Antioxidant and Anti-Inflammatory Activities in Relation to the Flavonoids Composition of Pepper (Capsicum annuum L.)

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Determination of 1,1-Diphenyl-2-Picrylhydrazyl (DPPH) Radical Scavenging Activity
2.3. 2,2′-Azino-Bis(3-Ethylbenzothiazoline-6-Sulfonic Acid (ABTS·+) Radical Scavenging Activity
2.4. Cells and Culture
2.5. Cell Viability
2.6. Determination of Nitric Oxide (NO) Production
2.7. Determination of Interleukin (IL)-6 and Tumor Necrosis Factor (TNF)-α
2.8. Determination of Reactive Oxygen Species (ROS)
2.9. Qualitative and Quantitative Analysis of Flavonoids Using UPLC-DAD/QTOF-MS
2.10. Statistical Analysis
3. Results
3.1. Radical Scavenging Activity of the PL and PF Extracts
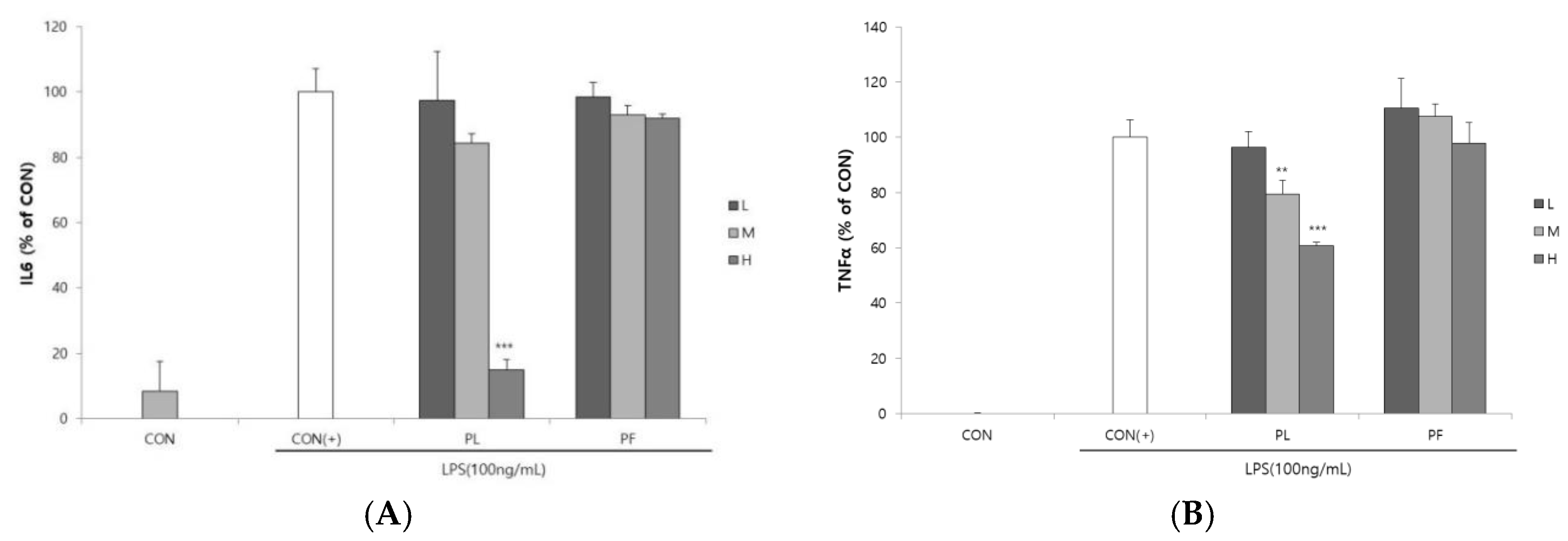
3.2. Effect of the PL and PF Extracts on Inflammation
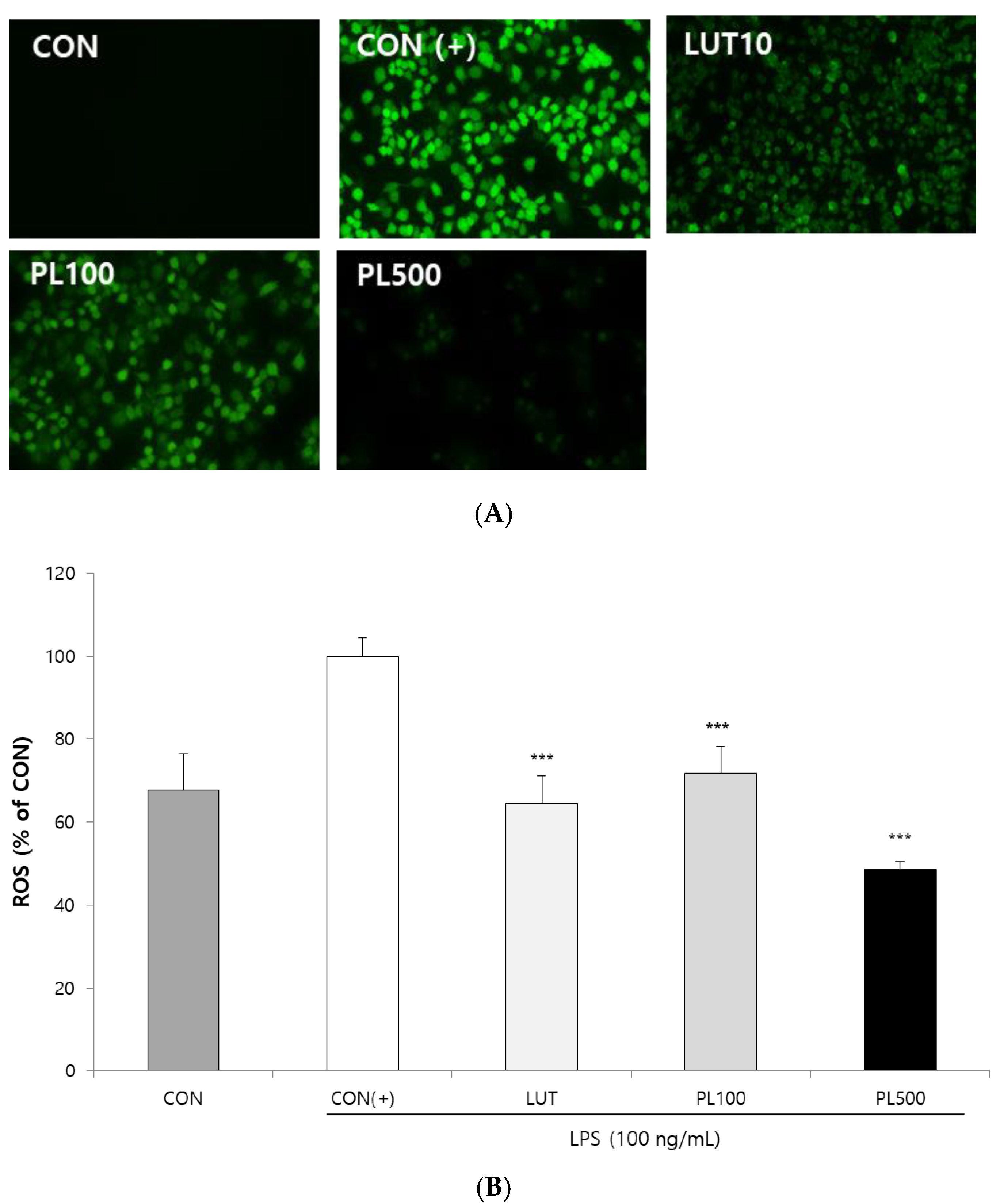
3.3. Effect of the PL and Luteolin on ROS Production
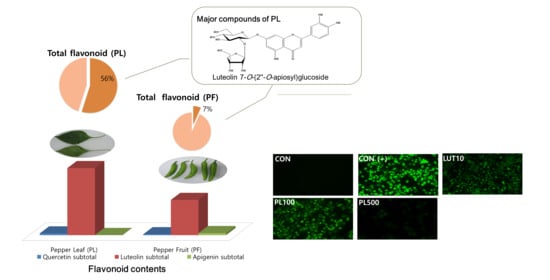
3.4. Flavonoid Content in the PL and PF Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bower, A.; Marquez, S.; de Mejia, E.G. The Health Benefits of Selected Culinary Herbs and Spices Found in the Traditional Mediterranean Diet. Crit. Rev. Food Sci. Nutr. 2016, 56, 2728–2746. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.A. Health Benefits of Culinary Herbs and Spices. J. AOAC Int. 2019, 102, 395–411. [Google Scholar] [CrossRef]
- Howard, L.R.; Talcott, S.T.; Brenes, C.H.; Villalon, B. Changes in Phytochemical and Antioxidant Activity of Selected Pepper Cultivars (Capsicum Species) As Influenced by Maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef]
- Marín, A.; Ferreres, F.; Tomás-Barberán, F.A.; Gil, M.I. Characterization and Quantitation of Antioxidant Constituents of Sweet Pepper (Capsicum annuum L.). J. Agric. Food Chem. 2004, 52, 3861–3869. [Google Scholar] [CrossRef] [PubMed]
- Materska, M.; Perucka, I. Antioxidant Activity of the Main Phenolic Compounds Isolated from Hot Pepper Fruit (Capsicum annuum L.). J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef]
- Srinivasan, K. Biological Activities of Red Pepper (Capsicum annuum) and Its Pungent Principle Capsaicin: A Review. Crit. Rev. Food Sci. Nutr. 2016, 56, 1488–1500. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.-R.; Kim, E.O.; Kang, K.; Oidovsambuu, S.; Jung, S.H.; Kim, B.S.; Nho, C.W.; Um, B.-H. Antioxidant Activity of Phenolics in Leaves of Three Red Pepper (Capsicum annuum) Cultivars. J. Agric. Food Chem. 2014, 62, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Miean, K.H.; Mohamed, S. Flavonoid (Myricetin, Quercetin, Kaempferol, Luteolin, and Apigenin) Content of Edible Tropical Plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef]
- González-Paramás, A.M.; Ayuda-Durán, B.; Martínez, S.; González-Manzano, S.; Santos-Buelga, C. The Mechanisms Behind the Biological Activity of Flavonoids. Curr. Med. Chem. 2019, 26, 6976–6990. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Knekt, P.; Kumpulainen, J.; Järvinen, R.; Rissanen, H.; Heliövaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-M.; Liu, Y.-J.; Huang, Y.; Yu, H.-J.; Yuan, S.; Tang, B.-W.; Wang, P.-G.; He, Q.-Q. Dietary total flavonoids intake and risk of mortality from all causes and cardiovascular disease in the general population: A systematic review and meta-analysis of cohort studies. Mol. Nutr. Food Res. 2017, 61, 1601003. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Yamamoto, Y.; He, P.; Klein, T.W.; Friedman, H. Endotoxin induced cytotoxicity of macrophages is due to apoptosis caused by nitric oxide production. J. Endotoxin Res. 1994, 1, 181–187. [Google Scholar] [CrossRef]
- Chen, L.; Kang, Y.-H. Anti-inflammatory and antioxidant activities of red pepper (Capsicum annuum L.) stalk extracts: Comparison of pericarp and placenta extracts. J. Funct. Foods 2013, 5, 1724–1731. [Google Scholar] [CrossRef]
- Sim, K.H.; Sil, H.Y. Antioxidant activities of red pepper (Capsicum annuum) pericarp and seed extracts. Int. J. Food Sci. Technol. 2008, 43, 1813–1823. [Google Scholar] [CrossRef]
- Choi, J. DPPH Radical Scavenging Activity and Isolation of Apigenins from the Leaves of Capsicum Annuum. Master‘s Thesis, Sunchon National University, Sunchon, Korea, 2006. [Google Scholar]
- Ku, K.M.; Kim, H.S.; Kim, B.S.; Kang, Y.H. Antioxidant activities and antioxidant constituents of pepper leaves from various cultivars and correlation between antioxidant activities and antioxidant constituents. J. Appl. Biol. Chem. 2009, 52, 70–76. [Google Scholar] [CrossRef][Green Version]
- Škrovánková, S.; Misurcova, L.; Machů, L. Antioxidant Activity and Protecting Health Effects of Common Medicinal Plants. Adv. Food Nutr. Res. 2012, 67, 75–139. [Google Scholar] [CrossRef]
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Dietary intake and blood concentrations of antioxidants and the risk of cardiovascular disease, total cancer, and all-cause mortality: A systematic review and dose-response meta-analysis of prospective studies. Am. J. Clin. Nutr. 2018, 108, 1069–1091. [Google Scholar] [CrossRef]
- Laflamme, N.; Rivest, S. Toll-like receptor 4: The missing link of the cerebral innate immune response triggered by circulating gram-negative bacterial cell wall components. FASEB J. 2001, 15, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Huang, Y.; So, Y.-J.; Hwang, J.R.; Yoo, K.-M.; Lee, K.-W.; Lee, Y.-J.; Hwang, I.K. Comparative studies on phytochemicals and bioactive activities in 24 new varieties of red pepper. Korean J. Food Sci. Technol. 2014, 46, 395–403. [Google Scholar] [CrossRef]
- Jeong, W.Y.; Jin, J.S.; Cho, Y.A.; Lee, J.H.; Park, S.; Jeong, S.W.; Kim, Y.H.; Lim, C.S.; El-Aty, A.A.; Kim, G.S. Determination of polyphenols in three Capsicum annuum L. (bell pepper) varieties using high-performance liquid chromatography-tandem mass spectrometry: Their contribution to overall antioxidant and anticancer activity. J. Sep. Sci. 2011, 34, 2967–2974. [Google Scholar] [CrossRef] [PubMed]
- Materska, M. Flavone C-glycosides from Capsicum annuum L.: Relationships between antioxidant activity and lipophilicity. Eur. Food Res. Technol. 2015, 240, 549–557. [Google Scholar] [CrossRef]
- Lee, Y.; Howard, L.; Villalon, B. Flavonoids and antioxidant activity of fresh pepper (Capsicum annuum) cultivars. J. Food Sci. 1995, 60, 473–476. [Google Scholar] [CrossRef]
- Galvez, J.; de la Cruz, J.P.; Zarzuelo, A.; de la Cuesta, F.S. Flavonoid inhibition of enzymic and nonenzymic lipid peroxidation in rat liver differs from its influence on the glutathione-related enzymes. Pharmacology 1995, 51, 127–133. [Google Scholar] [CrossRef]
- Xagorari, A.; Papapetropoulos, A.; Mauromatis, A.; Economou, M.; Fotsis, T.; Roussos, C. Luteolin inhibits an endotoxin-stimulated phosphorylation cascade and proinflammatory cytokine production in macrophages. J. Pharmacol. Exp. Ther. 2001, 296, 181–187. [Google Scholar]
- Ha, S.-H.; Kim, J.-B.; Park, J.-S.; Lee, S.-W.; Cho, K.-J. A comparison of the carotenoid accumulation in Capsicum varieties that show different ripening colours: Deletion of the capsanthin-capsorubin synthase gene is not a prerequisite for the formation of a yellow pepper. J. Exp. Bot. 2007, 58, 3135–3144. [Google Scholar] [CrossRef]
- Lee, J.J.; Crosby, K.M.; Pike, L.M.; Yoo, K.S.; Leskovar, D.I. Impact of genetic and environmental variation on development of flavonoids and carotenoids in pepper (Capsicum spp.). J. Sci. Hortic. 2005, 106, 341–352. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | DPPH radical scavenging activity (%) | |||
| EC50 (µg/mL) | 100 µg/mL | 200 µg/mL | 500 µg/mL | |
| Pepper leaf | 159.1 ± 8.4 | 42.8 ± 0.5 | 61.4 ± 2.1 | 90.1 ± 1.0 |
| Pepper fruit | 653.4 ± 30.4 | 26.6 ± 2.0 | 32.1 ± 1.4 | 42.7 ± 0.7 |
| Luteolin | 2.1 ± 0.1 | - | - | - |
| Ascorbic acid | 6.4 ± 0.9 | - | - | - |
| Samples | ABTS·+ radical scavenging activity (%) | |||
| EC50 (µg/mL) | 100 µg/mL | 200 µg/mL | 500 µg/mL | |
| Pepper leaf | 157.6 ± 14.6 | 45.6 ± 2.0 | 71.7 ± 2.8 | 92.1 ± 4.1 |
| Pepper fruit | 357.1 ± 21.7 | 25.9 ± 1.4 | 37.7 ± 1.1 | 59.5 ± 8.1 |
| Luteolin | 3.3 ± 0.2 | - | - | - |
| Ascorbic acid | 23.7 ± 0.1 | - | - | - |
| Classes | Sub-Classes | Individual Flavonoids | ESI(+)-QToF–MS | Contents (mg/100g Dry Weight) | |
|---|---|---|---|---|---|
| Fragmentation of [M + H]+ (m/z) | PL | PF | |||
| Flavones | Apigenin | Apigenin 6,8-di-C-glucoside (vicenin-2) | 617, 595, 577, 559, 523 | ND 2 | 38.7 ± 2.6 |
| Apigenin 6-C-arabinoside-8-C-glucoside (isoschaftoside) 3 | 587, 565, 547, 529, 335 | ND | 24.1 ± 1.7 | ||
| Apigenin 6-C-glucoside-8-C-arabinoside (schaftoside) 3 | 587, 565, 547, 529, 445, 335 | ND | 18.4 ± 0.8 | ||
| Apigenin 7-O-glucoside (cosmosiin) 3 | 455, 433, 271 | 5.9 ± 0.9 | ND | ||
| Sub total | 5.9 ± 0.9 | 81.2 ± 5.1 | |||
| Luteolin | Luteolin 6,8-di-C-glucoside (lucenin-2) | 633, 611, 593, 575, 401, 373 | ND | 13.0 ± 0.9 | |
| Luteolin 6-C-arabinoside-8-C-glucoside | 603, 581, 563, 545, 527, 371 | ND | 3.1 ± 0.3 | ||
| Luteolin 6-C-glucoside-8-C-arabinoside (carlinoside) | 603, 581, 563, 545, 461, 371 | ND | 12.9 ± 0.9 | ||
| Luteolin 8-C-glucoside (orientin) 3 | 471, 449, 431, 413, 329 | ND | 60.7 ± 2.2 | ||
| Luteolin 6-C-glucoside (isoorientin) 3 | 471, 449, 431, 329 | ND | 16.6 ± 0.4 | ||
| Luteolin 7-O-(2″-O-apiosyl)glucoside | 603, 581, 449, 287 | 1472.6 ± 107.2 | 98.2 ± 2.3 | ||
| Luteolin 7-O-apiosylmalonylglucoside 4 | 689, 667, 535, 287 | 1136.3 ± 18.5 | 1166.7 ± 63.1 | ||
| Sub total | 2608.9 ± 116.2 | 1371.1 ± 61.7 | |||
| Flavonols | Quercetin | Quercetin 3-O-rhamnoside-7-O-glucoside | 633, 611, 465, 449, 303 | ND | ND |
| Quercetin 3-O-glucoside (isoquercitrin) 3 | 487, 465, 303 | ND | 24.3 ± 0.8 | ||
| Quercetin 3-O-rhamnoside (quercitrin) 3 | 471, 449, 303 | 66.0 ± 1.6 | 10.9 ± 0.3 | ||
| Sub total | 66.0 ± 1.6 | 35.3 ± 1.1 | |||
| TOTAL | 2680.8 ± 117.2 | 1487.6 ± 58.8 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, S.-Y.; Kim, H.-W.; Lee, M.-K.; Kim, H.-J.; Kim, J.-B.; Choe, J.-S.; Lee, Y.-M.; Jang, H.-H. Antioxidant and Anti-Inflammatory Activities in Relation to the Flavonoids Composition of Pepper (Capsicum annuum L.). Antioxidants 2020, 9, 986. https://doi.org/10.3390/antiox9100986
Cho S-Y, Kim H-W, Lee M-K, Kim H-J, Kim J-B, Choe J-S, Lee Y-M, Jang H-H. Antioxidant and Anti-Inflammatory Activities in Relation to the Flavonoids Composition of Pepper (Capsicum annuum L.). Antioxidants. 2020; 9(10):986. https://doi.org/10.3390/antiox9100986
Chicago/Turabian StyleCho, Soo-Yeon, Heon-Woong Kim, Min-Ki Lee, Hyeon-Jung Kim, Jung-Bong Kim, Jeong-Sook Choe, Young-Min Lee, and Hwan-Hee Jang. 2020. "Antioxidant and Anti-Inflammatory Activities in Relation to the Flavonoids Composition of Pepper (Capsicum annuum L.)" Antioxidants 9, no. 10: 986. https://doi.org/10.3390/antiox9100986
APA StyleCho, S.-Y., Kim, H.-W., Lee, M.-K., Kim, H.-J., Kim, J.-B., Choe, J.-S., Lee, Y.-M., & Jang, H.-H. (2020). Antioxidant and Anti-Inflammatory Activities in Relation to the Flavonoids Composition of Pepper (Capsicum annuum L.). Antioxidants, 9(10), 986. https://doi.org/10.3390/antiox9100986