of the KEAP1 promoter region may lead to NRF2 activation. For instance, the tumor-specific hypermethylation of the KEAP1 promoter region was suggested as a specific feature of a clear cell renal carcinoma [64].
Crosstalk between the NRF2 pathway and other proteins can contribute to carcinogenesis and therapy resistance. Hence, it was shown that the progression of HCC requires metabolic changes involving TRAP1 and NRF2 as an early event, leading to an expression pattern of glucose-6-phosphate dehydrogenase that correlates with grading, metastasis, and poor prognosis [65]. Another example of this is the overexpression of TRIM25 (Tripartite motif-containing protein 25) and NRF2, associated with the protection of HCC cells upon ER-induced ROS [66]. The association of BACH1 and NRF2 was shown to promote lung cancer metastasis. Mechanistically, the mutations in KEAP1 found in lung adenocarcinoma patients lead to the activation of the NRF2 pathway and an increase in HMOX-1. Since the degradation of BACH1 by F-box only protein 22 (FBOXO22) necessities heme, the NRF2-induced HMOX-1 reduces heme levels, thus stabilizing BACH1 and promoting the transcription of the pro-metastatic genes. The antioxidant supplementation of N-acetyl cysteine and vitamin E mimics the NRF2-HMOX-1 action, leading to the BACH1 stabilization and glycolysis induction in KRAS-mutant lung cancer. Therefore, lung adenocarcinoma patients with a high BACH1 signature are associated with increased metastasis and worsened survival [67,68]. Conversely, FBXO22 promotes carcinogenesis in colorectal cancer [69] and HCC [70] by degrading PTEN (phosphatidylinositol 3,4,5-trisphosphate 3-phosphatase and dual-specificity protein phosphatase) and p21, respectively, while revealing both roles (protumorigenic and antimetastatic) in breast cancer [71].
3.2. The Thioredoxin System and Thioredoxin-Domain-Containing Protein Family in Tumorigenesis
The thioredoxin system, one of the key regulators of cellular redox homeostasis, comprises TRX, thioredoxin reductase (TRXR), nicotinamide adenine dinucleotide phosphate (NADPH), and thioredoxin interacting protein (TXNIP) (Figure 2). The TRX system requires NADPH to reduce the oxidized form of TRX. Reduced TRX is needed for the recycling of oxidized thioredoxin peroxidase/peroxiredoxin [139]. The family of thioredoxin-domain containing proteins (TXNC) are also redox regulators, and today 17 members of the TXNDC family are known [140]. When an oxidized substrate is reduced by the action of Cys32 and Cys35 of the reduced dithiol form of TRX, oxidized disulfide TRX is generated. Oxidized TRX is converted back to an active reduced TRX by TRXR at the expense of NADPH [141]. Thus, an adequate supply of NADPH and TRXR activity is crucial for this process.
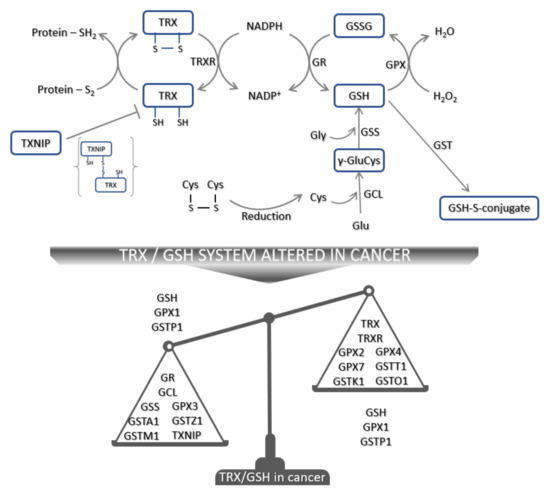
Figure 2.
Schematic presentation of TRX and GSH antioxidant defense mechanisms. Oxidized TRX is reduced to the active reduced form of TRX by TRXR that uses NADPH as the main electron source. Reduced TRX reduces oxidized substrates. However, this can be inhibited by TXNIP, resulting in TXNIP-TRX complex formation. Moreover, extracellular cystine once imported in the cell can be reduced by the TRX system and used for GSH synthesis. GCL catalyzes the formation of γ-GluCys from Cys and Glu. Further addition of Gly by GSS produces GSH. During the detoxification of ROS, GSH is converted to GSSG and its recycling depends on GR and NADPH. In addition, during the detoxification of other molecules—e.g., 4-HNE—GSH conjugation by GST is the main step. In cancer, TRX and/or GSH systems are altered and their expression in cancer is schematically presented. Abbreviations: Cys—cysteine, γ-GluCys—gamma glutamylcysteine, GCL—glutamate cysteine ligase, Glu—glutamine, Gly—glycine, GSH—glutathione, GSS—glutathione synthetase, GPX—glutathione peroxidase, GR—glutathione reductase, GSH—reduced glutathione, GSSG—oxidized glutathione, GST—glutathione S-transferase, NADP+—oxidized NADPH, NADPH—nicotinamide adenine dinucleotide phosphate, TXNIP—thioredoxin interacting protein, TRX—thioredoxin, TRXR—thioredoxin reductase.
In the last few decades, numerous studies have investigated the role of the TRX system and some members of the TXNDC protein family in tumorigenesis (Table 2). The overexpression and hyperactivation of cytoplasmic TRXR (TRXR1) have been reported for various cancer types, such as brain cancer [142], breast cancer [143], HCC [144], lung cancer [144,145], oral [146,147], and tongue squamous cell carcinoma [148] (Table 2). Mitochondrial TRXR (TRXR2) was also found to be upregulated in tumor tissue [149]. Moreover, tumor cells overexpress TRX [143,148,150,151,152,153] to cope with excessive ROS, and its expression is closely related to the pathological grade of the tumor [154,155,156,157,158]. Additionally, the downregulation of TXNIP, a TRX inhibitor, will result in a decrease in TXNIP-TRX complexes, protecting cells from the effects of excessive ROS and resistance to therapy. Indeed, it is common for various types of cancers to downregulate TXNIP [159,160]. The TXNIP was suggested to be used as a prognostic marker as its expression inversely correlates with the pathological grade of tumor [142,161], while its overexpression can indicate a longer survival of cancer patients [156,162,163,164]. Among the TXNDC protein family, TXNDC5 has been studied the most and was found to be overexpressed in tumor tissues [165,166,167], and its high expression correlates with poor survival [168]. Other TXNDCs reported to be altered in tumors are TXNDC9 [169,170] and TXNDC17 [171].
Table 2.
The implication of the TRX system and TXNDC protein family in cancer.
In addition to the important role of the TRX system and TXNDCs as guardians of cellular redox homeostasis, they also modulate various cellular pathways that might affect tumor development (Table 3), and some of the mechanisms are described below. TRX, mainly located in the cytosol and frequently referred to as cytoplasmic TRX (TRX1), interacts with different proteins in order to maintain cellular redox homeostasis, while in the state of oxidative stress it translocates to the nucleus modulating the activity of transcription factors. The TRX1 induces nitric oxide synthase type 3 and the S-nitrosylation of death receptor CD95, modulating apoptosis [187]. Nitric oxide synthase 2 overexpression induces the S-nitrosylation of mitochondrial TRX (TRX2) and caspase 3, altering their activity and promoting tumor growth [188]. TRX1 induces S100P, leading to an additional increase in TRX1 through a positive feedback mechanism via the upregulation of phosphorylated ERK1/2 and the downregulation of TXNIP [155]. TRX1 overexpression decreases tumor suppressor PTEN [157,189] and causes an increase in phosphorylated AKT (protein kinase B) that can consequently induce S100A4 and promote epithelial to mesenchymal transition (EMT), migration, and invasion of tumor cells [190]. Recently identified mitochondrial TRXR isoform (TRXR3) was found to reduce TRX2 and promote tumor cell survival [191]. On the contrary, the inhibition of TRXR alters the mitochondrial membrane, reduces tumor growth, and induces apoptosis [149,192,193]. Several microRNAs (miR) are known to inhibit TRXR, such as miR-125b-5p [194], miR-124 [195], and miR-17-3p [196]. Similarly to TRX, TRXR can be inactivated by nitrosylation [197], while its acetylation increases catalytic activity [198].
Table 3.
Mechanisms of action of the TRX system or TXNDC protein family in tumorigenesis.
Furthermore, metabolic and oxidative stress as well as hypoxia or hyperglycemia can promote TXNIP expression [199,200,201], which is also considered as a tumor suppressor [161]. The overexpression of TXNIP induces mitochondrial ROS generation, activates MAPK [159], and promotes apoptosis and cell cycle arrest [160,163]. Thus, the downregulation of TXNIP is frequent in tumor cells, and some of the mechanisms by which that is accomplished include targeting the TXNIP N-terminus [202], affecting the TXNIP promoter [177,203], or binding to the 3′-untranslated region (3′-UTR) of TXNIP [204,205]. The inhibition of TXNIP promotes tumor cell proliferation, EMT, and metastasis [204,205,206]. However, TXNIP can be upregulated by inhibiting histone deacetylases [207,208], bromodomain and extra-terminal domain [209], phosphatidylinositol-3-kinase (PI3K)/AKT pathway [210], the downregulation of HER1/2 [163], or focal adhesion kinase [211]. The poor survival of triple-negative breast cancer patients correlates with elevated c-MYC and decreased expression of TXNIP, which is probably due to c-MYC binding to the TXNIP promoter [177]. The secretome of monocyte-derived foam cells contains 4-HNE and was shown to increase the TXNIP expression of endothelial cells [212], which could be attributed to 4-HNE’s ability to inhibit the expression of c-MYC [213].
Among the TXNDCs, TXNDC5 can have a role in both tumor progression and tumor suppression. The inactivation or downregulation of nuclear receptor 4A1 (NR4A1) downregulates TXNDC5, isocitrate dehydrogenase 1, and the mTOR (mammalian target of rapamycin) pathway, further promoting ROS generation, inducing apoptosis, and inhibiting tumor growth [214,215,216]. The downregulation of TXNDC5 was also reported to inhibit angiogenesis [217]. On the other hand, hypoxia-induced TXNDC5 via hypoxia inducible factor 1α (HIF1α) promotes tumorigenesis [166,218].
3.3. The Glutathione System in Tumorigenesis
The glutathione system, another key regulator of cellular redox homeostasis, comprises glutamate-cysteine ligase (GCL), glutathione synthetase (GSS), reduced glutathione (GSH), oxidized glutathione (GSSG, glutathione disulfide), GPX, glutathione reductase (GR), NADPH, and glutathione S-transferase (GST) (Figure 2). GSH is ubiquitously distributed within the cell and the availability and level of GSH depend on its synthesis by GCL and GSS, the recycling of GSSG by GR and NADPH, and its detoxification activity via GST-mediated conjugation to molecules. GST belongs to phase II metabolism, and today seven classes of GST are known [232]. The detoxification role of GSH and GST is crucial to enable cells to cope with various stressors. However, alterations in GSH systems can promote tumorigenesis (Table 4). Decreased blood GSH was seen in cancer patients [233,234]. Interestingly, the GSH level was found to be increased in head and neck carcinoma [234] while the opposite was reported for colorectal cancer [235]. Moreover, cancer patients frequently have a decreased blood GSH and GPX activity [233,236,237,238,239]. Remarkably, the expression of GPX in tumor tissue depends on the GPX isoform. In that regard, GPX1, GPX3, and GPX7 expression were found to be decreased [144,240,241,242,243] in tumor tissues, while the expression of GPX2 and GPX4 was found to be upregulated [144,146,244,245,246,247]. In addition, GSTA1, GSTM1, and GSTZ1 are reported to be downregulated in tumor tissue and can correlate with a poor prognosis [63,248,249,250,251], while GSTT1, GSTO1, and GSTK1 are mostly reported to be upregulated in tumor tissue compared to the normal surrounding tissue [252,253]. The expression level of GSTP1 in tumor tissue is controversial [144,253,254,255], and its involvement in tumorigenesis could at least in part depend on its hypermethylation [256,257,258,259,260].
Table 4.
The implication of the GSH system in cancer.
Tumor cells tend to modulate the GSH system in order to survive (Table 5), and some mechanisms are described below. The upregulation of the NRF2 pathway promotes GCL, yielding elevated GSH and promoting tumorigenesis [110,272]. In the case that GSH is depleted, tumor cells overcome protein homeostasis by deubiquitinases [273]. Thus, the combined inhibition of deubiquitinases and GSH generation would be needed for malignant destruction [273]. The methylation of GPX1 promotor downregulates GPX1 [274], which can induce the activation of the AKT/GSK-3β/SNAIL pathway, promoting EMT [243]. GPX2 overexpression is also implicated in EMT [245]. However, miR-17-3p can inhibit GPX2, altering mitochondrial respiration and consequently rendering tumor cells susceptible to anticancer therapy [196]. Several other miRs also modulate the GSH system. Thus, miR-196a targets GPX3 [242], affecting GPX3-mediated cell death and promoting tumorigenesis [275]. Additionally, miR-133b downregulates GSTP1 by targeting GSTP1 3′-UTR [276]. GSTP1 is also downregulated by the methylation of the CpG island [277], while the recruitment of early B cell factor 1 to its promoter upregulates GSTP1 [255]. The overexpression of GSTP1 can induce cell cycle arrest [254], as well as GPX4 deficiency [278], and can thus be used as potential targets in anticancer therapy. GSTPs catalytic activity varies depending on the target. For example, GSTA4-4 has the highest activity towards 4-HNE and also the lowest rate with respect to 4-HNE adduction compared to others [279]. The conjugation of 4-HNE with GSH is the major route for 4-HNE elimination from cells.
Table 5.
Mechanisms of action of the GSH system in tumorigenesis.
4. Modulation of Antioxidant Defense Systems in Anticancer Therapy
The perception that antioxidants can protect cells from detrimental levels of ROS has led to several large-scale studies with somewhat disappointing results. Data suggesting the beneficial effects of antioxidant supplementation are limited [287,288], while more show no effects or even imply that antioxidants can increase cancer risk [289]. Among these, a Finnish trial on the effect of vitamin E and beta carotene on the incidence of lung cancer and other cancers in male smokers raised a lot of skepticism against the possible beneficial effects of the fat-soluble vitamins, stressing mostly their negative effects, especially an increase in the lung cancer incidence [290]. However, the authors of the trial have overseen the fact that their treatment resulted in a several-fold overload of the supplemented antioxidants in plasma, indicating their even higher potential overload in the lungs. In the case of persistent oxidative stress, as is the one occurring in heavy smokers, these antioxidants, especially beta-carotene, could be oxidized into multiple toxic, mutagenic, and likely carcinogenic products, although at the same time beta-carotene could further maintain its antioxidant capacity [291], in particular for the lung cells. Similarly, the Alpha-tocopherol, beta-carotene cancer prevention (ATBC) study in 90′ linked the supplementation of beta-carotene with the increased risk of lung cancer among smokers [292], while the more recent findings attenuated so negative results showing that supplementary alfa-tocopherol and beta-carotene have no late effects on cancer incidence. Thus, after two decades, the authors of the ATBC trial revealed the preventive effect of moderate-dose alpha-tocopherol on prostate cancer that continued for several years after the trial, reducing prostate cancer mortality [293]. However, the issue of the use of antioxidants as adjuvant therapy is still under debate favoring the laconic “no use” approach [294,295], while the increased recurrence and mortality of breast cancer in patients who concurrently took dietary supplements with radio- and chemotherapy is alarming [296].
While considering the possible benefits or risks of using antioxidants in oncology, one should recall the pathophysiology of oxidative stress, which is aware of beneficial roles played by endogenous antioxidants, especially in the case of acute stress response (such as those induced by irradiation, toxic compounds, or ischemia/reperfusion), for which optimal interplay between water-resistant and water-soluble antioxidants is essential. If that is not the case, many antioxidants (in particular, those water-resistant—i.e., lipid-soluble—present in biomembranes) could become free radicals, even more harmful than the initial ROS that started oxidative stress. That is even more relevant for chronic oxidative stress, as occurs in cancer and inflammation, while the long-lasting or excessive use of exogenous antioxidants even bears a risk of uncontrolled interference with endogenous antioxidants, which is the main reason why antioxidants cannot be considered as panacea. Similarly, even in the case of mild and acute oxidative stress (like in exercise) the use “preventive” exogeneous antioxidants can block desirable hormetic feedback effects of exercise, that can cause a rise in the endogenous antioxidant capacities, mostly affecting NRF2, which may eventually increase the resistance of oxidative-stress associated disorders.
The NRF2 pathway is the main cellular defense mechanism activated upon exposure to oxidative stress, electrophilic stress, and xenobiotics. It regulates the expression of a great variety of cytoprotective genes, enabling cells to withstand unfavorable conditions and restore the homeostatic state. In an effort to reduce cancer incidence, past research has been focused on the activation of what was considered as a cancer-protective mechanism, the NRF2 pathway. Thus, the extensive search for activators of the NRF2 pathways begun. Many candidates emerged, some reaching clinical trials. Sulforaphane is one such candidate abundantly present in broccoli sprouts. Clinical trials with broccoli sprout extracts revealed the attenuation of cancer risk in individuals exposed to aflatoxins and air-borne toxins [297] and insufficient, although safe, anticancer activity in prostate cancer patients [298], while others are still ongoing (extensively reviewed in [299]). Curcumin is another NRF2 activator with pleiotropic activity [300]. To date, clinical trials investigating the effectiveness of curcumin treatment in diverse cancers revealed the omission of the expected effect. Yet, curcumin was found to be well-tolerable and safe [301]. Other NRF2 activators enrolled in clinical trials include resveratrol, bardoxolone-methyl (CDDO-Me), oltipraz, RTA-408 (omaveloxolone), etc. (reviewed in [302]).
While the translation of in vitro and in vivo observed NRF2 activator-induced benefits to the clinic are still scarce, caution concerning their usage arises. The negative effect of NRF2 activation was observed in diabetic patients. Standard diabetic drugs, saxagliptin and sitagliptin, were shown to increase the risk of a metastatic spread in patients who already have cancer, although they did not enhance cancer incidence. The underlying mechanism included prolonged NRF2 activation [303]. Therefore, the use of NRF2 activators should be cautiously evaluated, particularly in cancer patients, considering the hyperactivation of the NRF2 pathway observed in the vast majority of cancers.
The constitutive activation of NRF2, observed in many cancers, causes research to rethink the new possibilities for treating cancer that can amend acquired resistance to conventional therapy. Inhibitors of the NRF2 activation, and its target products such as TRX and GSH, have become a focus of this research. All-trans retinoic acid (ATRA), clobetasol propionate (CP), and apigenin are some of the examples of the NRF2 inhibitors under clinical investigation, while others show promise in cellular and animal models, including ARE expression modulator 1 (AEM1), ML385, 1-(2-cyclohexylethoxy)aniline (IM3829), malabaricone-A (MAL-A), etc. [39,47]. ATRA forms a complex with retinoic acid receptors (RARs) that bind with NRF2, thus interfering with the binding of NRF2 to ARE sequences and blocking the activation of the NRF2 pathway [304]. Although already in use for treating acute promyelocytic leukemia patients, its applicability in the treatment of solid tumors is still ambiguous and heavily ongoing (reviewed in [305]). CP is another NRF2 inhibitor currently evaluated in phase 2 clinical trials (NCT02368886) in patients with refractory metastatic colorectal cancer. CP prevents nuclear accumulation and promotes the β-TRCP-dependent degradation of NRF2 in a glucocorticoid receptor- and a GSK-3-dependent manner [306]. While apigenin was shown to inhibit NRF2 activation [307], further research revealed its multiple modes of action [308]. Apigenin did reach the clinical trial (NCT00609310) investigating the prevention of neoplasia recurrence, yet the study has been suspended. Auranofin, an FDA-approved drug for the treatment of rheumatoid arthritis, and buthionine sulfoximine (BSO), an inhibitor of GSH synthesis, have been in consideration as anticancer agents as well. While BSO failed in the clinic, auranofin is still under investigation in several clinical trials [309]. Its mode of action includes the inhibition of the activity of TRXR and thus the disruption of the TRX system [310].
The recent opinion suggests combinational therapy to be more adequate in cancer treatment because anticancer agents usually affect different pathways, not offering cancer cells to adapt so quickly as in monotherapy and is thus reducing the occurrence of chemoresistance. In addition, the mutual effect lowers the effective dose, which in turn attenuates the unwanted side effects of some drugs [311]. PI3K/AKT inhibitors, such as MK2206 (Merck), have shown some promise in clinical trials. Yet, in vitro and in vivo studies have revealed the non-responsiveness of such monotherapy in NSCLC, suggesting combinational therapy as an advantageous strategy. Thus, a synthetic lethality induced by MK2206 and TXNRD1 inhibitor auranofin, found to be dependent on the genetic status of KEAP1, shows promise in KEAP1 wild-type over mutant [312].
A more personalized approach, such as the analysis of patients’ mutational status and putting them in context with the known mechanisms of the anticancer treatments, could improve patients’ outcomes. For example, combining clinical data and in vivo and in vitro approaches revealed the importance of KEAP1 and NFE2L2 mutations in lung adenocarcinoma, correlating them with advanced stages and worse survival. Associating these mutations with KRAS mutations revealed the high dependence on glutaminolysis of KRAS/KEAP1 or KRAS/NFE2L2 mutants, a potentially exploitable feature in future therapy [313]. In addition, KRAS/KEAP1 mutants were found to arise from a bronchiolar cell of origin and keep the pentose phosphate pathway active, another possible exploitable feature in therapy [314]. Moreover, in lung adenocarcinoma tumors co-occurring KEAP1 mutations and STK11/LKB1 loss lead to metabolic reprogramming (glutamine metabolism), activating the pentose phosphate pathway and the tricarboxylic acid (TCA) cycle to maintain redox balance, suggesting a glutaminase inhibitor as a possible treatment strategy [56]. Xu et al. investigated the relationship between NFE2L2/KEAP1 mutations, tumor mutational burden (TMB), and programmed death ligand 1 (PD-L1) expression. They found that NFE2L2/KEAP1 mutations are present in various cancers, with the highest incidence found in lung squamous cell carcinoma. These mutations were linked with higher TMB and PD-L1 expression. Since the NFE2L2/KEAP1 mutations in cancer are often associated with poorer overall survival, a survival analysis of NSCLC patients receiving immunotherapy revealed improved clinical outcomes in comparison to other treatments, suggesting its possible beneficial use for patients with mutations in NFE2L2/KEAP1 [315]. In addition, NFE2L2/KEAP1 mutations cause the constitutive activation of the NRF2 pathway and enhanced ARE activity, a feature suggested as being exploitable for a cancer suicide gene therapy. Leinonen et al. used a lentiviral vector expressing herpes simplex virus thymidine kinase (HSV-TK/GCV) under the regulation of ARE. They evaluated this approach in human lung adenocarcinoma cells. They showed this approach to be effective in both in vitro and in vivo and suggested it as a promising treatment in conjunction with traditional therapies [316,317]. Table 6 contains a summary of antioxidant defense system modulators and strategies in cancer incidence and therapy.
Table 6.
Summary of the described antioxidant defense mechanisms in cancer incidence and anticancer therapy.
5. Conclusions
Cancer biology is a very complex process that includes the multifaceted interplay between antioxidant systems and ROS in determining cancer development, progression, metastasis, and regression. Such complexity is sometimes overseen while evaluating the effects of potential anticancer antioxidants, as in the ATBC trial in the 1990s. In the modern era, realizing that a more personalized and integrative biomedical approach could give more benefits for the prevention and therapy of cancer, we should focus on altered oxidative homeostasis in cancer cells.
Therefore, the relevance of the NRF2 pathway and TRX and GSH systems in carcinogenesis and in anticancer therapies has been extensively investigated. Targeting only one system can be beneficial, while the combined modulation of multiple antioxidant systems can give better anticancer results. Besides various synthetic agents, miRs should also be considered in anticancer therapies, as they have been shown to have an important role in the modulation of the NRF2 pathway and TRX and GSH antioxidant system efficiencies.
Author Contributions
Conceptualization, M.J. and L.M.; investigation, M.J., L.M., and S.B.S.; writing—original draft preparation, M.J., L.M., S.B.S., and N.Z.; writing—review and editing, M.J., L.M., and N.Z.; visualization, M.J. and L.M.; supervision, N.Z. All the authors have read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA. Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Cipak Gasparovic, A.; Cindric, M.; Mouthuy, P.A.; Zarkovic, N. Short Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef] [PubMed]
- Dodson, M.; de la Vega, M.R.; Cholanians, A.B.; Schmidlin, C.J.; Chapman, E.; Zhang, D.D. Modulating NRF2 in Disease: Timing Is Everything. Annu. Rev. Pharmacol. Toxicol. 2019, 59, 555–575. [Google Scholar] [CrossRef] [PubMed]
- Zińczuk, J.; Maciejczyk, M.; Zaręba, K.; Pryczynicz, A.; Dymicka-Piekarska, V.; Kamińska, J.; Koper-Lenkiewicz, O.; Matowicka-Karna, J.; Kędra, B.; Zalewska, A.; et al. Pro-oxidant enzymes, redox balance and oxidative damage to proteins, lipids and DNA in colorectal cancer tissue. Is oxidative stress dependent on tumour budding and inflammatory infiltration? Cancers 2020, 12, 1636. [Google Scholar] [CrossRef]
- Harris, I.S.; Treloar, A.E.; Inoue, S.; Sasaki, M.; Gorrini, C.; Lee, K.C.; Yung, K.Y.; Brenner, D.; Knobbe-Thomsen, C.B.; Cox, M.A.; et al. Glutathione and thioredoxin antioxidant pathways synergize to drive cancer initiation and progression. Cancer Cell 2015, 27, 211–222. [Google Scholar] [CrossRef]
- Jaganjac, M.; Cipak, A.; Schaur, R.J.; Zarkovic, N. Pathophysiology of neutrophil-mediated extracellular redox reactions. Front. Biosci. Landmark 2016, 21, 839–855. [Google Scholar] [CrossRef]
- Jaganjac, M.; Milkovic, L.; Gegotek, A.; Cindric, M.; Zarkovic, K.; Skrzydlewska, E.; Zarkovic, N. The relevance of pathophysiological alterations in redox signaling of 4-hydroxynonenal for pharmacological therapies of major stress-associated diseases. Free Radic. Biol. Med. 2019. [Google Scholar] [CrossRef]
- Pi, J.; Bai, Y.; Zhang, Q.; Wong, V.; Floering, L.M.; Daniel, K.; Reece, J.M.; Deeney, J.T.; Andersen, M.E.; Corkey, B.E.; et al. Reactive oxygen species as a signal in glucose-stimulated insulin secretion. Diabetes 2007, 56, 1783–1791. [Google Scholar] [CrossRef]
- Li, N.; Brun, T.; Cnop, M.; Cunha, D.A.; Eizirik, D.L.; Maechler, P. Transient oxidative stress damages mitochondrial machinery inducing persistent β-cell dysfunction. J. Biol. Chem. 2009, 284, 23602–23612. [Google Scholar] [CrossRef]
- Jaganjac, M.; Tirosh, O.; Cohen, G.; Sasson, S.; Zarkovic, N. Reactive aldehydes-second messengers of free radicals in diabetes mellitus. Free Radic. Res. 2013, 47, 39–48. [Google Scholar] [CrossRef]
- Zivkovic, M.; Poljak-Blazi, M.; Egger, G.; Sunjic, S.B.; Schaur, R.J.; Zarkovic, N. Oxidative burst and anticancer activities of rat neutrophils. BioFactors 2005, 24, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, M.; Poljak-Blazi, M.; Zarkovic, K.; Mihaljevic, D.; Schaur, R.J.; Zarkovic, N. Oxidative burst of neutrophils against melanoma B16-F10. Cancer Lett. 2007, 246, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Poljak-Blazi, M.; Zarkovic, K.; Schaur, R.J.; Zarkovic, N. The involvement of granulocytes in spontaneous regression of Walker 256 carcinoma. Cancer Lett. 2008, 260, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Poljak-Blazi, M.; Schaur, R.J.; Zarkovic, K.; Borovic, S.; Cipak, A.; Cindric, M.; Uchida, K.; Waeg, G.; Zarkovic, N. Elevated neutrophil elastase and acrolein-protein adducts are associated with W256 regression. Clin. Exp. Immunol. 2012, 170, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Poljak-Blazi, M.; Kirac, I.; Borovic, S.; Joerg Schaur, R.; Zarkovic, N. Granulocytes as effective anticancer agent in experimental solid tumor models. Immunobiology 2010, 215, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Matijevic Glavan, T.; Zarkovic, N. The Role of Acrolein and NADPH Oxidase in the Granulocyte-Mediated Growth-Inhibition of Tumor Cells. Cells 2019, 8, 292. [Google Scholar] [CrossRef]
- Al-Thani, A.M.; Voss, S.C.; Al-Menhali, A.S.; Barcaru, A.; Horvatovich, P.; Al Jaber, H.; Nikolovski, Z.; Latiff, A.; Georgakopoulos, C.; Merenkov, Z.; et al. Whole blood storage in CPDA1 blood bags alters erythrocyte membrane proteome. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef]
- Ludtmann, M.H.R.; Angelova, P.R.; Horrocks, M.H.; Choi, M.L.; Rodrigues, M.; Baev, A.Y.; Berezhnov, A.V.; Yao, Z.; Little, D.; Banushi, B.; et al. α-synuclein oligomers interact with ATP synthase and open the permeability transition pore in Parkinson’s disease. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef]
- Poljak-Blazi, M.; Jaganjac, M.; Sabol, I.; Mihaljevic, B.; Matovina, M.; Grce, M. Effect of ferric ions on reactive oxygen species formation, cervical cancer cell lines growth and E6/E7 oncogene expression. Toxicol. Vitr. 2011, 25, 160–166. [Google Scholar] [CrossRef]
- Kukulj, S.; Jaganjac, M.; Boranic, M.; Krizanac, S.; Santic, Z.; Poljak-Blazi, M. Altered iron metabolism, inflammation, transferrin receptors, and ferritin expression in non-small-cell lung cancer. Med. Oncol. 2010, 27, 268–277. [Google Scholar] [CrossRef]
- Toyokuni, S.; Ito, F.; Yamashita, K.; Okazaki, Y.; Akatsuka, S. Iron and thiol redox signaling in cancer: An exquisite balance to escape ferroptosis. Free Radic. Biol. Med. 2017, 108, 610–626. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Sunjic, S.B.; Zarkovic, N. Utilizing iron for targeted lipid peroxidation as anticancer option of integrative biomedicine: A short review of nanosystems containing iron. Antioxidants 2020, 9, 191. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.S.; Evans, M.D.; Dizdaroglu, M.; Lunec, J. Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB J. 2003, 17, 1195–1214. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, X.; Li, Z.; Ye, W.; Ding, H.; Li, P.; Aung, L.H.H. Role of RNA oxidation in neurodegenerative diseases. Int. J. Mol. Sci. 2020, 21, 5022. [Google Scholar] [CrossRef] [PubMed]
- Schauenstein, E. Autoxidation of polyunsaturated esters in water: Chemical structure and biological activity of the products. J. Lipid Res. 1967, 8, 417–428. [Google Scholar] [PubMed]
- Schaur, R.J. Basic aspects of the biochemical reactivity of 4-hydroxynonenal. Mol. Asp. Med. 2003, 24, 149–159. [Google Scholar] [CrossRef]
- Zarkovic, N.; Cipak, A.; Jaganjac, M.; Borovic, S.; Zarkovic, K. Pathophysiological relevance of aldehydic protein modifications. J. Proteom. 2013, 92, 239–247. [Google Scholar] [CrossRef]
- Gentile, F.; Arcaro, A.; Pizzimenti, S.; Daga, M.; Cetrangolo, G.P.; Dianzani, C.; Lepore, A.; Graf, M.; Ames, P.R.J.; Barrera, G. DNA damage by lipid peroxidation products: Implications in cancer, inflammation and autoimmunity. AIMS Genet. 2017, 4, 103–137. [Google Scholar] [CrossRef]
- Jaganjac, M.; Čačev, T.; Čipak, A.; Kapitanović, S.; Trošelj, K.G.; Žarković, N. Even stressed cells are individuals: Second messengers of free radicals in pathophysiology of cancer. Croat. Med. J. 2012, 53, 304–309. [Google Scholar] [CrossRef]
- Al-Menhali, A.S.; Banu, S.; Angelova, P.R.; Barcaru, A.; Horvatovich, P.; Abramov, A.Y.; Jaganjac, M. Lipid peroxidation is involved in calcium dependent upregulation of mitochondrial metabolism in skeletal muscle. Biochim. Biophys. Acta Gen. Subj. 2020, 1864. [Google Scholar] [CrossRef]
- Elrayess, M.A.; Almuraikhy, S.; Kafienah, W.; Al-Menhali, A.; Al-Khelaifi, F.; Bashah, M.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Alsayrafi, M.; et al. 4-hydroxynonenal causes impairment of human subcutaneous adipogenesis and induction of adipocyte insulin resistance. Free Radic. Biol. Med. 2017, 104, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Almuraikhy, S.; Al-Khelaifi, F.; Al-Jaber, M.; Bashah, M.; Mazloum, N.A.; Zarkovic, K.; Zarkovic, N.; Waeg, G.; Kafienah, W.; et al. Combined metformin and insulin treatment reverses metabolically impaired omental adipogenesis and accumulation of 4-hydroxynonenal in obese diabetic patients. Redox Biol. 2017, 12, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Mrakovcic, L.; Wildburger, R.; Jaganjac, M.; Cindric, M.; Cipak, A.; Sunjic, S.B.; Waeg, G.; Milankovic, A.M.; Zarkovic, N. Lipid peroxidation product 4-hydroxynonenal as factor of oxidative homeostasis supporting bone regeneration with bioactive glasses. Acta Biochim. Pol. 2010, 57, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Cipak, A.; Mrakovcic, L.; Ciz, M.; Lojek, A.; Mihaylova, B.; Goshev, I.; Jaganjac, M.; Cindric, M.; Sitic, S.; Margaritoni, M.; et al. Growth suppression of human breast carcinoma stem cells by lipid peroxidation product 4-hydroxy-2-nonenal and hydroxyl radical-modified collagen. Acta Biochim. Pol. 2010, 57, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Semlitsch, T.; Tillian, H.M.; Zarkovic, N.; Borovic, S.; Purtscher, M.; Hohenwarter, O.; Schaur, R.J. Differential influence of the lipid peroxidation product 4-hydroxynonenal on the growth of human lymphatic leukaemia cells and human periopherial blood lymphocytes. Anticancer Res. 2002, 22, 1689–1697. [Google Scholar] [PubMed]
- Kreuzer, T.; Zarković, N.; Grube, R.; Schaur, R.J. Inhibition of HeLa cell proliferation by 4-hydroxynonenal is associated with enhanced expression of the c-fos oncogene. Cancer Biother. Radiopharm. 1997, 12, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, N.; Chisci, E.; Giovannoni, R. The Role of Hydrogen Peroxide in Redox-Dependent Signaling: Homeostatic and Pathological Responses in Mammalian Cells. Cells 2018, 7, 156. [Google Scholar] [CrossRef]
- Chan, J.Y.H.; Chan, S.H.H. Activation of endogenous antioxidants as a common therapeutic strategy against cancer, neurodegeneration and cardiovascular diseases: A lesson learnt from DJ-1. Pharmacol. Ther. 2015, 156, 69–74. [Google Scholar] [CrossRef]
- Milkovic, L.; Zarkovic, N.; Saso, L. Controversy about pharmacological modulation of Nrf2 for cancer therapy. Redox Biol. 2017, 12, 727–732. [Google Scholar]
- Chowdhry, S.; Zhang, Y.; McMahon, M.; Sutherland, C.; Cuadrado, A.; Hayes, J.D. Nrf2 is controlled by two distinct β-TrCP recognition motifs in its Neh6 domain, one of which can be modulated by GSK-3 activity. Oncogene 2013, 32, 3765–3781. [Google Scholar] [CrossRef]
- Wu, T.; Zhao, F.; Gao, B.; Tan, C.; Yagishita, N.; Nakajima, T.; Wong, P.K.; Chapman, E.; Fang, D.; Zhang, D.D. Hrd1 suppresses Nrf2-mediated cellular protection during liver cirrhosis. Genes Dev. 2014, 28, 708–722. [Google Scholar] [CrossRef] [PubMed]
- Dhakshinamoorthy, S.; Jain, A.K.; Bloom, D.A.; Jaiswal, A.K. Bach1 competes with Nrf2 leading to negative regulation of the antioxidant response element (ARE)-mediated NAD(P)H:quinone oxidoreductase 1 gene expression and induction in response to antioxidants. J. Biol. Chem. 2005, 280, 16891–16900. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jia, Z.; Zhu, H. Regulation of Nrf2 Signaling. React. Oxyg. Species 2019, 8, 312–322. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [PubMed]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Harder, B.; Jiang, T.; Wu, T.; Tao, S.; De La Vega, M.R.; Tian, W.; Chapman, E.; Zhang, D.D. Molecular mechanisms of Nrf2 regulation and how these influence chemical modulation for disease intervention. Biochem. Soc. Trans. 2015, 43, 680–686. [Google Scholar] [CrossRef]
- Cloer, E.W.; Goldfarb, D.; Schrank, T.P.; Weissman, B.E.; Major, M.B. Nrf2 activation in cancer: From DNA to protein. Cancer Res. 2019, 79, 889–898. [Google Scholar] [CrossRef]
- Xu, C.; Huang, M.T.; Shen, G.; Yuan, X.; Lin, W.; Khor, T.O.; Conney, A.H.; Kong, A.N.T. Inhibition of 7,12-dimethylbenz(a)anthracene-induced skin tumorigenesis in C57BL/6 mice by sulforaphane is mediated by nuclear factor E2-related factor 2. Cancer Res. 2006, 66, 8293–8296. [Google Scholar] [CrossRef]
- Becks, L.; Prince, M.; Burson, H.; Christophe, C.; Broadway, M.; Itoh, K.; Yamamoto, M.; Mathis, M.; Orchard, E.; Shi, R.; et al. Aggressive mammary carcinoma progression in Nrf2 knockout mice treated with 7,12-dimethylbenz[a]anthracene. BMC Cancer 2010, 10, 540. [Google Scholar] [CrossRef]
- Ramos-Gomez, M.; Kwak, M.K.; Dolan, P.M.; Itoh, K.; Yamamoto, M.; Talalay, P.; Kensler, T.W. Sensitivity to carcinogenesis is increased and chemoprotective efficacy of enzyme inducers is lost in nrf2 transcription factor-deficient mice. Proc. Natl. Acad. Sci. USA 2001, 98, 3410–3415. [Google Scholar] [CrossRef]
- Suzuki, T.; Shibata, T.; Takaya, K.; Shiraishi, K.; Kohno, T.; Kunitoh, H.; Tsuta, K.; Furuta, K.; Goto, K.; Hosoda, F.; et al. Regulatory Nexus of Synthesis and Degradation Deciphers Cellular Nrf2 Expression Levels. Mol. Cell. Biol. 2013, 33, 2402–2412. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.C.; Ambrosone, C.B.; Ahn, J.; Choi, J.Y.; McCullough, M.L.; Stevens, V.L.; Rodriguez, C.; Thun, M.J.; Calle, E.E. Genetic variability in iron-related oxidative stress pathways (Nrf2, NQ01, NOS3, and HO-1), iron intake, and risk of postmenopausal breast cancer. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1784–1794. [Google Scholar] [CrossRef]
- Yang, X.; Wang, D.; Ma, Y.; Xu, X.; Zhu, Z.; Wang, X.; Deng, H.; Li, C.; Chen, M.; Tong, J.; et al. Continuous activation of Nrf2 and its target antioxidant enzymes leads to arsenite-induced malignant transformation of human bronchial epithelial cells. Toxicol. Appl. Pharmacol. 2015, 289, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Levings, D.C.; Wang, X.; Kohlhase, D.; Bell, D.A.; Slattery, M. A distinct class of antioxidant response elements is consistently activated in tumors with NRF2 mutations. Redox Biol. 2018, 19, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Hellyer, J.A.; Stehr, H.; Das, M.; Padda, S.K.; Ramchandran, K.; Neal, J.W.; Diehn, M.; Wakelee, H.A. Impact of KEAP1/NFE2L2/CUL3 mutations on duration of response to EGFR tyrosine kinase inhibitors in EGFR mutated non-small cell lung cancer. Lung Cancer 2019, 134, 42–45. [Google Scholar] [CrossRef]
- Galan-Cobo, A.; Sitthideatphaiboon, P.; Qu, X.; Poteete, A.; Pisegna, M.A.; Tong, P.; Chen, P.H.; Boroughs, L.K.; Rodriguez, M.L.M.; Zhang, W.; et al. LKB1 and KEAP1/NRF2 pathways cooperatively promote metabolic reprogramming with enhanced glutamine dependence inKRAS-mutant lung adenocarcinoma. Cancer Res. 2019, 79, 3251–3267. [Google Scholar] [CrossRef]
- Qian, Z.; Zhou, T.; Gurguis, C.I.; Xu, X.; Wen, Q.; Lv, J.; Fang, F.; Hecker, L.; Cress, A.E.; Natarajan, V.; et al. Nuclear factor, erythroid 2-like 2-associated molecular signature predicts lung cancer survival. Sci. Rep. 2015, 5, 16889. [Google Scholar] [CrossRef]
- Sasaki, H.; Shitara, M.; Yokota, K.; Hikosaka, Y.; Moriyama, S.; Yano, M.; Fujii, Y. MRP3 gene expression correlates with NRF2 mutations in lung squamous cell carcinomas. Mol. Med. Rep. 2012, 6, 705–708. [Google Scholar] [CrossRef]
- Hast, B.E.; Goldfarb, D.; Mulvaney, K.M.; Hast, M.A.; Siesser, P.F.; Yan, F.; Hayes, D.N.; Major, M.B. Proteomic analysis of ubiquitin ligase KEAP1 reveals associated proteins that inhibit NRF2 ubiquitination. Cancer Res. 2013, 73, 2199–2210. [Google Scholar] [CrossRef]
- Wang, Q.; Ma, J.; Lu, Y.; Zhang, S.; Huang, J.; Chen, J.; Bei, J.X.; Yang, K.; Wu, G.; Huang, K.; et al. CDK20 interacts with KEAP1 to activate NRF2 and promotes radiochemoresistance in lung cancer cells. Oncogene 2017, 36, 5321–5330. [Google Scholar] [CrossRef]
- Umemura, A.; He, F.; Taniguchi, K.; Nakagawa, H.; Yamachika, S.; Font-Burgada, J.; Zhong, Z.; Subramaniam, S.; Raghunandan, S.; Duran, A.; et al. p62, Upregulated during Preneoplasia, Induces Hepatocellular Carcinogenesis by Maintaining Survival of Stressed HCC-Initiating Cells. Cancer Cell 2016, 29, 935–948. [Google Scholar] [CrossRef]
- Saito, T.; Ichimura, Y.; Taguchi, K.; Suzuki, T.; Mizushima, T.; Takagi, K.; Hirose, Y.; Nagahashi, M.; Iso, T.; Fukutomi, T.; et al. P62/Sqstm1 promotes malignancy of HCV-positive hepatocellular carcinoma through Nrf2-dependent metabolic reprogramming. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef]
- Yang, F.; Li, J.; Deng, H.; Wang, Y.; Lei, C.; Wang, Q.; Xiang, J.; Liang, L.; Xia, J.; Pan, X.; et al. GSTZ 1-1 Deficiency Activates NRF 2/IGF 1R Axis in HCC via Accumulation of Oncometabolite Succinylacetone. EMBO J. 2019, 38, e101964. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, F.P.; Costantini, M.; Copetti, M.; la Torre, A.; Sparaneo, A.; Fontana, A.; Poeta, L.; Gallucci, M.; Sentinelli, S.; Graziano, P.; et al. Keap1/Nrf2 pathway in kidney cancer: Frequent methylation of KEAP1 gene promoter in clear renal cell carcinoma. Oncotarget 2017, 8, 11187–11198. [Google Scholar] [CrossRef] [PubMed]
- Kowalik, M.A.; Guzzo, G.; Morandi, A.; Perra, A.; Menegon, S.; Masgras, I.; Trevisan, E.; Angioni, M.M.; Fornari, F.; Quagliata, L.; et al. Metabolic reprogramming identifies the most aggressive lesions at early phases of hepatic carcinogenesis. Oncotarget 2016, 7, 32375–32393. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tao, S.; Liao, L.; Li, Y.; Li, H.; Li, Z.; Lin, L.; Wan, X.; Yang, X.; Chen, L. TRIM25 promotes the cell survival and growth of hepatocellular carcinoma through targeting Keap1-Nrf2 pathway. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Lignitto, L.; LeBoeuf, S.E.; Homer, H.; Jiang, S.; Askenazi, M.; Karakousi, T.R.; Pass, H.I.; Bhutkar, A.J.; Tsirigos, A.; Ueberheide, B.; et al. Nrf2 Activation Promotes Lung Cancer Metastasis by Inhibiting the Degradation of Bach1. Cell 2019, 178, 316–329.e18. [Google Scholar] [CrossRef]
- Wiel, C.; Le Gal, K.; Ibrahim, M.X.; Jahangir, C.A.; Kashif, M.; Yao, H.; Ziegler, D.V.; Xu, X.; Ghosh, T.; Mondal, T.; et al. BACH1 Stabilization by Antioxidants Stimulates Lung Cancer Metastasis. Cell 2019, 178, 330–345.e22. [Google Scholar] [CrossRef]
- Ge, M.K.; Zhang, N.; Xia, L.; Zhang, C.; Dong, S.S.; Li, Z.M.; Ji, Y.; Zheng, M.H.; Sun, J.; Chen, G.Q.; et al. FBXO22 degrades nuclear PTEN to promote tumorigenesis. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, J.; Ning, D.; Liu, Q.; Wang, C.; Zhang, Z.; Chu, L.; Yu, C.; Liang, H.F.; Zhang, B.; et al. FBXO22 promotes the development of hepatocellular carcinoma by regulating the ubiquitination and degradation of p21. J. Exp. Clin. Cancer Res. 2019, 38, 101. [Google Scholar] [CrossRef]
- Sun, R.; Xie, H.Y.; Qian, J.X.; Huang, Y.N.; Yang, F.; Zhang, F.L.; Shao, Z.M.; Li, D.Q. FBXO22 possesses both protumorigenic and antimetastatic roles in breast cancer progression. Cancer Res. 2018, 78, 5274–5286. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, S.A.; Zaitseva, L.; Murray, M.Y.; Shah, N.M.; Bowles, K.M.; MacEwan, D.J. The high Nrf2 expression in human acute myeloid leukemia is driven by NF-κB and underlies its chemo-resistance. Blood 2012, 120, 5188–5198. [Google Scholar] [CrossRef] [PubMed]
- Rushworth, S.A.; Bowles, K.M.; MacEwan, D.J. High basal nuclear levels of Nrf2 in acute myeloid leukemia reduces sensitivity to proteasome inhibitors. Cancer Res. 2011, 71, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Ren, Y.; Yan, X.; Luo, Y.; Zhang, H.; Kesarwani, M.; Bu, J.; Zhan, D.; Zhou, Y.; Tang, Y.; et al. The high NRF2 expression confers chemotherapy resistance partly through up-regulated DUSP1 in myelodysplastic syndromes. Haematologica 2019, 104, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.P.; Hayashi, T.; Cottam, H.B.; Jin, G.; Yao, S.; Wu, C.C.N.; Rosenbach, M.D.; Corr, M.; Schwab, R.B.; Carson, D.A. Nrf2 responses and the therapeutic selectivity of electrophilic compounds in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2010, 107, 7479–7484. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Ghia, E.M.; Antonucci, L.; Sharma, N.; Rassenti, L.Z.; Xu, J.; Sun, B.; Kipps, T.J.; Karin, M. NF-κB-p62-NRF2 survival signaling is associated with high ROR1 expression in chronic lymphocytic leukemia. Cell Death Differ. 2020, 27, 2206–2216. [Google Scholar] [CrossRef]
- Yi, X.; Zhao, Y.; Xue, L.; Zhang, J.; Qiao, Y.; Jin, Q.; Li, H. Expression of keap1 and nrf2 in diffuse large b-cell lymphoma and its clinical significance. Exp. Ther. Med. 2018, 16, 573–578. [Google Scholar] [CrossRef]
- Kari, E.; Teppo, H.R.; Haapasaari, K.M.; Kuusisto, M.E.L.; Lemma, A.; Karihtala, P.; Pirinen, R.; Soini, Y.; Jantunen, E.; Turpeenniemi-Hujanen, T.; et al. Nuclear factor erythroid 2-related factors 1 and 2 are able to define the worst prognosis group among high-risk diffuse large B cell lymphomas treated with R-CHOEP. J. Clin. Pathol. 2019, 72, 316–321. [Google Scholar] [CrossRef]
- Frank, R.; Scheffler, M.; Merkelbach-Bruse, S.; Ihle, M.A.; Kron, A.; Rauer, M.; Ueckeroth, F.; Konig, K.; Michels, S.; Fischer, R.; et al. Clinical and pathological characteristics of KEAP1- and NFE2L2-mutated Non–Small Cell Lung Carcinoma (NSCLC). Clin. Cancer Res. 2018, 24, 3087–3096. [Google Scholar] [CrossRef]
- Goeman, F.; De Nicola, F.; Scalera, S.; Sperati, F.; Gallo, E.; Ciuffreda, L.; Pallocca, M.; Pizzuti, L.; Krasniqi, E.; Barchiesi, G.; et al. Mutations in the KEAP1-NFE2L2 Pathway Define a Molecular Subset of Rapidly Progressing Lung Adenocarcinoma. J. Thorac. Oncol. 2019, 14, 1924–1934. [Google Scholar] [CrossRef]
- Aljohani, H.M.; Aittaleb, M.; Furgason, J.M.; Amaya, P.; Deeb, A.; Chalmers, J.J.; Bahassi, E.M. Genetic mutations associated with lung cancer metastasis to the brain. Mutagenesis 2018, 33, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, H.L.; Lee, K.B.; Park, J.H.; Chung, W.Y.; Lee, K.S.; Sheen, S.S.; Park, K.J.; Hwang, S.C. Nuclear factor E2-related factor 2 dependent overexpression of sulfiredoxin and peroxiredoxin III in human lung cancer. Korean J. Intern. Med. 2011, 26, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Hikosaka, Y.; Okuda, K.; Kawano, O.; Moriyama, S.; Yano, M.; Fujii, Y. NFE2L2 gene mutation in male japanese squamous cell carcinoma of the lung. J. Thorac. Oncol. 2010, 5, 786–789. [Google Scholar] [CrossRef] [PubMed]
- Arbour, K.C.; Jordan, E.; Kim, H.R.; Dienstag, J.; Yu, H.A.; Sanchez-Vega, F.; Lito, P.; Berger, M.; Solit, D.B.; Hellmann, M.; et al. Effects of co-occurring genomic alterations on outcomes in patients with KRAS-mutant non-small cell lung cancer. Clin. Cancer Res. 2018, 24, 334–340. [Google Scholar] [CrossRef]
- Jeong, Y.; Hoang, N.T.; Lovejoy, A.; Stehr, H.; Newman, A.M.; Gentles, A.J.; Kong, W.; Truong, D.; Martin, S.; Chaudhuri, A.; et al. Role of KEAP1/NRF2 and TP53 mutations in lung squamous cell carcinoma development and radiation resistance. Cancer Discov. 2017, 7, 86–101. [Google Scholar] [CrossRef]
- Yu, S.; Cheng, C.; Wang, J.; Wang, J.; Qu, Z.; Ren, H.; Li, Y.; Ning, Q.; Chen, M.; Hu, T. Loss of Beclin1 Expression and Nrf2 Overexpression are Associated with Poor Survival of Patients with Non-Small Cell Lung Cancer. Anticancer Agents Med. Chem. 2018, 18, 1680–1687. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Koh, Y.; Ando, M.; Ito, N.; Takeo, S.; Adachi, H.; Tagawa, T.; Kakegawa, S.; Yamashita, M.; Kataoka, K.; et al. Prospective analysis of oncogenic driver mutations and environmental factors: Japan molecular epidemiology for lung cancer study. J. Clin. Oncol. 2016, 34, 2247–2257. [Google Scholar] [CrossRef]
- Choi, M.; Kadara, H.; Zhang, J.; Parra, E.R.; Rodriguez-Canales, J.; Gaffney, S.G.; Zhao, Z.; Behrens, C.; Fujimoto, J.; Chow, C.; et al. Mutation profiles in early-stage lung squamous cell carcinoma with clinical follow-up and correlation with markers of immune function. Ann. Oncol. 2017, 28, 83–89. [Google Scholar] [CrossRef]
- Cescon, D.W.; She, D.; Sakashita, S.; Zhu, C.Q.; Pintilie, M.; Shepherd, F.A.; Tsao, M.S. NRF2 pathway activation and adjuvant chemotherapy benefit in lung squamous Cell Carcinoma. Clin. Cancer Res. 2015, 21, 2499–2505. [Google Scholar] [CrossRef]
- Cardnell, R.J.G.; Behrens, C.; Diao, L.; Fan, Y.H.; Tang, X.; Tong, P.; Minna, J.D.; Mills, G.B.; Heymach, J.V.; Wistuba, I.I.; et al. An integrated molecular analysis of lung adenocarcinomas identifies potential therapeutic targets among TTF1-negative tumors, including DNA Repair Proteins and Nrf2. Clin. Cancer Res. 2015, 21, 3480–3491. [Google Scholar] [CrossRef]
- Solis, L.M.; Behrens, C.; Dong, W.; Suraokar, M.; Ozburn, N.C.; Moran, C.A.; Corvalan, A.H.; Biswal, S.; Swisher, S.G.; Bekele, B.N.; et al. Nrf2 and Keap1 abnormalities in non-small cell lung carcinoma and association with clinicopathologic features. Clin. Cancer Res. 2010, 16, 3743–3753. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gao, Z.; Li, F.; Li, X.; Sun, Y.; Wang, M.; Li, D.; Wang, R.; Li, F.; Fang, R.; et al. Whole exome sequencing identifies frequent somatic mutations in cell-cell adhesion genes in Chinese patients with lung squamous cell carcinoma. Sci. Rep. 2015, 5, 14237. [Google Scholar] [CrossRef] [PubMed]
- Hammerman, P.S.; Voet, D.; Lawrence, M.S.; Voet, D.; Jing, R.; Cibulskis, K.; Sivachenko, A.; Stojanov, P.; McKenna, A.; Lander, E.S.; et al. Comprehensive genomic characterization of squamous cell lung cancers. Nature 2012, 489, 519–525. [Google Scholar] [CrossRef]
- Singh, A.; Misra, V.; Thimmulappa, R.K.; Lee, H.; Ames, S.; Hoque, M.O.; Herman, J.G.; Baylin, S.B.; Sidransky, D.; Gabrielson, E.; et al. Dysfunctional KEAP1-NRF2 interaction in non-small-cell lung cancer. PLoS Med. 2006, 3, 1865–1876. [Google Scholar] [CrossRef]
- Li, Q.K.; Singh, A.; Biswal, S.; Askin, F.; Gabrielson, E. KEAP1 gene mutations and NRF2 activation are common in pulmonary papillary adenocarcinoma. J. Hum. Genet. 2011, 56, 230–234. [Google Scholar] [CrossRef]
- Tung, M.C.; Lin, P.L.; Wang, Y.C.; He, T.Y.; Lee, M.C.; Der Yeh, S.; Chen, C.Y.; Lee, H. Mutant p53 confers chemoresistance in non-small cell lung cancer by upregulating Nrf2. Oncotarget 2015, 6, 41692–41705. [Google Scholar] [CrossRef]
- Sasaki, H.; Suzuki, A.; Shitara, M.; Hikosaka, Y.; Okuda, K.; Moriyama, S.; Yano, M.; Fujii, Y. Genotype analysis of the nrf2 gene mutation in lung cancer. Int. J. Mol. Med. 2013, 31, 1135–1138. [Google Scholar] [CrossRef]
- Onodera, Y.; Motohashi, H.; Takagi, K.; Miki, Y.; Shibahara, Y.; Watanabe, M.; Ishida, T.; Hirakawa, H.; Sasano, H.; Yamamoto, M.; et al. NRF2 immunolocalization in human breast cancer patients as a prognostic factor. Endocr. Relat. Cancer 2014, 21, 241–252. [Google Scholar] [CrossRef]
- Hartikainen, J.M.; Tengström, M.; Winqvist, R.; Jukkola-Vuorinen, A.; Pylkäs, K.; Kosma, V.M.; Soini, Y.; Mannermaa, A. KEAP1 genetic polymorphisms associate with breast cancer risk and survival outcomes. Clin. Cancer Res. 2015, 21, 1591–1601. [Google Scholar] [CrossRef]
- Hartikainen, J.M.; Tengström, M.; Kosma, V.M.; Kinnula, V.L.; Mannermaa, A.; Soini, Y. Genetic polymorphisms and protein expression of NRF2 and sulfiredoxin predict survival outcomes in breast cancer. Cancer Res. 2012, 72, 5537–5546. [Google Scholar] [CrossRef]
- Wolf, B.; Goebel, G.; Hackl, H.; Fiegl, H. Reduced mRNA expression levels of NFE2L2 are associated with poor outcome in breast cancer patients. BMC Cancer 2016, 16, 821. [Google Scholar] [CrossRef]
- Almeida, M.; Soares, M.; Ramalhinho, A.C.; Moutinho, J.F.; Breitenfeld, L. Prognosis of hormone-dependent breast cancer seems to be influenced by KEAP1, NRF2 and GSTM1 genetic polymorphisms. Mol. Biol. Rep. 2019, 46, 3213–3224. [Google Scholar] [CrossRef] [PubMed]
- Seibold, P.; Hall, P.; Schoof, N.; Nevanlinna, H.; Heikkinen, T.; Benner, A.; Liu, J.; Schmezer, P.; Popanda, O.; Flesch-Janys, D.; et al. Polymorphisms in oxidative stress-related genes and mortality in breast cancer patients—Potential differential effects by radiotherapy? Breast 2013, 22, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Roy Chowdhury, S.; Mandal, G.; Purohit, S.; Gupta, A.; Bhattacharyya, A. RelA driven co-expression of CXCL13 and CXCR5 is governed by a multifaceted transcriptional program regulating breast cancer progression. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, E.S.; Koo, J.S. Expression of pentose phosphate pathway-related proteins in breast cancer. Dis. Markers 2018, 2018. [Google Scholar] [CrossRef]
- Hart, P.C.; Ratti, B.A.; Mao, M.; Ansenberger-Fricano, K.; Shajahan-Haq, A.N.; Tyner, A.L.; Minshall, R.D.; Bonini, M.G. Caveolin-1 regulates cancer cell metabolism via scavenging Nrf2 and suppressing MnSOD-driven glycolysis. Oncotarget 2016, 7, 308–322. [Google Scholar] [CrossRef]
- Wang, K.; Johnson, A.; Ali, S.M.; Klempner, S.J.; Bekaii-Saab, T.; Vacirca, J.L.; Khaira, D.; Yelensky, R.; Chmielecki, J.; Elvin, J.A.; et al. Comprehensive Genomic Profiling of Advanced Esophageal Squamous Cell Carcinomas and Esophageal Adenocarcinomas Reveals Similarities and Differences. Oncologist 2015, 20, 1132–1139. [Google Scholar] [CrossRef]
- Erkizan, H.V.; Johnson, K.; Ghimbovschi, S.; Karkera, D.; Trachiotis, G.; Adib, H.; Hoffman, E.P.; Wadleigh, R.G. African-American esophageal squamous cell carcinoma expression profile reveals dysregulation of stress response and detox networks. BMC Cancer 2017, 17, 426. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, M.; Ying, S.; Zhang, C.; Lin, R.; Zheng, J.; Zhang, G.; Tian, D.; Guo, Y.; Du, C.; et al. Genetic Alterations in Esophageal Tissues from Squamous Dysplasia to Carcinoma. Gastroenterology 2017, 153, 166–177. [Google Scholar] [CrossRef]
- Kitano, Y.; Baba, Y.; Nakagawa, S.; Miyake, K.; Iwatsuki, M.; Ishimoto, T.; Yamashita, Y.-I.Y.I.; Yoshida, N.; Watanabe, M.; Nakao, M.; et al. Nrf2 promotes oesophageal cancer cell proliferation via metabolic reprogramming and detoxification of reactive oxygen species. J. Pathol. 2018, 244, 346–357. [Google Scholar] [CrossRef]
- Hao, J.J.; Lin, D.C.; Dinh, H.Q.; Mayakonda, A.; Jiang, Y.Y.; Chang, C.; Jiang, Y.; Lu, C.C.; Shi, Z.Z.; Xu, X.; et al. Spatial intratumoral heterogeneity and temporal clonal evolution in esophageal squamous cell carcinoma. Nat. Genet. 2016, 48, 1500–1507. [Google Scholar] [CrossRef]
- Deng, J.; Chen, H.; Zhou, D.; Zhang, J.; Chen, Y.; Liu, Q.; Ai, D.; Zhu, H.; Chu, L.; Ren, W.; et al. Comparative genomic analysis of esophageal squamous cell carcinoma between Asian and Caucasian patient populations. Nat. Commun. 2017, 8, 1533. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Kokubu, A.; Saito, S.; Narisawa-Saito, M.; Sasaki, H.; Aoyagi, K.; Yoshimatsu, Y.; Tachimori, Y.; Kushima, R.; Kiyono, T.; et al. NRF2 mutation confers malignant potential and resistance to chemoradiation therapy in advanced esophageal squamous cancer. Neoplasia 2011, 13, 864–873. [Google Scholar] [CrossRef]
- Gao, Y.B.; Chen, Z.L.; Li, J.G.; Hu, X.D.; Shi, X.J.; Sun, Z.M.; Zhang, F.; Zhao, Z.R.; Li, Z.T.; Liu, Z.Y.; et al. Genetic landscape of esophageal squamous cell carcinoma. Nat. Genet. 2014, 46, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, L.; Ou, Y.; Gao, Z.; Li, E.; Li, X.; Zhang, W.; Wang, J.; Xu, L.; Zhou, Y.; et al. Identification of genomic alterations in oesophageal squamous cell cancer. Nature 2014, 508, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, B.; Nakajima, Y.; Inazawa, J.; Inoue, J. miR-432 induces NRF2 stabilization by directly targeting KEAP1. Mol. Cancer Res. 2017, 15, 1570–1578. [Google Scholar] [CrossRef]
- Kawasaki, Y.; Ishigami, S.; Arigami, T.; Uenosono, Y.; Yanagita, S.; Uchikado, Y.; Kita, Y.; Nishizono, Y.; Okumura, H.; Nakajo, A.; et al. Clinicopathological significance of nuclear factor (erythroid-2)-related factor 2 (Nrf2) expression in gastric cancer. BMC Cancer 2015, 15, 5. [Google Scholar] [CrossRef]
- Soini, Y.; Eskelinen, M.; Juvonen, P.; Kärjä, V.; Haapasaari, K.M.; Saarela, A.; Karihtala, P. Nuclear Nrf2 expression is related to a poor survival in pancreatic adenocarcinoma. Pathol. Res. Pract. 2014, 210, 35–39. [Google Scholar] [CrossRef]
- Eichenmüller, M.; Trippel, F.; Kreuder, M.; Beck, A.; Schwarzmayr, T.; Häberle, B.; Cairo, S.; Leuschner, I.; Von Schweinitz, D.; Strom, T.M.; et al. The genomic landscape of hepatoblastoma and their progenies with HCC-like features. J. Hepatol. 2014, 61, 1312–1320. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, C.; Zhang, L.; Yang, Q.; Zhou, S.; Wen, Q.; Wang, J. Nrf2 is a potential prognostic marker and promotes proliferation and invasion in human hepatocellular carcinoma. BMC Cancer 2015, 15, 531. [Google Scholar] [CrossRef]
- Ma-on, C.; Sanpavat, A.; Whongsiri, P.; Suwannasin, S.; Hirankarn, N.; Tangkijvanich, P.; Boonla, C. Oxidative stress indicated by elevated expression of Nrf2 and 8-OHdG promotes hepatocellular carcinoma progression. Med. Oncol. 2017, 34, 57. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.L.; Lu, Y.F.; Chen, H.; Shen, Z.Y.; Liu, J. Liver expression of Nrf2-related genes in different liver diseases. Hepatobiliary Pancreat. Dis. Int. 2015, 14, 485–491. [Google Scholar] [CrossRef]
- Chen, J.; Yu, Y.; Ji, T.; Ma, R.; Chen, M.; Li, G.; Li, F.; Ding, Q.; Kang, Q.; Huang, D.; et al. Clinical implication of Keap1 and phosphorylated Nrf2 expression in hepatocellular carcinoma. Cancer Med. 2016, 5, 2678–2687. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Inoue, K.I.; Hachiya, H.; Shibuya, N.; Shimoda, M.; Kubota, K. Frequent alteration of the protein synthesis of enzymes for glucose metabolism in hepatocellular carcinomas. J. Gastroenterol. 2014, 49, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Cleary, S.P.; Jeck, W.R.; Zhao, X.; Chen, K.; Selitsky, S.R.; Savich, G.L.; Tan, T.X.; Wu, M.C.; Getz, G.; Lawrence, M.S.; et al. Identification of driver genes in hepatocellular carcinoma by exome sequencing. Hepatology 2013, 58, 1693–1702. [Google Scholar] [CrossRef]
- Shimokawa, M.; Yoshizumi, T.; Itoh, S.; Iseda, N.; Sakata, K.; Yugawa, K.; Toshima, T.; Harada, N.; Ikegami, T.; Mori, M. Modulation of Nqo1 activity intercepts anoikis resistance and reduces metastatic potential of hepatocellular carcinoma. Cancer Sci. 2020, 111, 1228–1240. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Stawiski, E.W.; Durinck, S.; Gowda, H.; Goldstein, L.D.; Barbhuiya, M.A.; Schröder, M.S.; Sreenivasamurthy, S.K.; Kim, S.W.; Phalke, S.; et al. Integrated genomic analysis reveals mutated ELF3 as a potential gallbladder cancer vaccine candidate. Nat. Commun. 2020, 11, 4225. [Google Scholar] [CrossRef]
- Zhan, M.; Wang, H.; Xu, S.W.; Yang, L.H.; Chen, W.; Zhao, S.X.; Shen, H.; Liu, Q.; Yang, R.M.; Wang, J. Variants in oxidative stress-related genes affect the chemosensitivity through Nrf2-mediated signaling pathway in biliary tract cancer. EBioMedicine 2019, 48, 143–160. [Google Scholar] [CrossRef]
- Chang, L.C.; Fan, C.W.; Tseng, W.K.; Chen, J.R.; Chein, H.P.; Hwang, C.C.; Hua, C.C. Immunohistochemical study of the Nrf2 pathway in colorectal cancer: Nrf2 expression is closely correlated to keap1 in the tumor and bach1 in the normal tissue. Appl. Immunohistochem. Mol. Morphol. 2013, 21, 511–517. [Google Scholar] [CrossRef]
- Chang, L.-C.; Fan, C.-W.; Tseng, W.-K.; Chein, H.-P.; Hsieh, T.-Y.; Chen, J.-R.; Hwang, C.-C.; Hua, C.-C. The Ratio of Hmox1/Nrf2 mRNA Level in the Tumor Tissue Is a Predictor of Distant Metastasis in Colorectal Cancer. Dis. Markers 2016, 2016, 8143465. [Google Scholar] [CrossRef]
- El-Deek, H.E.M.; Ahmed, A.M.; Mohammed, R.A.A. Aberration of Nrf2-Bach1 pathway in colorectal carcinoma; role in carcinogenesis and tumor progression. Ann. Diagn. Pathol. 2019, 38, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Czogalla, B.; Kahaly, M.; Mayr, D.; Schmoeckel, E.; Niesler, B.; Kolben, T.; Burges, A.; Mahner, S.; Jeschke, U.; Trillsch, F. Interaction of ERα and NRF2 impacts survival in ovarian cancer patients. Int. J. Mol. Sci. 2019, 20, 112. [Google Scholar] [CrossRef] [PubMed]
- Beinse, G.; Just, P.A.; Rance, B.; Izac, B.; Letourneur, F.; Saidu, N.E.B.; Chouzenoux, S.; Nicco, C.; Goldwasser, F.; Pasmant, E.; et al. The NRF2 transcriptional target NQO1 has low mRNA levels in TP53-mutated endometrial carcinomas. PLoS ONE 2019, 14, e0214416. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Yi, X.; Abushahin, N.; Pang, S.; Zhang, D.; Kong, B.; Zheng, W. Nrf2 expression in endometrial serous carcinomas and its precancers. Int. J. Clin. Exp. Pathol. 2011, 4, 85–96. [Google Scholar]
- Stacy, D.R.; Ely, K.; Massion, P.P.; Yarbrough, W.G.; Hallahan, D.E.; Sekhar, K.R.; Freeman, M.L. Increased expression of nuclear factor E2 p45-related factor 2 (NRF2) in head and neck squamous cell carcinomas. Head Neck 2006, 28, 813–818. [Google Scholar] [CrossRef]
- Namani, A.; Matiur Rahaman, M.; Chen, M.; Tang, X. Gene-expression signature regulated by the KEAP1-NRF2-CUL3 axis is associated with a poor prognosis in head and neck squamous cell cancer. BMC Cancer 2018, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- Hämälaïnen, M.; Teppo, H.R.; Skarp, S.; Haapasaari, K.M.; Porvari, K.; Vuopala, K.; Kietzmann, T.; Karihtala, P. NRF1 and NRF2 mRNA and Protein Expression Decrease Early during Melanoma Carcinogenesis: An Insight into Survival and MicroRNAs. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Kim, Y.R.; Oh, J.E.; Kim, M.S.; Kang, M.R.; Park, S.W.; Han, J.Y.; Eom, H.S.; Yoo, N.J.; Lee, S.H. Oncogenic NRF2 mutations in squamous cell carcinomas of oesophagus and skin. J. Pathol. 2010, 220, 446–451. [Google Scholar] [CrossRef]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen Peroxide Sensing and Signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef]
- Hanschmann, E.-M.; Godoy, J.R.; Berndt, C.; Hudemann, C.; Lillig, C.H. Thioredoxins, glutaredoxins, and peroxiredoxins-molecular mechanisms and health significance: From cofactors to antioxidants to redox signaling. Antioxid. Redox Signal. 2013, 19, 1539–1605. [Google Scholar] [CrossRef]
- Holmgren, A. Thioredoxin. Annu. Rev. Biochem. 1985, 54, 237–271. [Google Scholar] [CrossRef] [PubMed]
- Yao, A.; Storr, S.J.; Al-hadyan, K.; Rahman, R.; Smith, S.; Grundy, R.; Paine, S.; Martin, S.G. Thioredoxin System Protein Expression Is Associated with Poor Clinical Outcome in Adult and Paediatric Gliomas and Medulloblastomas. Mol. Neurobiol. 2020, 57, 2889–2901. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; McGrath, K.L.; Di Trapani, G.; Charoentong, P.; Shah, F.; King, M.M.; Clarke, F.M.; Tonissen, K.F. The thioredoxin system in breast cancer cell invasion and migration. Redox Biol. 2016, 8, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Tobe, R.; Carlson, B.A.; Tsuji, P.A.; Lee, B.J.; Gladyshev, V.N.; Hatfield, D.L. Differences in redox regulatory systems in human lung and liver tumors suggest different avenues for therapy. Cancers 2015, 7, 2262–2276. [Google Scholar] [CrossRef]
- Ran, D.-M.; Zhang, Q.-W.; Su, H.-L.; Wang, C.; Gao, F.-H. Expression of thioredoxin reductase-1 and its effect in non-small cell lung cancer. Int. J. Clin. Exp. Med. 2016, 9, 7608–7614. [Google Scholar]
- Lee, J.R.; Roh, J.-L.; Lee, S.M.; Park, Y.; Cho, K.-J.; Choi, S.-H.; Nam, S.Y.; Kim, S.Y. Overexpression of glutathione peroxidase 1 predicts poor prognosis in oral squamous cell carcinoma. J. Cancer Res. Clin. Oncol. 2017, 143, 2257–2265. [Google Scholar] [CrossRef]
- Iwasawa, S.; Yamano, Y.; Takiguchi, Y.; Tanzawa, H.; Tatsumi, K.; Uzawa, K. Upregulation of thioredoxin reductase 1 in human oral squamous cell carcinoma. Oncol. Rep. 2011, 25, 637–644. [Google Scholar] [CrossRef]
- Zhu, X.; Huang, C.; Peng, B. Overexpression of thioredoxin system proteins predicts poor prognosis in patients with squamous cell carcinoma of the tongue. Oral Oncol. 2011, 47, 609–614. [Google Scholar] [CrossRef]
- Bu, L.; Li, W.; Ming, Z.; Shi, J.; Fang, P.; Yang, S. Inhibition of TrxR2 suppressed NSCLC cell proliferation, metabolism and induced cell apoptosis through decreasing antioxidant activity. Life Sci. 2017, 178, 35–41. [Google Scholar] [CrossRef]
- Yoon, B.-I.; Kim, Y.H.; Yi, J.-Y.; Kang, M.-S.; Jang, J.-J.; Joo, K.-H.; Kim, Y.; McHugh Law, J.; Kim, D.-Y. Expression of thioredoxin during progression of hamster and human cholangiocarcinoma. Cancer Sci. 2010, 101, 281–288. [Google Scholar] [CrossRef]
- Nagano, M.; Hatakeyama, K.; Kai, M.; Nakamura, H.; Yodoi, J.; Asada, Y.; Chijiiwa, K. Nuclear expression of thioredoxin-1 in the invasion front is associated with outcome in patients with gallbladder carcinoma. HPB 2012, 14, 573–582. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Abdel-Hamid, N.M.; Mahmoud, T.K.; Abass, S.A.; El-Shishtawy, M.M. Expression of thioredoxin and glutaredoxin in experimental hepatocellular carcinoma—Relevance for prognostic and diagnostic evaluation. Pathophysiology 2018, 25, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, D.T.; Al-Yatama, F.; Mohammed, F.M.A.; Al-Banaw, A.G.; Al-Bader, M.; Burge, M.; Sinowatz, F.; Singal, P.K. Thioredoxin and thioredoxin reductase expression in thyroid cancer depends on tumour aggressiveness. Anticancer Res. 2010, 30, 767–776. [Google Scholar] [PubMed]
- Gollapalli, K.; Ghantasala, S.; Atak, A.; Rapole, S.; Moiyadi, A.; Epari, S.; Srivastava, S. Tissue Proteome Analysis of Different Grades of Human Gliomas Provides Major Cues for Glioma Pathogenesis. Omics J. Integr. Biol. 2017, 21, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Zhang, P.; Zuo, Z.; Wang, F.; Bi, R.; Shang, W.; Wu, A.; Ye, J.; Li, S.; Sun, X.; et al. Thioredoxin-1 promotes colorectal cancer invasion and metastasis through crosstalk with S100P. Cancer Lett. 2017, 401, 1–10. [Google Scholar] [CrossRef]
- Lim, J.Y.; Yoon, S.O.; Hong, S.W.; Kim, J.W.; Choi, S.H.; Cho, J.Y. Thioredoxin and thioredoxin-interacting protein as prognostic markers for gastric cancer recurrence. World J. Gastroenterol. 2012, 18, 5581–5588. [Google Scholar] [CrossRef]
- Shang, W.; Xie, Z.; Lu, F.; Fang, D.; Tang, T.; Bi, R.; Chen, L.; Jiang, L. Increased Thioredoxin-1 Expression Promotes Cancer Progression and Predicts Poor Prognosis in Patients with Gastric Cancer. Oxid. Med. Cell. Longev. 2019, 2019. [Google Scholar] [CrossRef]
- Samaranayake, G.J.; Troccoli, C.I.; Huynh, M.; Lyles, R.D.Z.; Kage, K.; Win, A.; Lakshmanan, V.; Kwon, D.; Ban, Y.; Chen, S.X.; et al. Thioredoxin-1 protects against androgen receptor-induced redox vulnerability in castration-resistant prostate cancer. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Li, J.; Yue, Z.; Xiong, W.; Sun, P.; You, K.; Wang, J. TXNIP overexpression suppresses proliferation and induces apoptosis in SMMC7221 cells through ROS generation and MAPK pathway activation. Oncol. Rep. 2017, 37, 3369–3376. [Google Scholar] [CrossRef]
- Xie, M.H.; Xie, R.; Xie, S.; Wu, Y.; Wang, W.; Li, X.; Xu, Y.; Liu, B.; Zhou, Y.; Wang, T.; et al. Thioredoxin interacting protein (TXNIP) acts as a tumor suppressor in human prostate cancer. Cell Biol. Int. 2020. [Google Scholar] [CrossRef]
- Morrison, J.A.; Pike, L.A.; Sams, S.B.; Sharma, V.; Zhou, Q.; Severson, J.J.; Tan, A.-C.; Wood, W.M.; Haugen, B.R. Thioredoxin interacting protein (TXNIP) is a novel tumor suppressor in thyroid cancer. Mol. Cancer 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Noura, M.; Matsuo, H.; Koyama, A.; Adachi, S.; Masutani, H. TXNIP induces growth arrest and enhances ABT263-induced apoptosis in mixed-lineage leukemia-rearranged acute myeloid leukemia cells. FEBS Open Bio 2020, 10, 1532–1541. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; Huang, W.; Zhang, W.; Xu, J.; Song, W.; Wang, Y.; Zhu, A.; Luo, J.; Huang, G.; Wang, Y.; et al. TXNIP interaction with the Her-1/2 pathway contributes to overall survival in breast cancer. Oncotarget 2015, 6, 3003–3012. [Google Scholar] [CrossRef] [PubMed]
- da Motta, L.L.; De Bastiani, M.A.; Stapenhorst, F.; Klamt, F. Oxidative stress associates with aggressiveness in lung large-cell carcinoma. Tumor Biol. 2015, 36, 4681–4688. [Google Scholar] [CrossRef]
- Mo, R.; Peng, J.; Xiao, J.; Ma, J.; Li, W.; Wang, J.; Ruan, Y.; Ma, S.; Hong, Y.; Wang, C.; et al. High TXNDC5 expression predicts poor prognosis in renal cell carcinoma. Tumor Biol. 2016, 37, 9797–9806. [Google Scholar] [CrossRef]
- Tan, F.; Zhu, H.; He, X.; Yu, N.; Zhang, X.; Xu, H.; Pei, H. Role of TXNDC5 in tumorigenesis of colorectal cancer cells: In vivo and in vitro evidence. Int. J. Mol. Med. 2018, 42, 935–945. [Google Scholar] [CrossRef]
- Vincent, E.E.; Elder, D.J.E.; Phillips, L.; Heesom, K.J.; Pawade, J.; Luckett, M.; Sohail, M.; May, M.T.; Hetzel, M.R.; Tavaré, J.M. Overexpression of the TXNDC5 protein in non-small cell lung carcinoma. Anticancer Res. 2011, 31, 1577–1582. [Google Scholar]
- Wu, Z.; Zhang, L.; Li, N.; Sha, L.; Zhang, K. An immunohistochemical study of thioredoxin domain-containing 5 expression in gastric adenocarcinoma. Oncol. Lett. 2015, 9, 1154–1158. [Google Scholar] [CrossRef][Green Version]
- Lu, A.; Wangpu, X.; Han, D.; Feng, H.; Zhao, J.; Ma, J.; Qu, S.; Chen, X.; Liu, B.; Zheng, M. TXNDC9 expression in colorectal cancer cells and its influence on colorectal cancer prognosis. Cancer Investig. 2012, 30, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Zhao, R.; Sun, F.; Lu, Q.; Wang, X.; Hu, J.; Wang, S.; Gao, L.; Zhou, Q.; Xiong, X.; et al. TXNDC9 regulates oxidative stress-induced androgen receptor signaling to promote prostate cancer progression. Oncogene 2020, 39, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-F.; Wang, X.-Y.; Fu, Z.-Q.; Peng, Q.-H.; Zhang, J.-Y.; Ye, F.; Fu, Y.-F.; Zhou, C.-Y.; Lu, W.-G.; Cheng, X.-D.; et al. TXNDC17 promotes paclitaxel resistance via inducing autophagy in ovarian cancer. Autophagy 2015, 11, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Sobhani, M.; Taheri, A.-R.; Jafarian, A.-H.; Hashemy, S.I. The activity and tissue distribution of thioredoxin reductase in basal cell carcinoma. J. Cancer Res. Clin. Oncol. 2016, 142, 2303–2307. [Google Scholar] [CrossRef] [PubMed]
- Söderberg, A.; Sahaf, B.; Rosén, A. Thioredoxin reductase, a redox-active selenoprotein, is secreted by normal and neoplastic cells: Presence in human plasma. Cancer Res. 2000, 60, 2281–2289. [Google Scholar] [PubMed]
- Shao, L.-E.; Tanaka, T.; Gribi, R.; Yu, J.; Diccianni, M.B.; Yu, A.L.; Pullen, J.D.; Camitta, B.M. Thioredoxin expression in primary T-cell acute lymphoblastic leukemia and its therapeutic implication. Cancer Res. 2001, 61, 7333–7338. [Google Scholar]
- Cai, Z.; Zhang, C.; Zou, Y.; Lu, C.; Hu, H.; Qian, J.; Jiang, L.; Hu, G. Tissue thioredoxin-interacting protein expression predicted recurrence in patients with meningiomas. Int. J. Clin. Oncol. 2017, 22, 660–666. [Google Scholar] [CrossRef]
- Cadenas, C.; Franckenstein, D.; Schmidt, M.; Gehrmann, M.; Hermes, M.; Geppert, B.; Schormann, W.; Maccoux, L.J.; Schug, M.; Schumann, A.; et al. Role of thioredoxin reductase 1 and thioredoxin interacting protein in prognosis of breast cancer. Breast Cancer Res. 2010, 12. [Google Scholar] [CrossRef]
- Shen, L.; O’Shea, J.M.; Kaadige, M.R.; Cunha, S.; Wilde, B.R.; Cohen, A.L.; Welm, A.L.; Ayer, D.E. Metabolic reprogramming in triple-negative breast cancer through Myc suppression of TXNIP. Proc. Natl. Acad. Sci. USA 2015, 112, 5425–5430. [Google Scholar] [CrossRef]
- Zhu, H.; Tao, X.; Zhou, L.; Sheng, B.; Zhe, X.; Zhu, X. Expression of thioredoxin 1 and peroxiredoxins in squamous cervical carcinoma and its predictive role in NACT. BMC Cancer 2019, 19. [Google Scholar] [CrossRef]
- Lu, Y.; Zhao, X.; Li, K.; Luo, G.; Nie, Y.; Shi, Y.; Zhou, Y.; Ren, G.; Feng, B.; Liu, Z.; et al. Thioredoxin-like protein 2 is overexpressed in colon cancer and promotes cancer cell metastasis by interaction with ran. Antioxid. Redox Signal. 2013, 19, 899–911. [Google Scholar] [CrossRef]
- Peng, W.; Zhou, Z.; Zhong, Y.; Sun, Y.; Wang, Y.; Zhu, Z.; Jiao, W.; Bai, M.; Sun, J.; Yin, H.; et al. Plasma activity of Thioredoxin Reductase as a Novel Biomarker in Gastric Cancer. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Zhang, W.-L.; Sun, K.-Y.; Tan, Q.; Diao, X.-F.; Liu, T. Expression and prognosis of TrxR in non-small cell lung cancer tissues. Chin. J. Cancer Prev. Treat. 2016, 23, 1224–1228. [Google Scholar]
- Fernandes, A.P.; Capitanio, A.; Selenius, M.; Brodin, O.; Rundlöf, A.-K.; Björnstedt, M. Expression profiles of thioredoxin family proteins in human lung cancer tissue: Correlation with proliferation and differentiation. Histopathology 2009, 55, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Pylväs, M.; Puistola, U.; Kauppila, S.; Soini, Y.; Karihtala, P. Oxidative stress-induced antioxidant enzyme expression is an early phenomenon in ovarian carcinogenesis. Eur. J. Cancer 2010, 46, 1661–1667. [Google Scholar] [CrossRef]
- Chaiswing, L.; Zhong, W.; Oberley, T.D. Increasing discordant antioxidant protein levels and enzymatic activities contribute to increasing redox imbalance observed during human prostate cancer progression. Free Radic. Biol. Med. 2014, 67, 342–352. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Metere, A.; Frezzotti, F.; Graves, C.E.; Vergine, M.; De Luca, A.; Pietraforte, D.; Giacomelli, L. A possible role for selenoprotein glutathione peroxidase (GPx1) and thioredoxin reductases (TrxR1) in thyroid cancer: Our experience in thyroid surgery. Cancer Cell Int. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, P.; Ramasamy, P.; Ramasamy, P.; Larkin, A.-M.; Larkin, A.-M.; Linge, A.; Tiernan, D.; McAree, F.; Horgan, N.; Moriarty, P.; et al. PRDX3 is associated with metastasis and poor survival in uveal melanoma. J. Clin. Pathol. 2020, 73, 408–412. [Google Scholar] [CrossRef]
- González, R.; Rodríguez-Hernández, M.A.; Negrete, M.; Ranguelova, K.; Rossin, A.; Choya-Foces, C.; Cruz-Ojeda, P.D.L.; Miranda-Vizuete, A.; Martínez-Ruiz, A.; Rius-Pérez, S.; et al. Downregulation of thioredoxin-1-dependent CD95 S-nitrosation by Sorafenib reduces liver cancer. Redox Biol. 2020, 34. [Google Scholar] [CrossRef]
- Shen, X.; Burguillos, M.A.; Osman, A.M.; Frijhoff, J.; Carrillo-Jiménez, A.; Kanatani, S.; Augsten, M.; Saidi, D.; Rodhe, J.; Kavanagh, E.; et al. Glioma-induced inhibition of caspase-3 in microglia promotes a tumor-supportive phenotype. Nat. Immunol. 2016, 17, 1282–1290. [Google Scholar] [CrossRef]
- Meuillet, E.J.; Mahadevan, D.; Berggren, M.; Coon, A.; Powis, G. Thioredoxin-1 binds to the C2 domain of PTEN inhibiting PTEN’s lipid phosphatase activity and membrane binding: A mechanism for the functional loss of PTEN’s tumor suppressor activity. Arch. Biochem. Biophys. 2004, 429, 123–133. [Google Scholar] [CrossRef]
- Zuo, Z.; Zhang, P.; Lin, F.; Shang, W.; Bi, R.; Lu, F.; Wu, J.; Jiang, L. Interplay between Trx-1 and S100P promotes colorectal cancer cell epithelial–mesenchymal transition by up-regulating S100A4 through AKT activation. J. Cell. Mol. Med. 2018, 22, 2430–2441. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Lu, W.; Han, Y.; Yang, J.; Jiang, W.; You, X.; Luo, Y.; Wen, S.; Hu, Y.; et al. Mitochondrial TXNRD3 confers drug resistance via redox-mediated mechanism and is a potential therapeutic target in vivo. Redox Biol. 2020, 36. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Wang, G.; Zhang, J.; Han, Q. Inhibiting TrxR suppresses liver cancer by inducing apoptosis and eliciting potent antitumor immunity. Oncol. Rep. 2018, 40, 3447–3457. [Google Scholar] [CrossRef] [PubMed]
- Raninga, P.V.; Di Trapani, G.; Vuckovic, S.; Tonissen, K.F. Cross-talk between two antioxidants, thioredoxin reductase and heme oxygenase-1, and therapeutic implications for multiple myeloma. Redox Biol. 2016, 8, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.; Quan, Y.; Zhan, M.; Liao, H.; Li, Y.; Lu, L. MiR-125b-5p inhibits cell proliferation, migration, and invasion in hepatocellular carcinoma via targeting TXNRD1. Cancer Cell Int. 2019, 19. [Google Scholar] [CrossRef]
- Hao, C.; Xu, X.; Ma, J.; Xia, J.; Dai, B.; Liu, L.; Ma, Y. MicroRNA-124 regulates the radiosensitivity of non-small cell lung cancer cells by targeting TXNRD1. Oncol. Lett. 2017, 13, 2071–2078. [Google Scholar] [CrossRef]
- Xu, Z.; Zhang, Y.; Ding, J.; Hu, W.; Tan, C.; Wang, M.; Tang, J.; Xu, Y. miR-17-3p Downregulates Mitochondrial Antioxidant Enzymes and Enhances the Radiosensitivity of Prostate Cancer Cells. Mol. Ther. Nucleic Acids 2018, 13, 64–77. [Google Scholar] [CrossRef]
- Engelman, R.; Ziv, T.; Arnér, E.S.J.; Benhar, M. Inhibitory nitrosylation of mammalian thioredoxin reductase 1: Molecular characterization and evidence for its functional role in cellular nitroso-redox imbalance. Free Radic. Biol. Med. 2016, 97, 375–385. [Google Scholar] [CrossRef]
- Wright, D.E.; Altaany, Z.; Bi, Y.; Alperstein, Z.; O’Donoghue, P. Acetylation regulates thioredoxin reductase oligomerization and activity. Antioxid. Redox Signal. 2018, 29, 377–388. [Google Scholar] [CrossRef]
- Nagaraj, K.; Lapkina-Gendler, L.; Sarfstein, R.; Gurwitz, D.; Pasmanik-Chor, M.; Laron, Z.; Yakar, S.; Werner, H. Identification of thioredoxin-interacting protein (TXNIP) as a downstream target for IGF1 action. Proc. Natl. Acad. Sci. USA 2018, 115, 1045–1050. [Google Scholar] [CrossRef]
- Li, Y.; Miao, L.-Y.; Xiao, Y.-L.; Huang, M.; Yu, M.; Meng, K.; Cai, H.-R. Hypoxia induced high expression of thioredoxin interacting protein (TXNIP) in non-small cell lung cancer and its prognostic effect. Asian Pac. J. Cancer Prev. 2015, 16, 2953–2958. [Google Scholar] [CrossRef]
- Li, W.; Wu, Z.; Ma, Q.; Liu, J.; Xu, Q.; Han, L.; Duan, W.; Lv, Y.; Wang, F.; Reindl, K.M.; et al. Hyperglycemia regulates TXNIP/TRX/ROS axis via p38 MAPK and ERK pathways in pancreatic cancer. Curr. Cancer Drug Targets 2014, 14, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Ayer, D.E. Ras suppresses TXNIP expression by restricting ribosome translocation. Mol. Cell. Biol. 2018, 38. [Google Scholar] [CrossRef] [PubMed]
- Kuljaca, S.; Liu, T.; Dwarte, T.; Kavallaris, M.; Haber, M.; Norris, M.D.; Martin-Caballero, J.; Marshall, G.M. The cyclin-dependent kinase inhibitor, p21WAF1, promotes angiogenesis by repressing gene transcription of thioredoxin-binding protein 2 in cancer cells. Carcinogenesis 2009, 30, 1865–1871. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Zhou, L.; Liu, H.; Shan, Y.; Zhang, X. MicroRNA-224 promotes pancreatic cancer cell proliferation and migration by targeting the TXNIP-mediated HIF1α pathway. Cell. Physiol. Biochem. 2018, 48, 1735–1746. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Dang, B.-L.; Huang, J.; Chen, M.; Wu, D.; Xu, M.-L.; Li, R.; Yan, G.-R. MiR-373 drives the epithelial-to-mesenchymal transition and metastasis via the miR-373-TXNIP-HIF1a-TWIST signaling axis in breast cancer. Oncotarget 2015, 6, 32701–32712. [Google Scholar] [CrossRef]
- Masaki, S.; Masutani, H.; Yoshihara, E.; Yodoi, J. Deficiency of Thioredoxin binding protein-2 (TBP-2) enhances TGF-β signaling and promotes epithelial to mesenchymal transition. PLoS ONE 2012, 7, e39900. [Google Scholar] [CrossRef]
- Zhang, B.; Lyu, J.; Yang, E.J.; Liu, Y.; Wu, C.; Pardeshi, L.; Tan, K.; Chen, Q.; Xu, X.; Deng, C.-X.; et al. Class I histone deacetylase inhibition is synthetic lethal with BRCA1 deficiency in breast cancer cells. Acta Pharm. Sin. B 2020, 10, 615–627. [Google Scholar] [CrossRef]
- Lee, J.-H.; Jeong, E.-G.; Choi, M.-C.; Kim, S.-H.; Park, J.-H.; Song, S.-H.; Park, J.; Bang, Y.-J.; Kim, T.-Y. Inhibition of histone deacetylase 10 induces thioredoxin-interacting protein and causes accumulation of reactive oxygen species in SNU-620 human gastric cancer cells. Mol. Cells 2010, 30, 107–112. [Google Scholar] [CrossRef]
- Zhang, B.; Lyu, J.; Liu, Y.; Wu, C.; Yang, E.J.; Pardeshi, L.; Tan, K.; Wong, K.H.; Chen, Q.; Xu, X.; et al. BRCA1 deficiency sensitizes breast cancer cells to bromodomain and extra-terminal domain (BET) inhibition. Oncogene 2018, 37, 6341–6356. [Google Scholar] [CrossRef]
- Hong, S.Y.; Yu, F.-X.; Luo, Y.; Hagen, T. Oncogenic activation of the PI3K/Akt pathway promotes cellular glucose uptake by downregulating the expression of thioredoxin-interacting protein. Cell. Signal. 2016, 28, 377–383. [Google Scholar] [CrossRef]
- Ho, B.; Huang, G.; Golubovskaya, V.M. Focal adhesion kinase regulates expression of thioredoxin-interacting protein (TXNIP) in cancer cells. Anticancer Agents Med. Chem. 2014, 14, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Riahi, Y.; Kaiser, N.; Cohen, G.; Abd-Elrahman, I.; Blum, G.; Shapira, O.M.; Koler, T.; Simionescu, M.; Sima, A.V.; Zarkovic, N.; et al. Foam cell-derived 4-hydroxynonenal induces endothelial cell senescence in a TXNIP-dependent manner. J. Cell. Mol. Med. 2015, 19, 1887–1899. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G.; Martinotti, S.; Fazio, V.; Manzari, V.; Paradisi, L.; Parola, M.; Frati, L.; Dianzani, M.U. Effect of 4-Hydroxynonenal on c-myc Expression. Toxicol. Pathol. 1987, 15, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, E.; Mohankumar, K.; Lacey, A.; Safe, S. Inhibition of NR4A1 promotes ROS accumulation and IL24-dependent growth arrest in rhabdomyosarcoma. Mol. Cancer Res. 2019, 17, 2221–2232. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, E.; Lee, S.-O.; Kim, G.; Abdelrahim, M.; Jin, U.-H.; Safe, S.; Abudayyeh, A. Nuclear receptor 4A1 (NR4A1) as a drug target for renal cell adenocarcinoma. PLoS ONE 2015, 10, e0128308. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, E.; Lee, S.-O.; Doddapaneni, R.; Singh, M.; Safe, S. Nuclear receptor 4A1 as a drug target for breast cancer chemotherapy. Endocr. Relat. Cancer 2015, 22, 831–840. [Google Scholar] [CrossRef]
- Xu, B.; Li, J.; Liu, X.; Li, C.; Chang, X. TXNDC5 is a cervical tumor susceptibility gene that stimulates cell migration, vasculogenic mimicry and angiogenesis by downregulating SERPINF1 and TRAF1 expression. Oncotarget 2017, 8, 91009–91024. [Google Scholar] [CrossRef]
- Wang, L.; Song, G.; Chang, X.; Tan, W.; Pan, J.; Zhu, X.; Liu, Z.; Qi, M.; Yu, J.; Han, B. The role of TXNDC5 in castration-resistant prostate cancer—Involvement of androgen receptor signaling pathway. Oncogene 2015, 34, 4735–4745. [Google Scholar] [CrossRef]
- Gurjar, S.A.; Wheeler, J.X.; Wadhwa, M.; Thorpe, R.; Kimber, I.; Derrick, J.P.; Dearman, R.J.; Metcalfe, C. The impact of thioredoxin reduction of allosteric disulfide bonds on the therapeutic potential of monoclonal antibodies. J. Biol. Chem. 2019, 294, 19616–19634. [Google Scholar] [CrossRef]
- Roh, J.-L.; Jang, H.; Kim, E.H.; Shin, D. Targeting of the Glutathione, Thioredoxin, and Nrf2 Antioxidant Systems in Head and Neck Cancer. Antioxid. Redox Signal. 2017, 27, 106–114. [Google Scholar] [CrossRef]
- Chen, X.; Tang, W.; Liu, S.; Yu, L.; Chen, Z. Thioredoxin-1 phosphorylated at T100 is needed for its anti-apoptotic activity in HepG2 cancer cells. Life Sci. 2010, 87, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, M.; Kwon, Y.-W.; Yodoi, J.; Masutani, H. Thioredoxin regulates cell cycle via the ERK1/2-cyclin d1 pathway. Antioxid. Redox Signal. 2009, 11, 2957–2971. [Google Scholar] [CrossRef] [PubMed]
- Reynoso, E.; Liu, H.; Li, L.; Yuan, A.L.; Chen, S.; Wang, Z. Thioredoxin-1 actively maintains the pseudokinase MLKL in a reduced state to suppress disulfide bond-dependent MLKL polymer formation and necroptosis. J. Biol. Chem. 2017, 292, 17514–17524. [Google Scholar] [CrossRef] [PubMed]
- Farina, A.R.; Cappabianca, L.; Desantis, G.; Ianni, N.D.; Ruggeri, P.; Ragone, M.; Merolle, S.; Tonissen, K.F.; Gulino, A.; MacKay, A.R. Thioredoxin stimulates MMP-9 expression, de-regulates the MMP-9/TIMP-1 equilibrium and promotes MMP-9 dependent invasion in human MDA-MB-231 breast cancer cells. FEBS Lett. 2011, 585, 3328–3336. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Kan, G.; Mao, Y.; Wu, Z.; Tan, X.; He, H.; Lee, C. UHRF1 downmodulation enhances antitumor effects of histone deacetylase inhibitors in retinoblastoma by augmenting oxidative stress-mediated apoptosis. Mol. Oncol. 2020, 14, 329–346. [Google Scholar] [CrossRef]
- Chepikova, O.E.; Malin, D.; Strekalova, E.; Lukasheva, E.V.; Zamyatnin, A.A.; Cryns, V.L. Lysine oxidase exposes a dependency on the thioredoxin antioxidant pathway in triple-negative breast cancer cells. Breast Cancer Res. Treat. 2020. [Google Scholar] [CrossRef]
- Shin, B.; Feser, R.; Nault, B.; Hunter, S.; Maiti, S.; Ugwuagbo, K.C.; Majumder, M. miR526b and miR655 induce oxidative stress in breast cancer. Int. J. Mol. Sci. 2019, 20, 4039. [Google Scholar] [CrossRef]
- Yu, J.; Yang, M.; Zhou, B.; Luo, J.; Zhang, Z.; Zhang, W.; Yan, Z. CircRNA-104718 acts as competing endogenous RNA and promotes hepatocellular carcinoma progression through microRNA-218-5p/TXNDC5 signaling pathway. Clin. Sci. 2019, 133, 1487–1503. [Google Scholar] [CrossRef]
- Chawsheen, H.A.; Jiang, H.; Ying, Q.; Ding, N.; Thapa, P.; Wei, Q. The redox regulator sulfiredoxin forms a complex with thioredoxin domain–containing 5 protein in response to ER stress in lung cancer cells. J. Biol. Chem. 2019, 294, 8991–9006. [Google Scholar] [CrossRef]
- Kamitori, K.; Yamaguchi, F.; Dong, Y.; Hossain, A.; Katagi, A.; Noguchi, C.; Hirata, Y.; Tsukamoto, I.; Hatano, N.; Tokuda, M. Both Ser361 phosphorylation and the C-arrestin domain of thioredoxin interacting protein are important for cell cycle blockade at the G1/S checkpoint. FEBS Open Bio 2018, 8, 1804–1819. [Google Scholar] [CrossRef]
- Qu, X.; Sun, J.; Zhang, Y.; Li, J.; Hu, J.; Li, K.; Gao, L.; Shen, L. c-Myc-driven glycolysis via TXNIP suppression is dependent on glutaminase-MondoA axis in prostate cancer. Biochem. Biophys. Res. Commun. 2018, 504, 415–421. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, C.C.; Townsend, D.M.; Tew, K.D. Glutathione S-transferase polymorphisms: Cancer incidence and therapy. Oncogene 2006, 25, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Mukthapura, A.; Shimogga, A.; Sudha, K.V.; Shetty, B.; Rao, G.M. Oxidative products of proteins and antioxidant potential of thiols in gastric carcinoma patients. J. Med. Biochem. 2010, 29, 102–106. [Google Scholar] [CrossRef]
- Khan, S.U.; Mahjabeen, I.; Malik, F.A.; Kayani, M.A. Prognostic significance of altered blood and tissue glutathione levels in head and neck squamous cell carcinoma cases. Asian Pac. J. Cancer Prev. 2014, 15, 7603–7609. [Google Scholar] [CrossRef] [PubMed]
- Murawaki, Y.; Tsuchiya, H.; Kanbe, T.; Harada, K.; Yashima, K.; Nozaka, K.; Tanida, O.; Kohno, M.; Mukoyama, T.; Nishimuki, E.; et al. Aberrant expression of selenoproteins in the progression of colorectal cancer. Cancer Lett. 2008, 259, 218–230. [Google Scholar] [CrossRef]
- Gupta, R.K.; Patel, A.K.; Kumari, R.; Chugh, S.; Shrivastav, C.; Mehra, S.; Sharma, A.N. Interactions between oxidative stress, lipid profile and antioxidants in breast cancer: A case control study. Asian Pac. J. Cancer Prev. 2012, 13, 6295–6298. [Google Scholar] [CrossRef]
- Himmetoglu, S.; Dincer, Y.; Ersoy, Y.E.; Bayraktar, B.; Celik, V.; Akcay, T. DNA oxidation and antioxidant status in breast cancer. J. Investig. Med. 2009, 57, 720–723. [Google Scholar] [CrossRef]
- Sehitogullari, A.; Aslan, M.; Sayir, F.; Kahraman, A.; Demir, H. Serum paraoxonase-1 enzyme activities and oxidative stress levels in patients with esophageal squamous cell carcinoma. Redox Rep. 2014, 19, 199–205. [Google Scholar] [CrossRef]
- Sharma, A.; Tripathi, M.; Satyam, A.; Kumar, L. Study of antioxidant levels in patients with multiple myeloma. Leuk. Lymphoma 2009, 50, 809–815. [Google Scholar] [CrossRef]
- Mohamed, M.M.; Sabet, S.; Peng, D.-F.; Nouh, M.A.; El-Shinawi, M.; El-Rifai, W. Promoter hypermethylation and suppression of glutathione peroxidase 3 are associated with inflammatory breast carcinogenesis. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef]
- Chen, Z.; Hu, T.; Zhu, S.; Mukaisho, K.; El-Rifai, W.; Peng, D.-F. Glutathione peroxidase 7 suppresses cancer cell growth and is hypermethylated in gastric cancer. Oncotarget 2017, 8, 54345–54356. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Bai, W.; Huang, F.; Tang, J.; Lin, X. Downregulation of microRNA-196a inhibits stem cell self-renewal ability and stemness in non-small-cell lung cancer through upregulating GPX3 expression. Int. J. Biochem. Cell Biol. 2019, 115. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Shi, S.; Liang, C.; Liang, D.; Hua, J.; Zhang, B.; Xu, J.; Yu, X. Abrogation of glutathione peroxidase-1 drives EMT and chemoresistance in pancreatic cancer by activating ROS-mediated Akt/GSK3β/snail signaling. Oncogene 2018, 37, 5843–5857. [Google Scholar] [CrossRef] [PubMed]
- Naiki, T.; Naiki-Ito, A.; Iida, K.; Etani, T.; Kato, H.; Suzuki, S.; Yamashita, Y.; Kawai, N.; Yasui, T.; Takahashi, S. GPX2 promotes development of bladder cancer with squamous cell differentiation through the control of apoptosis. Oncotarget 2018, 9, 15847–15859. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cao, P.; Alshwmi, M.; Jiang, N.; Xiao, Z.; Jiang, F.; Gu, J.; Wang, X.; Sun, X.; Li, S. GPX2 suppression of H2O2 stress regulates cervical cancer metastasis and apoptosis via activation of the β-catenin-WNT pathway. Onco Targets. Ther. 2019, 12, 6639–6651. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Sun, L.; Tong, J.; Chen, X.; Li, H.; Zhang, Q. Prognostic significance of glutathione peroxidase 2 in gastric carcinoma. Tumor Biol. 2017, 39. [Google Scholar] [CrossRef]
- Kinowaki, Y.; Kurata, M.; Ishibashi, S.; Ikeda, M.; Tatsuzawa, A.; Yamamoto, M.; Miura, O.; Kitagawa, M.; Yamamoto, K. Glutathione peroxidase 4 overexpression inhibits ROS-induced cell death in diffuse large B-cell lymphoma. Lab. Investig. 2018, 98, 609–619. [Google Scholar] [CrossRef]
- Liu, X.; Sui, X.; Zhang, C.; Wei, K.; Bao, Y.; Xiong, J.; Zhou, Z.; Chen, Z.; Wang, C.; Zhu, H.; et al. Glutathione S-transferase A1 suppresses tumor progression and indicates better prognosis of human primary hepatocellular carcinoma. J. Cancer 2020, 11, 83–91. [Google Scholar] [CrossRef]
- Lu, Y.; Zhou, J.; Zhang, J.; Wang, Z.; Yu, Y.; Miao, M.; Yao, Q. Dual roles of glutathione S-transferase mu 1 in the development and metastasis of hepatocellular carcinoma. Biomed. Pharmacother. 2019, 120. [Google Scholar] [CrossRef]
- Li, J.; Wang, Q.; Yang, Y.; Lei, C.; Yang, F.; Liang, L.; Chen, C.; Xia, J.; Wang, K.; Tang, N. GSTZ1 deficiency promotes hepatocellular carcinoma proliferation via activation of the KEAP1/NRF2 pathway. J. Exp. Clin. Cancer Res. 2019, 38, 438. [Google Scholar] [CrossRef]
- Gómez-Martín, A.; Martinez-Gonzalez, L.J.; Puche-Sanz, I.; Cozar, J.M.; Lorente, J.A.; Hernández, A.F.; Alvarez-Cubero, M.J. GSTM1 gene expression and copy number variation in prostate cancer patients—Effect of chemical exposures and physical activity. Urol. Oncol. Semin. Orig. Investig. 2019, 37, 290.e9–290.e15. [Google Scholar] [CrossRef] [PubMed]
- Djukic, T.; Simic, T.; Pljesa-Ercegovac, M.; Matic, M.; Suvakov, S.; Coric, V.; Dragicevic, D.; Savic-Radojevic, A. Upregulated glutathione transferase omega-1 correlates with progression of urinary bladder carcinoma. Redox Rep. 2017, 22, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Bulus, H.; Oguztuzun, S.; Güler Simsek, G.; Kilic, M.; Ada, A.O.; Göl, S.; Kocdogan, A.K.; Kaygın, P.; Bozer, B.; Iscan, M. Expression of CYP and GST in human normal and colon tumor tissues. Biotech. Histochem. 2019, 94, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tan, N.; Liao, H.; Pan, G.; Xu, Q.; Zhu, R.; Zou, L.; He, S.; Zhu, H. High GSTP1 inhibits cell proliferation by reducing Akt phosphorylation and is associated with a better prognosis in hepatocellular carcinoma. Oncotarget 2018, 9, 8957–8971. [Google Scholar] [CrossRef] [PubMed]
- Qiu, K.; Zheng, Z.; Huang, Y. Long intergenic noncoding RNA 00844 promotes apoptosis and represses proliferation of prostate cancer cells through upregulating GSTP1 by recruiting EBF1. J. Cell. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hafez, M.M.; Al-Shabanah, O.A.; Al-Rejaie, S.S.; Al-Harbi, N.O.; Hassan, Z.K.; Alsheikh, A.; Al Theyab, A.I.; Aldelemy, M.L.; Sayed-Ahmed, M.M. Increased Hypermethylation of Glutathione S-transferase P1, DNA-binding protein inhibitor, death associated protein kinase and paired box protein-5 genes in triple-negative breast cancer Saudi females. Asian Pac. J. Cancer Prev. 2015, 16, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Richiardi, L.; Fiano, V.; Grasso, C.; Zugna, D.; Delsedime, L.; Gillio-Tos, A.; Merletti, F. Methylation of APC and GSTP1 in Non-Neoplastic Tissue Adjacent to Prostate Tumour and Mortality from Prostate Cancer. PLoS ONE 2013, 8, e68162. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.-Y.; Kim, Y.-W.; Kang, H.-W.; Kim, W.T.; Yun, S.-J.; Lee, S.-C.; Kim, W.-J.; Kim, Y.-J. DNA methylation of GSTP1 in human prostate tissues: Pyrosequencing analysis. Korean J. Urol. 2012, 53, 200–205. [Google Scholar] [CrossRef]
- Dumache, R.; Puiu, M.; Motoc, M.; Vernic, C.; Dumitrascu, V. Prostate cancer molecular detection in plasma samples by glutathione S-transferase P1 (GSTP1) methylation analysis. Clin. Lab. 2014, 60, 847–852. [Google Scholar] [CrossRef]
- Mahon, K.L.; Qu, W.; Lin, H.-M.; Spielman, C.; Cain, D.; Jacobs, C.; Stockler, M.R.; Higano, C.S.; de Bono, J.S.; Chi, K.N.; et al. Serum Free Methylated Glutathione S-transferase 1 DNA Levels, Survival, and Response to Docetaxel in Metastatic, Castration-resistant Prostate Cancer: Post Hoc Analyses of Data from a Phase 3 Trial. Eur. Urol. 2019, 76, 306–312. [Google Scholar] [CrossRef]
- Hopkins, J.; Tudhope, G.R. Glutathione Peroxidase in Human Red Cells in Health and Disease. Br. J. Haematol. 1973, 25, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Devi, G.S.; Prasad, M.H.; Saraswathi, I.; Raghu, D.; Rao, D.N.; Reddy, P.P. Free radicals antioxidant enzymes and lipid peroxidation in different types of leukemias. Clin. Chim. Acta 2000, 293, 53–62. [Google Scholar] [CrossRef]
- Oltra, A.M.; Carbonell, F.; Tormos, C.; Iradi, A.; Sáez, G.T. Antioxidant enzyme activities and the production of MDA and 8-oxo-dG in chronic lymphocytic leukemia. Free Radic. Biol. Med. 2001, 30, 1286–1292. [Google Scholar] [CrossRef]
- Moscow, J.A.; Fairchild, C.R.; Madden, M.J.; Ransom, D.T.; Wieand, H.S.; O’Brien, E.E.; Poplack, D.G.; Cossman, J.; Myers, C.E.; Cowan, K.H. Expression of Anionic Glutathione-S-transferase and P-Glycoprotein Genes in Human Tissues and Tumors. Cancer Res. 1989, 49, 1422–1428. [Google Scholar]
- Tome, M.E.; Johnson, D.B.F.; Rimsza, L.M.; Roberts, R.A.; Grogan, T.M.; Miller, T.P.; Oberley, L.W.; Briehl, M.M. A redox signature score identifies diffuse large B-cell lymphoma patients with a poor prognosis. Blood 2005, 106, 3594–3601. [Google Scholar] [CrossRef]
- Kangari, P.; Farahany, T.Z.; Golchin, A.; Ebadollahzadeh, S.; Salmaninejad, A.; Mahboob, S.A.; Nourazarian, A. Enzymatic antioxidant and lipid peroxidation evaluation in the newly diagnosed breast cancer patients in Iran. Asian Pac. J. Cancer Prev. 2018, 19, 3511–3515. [Google Scholar] [CrossRef] [PubMed]
- Diakowska, D.; Nienartowicz, M.; Grabowski, K. Activities of antioxidant enzymes in erythrocytes and tumor tissue in colorectal cancer patients. Gastroenterol. Pol. 2013, 20, 10–14. [Google Scholar]
- Zhu, X.; Wang, J.; Li, L.; Deng, L.; Wang, J.; Liu, L.; Zeng, R.; Wang, Q.; Zheng, Y. GPX3 suppresses tumor migration and invasion via the FAK/AKT pathway in esophageal squamous cell carcinoma. Am. J. Transl. Res. 2018, 10, 1908–1920. [Google Scholar]
- Handayani, E.; Edianto, D.; Sahil, M.F.; Barus, R.P.; Tobing, C.L.; Ardiansyah, E.; Yaznil, M.R. Glutathione peroxidase in ovarian cancer patients in Indonesia. S. Afr. J. Obstet. Gynaecol. 2019, 25. [Google Scholar] [CrossRef]
- Agnani, D.; Camacho-Vanegas, O.; Camacho, C.; Lele, S.; Odunsi, K.; Cohen, S.; Dottino, P.; Martignetti, J.A. Decreased levels of serum glutathione peroxidase 3 are associated with papillary serous ovarian cancer and disease progression. J. Ovarian Res. 2011, 4. [Google Scholar] [CrossRef]
- Liu, L.-D.; Zhang, Y.-N.; Wang, L.-F. GPX7 promotes the growth of human papillary thyroid carcinoma via enhancement of cell proliferation and inhibition of cell apoptosis. Transl. Cancer Res. 2019, 8, 2570–2580. [Google Scholar] [CrossRef]
- Chew, S.H.; Okazaki, Y.; Akatsuka, S.; Wang, S.; Jiang, L.; Ohara, Y.; Ito, F.; Saya, H.; Sekido, Y.; Toyokuni, S. Rheostatic CD44 isoform expression and its association with oxidative stress in human malignant mesothelioma. Free Radic. Biol. Med. 2017, 106, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Harris, I.S.; Endress, J.E.; Coloff, J.L.; Selfors, L.M.; McBrayer, S.K.; Rosenbluth, J.M.; Takahashi, N.; Dhakal, S.; Koduri, V.; Oser, M.G.; et al. Deubiquitinases Maintain Protein Homeostasis and Survival of Cancer Cells upon Glutathione Depletion. Cell Metab. 2019, 29, 1166–1181. [Google Scholar] [CrossRef] [PubMed]
- Kulak, M.V.; Cyr, A.R.; Woodfield, G.W.; Bogachek, M.; Spanheimer, P.M.; Li, T.; Price, D.H.; Domann, F.E.; Weigel, R.J. Transcriptional regulation of the GPX1 gene by TFAP2C and aberrant CpG methylation in human breast cancer. Oncogene 2013, 32, 4043–4051. [Google Scholar] [CrossRef]
- Wang, H.; Luo, K.; Tan, L.-Z.; Ren, B.-G.; Gu, L.-Q.; Michalopoulos, G.; Luo, J.-H.; Yu, Y.P. p53-induced gene 3 mediates cell death induced by glutathione peroxidase 3. J. Biol. Chem. 2012, 287, 16890–16902. [Google Scholar] [CrossRef]
- Lin, C.; Xie, L.; Lu, Y.; Hu, Z.; Chang, J. MiR-133b reverses cisplatin resistance by targeting GSTP1 in cisplatin-resistant lung cancer cells. Int. J. Mol. Med. 2018, 41, 2050–2058. [Google Scholar] [CrossRef]
- Zhang, L.; Meng, X.; Pan, C.; Qu, F.; Gan, W.; Xiang, Z.; Han, X.; Li, D. piR-31470 epigenetically suppresses the expression of glutathione S-transferase pi 1 in prostate cancer via DNA methylation. Cell. Signal. 2020, 67. [Google Scholar] [CrossRef]
- Peng, G.; Tang, Z.; Xiang, Y.; Chen, W. Glutathione peroxidase 4 maintains a stemness phenotype, oxidative homeostasis and regulates biological processes in Panc-1 cancer stem-like cells. Oncol. Rep. 2019, 41, 1264–1274. [Google Scholar] [CrossRef]
- Shireman, L.M.; Kripps, K.A.; Balogh, L.M.; Conner, K.P.; Whittington, D.; Atkins, W.M. Glutathione transferase A4-4 resists adduction by 4-hydroxynonenal. Arch. Biochem. Biophys. 2010, 504, 182–189. [Google Scholar] [CrossRef][Green Version]
- Yang, Y.; Yang, L.; Jiang, S.; Yang, T.; Lan, J.; Lei, Y.; Tan, H.; Pan, K. HMGB1 mediates lipopolysaccharide-induced inflammation via interacting with GPX4 in colon cancer cells. Cancer Cell Int. 2020, 20. [Google Scholar] [CrossRef]
- Li, X.; Wu, J.; Zhang, X.; Chen, W. Glutathione reductase-mediated thiol oxidative stress suppresses metastasis of murine melanoma cells. Free Radic. Biol. Med. 2018, 129, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Dong, J.-L.; Chen, Y.-L.; Liu, Y.; Huang, S.-S.; Zhong, X.-L.; Cheng, Y.-H.; Wang, Z.-G. Nrf2 mediates the protective effects of homocysteine by increasing the levels of GSH content in HepG2 cells. Mol. Med. Rep. 2017, 16, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Ye, J.; You, J.; Shi, X.; Kang, W.; Wang, T. Role of the cystathionine β-synthase/H2S system in liver cancer cells and the inhibitory effect of quinolone-indolone conjugate QIC2 on the system. Oncol. Rep. 2017, 37, 3001–3009. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Song, L.; Wan, Q.; Wu, G.; Li, X.; Wang, Y.; Wang, J.; Liu, Z.; Zhong, X.; He, X.; et al. CMyc-mediated activation of serine biosynthesis pathway is critical for cancer progression under nutrient deprivation conditions. Cell Res. 2015, 25, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-L.; Lin, Z.-X.; Qin, Y.-S.; She, Y.-Q.; Chen, Y.; Chen, C.; Qiu, G.-D.; Zheng, J.-T.; Chen, Z.-L.; Zhang, S.-Y. Overexpression of long noncoding RNA LINC01419 in esophageal squamous cell carcinoma and its relation to the sensitivity to 5-fluorouracil by mediating GSTP1 methylation. Ther. Adv. Med. Oncol. 2019, 11. [Google Scholar] [CrossRef]
- Liu, C.J.; Yang, J.H.; Huang, F.Z.; Nie, W.P.; Liu, C.P.; Mao, X.H.; Yin, X.M.; Shen, X.B.; Peng, C.; Chen, M.F.; et al. Glutathione-s-transferase A 4 (GSTA4) suppresses tumor growth and metastasis of human hepatocellular carcinoma by targeting AKT pathway. Am. J. Transl. Res. 2017, 9, 301–315. [Google Scholar]
- Gaziano, J.M.; Sesso, H.D.; Christen, W.G.; Bubes, V.; Smith, J.P.; MacFadyen, J.; Schvartz, M.; Manson, J.E.; Glynn, R.J.; Buring, J.E. Multivitamins in the prevention of cancer in men: The Physicians’ Health Study II randomized controlled trial. JAMA 2012, 308, 1871–1880. [Google Scholar] [CrossRef]
- Qiao, Y.-L.; Dawsey, S.M.; Kamangar, F.; Fan, J.-H.; Abnet, C.C.; Sun, X.-D.; Johnson, L.L.; Gail, M.H.; Dong, Z.-W.; Yu, B.; et al. Total and cancer mortality after supplementation with vitamins and minerals: Follow-up of the Linxian General Population Nutrition Intervention Trial. J. Natl. Cancer Inst. 2009, 101, 507–518. [Google Scholar] [CrossRef]
- Myung, S.K.; Kim, Y.; Ju, W.; Choi, H.J.; Bae, W.K. Effects of antioxidant supplements on cancer prevention: Meta-analysis of randomized controlled trials. Ann. Oncol. 2009, 21, 166–179. [Google Scholar] [CrossRef]
- The Alpha-Tocopherol Beta Carotene Cancer Prevention Study Group. The effect of vitamin e and beta carotene on the incidence of lung cancer and other cancers in male smokers. N. Engl. J. Med. 1994, 330, 1029–1035. [Google Scholar] [CrossRef]
- Haider, C.; Ferk, F.; Bojaxhi, E.; Martano, G.; Stutz, H.; Bresgen, N.; Knasmüller, S.; Alija, A.; Eckl, P.M. Effects of β-carotene and its cleavage products in primary pneumocyte type II cells. Antioxidants 2017, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Albanes, D.; Heinonen, O.P.; Taylor, P.R.; Virtamo, J.; Edwards, B.K.; Rautalahti, M.; Hartman, A.M.; Palmgren, J.; Freedman, L.S.; Haapakoski, J.; et al. Alpha-Tocopherol and beta-carotene supplements and lung cancer incidence in the alpha-tocopherol, beta-carotene cancer prevention study: Effects of base-line characteristics and study compliance. J. Natl. Cancer Inst. 1996, 88, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- Virtamo, J.; Taylor, P.R.; Kontto, J.; Männistö, S.; Utriainen, M.; Weinstein, S.J.; Huttunen, J.; Albanes, D. Effects of α-tocopherol and β-carotene supplementation on cancer incidence and mortality: 18-Year postintervention follow-up of the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study. Int. J. Cancer 2014, 135, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Milkovic, L.; Siems, W.; Siems, R.; Zarkovic, N. Oxidative stress and antioxidants in carcinogenesis and integrative therapy of cancer. Curr. Pharm. Des. 2014, 20, 6529–6542. [Google Scholar] [CrossRef] [PubMed]
- Mut-Salud, N.; Álvarez, P.J.; Garrido, J.M.; Carrasco, E.; Aránega, A.; Rodríguez-Serrano, F. Antioxidant Intake and Antitumor Therapy: Toward Nutritional Recommendations for Optimal Results. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Jung, A.Y.; Cai, X.; Thoene, K.; Obi, N.; Jaskulski, S.; Behrens, S.; Flesch-Janys, D.; Chang-Claude, J. Antioxidant supplementation and breast cancer prognosis in postmenopausal women undergoing chemotherapy and radiation therapy. Am. J. Clin. Nutr. 2019, 109, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.-C.; Ji, J.-A.; Jiang, Z.-Y.; You, Q.-D. The Keap1-Nrf2-ARE Pathway as a Potential Preventive and Therapeutic Target: An Update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef]
- Alumkal, J.J.; Slottke, R.; Schwartzman, J.; Cherala, G.; Munar, M.; Graff, J.N.; Beer, T.M.; Ryan, C.W.; Koop, D.R.; Gibbs, A.; et al. A phase II study of sulforaphane-rich broccoli sprout extracts in men with recurrent prostate cancer. Investig. New Drugs 2015, 33, 480–489. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Fahey, J.W.; Kostov, R.V.; Kensler, T.W. KEAP1 and done? Targeting the NRF2 pathway with sulforaphane. Trends Food Sci. Technol. 2017, 69, 257–269. [Google Scholar] [CrossRef]
- Kujundžić, R.N.; Stepanić, V.; Milković, L.; Gašparović, A.Č.; Tomljanović, M.; Trošelj, K.G. Curcumin and its potential for systemic targeting of inflamm-aging and metabolic reprogramming in cancer. Int. J. Mol. Sci. 2019, 20, 1180. [Google Scholar] [CrossRef]
- Ferri, C.; West, K.; Otero, K.; Kim, Y.H. Effectiveness of Curcumin for Treating Cancer during Chemotherapy. Altern. Complementary Ther. 2018, 24, 13–18. [Google Scholar] [CrossRef]
- Robledinos-Antón, N.; Fernández-Ginés, R.; Manda, G.; Cuadrado, A. Activators and Inhibitors of NRF2: A Review of Their Potential for Clinical Development. Oxid. Med. Cell. Longev. 2019, 2019, 9372182. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, X.; Long, M.; Huang, Y.; Zhang, L.; Zhang, R.; Zheng, Y.; Liao, X.; Wang, Y.; Liao, Q.; et al. NRF2 activation by antioxidant antidiabetic agents accelerates tumor metastasis. Sci. Transl. Med. 2016, 8, 334ra51. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Hayes, J.D.; Henderson, C.J.; Wolf, C.R. Identification of retinoic acid as an inhibitor of transcription factor Nrf2 through activation of retinoic acid receptor alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 19589–19594. [Google Scholar] [CrossRef]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Retinoic Acids in the Treatment of Most Lethal Solid Cancers. J. Clin. Med. 2020, 9, 360. [Google Scholar] [CrossRef]
- Choi, E.J.; Jung, B.J.; Lee, S.H.; Yoo, H.S.; Shin, E.A.; Ko, H.J.; Chang, S.; Kim, S.Y.; Jeon, S.M. A clinical drug library screen identifies clobetasol propionate as an NRF2 inhibitor with potential therapeutic efficacy in KEAP1 mutant lung cancer. Oncogene 2017, 36, 5285–5295. [Google Scholar] [CrossRef]
- Gao, A.; Ke, Z.; Wang, J.; Yang, J.; Chen, S.-Y.; Chen, H. Apigenin sensitizes doxorubicin-resistant hepatocellular carcinoma BEL-7402/ADM cells to doxorubicin via inhibiting PI3K/Akt/Nrf2 pathway. Carcinogenesis 2013, 34, 1806–1814. [Google Scholar] [CrossRef]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef]
- Kirkpatrick, D.L.; Powis, G. Clinically Evaluated Cancer Drugs Inhibiting Redox Signaling. Antioxid. Redox Signal. 2017, 26, 262–273. [Google Scholar] [CrossRef]
- Onodera, T.; Momose, I.; Kawada, M. Potential Anticancer Activity of Auranofin. Chem. Pharm. Bull. (Tokyo) 2019, 67, 186–191. [Google Scholar] [CrossRef]
- Brenneisen, P.; Reichert, A.S. Nanotherapy and Reactive Oxygen Species (ROS) in Cancer: A Novel Perspective. Antioxidants 2018, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Dai, B.; Yoo, S.Y.; Bartholomeusz, G.; Graham, R.A.; Majidi, M.; Yan, S.; Meng, J.; Ji, L.; Coombes, K.; Minna, J.D.; et al. KEAP1-dependent synthetic lethality induced by AKT and TXNRD1 inhibitors in lung cancer. Cancer Res. 2013, 73, 5532–5543. [Google Scholar] [CrossRef] [PubMed]
- Romero, R.; Sayin, V.I.; Davidson, S.M.; Bauer, M.R.; Singh, S.X.; Leboeuf, S.E.; Karakousi, T.R.; Ellis, D.C.; Bhutkar, A.; Sánchez-Rivera, F.J.; et al. Keap1 loss promotes Kras-driven lung cancer and results in dependence on glutaminolysis. Nat. Med. 2017, 23, 1362–1368. [Google Scholar] [CrossRef] [PubMed]
- Best, S.A.; Ding, S.; Kersbergen, A.; Dong, X.; Song, J.Y.; Xie, Y.; Reljic, B.; Li, K.; Vince, J.E.; Rathi, V.; et al. Distinct initiating events underpin the immune and metabolic heterogeneity of KRAS-mutant lung adenocarcinoma. Nat. Commun. 2019, 10, 4190. [Google Scholar] [CrossRef]
- Xu, X.; Yang, Y.; Liu, X.; Cao, N.; Zhang, P.; Zhao, S.; Chen, D.; Li, L.; He, Y.; Dong, X.; et al. NFE2L2/KEAP1 Mutations Correlate with Higher Tumor Mutational Burden Value/PD-L1 Expression and Potentiate Improved Clinical Outcome with Immunotherapy. Oncologist 2020, 25, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonenn, A.L.; Levonen, A. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef]
- Leinonen, H.M.; Ruotsalainen, A.K.; Määttä, A.M.; Laitinen, H.M.; Kuosmanen, S.M.; Kansanen, E.; Pikkarainen, J.T.; Lappalainen, J.P.; Samaranayake, H.; Lesch, H.P.; et al. Oxidative stress-regulated lentiviral TK/GCV gene therapy for lung cancer treatment. Cancer Res. 2012, 72, 6227–6235. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).