Sigma-1 Receptor Engages an Anti-Inflammatory and Antioxidant Feedback Loop Mediated by Peroxiredoxin in Experimental Colitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Drug Preparations
2.2. Experimental Animals for the Induction of Colitis
2.3. Damage Score and Measurement of the Lesions
2.4. Determination of 3-NT, PRDX1, -2, -4, and -6 Levels in the Colon by ELISA
2.5. Determination of SOD Activity in the Colon
2.6. Determination of the Total GSH in the Colon
2.7. Protein Determination
2.8. Data Representation and Statistical Analysis
3. Results
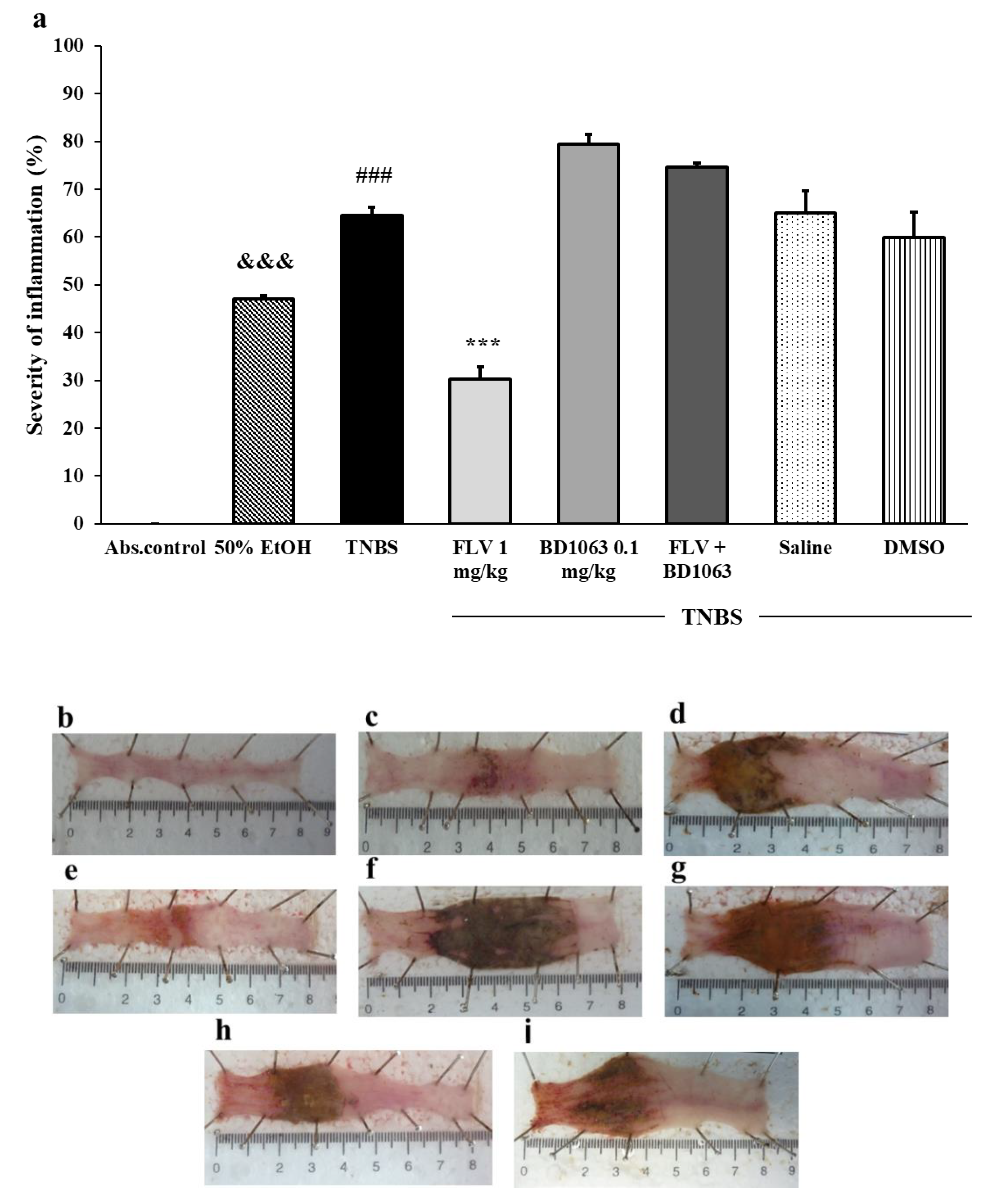
3.1. Severity of Inflammation Due to Sigma-1 Receptor Associated Treatments in TNBS-Induced Rat Colitis
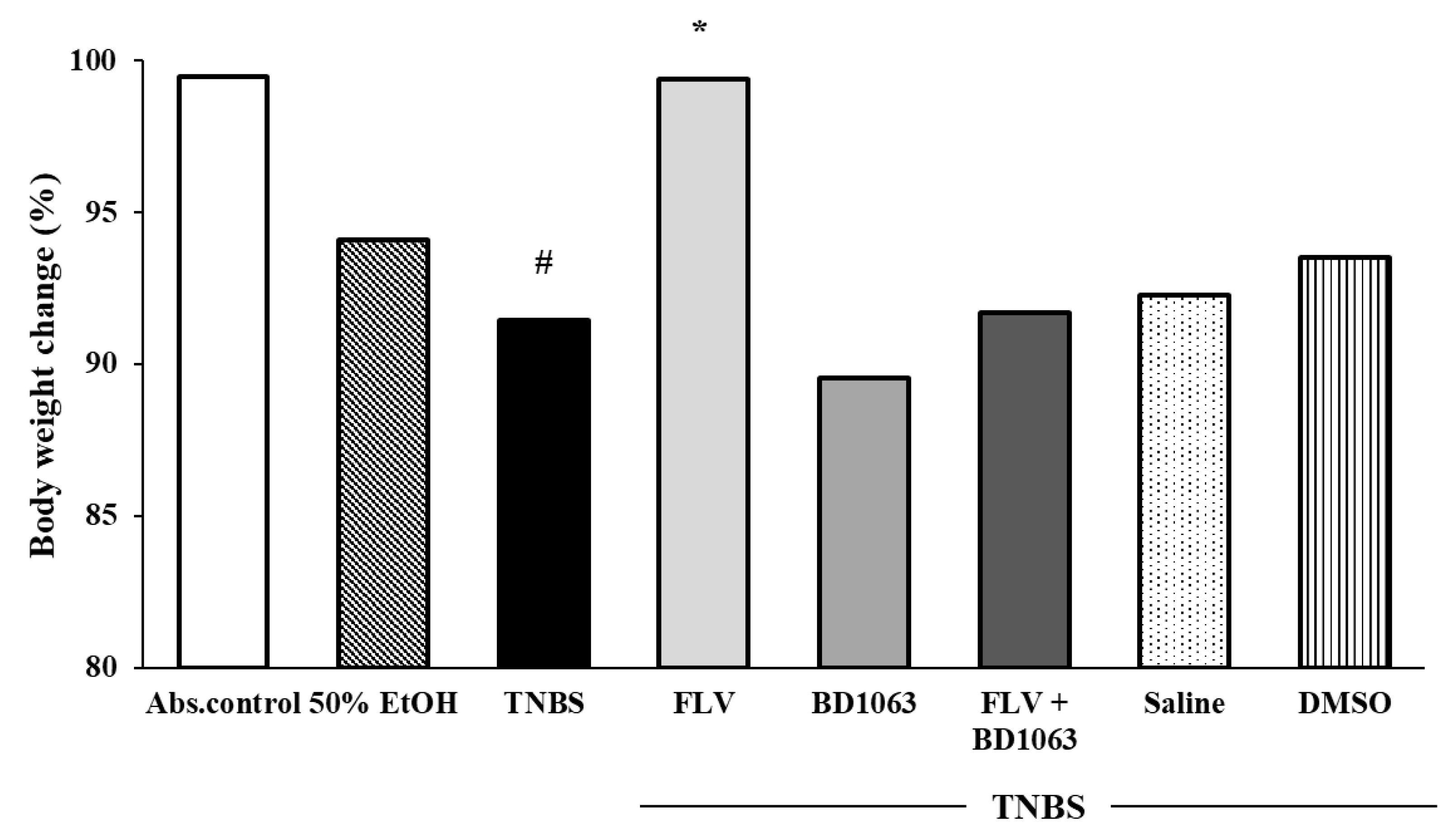
3.2. Body Weight Change of the Animals
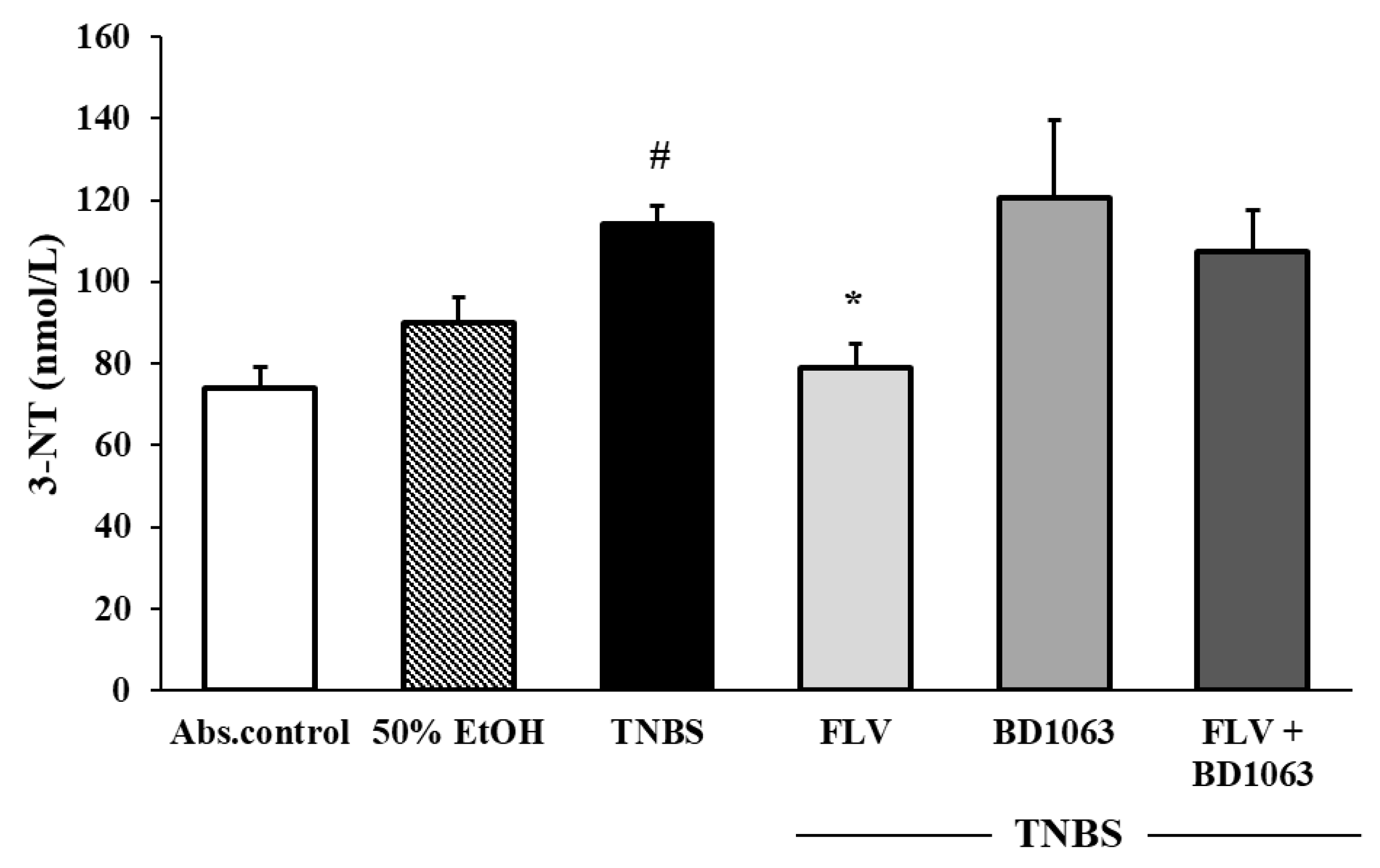
3.3. Sigma-1 Receptor Agonist FLV Decreased the Levels of 3-NT in TNBS Colitis
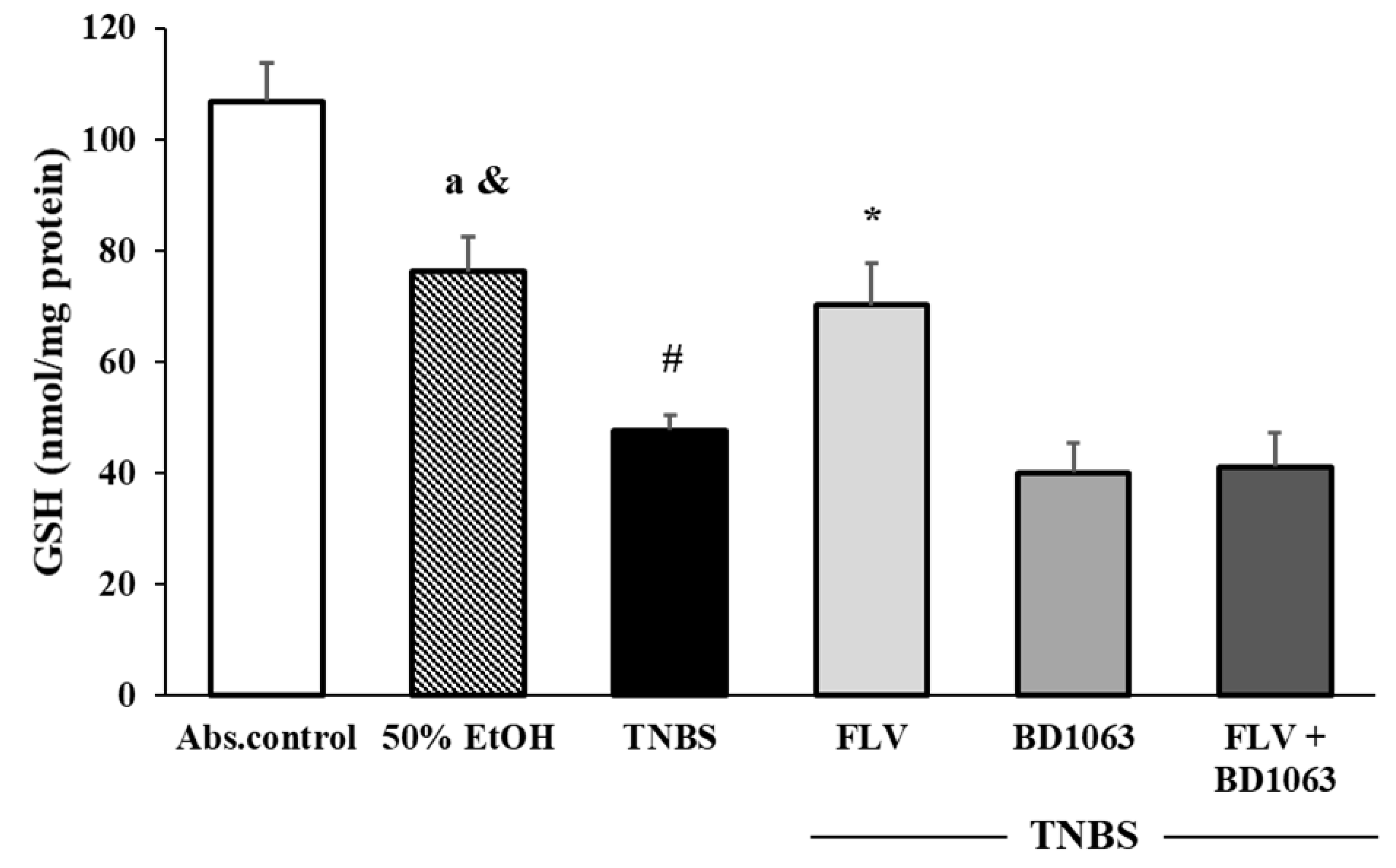
3.4. Effects of the Sigma-1 Receptor on the Levels of GSH in TNBS-Induced Colitis
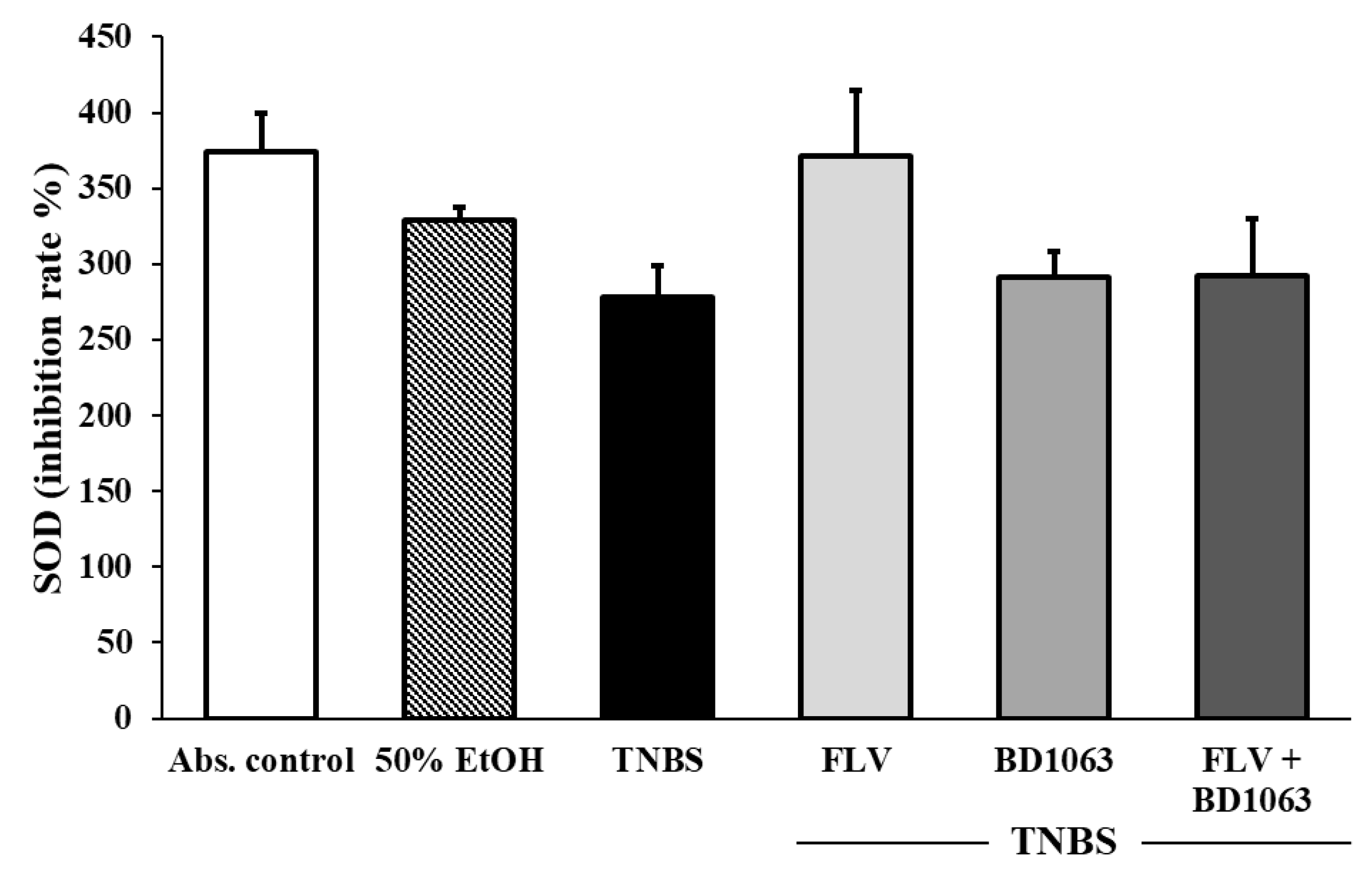
3.5. Determination of SOD Activity in Sigma-1 Receptor Ligand Treated Rat Colitis
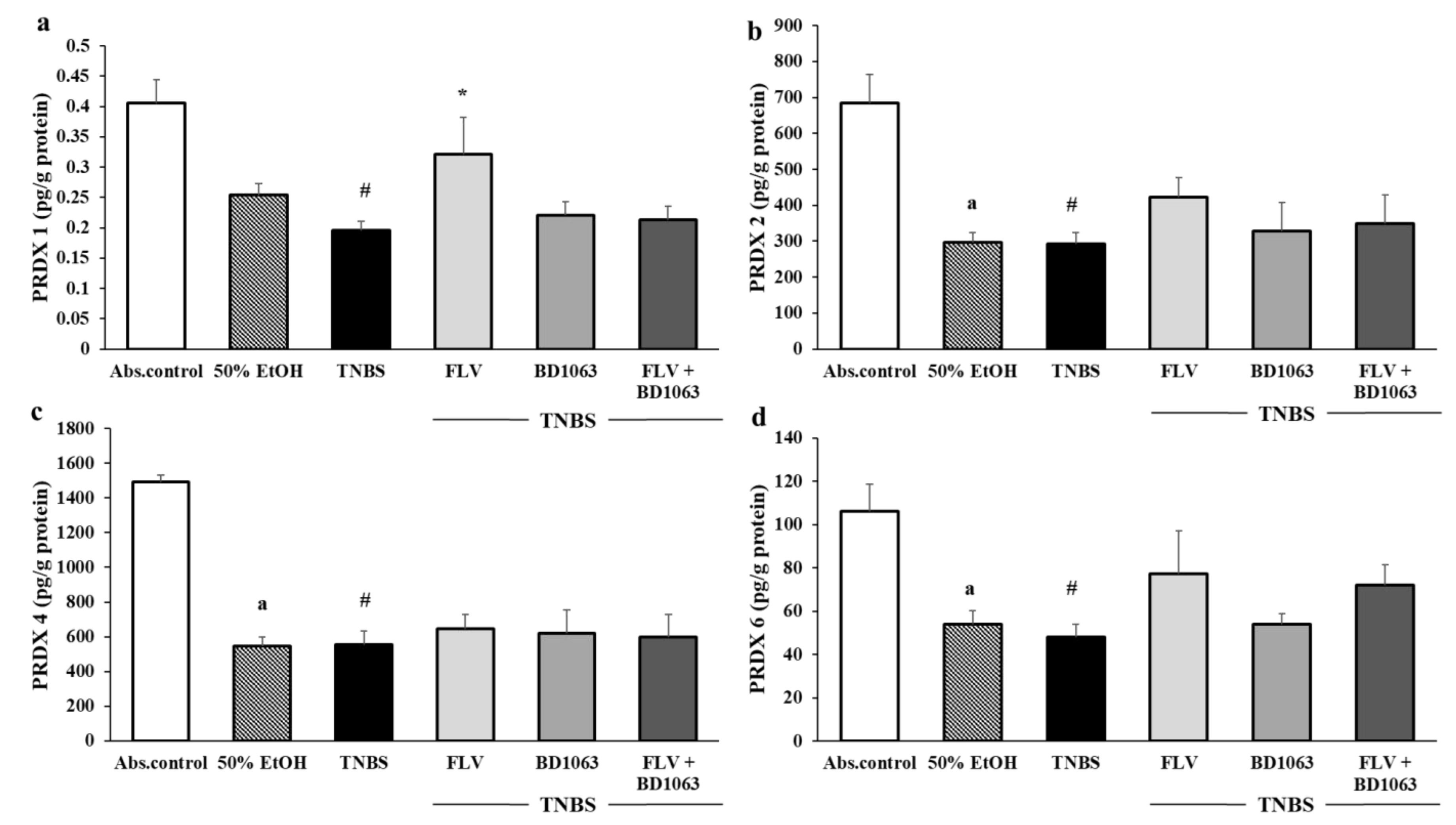
3.6. Changes in the Levels of PRDX1, -2, -4 and -6 Due to Sigma-1 Receptor Ligand Administration in Rat Colitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guan, Q. A Comprehensive review and update on the pathogenesis of inflammatory bowel disease. J. Immunol. Res. 2019, 2019, 7247238. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Ko, C.-O.; Ke, C.; Fa, F.; Rk, C. Faculty opinions recommendation of a state-of-the-art review of new and emerging therapies for the treatment of IBD. Inflamm. Bowel Dis. 2019, 25, 820–830. [Google Scholar] [CrossRef]
- Kim, D.H.; Cheon, J.H. Pathogenesis of inflammatory bowel disease and recent advances in biologic therapies. Immune Netw. 2017, 17, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Műzes, G. Changes of the cytokine profile in inflammatory bowel diseases. World J. Gastroenterol. 2012, 18, 5848–5861. [Google Scholar] [CrossRef]
- Zenewicz, L.A.; Antov, A.; Flavell, R.A. CD4 T-cell differentiation and inflammatory bowel disease. Trends Mol. Med. 2009, 15, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of oxidative stress in inflammatory bowel disease and potential antioxidant therapies. Oxidative Med. Cell. Longev. 2017, 2017, 4535194. [Google Scholar] [CrossRef]
- Dodda, D.; Chhajed, R.; Mishra, J.; Padhy, M. Targeting oxidative stress attenuates trinitrobenzene sulphonic acid induced inflammatory bowel disease like symptoms in rats: Role of quercetin. Indian J. Pharmacol. 2014, 46, 286–291. [Google Scholar] [CrossRef]
- Luceri, C.; Bigagli, E.; Agostiniani, S.; Giudici, F.; Zambonin, D.; Scaringi, S.; Ficari, F.; Lodovici, M.; Malentacchi, C. Analysis of oxidative stress-related markers in Crohn’s Disease patients at surgery and correlations with clinical findings. Antioxidants 2019, 8, 378. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxidative Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Ahsan, H. 3-Nitrotyrosine: A biomarker of nitrogen free radical species modified proteins in systemic autoimmunogenic conditions. Hum. Immunol. 2013, 74, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- De Barboza, G.D.; Guizzardi, S.; Moine, L.; De Talamoni, N.T. Oxidative stress, antioxidants and intestinal calcium absorption. World J. Gastroenterol. 2017, 23, 2841–2853. [Google Scholar] [CrossRef]
- Dickinson, D.A.; Forman, H.J. Cellular glutathione and thiols metabolism. Biochem. Pharmacol. 2002, 64, 1019–1026. [Google Scholar] [CrossRef]
- Buettner, G.R. Superoxide dismutase in redox biology: The roles of superoxide and hydrogen peroxide. Anti-Cancer Agents Med. Chem. 2011, 11, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G. Overview on peroxiredoxin. Mol. Cells 2016, 39, 1–5. [Google Scholar] [CrossRef]
- Horie, K.; Mikami, T.; Yoshida, T.; Sato, Y.; Okayasu, I. Peroxiredoxin 1 expression in active ulcerative colitis mucosa identified by proteome analysis and involvement of thioredoxin based on immunohistochemistry. Oncol. Lett. 2017, 15, 2364–2372. [Google Scholar] [CrossRef] [PubMed]
- Senhaji, N.; Zaid, Y.; El Khalfi, B.; Fahimi, M.; Martin, J.; Badre, W.; Nadifi, S.; Soukri, A. Peroxiredoxin-2 up-regulation in inflammatory bowel disease: Friend or foe? J. Gastroenterol. Hepatol. 2017, 32, 1212–1220. [Google Scholar] [CrossRef]
- Pfeuffer, S.; Trost, N.; Aden, K.; Schreiber, S.; Lipinski, S.; Rosenstiel, P. PWE-110 a role for peroxiredoxin-4 in a murine colitis model of intestinal inflammation. Gut 2014, 63, A172.2–A173. [Google Scholar] [CrossRef][Green Version]
- Melhem, H.; Spalinger, M.R.; Cosin-Roger, J.; Atrott, K.; Lang, S.; Wojtal, K.A.; Vavricka, S.R.; Rogler, G.; Frey-Wagner, I. Prdx6 deficiency ameliorates DSS colitis: Relevance of compensatory antioxidant mechanisms. J. Crohns Colitis 2017, 11, 871–884. [Google Scholar] [CrossRef]
- Martin, W.R.; Eades, C.G.; Thompson, J.A.; Huppler, R.E.; Gilbert, P.E. The effects of morphine- and nalorphine- like drugs in the nondependent and morphine-dependent chronic spinal dog. J. Pharmacol. Exp. Ther. 1976, 197, 517–532. [Google Scholar]
- Rousseaux, C.G.; Greene, S.F. Sigma receptors [σRs]: Biology in normal and diseased states. J. Recept. Signal Transduct. 2015, 36, 1–62. [Google Scholar] [CrossRef]
- Guitart, X.; Codony, X.; Monroy, X. Sigma receptors: Biology and therapeutic potential. Psychopharmacology 2004, 174, 301–319. [Google Scholar] [CrossRef]
- Schmidt, H.R.; Betz, R.M.; Dror, R.O.; Kruse, A.C. Structural basis for σ1 receptor ligand recognition. Nat. Struct. Mol. Biol. 2018, 25, 333765. [Google Scholar] [CrossRef]
- Omi, T.; Tanimukai, H.; Kanayama, D.; Sakagami, Y.; Tagami, S.; Okochi, M.; Morihara, T.; Sato, M.; Yanagida, K.; Kitasyoji, A.; et al. Fluvoxamine alleviates ER stress via induction of Sigma-1 receptor. Cell Death Dis. 2014, 5, e1332. [Google Scholar] [CrossRef]
- Urfer, R.; Moebius, H.J.; Skoloudik, D.; Santamarina, E.; Sato, W.; Mita, S.; Muir, K.W. Phase II trial of the sigma-1 receptor agonist cutamesine (SA4503) for recovery enhancement after acute ischemic stroke. Stroke 2014, 45, 3304–3310. [Google Scholar] [CrossRef]
- Pal, A.; Fontanilla, D.; Gopalakrishnan, A.; Chae, Y.-K.; Markley, J.L.; Ruoho, A.E. The sigma-1 receptor protects against cellular oxidative stress and activates antioxidant response elements. Eur. J. Pharmacol. 2012, 682, 12–20. [Google Scholar] [CrossRef]
- Rosen, D.A.; Seki, S.M.; Fernández-Castañeda, A.; Beiter, R.M.; Eccles, J.D.; Woodfolk, J.A.; Gaultier, A. Modulation of the sigma-1 receptor–IRE1 pathway is beneficial in preclinical models of inflammation and sepsis. Sci. Transl. Med. 2019, 11, eaau5266. [Google Scholar] [CrossRef]
- Almási, N.; Török, S.; Dvorácskó, S.; Tömböly, C.; Csonka, Á.; Baráth, Z.; Murlasits, Z.; Valkusz, Z.; Pósa, A.; Varga, C.; et al. Lessons on the sigma-1 receptor in TNBS-induced rat colitis: Modulation of the UCHL-1, IL-6 pathway. Int. J. Mol. Sci. 2020, 21, 4046. [Google Scholar] [CrossRef]
- Morris, G.P.; Beck, P.L.; Herridge, M.S.; Depew, W.T.; Szewczuk, M.R.; Wallace, J.L. Hapten-induced model of chronic inflammation and ulceration in the rat colon. Gastroenterology 1989, 96, 795–803. [Google Scholar] [CrossRef]
- Goguadze, N.; Zhuravliova, E.; Morin, D.; Mikeladze, D.; Maurice, T. Sigma-1 receptor agonists induce oxidative stress in mitochondria and enhance complex I activity in physiological condition but protect against pathological oxidative stress. Neurotox. Res. 2019, 35, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Su, T.-C.; Lin, S.-H.; Lee, P.-T.; Yeh, S.-H.; Hsieh, T.-H.; Chou, S.-Y.; Su, T.-P.; Hung, J.-J.; Chang, W.-C.; Lee, Y.-C.; et al. The sigma-1 receptor-zinc finger protein 179 pathway protects against hydrogen peroxide-induced cell injury. Neuropharmacology 2016, 105, 1–9. [Google Scholar] [CrossRef]
- Jeppesen, U.; Gram, L.F.; Vistisen, K.; Loft, S.; Poulsen, H.E.; Nielsen, F. Dose-dependent inhibition of CYP1A2, CYP2C19 and CYP2D6 by citalopram, fluoxetine, fluvoxamine and paroxetine. Eur. J. Clin. Pharmacol. 1996, 51, 73–78. [Google Scholar] [CrossRef]
- Antoniou, E.; Margonis, G.A.; Angelou, A.; Pikouli, A.; Argiri, P.; Karavokyros, I.; Papalois, A.; Pikoulis, E. The TNBS-induced colitis animal model: An overview. Ann. Med. Surg. 2016, 11, 9–15. [Google Scholar] [CrossRef]
- Wieczorek, I.; Schulz, C.; Jarry, H.; Lehnert, H. The effects of the selective serotonin reuptake-inhibitor fluvoxamine on body weight in Zucker rats are mediated by corticotropin-releasing hormone. Int. J. Obes. 2001, 25, 1566–1569. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dursun, H.; Bilici, M.; Albayrak, F.; Ozturk, C.; Saglam, M.B.; Alp, H.H.; Suleyman, H. Antiulcer activity of fluvoxamine in rats and its effect on oxidant and antioxidant parameters in stomach tissue. BMC Gastroenterol. 2009, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Weng, T.-Y.; Hung, D.T.; Su, T.-P.; Tsai, S.-Y.A. Loss of sigma-1 receptor chaperone promotes astrocytosis and enhances the Nrf2 antioxidant defense. Oxidative Med. Cell. Longev. 2017, 2017, 4582135. [Google Scholar] [CrossRef]
- Elsaed, W.M.; Alahmadi, A.M.; Al-Ahmadi, B.T.; Taha, J.A.; Tarabishi, R.M. Gastroprotective and antioxidant effects of fluvoxamine on stress-induced peptic ulcer in rats. J. Taibah Univ. Med. Sci. 2018, 13, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-J.; Leung, F.P.; Hsiao, W.W.; Tan, S.; Li, S.; Xu, H.-X.; Sung, J.J.; Bian, Z.-X. Proteome profiling of spinal cord and dorsal root ganglia in rats with trinitrobenzene sulfonic acid-induced colitis. World J. Gastroenterol. 2012, 18, 2914–2928. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.-Y.; Shih, T.-C.; Yeh, C.-Y.; Lin, C.-J.; Chou, Y.-Y.; Lee, Y.-S. Comparative proteomic studies on the pathogenesis of human ulcerative colitis. Proteomics 2006, 6, 5322–5331. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almási, N.; Török, S.; Valkusz, Z.; Tajti, M.; Csonka, Á.; Murlasits, Z.; Pósa, A.; Varga, C.; Kupai, K. Sigma-1 Receptor Engages an Anti-Inflammatory and Antioxidant Feedback Loop Mediated by Peroxiredoxin in Experimental Colitis. Antioxidants 2020, 9, 1081. https://doi.org/10.3390/antiox9111081
Almási N, Török S, Valkusz Z, Tajti M, Csonka Á, Murlasits Z, Pósa A, Varga C, Kupai K. Sigma-1 Receptor Engages an Anti-Inflammatory and Antioxidant Feedback Loop Mediated by Peroxiredoxin in Experimental Colitis. Antioxidants. 2020; 9(11):1081. https://doi.org/10.3390/antiox9111081
Chicago/Turabian StyleAlmási, Nikoletta, Szilvia Török, Zsuzsanna Valkusz, Máté Tajti, Ákos Csonka, Zsolt Murlasits, Anikó Pósa, Csaba Varga, and Krisztina Kupai. 2020. "Sigma-1 Receptor Engages an Anti-Inflammatory and Antioxidant Feedback Loop Mediated by Peroxiredoxin in Experimental Colitis" Antioxidants 9, no. 11: 1081. https://doi.org/10.3390/antiox9111081
APA StyleAlmási, N., Török, S., Valkusz, Z., Tajti, M., Csonka, Á., Murlasits, Z., Pósa, A., Varga, C., & Kupai, K. (2020). Sigma-1 Receptor Engages an Anti-Inflammatory and Antioxidant Feedback Loop Mediated by Peroxiredoxin in Experimental Colitis. Antioxidants, 9(11), 1081. https://doi.org/10.3390/antiox9111081