Relationship between the Length of Sperm Tail Mitochondrial Sheath and Fertility Traits in Boars Used for Artificial Insemination

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. General Approach
2.2. Animals and Fertility Data
2.3. Semen Collection and Processing
2.4. Mitochondrial Sheath Labelling and Measurement
2.5. Image-based Flow Cytometry (IBFC) Sample Acquisiation
2.6. IBFC Data Analysis
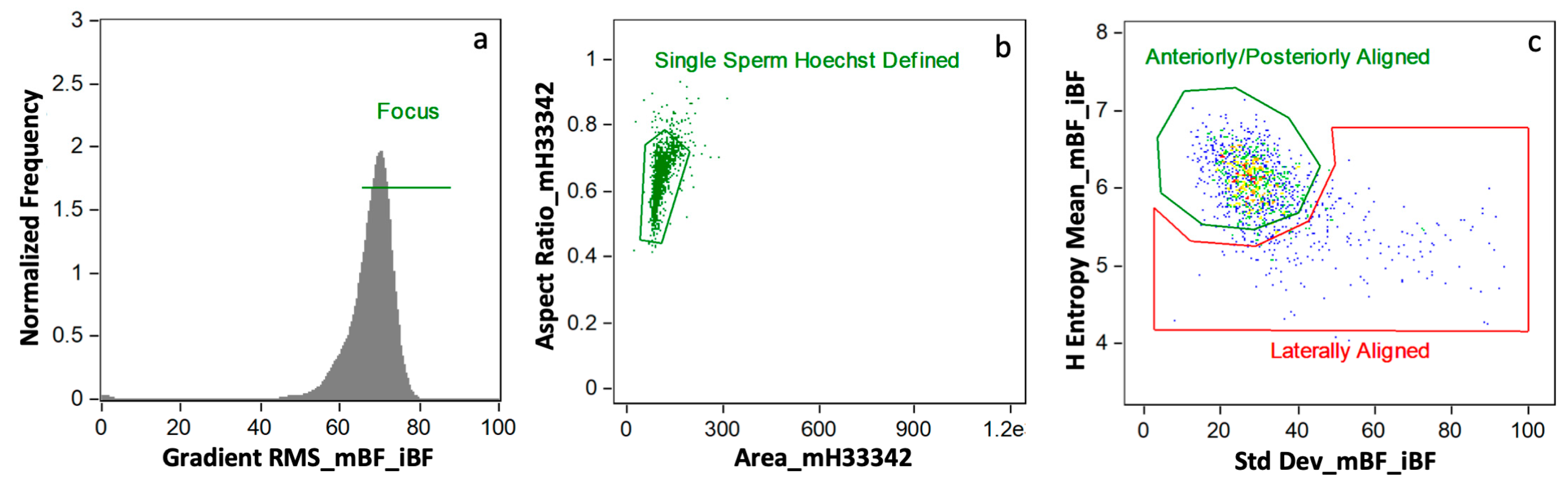
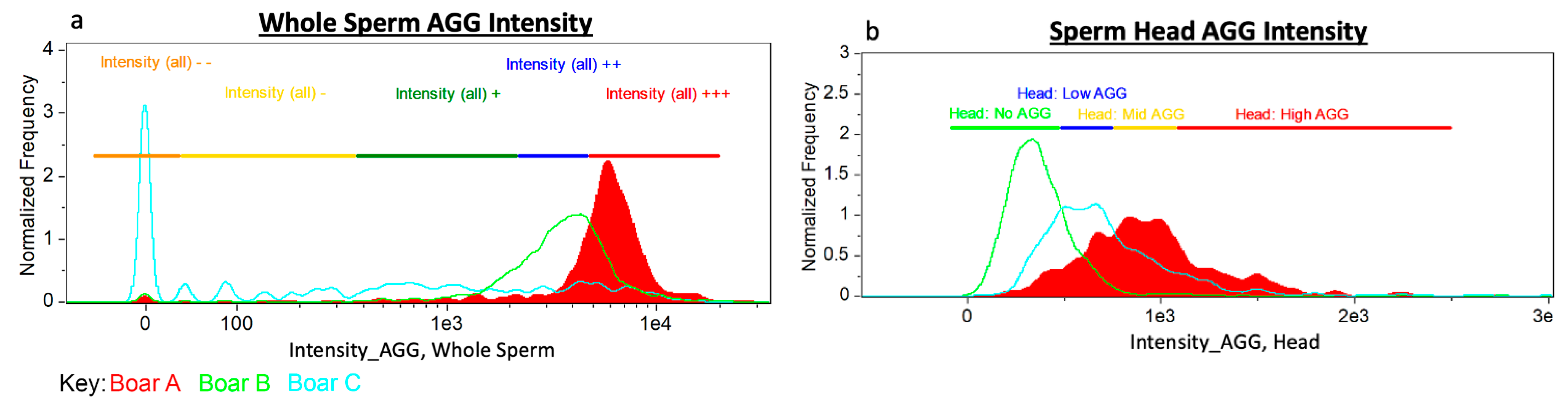
2.6.1. Masks
2.6.2. IBFC Definitions
2.6.3. Gating
2.7. Statistics
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fisher, H.S.; Jacobs-Palmer, E.; Lassance, J.-M.; Hoekstra, H.E. The genetic basis and fitness consequences of sperm midpiece size in deer mice. Nat. Commun. 2016, 7, 13652. [Google Scholar] [CrossRef] [PubMed]
- Firman, R.C.; Simmons, L.W. Sperm midpiece length predicts sperm swimming velocity in house mice. Biol. Lett. 2010, 6, 513–516. [Google Scholar] [CrossRef] [PubMed]
- Vladić, T.V.; Afzelius, B.A.; Bronnikov, G.E. Sperm quality as reflected through morphology in salmon alternative life histories. Biol. Reprod. 2002, 66, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Lüpold, S.; Calhim, S.; Immler, S.; Birkhead, T.R. Sperm morphology and sperm velocity in passerine birds. Proc. R. Soc. B Biol. Sci. 2008, 276, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Gomendio, M.; Roldan, E.R. Sperm competition influences sperm size in mammals. Proc. R. Soc. B Boil. Sci. 1991, 243, 181–185. [Google Scholar] [CrossRef]
- Lush, J.L. The possibility of sex control by artificial insemination with centrifuged spermatozoa. J. Agric. Res. 1925, 30, 893–913. [Google Scholar]
- Parra, C.M.; Sansegundo, M.; Ruiz, S.; García-Vázquez, F.A.; Gadea, J.; Romar, R.; Coy, P. Sperm treatment affects capacitation parameters and penetration ability of ejaculated and epididymal boar spermatozoa. Theriogenology 2010, 74, 1327–1340. [Google Scholar] [CrossRef]
- Garner, D.L.; Johnson, L.A.; Yue, S.T.; Roth, B.L.; Haugland, R.P. Dual DNA staining assessment of bovine sperm viability using SYBR-14 and propidium iodide. J. Androl. 1994, 15, 620–629. [Google Scholar]
- Kerns, K.; Zigo, M.; Drobnis, E.Z.; Sutovsky, M.; Sutovsky, P. Zinc ion flux during mammalian sperm capacitation. Nat. Commun. 2018, 9, 2061. [Google Scholar] [CrossRef]
- Garner, D.L.; Thomas, C.A.; Joerg, H.W.; DeJarnette, J.M.; Marshall, C.E. Fluorometric Assessments of Mitochondrial Function and Viability in Cryopreserved Bovine Spermatozoa1. Biol. Reprod. 1997, 57, 1401–1406. [Google Scholar] [CrossRef]
- Sellem, E.; Broekhuijse, M.; Chevrier, L.; Camugli, S.; Schmitt, E.; Schibler, L.; Koenen, E. Use of combinations of in vitro quality assessments to predict fertility of bovine semen. Theriogenology 2015, 84, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Broekhuijse, M.L.W.J.; Šoštarić, E.; Feitsma, H.; Gadella, B.M. Relationship of flow cytometric sperm integrity assesments with boar fertility performance under optimized field conditions. J. Anim. Sci. 2012, 90, 4327–4336. [Google Scholar] [CrossRef] [PubMed]
- Evenson, D.; Thompson, L.; Jost, L. Flow cytometric evaluation of boar semen by the sperm chromatin structure assay as related to cryopreservation and fertility. Theriogenology 1994, 41, 637–651. [Google Scholar] [CrossRef]
- Althouse, G.; Kuster, C.; Clark, S.; Weisiger, R. Field investigations of bacterial contaminants and their effects on extended porcine semen. Theriogenology 2000, 53, 1167–1176. [Google Scholar] [CrossRef]
- Kennedy, C.E.; Krieger, K.B.; Sutovsky, M.; Xu, W.; Vargovič, P.; Didion, B.A.; Ellersieck, M.R.; Hennessy, M.E.; Verstegen, J.; Oko, R.; et al. Protein expression pattern of PAWP in bull spermatozoa is associated with sperm quality and fertility following artificial insemination. Mol. Reprod. Dev. 2014, 81, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.A.; Dziuk, P.J. Assessment of relative fertility of males (cockerels and boars) by competitive mating. Reproduction 1977, 49, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Dziuk, P.J. Factors that influence the proportion of offspring sired by a male following heterospermic insemination. Anim. Reprod. Sci. 1996, 43, 65–88. [Google Scholar] [CrossRef]
- Parker, G.A. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- Gage, M.J.G. Mammalian sperm morphometry. Proc. R. Soc. B Biol. Sci. 1998, 265, 97–103. [Google Scholar] [CrossRef]
- Lüpold, S.; Fitzpatrick, J.L. Sperm number trumps sperm size in mammalian ejaculate evolution. Proc. R. Soc. B Boil. Sci. 2015, 282. [Google Scholar] [CrossRef]
- Sutovsky, P.; Navara, C.S.; Schatten, G. Fate of the sperm mitochondria, and the incorporation, conversion, and disassembly of the sperm tail structures during bovine fertilization. Biol. Reprod. 1996, 55, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P.; McCauley, T.C.; Day, B.N. Early Degradation of Paternal Mitochondria in Domestic Pig (Sus scrofa) Is Prevented by Selective Proteasomal Inhibitors Lactacystin and MG1321. Biol. Reprod. 2003, 68, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- Zuidema, D.; Sutovsky, P. The domestic pig as a model for the study of mitochondrial inheritance. Cell Tissue Res. 2020, 380, 263–271. [Google Scholar] [CrossRef]
- Sutovsky, P.; Moreno, R.D.; Ramalho-Santos, J.; Dominko, T.; Simerly, C.; Schatten, G. Ubiquitin tag for sperm mitochondria. Nature 1999, 402, 371–372. [Google Scholar] [CrossRef]
- Sutovsky, P.; Moreno, R.D.; Ramalho-Santos, J.; Dominko, T.; Simerly, C.; Schatten, G. Ubiquitinated sperm mitochondria, selective proteolysis, and the regulation of mitochondrial inheritance in mammalian embryos. Biol. Reprod. 2000, 63, 582–590. [Google Scholar] [CrossRef]
- Song, W.H.; Yi, Y.J.; Sutovsky, M.; Meyers, S.; Sutovsky, P. Autophagy and ubiquitin-proteasome system contribute to sperm mitophagy after mammalian fertilization. Proc. Natl. Acad. Sci. USA 2016. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Data | |
| Term | Definition |
| Mean Number of Matings/Sire | The average number of sows a boar was used to breed |
| Terminal Index (TI) | The genetic value assigned to a boar based on specific traits (internal Maschhoffs composite value compromised of wean-to-market traits, e.g., % lean, backfat depth, etc.) |
| Mean Conception Rate (CR) | The number of sows confirmed pregnant at 28 day pregnancy check that were bred |
| Average Total Number Born/Litter (TNB) | The number of pigs born (includes live, stillborn and mummified) per litter farrowed |
| Reproduction Index (RI) | CR × TNB × 100 |
| Semen Data | |
| Term | Definition |
| % Normal Morphology | Percent of spermatozoa with normal morphology |
| % Mero-negative | Percent of spermatozoa not stained with mero cyanine 540. Positive cells indicate changes in phospholipid arrangement and membrane fluidity, an early indicator of spermatozoa stress |
| % Viable & Intact Acrosomes | Percent viable spermatozoa with intact acrosomes not stained with propidium iodide or lectin PNA-FITC |
| % Depolarized mitochondria | Percent of spermatozoa with depolarized mitochondria (low membrane potential) as determined by JC-1 staining |
| Oxidation | Measures the intracellular level of reactive oxygen species (ROS) or free radicals in spermatozoa, calculated as the Basal:Induced ratio |
| Comp DNA | Measures the ability of the sperm chromatin to maintain structural integrity after undergoing acid stress |
| Parameter | All (n = 57) | 1st TERTILE (Low/Shortest) | 2nd TERTILE (Middle) | 3rd TERTILE (High/Longest) |
|---|---|---|---|---|
| Mean MS length ± SD (µm) | 7.26 ± 0.2 | 7.12 ± 0.09 | 7.26 ± 0.03 | 7.4 ± 0.1 |
| Mean MS length range (µm) | 1.01 ± 0.4 | 1.04 ± 0.4 | 0.85 ± 0.29 | 1.12 ± 0.46 |
| Mean shortest MS (µm) | 6.73 ± 0.4 | 6.55 ± 0.29 | 6.79 ± 0.29 | 6.86 ± 0.25 |
| Mean longest MS (µm) | 7.74 ± 0.33 | 7.59 ± 0.26 | 7.64 ± 0.09 | 7.97 ± 0.43 |
| Mean number of matings/sire | 113.27 ± 105.46 | 104.79 ± 93.15 | 91.39 ± 59.21 | 142.21 ± 141.21 |
| Terminal index | 129.23 ± 6.83 | 129.64 ± 7.11 | 128.47 ± 5.88 | 129.53 ± 7.65 |
| Mean conception rate (% CR) | 74.55 ± 1.0 | 74.79 ± 9.96 | 71.0 ± 13.3 | 77.73 ± 6.02 p23 * |
| Average total number born/liter | 13.72 ± 0.74 | 13.92 ± 0.71 | 13.61 ± 0.79 | 13.62 ± 0.7 |
| Reproduction index | 1024.18 ± 161.9 | 1041 ± 149.56 | 969.61 ± 205.2 | 1059.47 ± 109.11 |
| % Normal morphology | 71.39 ± 13.22 | 72.18 ± 12.11 | 71.03 ± 12.65 | 71.44 ± 15.58 |
| % Mero-negative | 85.89 ± 4.36 | 87.18 ± 2.85 | 84.62 ± 4.86 | 85.6 ± 4.92 |
| % Depolarized mitochondria | 18.12 ± 5.61 | 18.24 ± 2.85 | 17.61 ± 6.02 | 18.01 ± 4.41 |
| % Viable & intact acrosomes | 72.94 ± 13.92 | 75.27 ± 10.86 | 66.34 ± 20.17 | 76.72 ± 5.41 p23 * |
| Oxidation (Basal:Induced ratio) | 0.44 ± 0.25 | 0.54 ± 0.3 | 0.35 ± 0.22 p12 * | 0.44 ± 0.17 |
| Comp DNA | 5.37 ± 11.97 | 2.29 ± 0.94 p12 x | 11.2 ± 19.55 | 2.46 ± 0.96 p23 x |
| Number of matings per tertile | 6535 | 1991 | 1842 | 2702 |
| PARAMETER | ALL (n = 45) | 1st TERTILE (Low) | 2nd TERTILE (Middle) | 3rd TERTILE (High) | Grouping |
|---|---|---|---|---|---|
| Sperm head/nucleus circularity, Mean (FlowSight units) | 4.91 ± 0.38 | 5.11 ±0.47 | 4.71 ± 0.23 p12 ** | 4.90 ± 0.311 | Tertile: Reproduction Index 1 |
| Average total number born/litter | 13.77 ±0.74 | 14.31 ± 0.37 p12 *** | 13.5 ± 0.73 | 13.85 ± 0.79 | Tertile: MS length 2 |
| Average total number born/litter | 13.77 ± 0.74 | 14.05 ± 0.51 | 13.82 ± 0.70 p23 ** | 12.85 ± 0.1 p13 ** | Tertile: Head AGG Bright Detail Intensity R3 3 |
| % Viable with intact acrosome | 76.50 ± 8.45 | 78.88 ± 4.70 | 77.28 ± 7.17 p23 ** | 66.42 ± 15.15 p13 ** | Tertile: Head AGG Bright Detail Intensity R3 4 |
| % Intensity AGG +++ | 8.37 ± 7.68 | 15.99 ± 13.06 p13 ** | 5.94 ± 2.99 p23 ** | 6.5 ± 3.18 | Tertile: Terminal Index 5 |
| Oxidation (Basal:Induced ratio) | 0.47 ± 0.25 | 0.30 ± 0.11 | 0.50 ± 0.22 p12 ** | 0.65 ± 0.31 p13 ** | Tertile: Terminal Index 5 |
| Oxidation (Basal:Induced ratio) | 0.47 ± 0.25 | 0.53 ± 0.28 p12 * | N/A | 0.39 ± 0.18 | Quantile: MS length 6 |
| Side Scatter Symmetry_3, Mean | 12.15 ± 0.50 | 12.45 ± 0.45 p13 ** | 12.06 ± 0.42 p12 ** | 11.89 ± 0.55 | Tertile: Terminal Index 5 |
| H Correlation Mean Side Scatter_5, Std. Dev. | 0.199 ± 0.0096 | 0.205 ± 0.0123 p13 * | 0.197 ± 0.0064 p23 * | 0.193 ± 0.0064 | Tertile: Terminal Index 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerns, K.; Jankovitz, J.; Robinson, J.; Minton, A.; Kuster, C.; Sutovsky, P. Relationship between the Length of Sperm Tail Mitochondrial Sheath and Fertility Traits in Boars Used for Artificial Insemination. Antioxidants 2020, 9, 1033. https://doi.org/10.3390/antiox9111033
Kerns K, Jankovitz J, Robinson J, Minton A, Kuster C, Sutovsky P. Relationship between the Length of Sperm Tail Mitochondrial Sheath and Fertility Traits in Boars Used for Artificial Insemination. Antioxidants. 2020; 9(11):1033. https://doi.org/10.3390/antiox9111033
Chicago/Turabian StyleKerns, Karl, Jennifer Jankovitz, Julie Robinson, Amanda Minton, Chris Kuster, and Peter Sutovsky. 2020. "Relationship between the Length of Sperm Tail Mitochondrial Sheath and Fertility Traits in Boars Used for Artificial Insemination" Antioxidants 9, no. 11: 1033. https://doi.org/10.3390/antiox9111033
APA StyleKerns, K., Jankovitz, J., Robinson, J., Minton, A., Kuster, C., & Sutovsky, P. (2020). Relationship between the Length of Sperm Tail Mitochondrial Sheath and Fertility Traits in Boars Used for Artificial Insemination. Antioxidants, 9(11), 1033. https://doi.org/10.3390/antiox9111033