Role of Antioxidant Natural Products in Management of Infertility: A Review of Their Medicinal Potential



Abstract
1. Infertility
Infertility and Oxidative Stress
2. Plant Extract and Infertility
2.1. Natural Products That Reverse Male Infertility
2.1.1. Plant Derived Natural Products for Treatment of Male Infertility
In Vitro Studies
In Vivo Studies
In Vitro and in Vivo Studies
2.1.2. Animal Derived Natural Products for Treatment of Male Infertility
2.1.3. Fungus Derived Natural Products for Treatment of Male Infertility
2.1.4. Formula for Treatment of Male Infertility
2.2. Natural Products That Reverse Female Infertility
2.2.1. Plant Derived Natural Products for Treatment of Female Infertility
In Vitro Studies
In Vivo Studies
2.2.2. Animal Derived Natural Products for Treatment of Female Infertility
2.3. Natural Products and Clinical Studies
2.4. Natural Products with Adverse Effects
3. Methods
4. Discussion
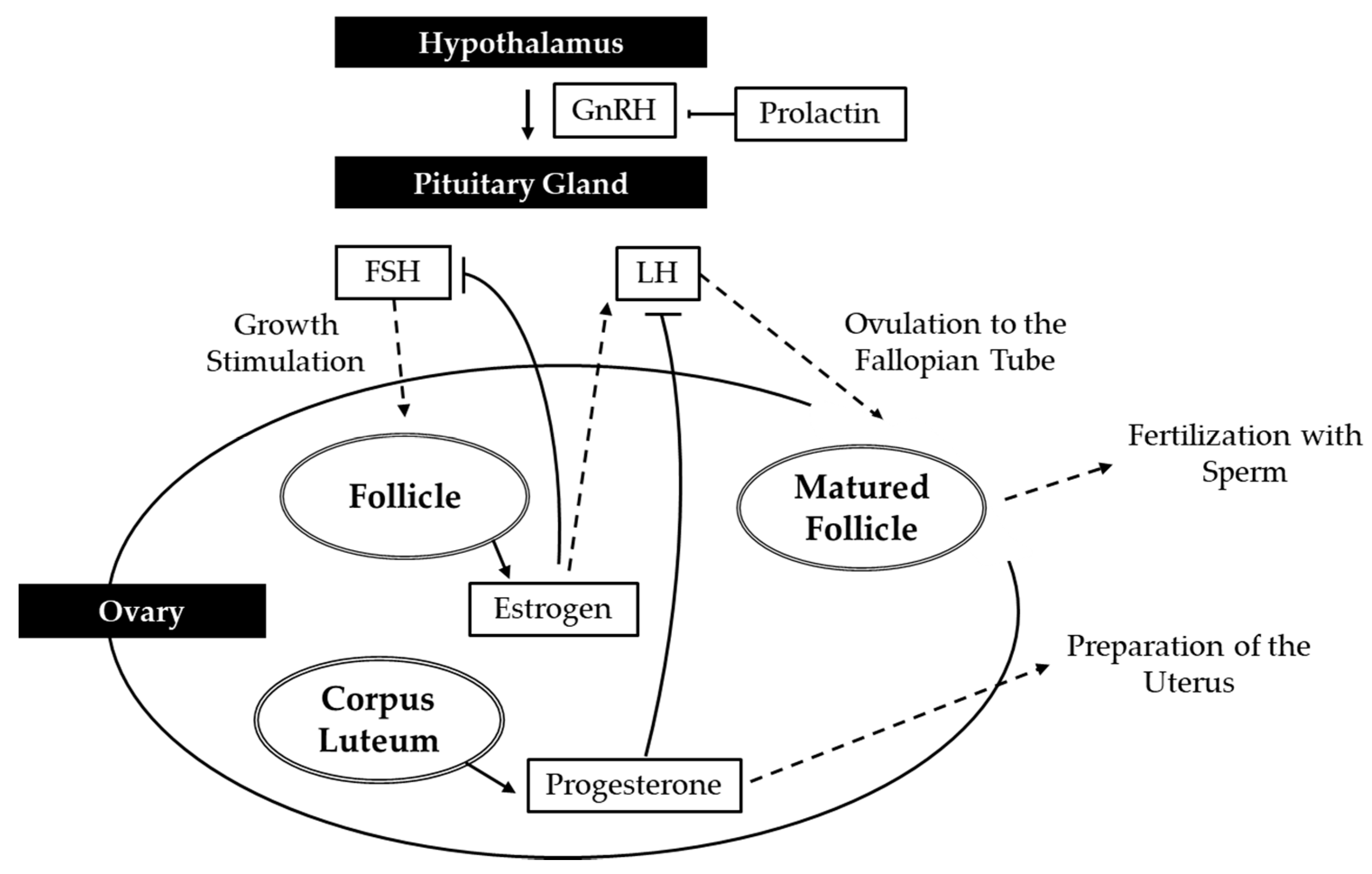
4.1. Male Reproductive System
4.2. Female Reproductive System
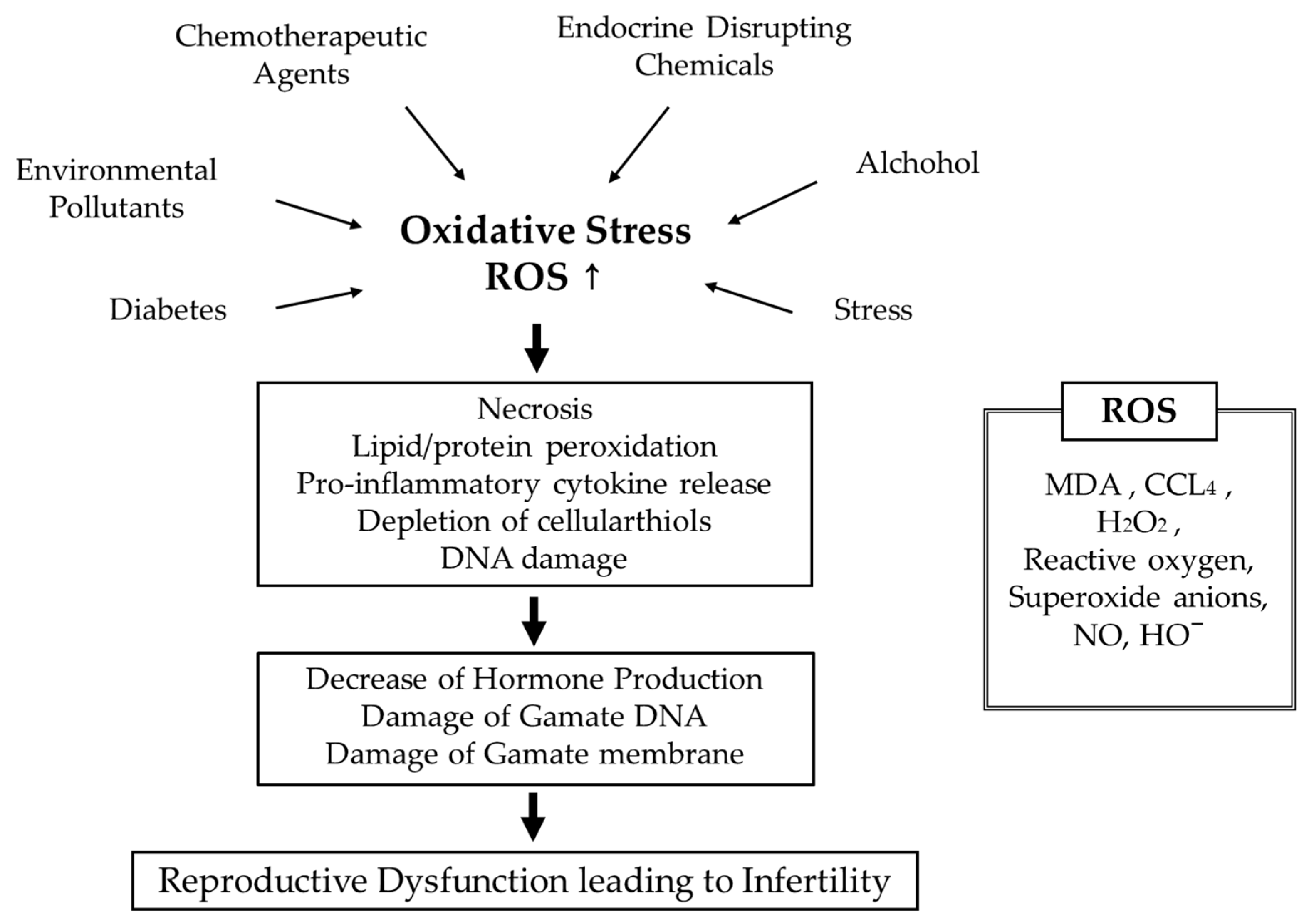
4.3. Oxidative Stress and Fertility
4.3.1. Causes of Oxidative Stress
4.3.2. Oxidative Stress and Reproductive Function
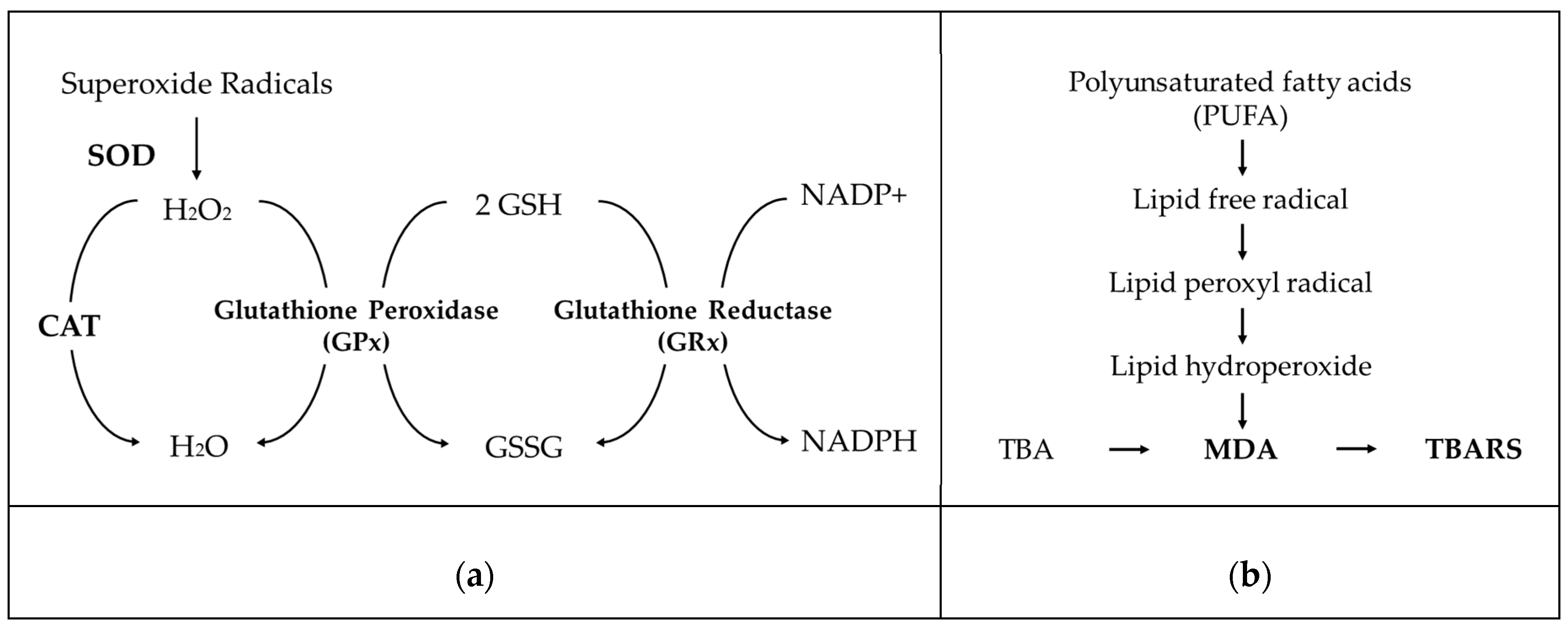
4.3.3. Oxidative Stress Related Enzymes
4.4. Diabetes and Fertility
4.5. Natural Products and Formulations with Significant Relevance to Male Fertility
4.6. Natural Products with Significant Relevance to Female Fertility
4.7. Natural Products with Adverse Effects
4.8. Insufficient Data of Existing Research
4.9. Limitations of This Review and Future Prospects
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kokubu, D.; Ooba, R.; Abe, Y.; Ishizaki, H.; Yoshida, S.; Asano, A.; Kashiwabara, S.I.; Miyazaki, H. Angelica keiskei (ashitaba) powder and its functional compound xanthoangelol prevent heat stress-induced impairment in sperm density and quality in mouse testes. J. Reprod. Dev. 2019, 65, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Troncoso, N.; Sanchez, F.; Garbarino, J.A.; Vanella, A. Propolis protects human spermatozoa from DNA damage caused by benzo[a]pyrene and exogenous reactive oxygen species. Life Sci. 2006, 78, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Adeoye, O.; Olawumi, J.; Opeyemi, A.; Christiania, O. Review on the role of glutathione on oxidative stress and infertility. JBRA Assist. Reprod. 2018, 22, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Rahim, S.M.; Taha, E.M.; Mubark, Z.M.; Aziz, S.S.; Simon, K.; Mazlan, A.G. Protective effect of Cymbopogon citratus on hydrogen peroxide-induced oxidative stress in the reproductive system of male rats. Syst. Biol. Reprod. Med. 2013, 59, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Koo, Y.K.; Park, M.J.; Hwang, Y.K.; Hwang, S.Y.; Park, N.C. Restoration of spermatogenesis using a new combined herbal formula of epimedium koreanum nakai and angelica gigas nakai in an luteinizing hormone-releasing hormone agonist-induced rat model of male infertility. World J. Men’s Health 2017, 35, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.Q.; Chen, D.L.; Lin, F.H.; Lin, L.; Shuai, O.; Wang, J.Y.; Qi, L.K.; Zhang, P. Effect of bajijiasu isolated from Morinda officinalis f. C. How on sexual function in male mice and its antioxidant protection of human sperm. J. Ethnopharmacol. 2015, 164, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Sahreen, S.; Khan, M.R.; Khan, R.A.; Shah, N.A. Effect of Carissa opaca leaves extract on lipid peroxidation, antioxidant activity and reproductive hormones in male rats. Lipids Health Dis. 2013, 12, 90. [Google Scholar] [CrossRef]
- Zhou, S.; Wen, Z.; Liang, A.; Zhang, S. Experimental research on therapeutic efficacy of traditional chinese medicine shengjing capsule extracts in treating spermatogenesis impairment induced by oxidative stress. Med. Sci. Monit. 2016, 22, 50–56. [Google Scholar] [CrossRef][Green Version]
- Pavin, N.F.; Izaguirry, A.P.; Soares, M.B.; Spiazzi, C.C.; Mendez, A.S.L.; Leivas, F.G.; dos Santos Brum, D.; Cibin, F.W.S. Tribulus terrestris protects against male reproductive damage induced by cyclophosphamide in mice. Oxidative Med. Cell. Longev. 2018, 2018, 5758191. [Google Scholar] [CrossRef]
- Nasr, A.Y. The impact of aged garlic extract on adriamycin-induced testicular changes in adult male wistar rats. Acta Histochem. 2017, 119, 648–662. [Google Scholar] [CrossRef]
- Chung, H.J.; Noh, Y.; Kim, M.S.; Jang, A.; Lee, C.E.; Myung, S.C. Steroidogenic effects of taraxacum officinale extract on the levels of steroidogenic enzymes in mouse leydig cells. Anim. Cells Syst. 2018, 22, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Chung, T.W.; Park, M.J.; Jung, Y.S.; Lee, S.O.; Kim, K.J.; Ha, K.T. Water-extracted tubers of cyperus rotundus l. Enhance endometrial receptivity through leukemia inhibitory factor-mediated expression of integrin alphavbeta3 and alphavbeta5. J. Ethnopharmacol. 2017, 208, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Uchewa, O.O. Countering the effects of lead as an environmental toxicant on the microanatomy. J. Trace Elem. Med. Biol. 2019, 52, 192–198. [Google Scholar] [CrossRef]
- Smits, R.M.; Mackenzie-Proctor, R.; Fleischer, K.; Showell, M.G. Antioxidants in fertility: Impact on male and female reproductive outcomes. Fertil. Steril. 2018, 110, 578–580. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Choi, H.J.; Chung, T.W.; Choi, J.Y.; Kim, H.S.; Jung, Y.S.; Lee, S.O.; Ha, K.T. Water-extracted perilla frutescens increases endometrial receptivity though leukemia inhibitory factor-dependent expression of integrins. J. Pharmacol. Sci. 2016, 131, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.X.; Roy, A.C.; Chan, C.; Arulkumaran, S.; Ratnam, S.S. A new molecular variant of luteinizing hormone associated with female infertility. Fertil. Steril. 1998, 69, 102–106. [Google Scholar] [CrossRef]
- Ajuogu, P.K.; Mgbere, O.O.; Bila, D.S.; McFarlane, J.R. Hormonal changes, semen quality and variance in reproductive activity outcomes of post pubertal rabbits fed Moringa oleifera lam. Leaf powder. J. Ethnopharmacol. 2019, 233, 80–86. [Google Scholar] [CrossRef]
- Mvondo, M.A.; Touomo Sakock, A.J.; Ateba, S.B.; Awounfack, C.F.; Nanbo Gueyo, T.; Njamen, D. Emmenagogue properties of Milicia excelsa (welw.) c.C. Berg (moraceae) based, at least in part, on its ability to correlate the activity of the hypothalamic-pituitary axis to that of the ovaries. J. Ethnopharmacol. 2017, 206, 283–289. [Google Scholar] [CrossRef]
- Naz, M.; Kamal, M. Classification, causes, diagnosis and treatment of male infertility: A review. Orient. Pharm. Exp. Med. 2017, 17, 89–109. [Google Scholar] [CrossRef]
- Lienou, L.L.; Telefo, B.P.; Bale, B.; Yemele, D.; Tagne, R.S.; Goka, S.C.; Lemfack, C.M.; Mouokeu, C.; Moundipa, P.F. Effect of the aqueous extract of Senecio biafrae (oliv. & hiern) j. Moore on sexual maturation of immature female rat. BMC Complementary Altern. Med. 2012, 12, 36. [Google Scholar]
- AM, M.; HA, D.S.; MY, K.J. Effects of gelam honey on sperm quality and testis of rat. Sains Malays. 2011, 40, 1243–1246. [Google Scholar]
- Kong, Z.L.; Johnson, A.; Ko, F.C.; He, J.L.; Cheng, S.C. Effect of cistanche tubulosa extracts on male reproductive function in streptozotocin(-)nicotinamide-induced diabetic rats. Nutrients 2018, 10, 1562. [Google Scholar] [CrossRef] [PubMed]
- Azza, M.; Elhabibi, E.-S.M.; El-Ghany, E.A. Preventing male infertility by marjoram and sage essential oils through modulating testicular lipid accumulation and androgens biosynthesis disruption in a rat model of dietary obesity. Egypt. J. Basic Appl. Sci. 2015, 2, 167–175. [Google Scholar]
- Hong, S.H.; Li, M.; Jeung, E.B.; Lee, G.S.; Hong, E.J.; Choi, Y.W.; An, B.S. Therapeutic effects of schisandra chinensis on the hyperprolactinemia in rat. Int. J. Oncol. 2017, 50, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Afrigan, L.; Jafari Anarkooli, I.; Sohrabi, D.; Abdanipour, A.; Yazdinezhad, A.; Sayyar, Z.; Ghorbanlou, M.; Arianmanesh, M.J.A. The effect of hydroethanolic extract of Matricaria chamomilla on the reproductive system of male rats exposed to formaldehyde. Andrologia 2019, 51, e13362. [Google Scholar] [CrossRef]
- Arafa, N.M. Efficacy of echinacea on the action of cyproterone acetate in male rats. Pak. J. Biol. Sci. 2010, 13, 966–976. [Google Scholar] [CrossRef]
- Yari, A.; Sarveazad, A.; Asadi, E.; Raouf Sarshoori, J.; Babahajian, A.; Amini, N.; Amidi, F.; Bahadoran, H.; Joghataei, M.T.; Asadi, M.H.; et al. Efficacy of Crocus sativus L. On reduction of cadmium-induced toxicity on spermatogenesis in adult rats. Andrologia 2016, 48, 1244–1252. [Google Scholar] [CrossRef]
- Nam, E.Y.; Kim, S.A.; Kim, H.; Kim, S.H.; Han, J.H.; Lee, J.H.; Kim, D.I. Akt activation by evodiae fructus extract protects ovary against 4-vinylcyclohexene diepoxide-induced ovotoxicity. J. Ethnopharmacol. 2016, 194, 733–739. [Google Scholar] [CrossRef]
- Mozafari, A.A.; Shahrooz, R.; Ahmadi, A.; Malekinjad, H.; Mardani, K. Protective Effect of Ethyl Pyruvate on Mice Sperm Parameters in Phenylhydrazine Induced Hemolytic Anemia; Veterinary Research Forum; Faculty of Veterinary Medicine, Urmia University: Urmia, Iran, 2016; p. 63. [Google Scholar]
- Ji, H.J.; Wang, D.M.; Wu, Y.P.; Niu, Y.Y.; Jia, L.L.; Liu, B.W.; Feng, Q.J.; Feng, M.L. Wuzi yanzong pill, a chinese polyherbal formula, alleviates testicular damage in mice induced by ionizing radiation. BMC Complementary Altern. Med. 2016, 16, 509. [Google Scholar] [CrossRef]
- Tiwari, A.; Singh, P.; Jaitley, P.; Sharma, S.; Prakash, A.; Mandil, R.; Choudhury, S.; Gangwar, N.K.; Garg, S.K. Eucalyptus robusta leaves methanolic extract suppresses inflammatory mediators by by specifically targeting TLR4/TLR9, MPO, COX2, iNOS and inflammatory cytokines in experimentally-induced endometritis in rats. J. Ethnopharmacol. 2018, 213, 149–158. [Google Scholar] [CrossRef]
- Rahmouni, F.; Daoud, S.; Rebai, T. Teucrium polium attenuates carbon tetrachloride-induced toxicity in the male reproductive system of rats. Andrologia 2019, 51, e13182. [Google Scholar] [CrossRef] [PubMed]
- Askaripour, M.; Hasanpour, A.; Hosseini, F.; Moshrefi, M.; Moshtaghi, G.; Hasannejad, M.; Rajabi, S.; Nematollahi-Mahani, S.N. The effect of aqueous extract of Rosa damascena on formaldehyde-induced toxicity in mice testes. Pharm. Biol. 2018, 56, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Fakher, S.; Seghatoleslam, A.; Noorafshan, A.; Karbalay-Doust, S.; Rahmanifard, M.; Rashidi, M. The impact of echium amoenum distillate on naturally boosting fertility: Potential ameliorative role in male mice reproductive parameters. Iran. J. Med. Sci. 2019, 44, 227. [Google Scholar] [PubMed]
- Orkhon, B. Astragalus root induces ovarian betaoxidation and suppresses estrogendependent. Mol. Med. Rep. 2018, 18, 5198–5206. [Google Scholar] [PubMed]
- Saleem, M.A.; Al-Attar, M.S. Protective effects of mentha spicata aqueous extract against ifosfamide induced chromosomal aberrations and sperm abnormalities in male albino mice. Trends Biotechnol. Res. 2013, 2, 1. [Google Scholar]
- Sm, S.; Mahaboob Basha, P. Fluoride exposure aggravates the testicular damage and sperm quality in diabetic mice: Protective role of ginseng and banaba. Biol. Trace Elem. Res. 2017, 177, 331–344. [Google Scholar] [CrossRef]
- Heo, J. Dongui Bogam; Namsandang: Seoul, Korea, 1994; Volume 90. [Google Scholar]
- Agbodjento, E.; Klotoé, J.R.; Sacramento, T.I.; Dougnon, V.; Tchabi, F.L.; Déguénon, E.; Atègbo, J.-M. Ethnobotanical knowledge of medicinal plants used in the treatment of male infertility in southern benin. Adv. Tradit. Med. 2020. [Google Scholar] [CrossRef]
- Mahaldashtian, M.; Naghdi, M.; Ghorbanian, M.T.; Makoolati, Z.; Movahedin, M.; Mohamadi, S.M. In vitro effects of date palm (Phoenix dactylifera L.) pollen on colonization of neonate mouse spermatogonial stem cells. J. Ethnopharmacol. 2016, 186, 362–368. [Google Scholar] [CrossRef]
- Jung, S.E.; Kim, Y.H.; Cho, S.; Kim, B.J.; Lee, H.S.; Hwang, S.; Kim, G.B.; Kim, Y.H.; Pang, M.G.; Lee, S.; et al. A phytochemical approach to promotion of self-renewal in murine spermatogonial stem cell by using sedum sarmentosum extract. Sci. Rep. 2017, 7, 11441. [Google Scholar] [CrossRef]
- Wang, C.; Jin, Y.; Jin, Y. Promoting effect of licorice extract on spermatogonial proliferation and spermatocytes differentiation of neonatal mice in vitro. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 149–155. [Google Scholar] [CrossRef]
- Yang, F.; Wei, Y.; Liao, B.; Wei, G.; Qin, H.; Pang, X.; Wang, J. Lycium barbarum polysaccharide prevents cisplatin-induced mltc-1 cell apoptosis and autophagy via regulating endoplasmic reticulum stress pathway. Drug Des. Dev. Ther. 2018, 12, 3211–3219. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.-S.; Kim, W.-N.; Yang, W.-M.; Kim, H.-Y.; Oh, J.-H.; Park, S.-K. Cytoprotective effects of Morinda officinalis against hydrogen peroxide-induced oxidative stress in leydig tm3 cells. Asian J. Androl. 2008, 10, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Ilfergane, A.; Henkel, R.R. Effect of Typha capensis (rohrb.)n.E.Br. Rhizome extract f1 fraction on cell viability, apoptosis induction and testosterone production in tm3-leydig cells. Andrologia 2018, 50, e12854. [Google Scholar] [CrossRef] [PubMed]
- Afsar, T.; Razak, S.; Khan, M.R.; Almajwal, A. Acacia hydaspica ethyl acetate extract protects against cisplatin-induced DNA damage, oxidative stress and testicular injuries in adult male rats. BMC Cancer 2017, 17, 883. [Google Scholar] [CrossRef]
- Salahipour, M.H.; Hasanzadeh, S.; Malekinejad, H. Ameliorative effects of Achillea millefolium inflorescences alcoholic extract against nicotine-induced reproductive failure in rat. Exp. Toxicol. Pathol. 2017, 69, 504–516. [Google Scholar] [CrossRef]
- NS, A.E.R.; Al-Ahmary, S.H. Effect of sapogenin extract on anti-infertility induced by aluminium chloride in male rats. Adv. Med. Plant Res. 2013, 1, 77–85. [Google Scholar]
- Zakaria, F.H.; Haron, M.N. Effect of bee bread on male reproductive system of sprague dawley rat. J. Agrobiotechnol. 2018, 9, 12–17. [Google Scholar]
- Jiang, Z.; Wang, J.; Li, X.; Zhang, X. Echinacoside and Cistanche tubulosa (schenk) R. Wight ameliorate bisphenol a-induced testicular and sperm damage in rats through gonad axis regulated steroidogenic enzymes. J. Ethnopharmacol. 2016, 193, 321–328. [Google Scholar] [CrossRef]
- Oguzturk, H.; Ciftci, O.; Aydin, M.; Timurkaan, N.; Beytur, A.; Yilmaz, F. Ameliorative effects of curcumin against acute cadmium toxicity on male reproductive system in rats. Andrologia 2012, 44, 243–249. [Google Scholar] [CrossRef]
- Tvrdá, E.; Lukác, N.; Jambor, T.; Lukácová, J.; Massányi, P. Curcumin in male fertility: Effects on spermatozoa vitality and oxidative balance. J. Microbiol. Biotechnol. Food Sci. 2015, 4, 120. [Google Scholar] [CrossRef]
- El-Kashlan, A.M.; Nooh, M.M.; Hassan, W.A.; Rizk, S.M. Therapeutic potential of date palm pollen for testicular dysfunction induced by thyroid disorders in male rats. PLoS ONE 2015, 10, e0139493. [Google Scholar] [CrossRef] [PubMed]
- Hassan, E.; Kahilo, K.; Kamal, T.; El-Neweshy, M.; Hassan, M.J. Protective effect of diallyl sulfide against lead-mediated oxidative damage, apoptosis and down-regulation of cyp19 gene expression in rat testes. Life Sci. 2019, 226, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhou, B.; Li, X.; Kirby, G.M.; Zhang, X. Echinacoside increases sperm quantity in rats by targeting the hypothalamic androgen receptor. Sci. Rep. 2018, 8, 3839. [Google Scholar] [CrossRef] [PubMed]
- Wahab, N.A.; Mokhtar, N.M.; Halim, W.N.; Das, S. The effect of Eurycoma longifolia jack on spermatogenesis in estrogen-treated rats. Clinics (Sao Paulo Braz.) 2010, 65, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.I.; Lasheen, N.N.; El-Zawahry, K.M. Ginkgo biloba ameliorates subfertility induced by testicular ischemia/reperfusion injury in adult wistar rats: A possible new mitochondrial mechanism. Oxidative Med. Cell. Longev. 2016, 2016, 6959274. [Google Scholar] [CrossRef] [PubMed]
- Khaki, A.; Fathiazad, F.; Nouri, M.; Khaki, A.A.; Ozanci, C.C.; Ghafari-Novin, M.; Hamadeh, M. The effects of ginger on spermatogenesis and sperm parameters of rat. Int. J. Reprod. BioMed. 2009, 7, 7–12. [Google Scholar] [CrossRef]
- Gang, L.S.; Song, D.Y.; Qiang, N.; Zhi, X.S.; Juan, P.L.; Lin, M.R.; Xia, J.M.; Ling, F.G.; Ming, L.J.; Xia, G.S.J. Grape seed proanthocyanidin extract alleviates arsenic-induced oxidative reproductive toxicity in male mice. Biomed. Environ. Sci. 2015, 28, 272–280. [Google Scholar]
- Chenniappan, K.; Murugan, K. Therapeutic and fertility restoration effects of ionidium suffruticosum on sub-fertile male albino wistar rats: Effects on testis and caudal spermatozoa. Pharm. Biol. 2017, 55, 946–957. [Google Scholar] [CrossRef]
- Shah, N.A.; Khan, M.R. Increase of glutathione, testosterone and antioxidant effects of Jurenia dolomiaea on ccl4 induced testicular toxicity in rat. BMC Complementary Altern. Med. 2017, 17, 206. [Google Scholar] [CrossRef]
- Ebokaiwe, A.P.; Ijomone, O.M.; Osawe, S.O.; Chukwu, C.J.; Ejike, C.; Zhang, G.; Wang, F. Alteration in sperm characteristics, endocrine balance and redox status in rats rendered diabetic by streptozotocin treatment: Attenuating role of Loranthus micranthus. Redox Rep. 2018, 23, 194–205. [Google Scholar] [CrossRef]
- Shi, G.J.; Zheng, J.; Wu, J.; Qiao, H.Q.; Chang, Q.; Niu, Y.; Sun, T.; Li, Y.X.; Yu, J.Q. Protective effects of lycium barbarum polysaccharide on male sexual dysfunction and fertility impairments by activating hypothalamic pituitary gonadal axis in streptozotocin-induced type-1 diabetic male mice. Endocr. J. 2017, 64, 907–922. [Google Scholar] [CrossRef] [PubMed]
- Onaolapo, A.Y.; Oladipo, B.P.; Onaolapo, O.J. Cyclophosphamide-induced male subfertility in mice: An assessment of the potential benefits of maca supplement. Andrologia 2018, 50, e12911. [Google Scholar] [CrossRef]
- Valdivia Cuya, M.; Yarasca De La Vega, K.; Levano Sanchez, G.; Vasquez Cavero, J.; Temoche Garcia, H.; Torres Torres, L.; Cruz Ornetta, V. Effect of Lepidium meyenii (maca) on testicular function of mice with chemically and physically induced subfertility. Andrologia 2016, 48, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Zade, V.S.; Dabhadkar, D.K.; Thakare, V.G.; Pare, S.R. Effect of aqueous extract of Moringa oleifera seed on sexual activity of male albino rats. Int. J. Bio Forum 2013, 5, 129–140. [Google Scholar]
- Tekayev, M.; Bostancieri, N.; Saadat, K.; Turker, M.; Yuncu, M.; Ulusal, H.; Cicek, H.; Arman, K. Effects of Moringa oleifera lam extract (mole) in the heat shock protein 70 expression and germ cell apoptosis on experimentally induced cryptorchid testes of rats. Gene 2019, 688, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Adana, M.Y.; Akang, E.N.; Naidu, E.C.S.; Aniekan, P.I.; Kouame, K.; Offor, U.; Ogedengbe, O.O.; Azu, O.O. Testicular microanatomical and hormonal alterations following use of antiretroviral therapy in sprague dawley rats: Role of naringenin. Andrologia 2018, 50, e13137. [Google Scholar] [CrossRef]
- Haseena, S.; Aithal, M.; Das, K.K.; Saheb, S.H. Effect of Nigella sativa seed powder on testosterone and lh levels in sterptozotocine induced diabetes male albino rats. J. Pharm. Sci. Res. 2015, 7, 234. [Google Scholar]
- Sharma, M.; Arya, D.; Bhagour, K.; Gupta, R.S. Modulatory effects of methanolic fruit fraction of Pedalium murex on sulphasalazine-induced male reproductive disruption. Andrologia 2019, 51, e13190. [Google Scholar] [CrossRef]
- Kang, H.R.; Lee, Y.A.; Kim, Y.H.; Lee, D.G.; Kim, B.J.; Kim, K.J.; Kim, B.G.; Oh, M.G.; Han, C.K.; Lee, S.; et al. Petasites japonicus stimulates the proliferation of mouse spermatogonial stem cells. PLoS ONE 2015, 10, e0133077. [Google Scholar] [CrossRef]
- Bahmanpour, S.; Talaei, T.; Vojdani, Z.; Panjehshahin, M.; Poostpasand, A.; Zareei, S.; Ghaeminia, M.J. Effect of Phoenix dactylifera pollen on sperm parameters and reproductive system of adult male rats. Iran. J. Med. Sci. 2006, 31, 208–212. [Google Scholar]
- Arun, S.; Burawat, J.; Yannasithinon, S.; Sukhorum, W.; Limpongsa, A.; Iamsaard, S. Phyllanthus emblica leaf extract ameliorates testicular damage in rats with chronic stress. J. Zhejiang Univ. Sci. B 2018, 19, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Heidari, R.; Alizadeh, R.; Abbasi, N.; Pasbakhsh, P.; Hedayatpour, A.; Farajpour, M.; Khaleghi, M.R.; Abbasi, M.; Dehpour, A.R. Do Pilea microphylla improve sperm DNA fragmentation and sperm parameters in varicocelized rats? Acta Med. Iran. 2015, 53, 547–554. [Google Scholar] [PubMed]
- Ola-Davies, O.; Ajani, O.S. Semen characteristics and sperm morphology of Pistia stratiotes linn. (araceae) protected male albino rats (wistar strain) exposed to sodium arsenite. J. Complementary Integr. Med. 2016, 13, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Singhal, S.; Kumar, N.; Rao, C.M.; Sumalatha, S.; Dave, J.; Dave, R.; Nandakumar, K. Standardised extract of safed musli (Chlorophytum borivilianum) increases aphrodisiac potential besides being safe in male wistar rats. Andrologia 2016, 48, 1236–1243. [Google Scholar] [CrossRef]
- Luthfi, M.J.F. Effect of Lunasia amara blanco on sperm number, sperm motility, and testicular histology of male rats. Biol. Med. Nat. Prod. Chem. 2015, 4, 31–33. [Google Scholar] [CrossRef]
- Mishra, R.K.; Jain, A.; Singh, S.K. Profertility effects of shilajit on cadmium-induced infertility in male mice. Andrologia 2018, 50, e13064. [Google Scholar] [CrossRef]
- Abedi, H.; Jahromi, H.K.; Hashemi, S.A.; Jashni, H.K.; Jahromi, Z.K.; Pourahmadi, M. The effect of silymarin on spermatogenesis process in rats. Int. J. Med. Res. Health Sci. 2016, 5, 146–150. [Google Scholar]
- Kanedi, M. Plant extracts of suruhan (Peperomia pellucida L. Kunth) ameliorate infertility of male mice with alloxan-induced hyperglycemia. Int. J. Biomed. Res. 2019, 10, 1–4. [Google Scholar]
- Akomolafe, S.F.; Oboh, G.; Akindahunsi, A.A.; Afolayan, A.J. Ethanol-induced male infertility: Effects of aqueous leaf extract of Tetracarpidium conophorum. Andrologia 2017, 49, e12759. [Google Scholar] [CrossRef]
- Akomolafe, S.F.; Oboh, G. Walnut leaf extract acts as a fertility agent in male wistar albino rats—A search for herbal male fertility enhancer. J. Complementary Integr Med. 2017, 15. [Google Scholar] [CrossRef]
- Guesmi, F.; Beghalem, H.; Tyagi, A.K.; Ali, M.B.; Mouhoub, R.B.; Bellamine, H.; Landoulsi, A. Prevention of H2O2 induced oxidative damages of rat testis by Thymus algeriensis. Biomed. Environ Sci. 2016, 29, 275–285. [Google Scholar] [PubMed]
- Oliveira, P.F.; Tomas, G.D.; Dias, T.R.; Martins, A.D.; Rato, L.; Alves, M.G.; Silva, B.M. White tea consumption restores sperm quality in prediabetic rats preventing testicular oxidative damage. Reprod. Biomed. Online 2015, 31, 544–556. [Google Scholar] [CrossRef] [PubMed]
- Oridupa, O.A.; Folasire, O.F.; Owolabi, A.J.; Aina, O. Effect of traditional treatment of diabetes mellitus with xanthosoma sagittifolium on the male reproductive system of alloxan-induced diabetic wistar rats. Drug Res. (Stuttg.) 2017, 67, 337–342. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Akomolafe, S.F.; Oboh, G.; Akindahunsi, A.A.; Afolayan, A.J. Tetracarpidium conophorum ameliorates oxidative reproductive toxicity induced by ethanol in male rats. BMC Complementary Altern. Med. 2015, 15, 439. [Google Scholar] [CrossRef]
- Saeid, J.; Shanoon, A.; Marbut, M.J. Effects of Zingiber officinale aqueous extract on semen characteristic and some blood plasma, semen plasma parameters in the broilers breeder male. Int. J. Poult. Sci. 2011, 10, 629–633. [Google Scholar] [CrossRef]
- Mao, C.F.; Zhang, X.R.; Johnson, A.; He, J.L.; Kong, Z.L. Modulation of diabetes mellitus-induced male rat reproductive dysfunction with micro-nanoencapsulated Echinacea purpurea ethanol extract. Biomed. Res. Int. 2018, 2018, 4237354. [Google Scholar] [CrossRef] [PubMed]
- Seres, A.B.; Ducza, E.; Bathori, M.; Hunyadi, A.; Beni, Z.; Dekany, M.; Hajagos-Toth, J.; Verli, J.; Gaspar, R. Androgenic effect of honeybee drone milk in castrated rats: Roles of methyl palmitate and methyl oleate. J. Ethnopharmacol. 2014, 153, 446–453. [Google Scholar] [CrossRef]
- Kumari, S.; Nayak, G.; Lukose, S.T.; Kalthur, S.G.; Bhat, N.; Hegde, A.R.; Mutalik, S.; Kalthur, G.; Adiga, S.K. Indian propolis ameliorates the mitomycin c-induced testicular toxicity by reducing DNA damage and elevating the antioxidant activity. Biomed. Pharmacother. 2017, 95, 252–263. [Google Scholar] [CrossRef]
- Martinez, G.; Hograindleur, J.P.; Voisin, S.; Abi Nahed, R.; Abd El Aziz, T.M.; Escoffier, J.; Bessonnat, J.; Fovet, C.M.; De Waard, M.; Hennebicq, S.; et al. Spermaurin, an la1-like peptide from the venom of the scorpion Scorpio maurus palmatus, improves sperm motility and fertilization in different mammalian species. Mol. Hum. Reprod. 2017, 23, 116–131. [Google Scholar]
- Johnson, A.; Cheng, S.C.; Tsou, D.; Kong, Z.L. Attenuation of reproductive dysfunction in diabetic male rats with timber cultured Antrodia cinnamomea ethanol extract. Biomed. Pharmacother. 2019, 112, 108684. [Google Scholar] [CrossRef]
- Qu, N.; Kuramasu, M.; Hirayanagi, Y.; Nagahori, K.; Hayashi, S.; Ogawa, Y.; Terayama, H.; Suyama, K.; Naito, M.; Sakabe, K.; et al. Gosha-jinki-gan recovers spermatogenesis in mice with busulfan-induced aspermatogenesis. Int. J. Mol. Sci. 2018, 19, 2606. [Google Scholar] [CrossRef] [PubMed]
- Soni, K.K.; Zhang, L.T.; Choi, B.R.; Karna, K.K.; You, J.H.; Shin, Y.S.; Lee, S.W.; Kim, C.Y.; Zhao, C.; Chae, H.J.; et al. Protective effect of motiliperm in varicocele-induced oxidative injury in rat testis by activating phosphorylated inositol requiring kinase 1alpha (p-ire1alpha) and phosphorylated c-jun n-terminal kinase (p-jnk) pathways. Pharm. Biol. 2018, 56, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Zang, Z.J.; Ji, S.Y.; Zhang, Y.N.; Gao, Y.; Zhang, B. Effects of saikokaryukotsuboreito on spermatogenesis and fertility in aging male mice. Chin. Med. J. (Engl.) 2016, 129, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Zhang, Y.S.; Li, Y.Q.; Wu, X.N.; Chai, L.M.; Yue, L.F.; Juan, L.; Ding, J.; Li, X.R.; Chen, M.; et al. Effect of yishenjianpi recipe on semen quality and sperm mitochondria in mice with oligoasthenozoospermia induced by tripterygium glycosides. Afr. J. Tradit. Complementary Altern. Med. 2017, 14, 87–95. [Google Scholar] [CrossRef]
- Chantal, N.M.; Paul Désiré, D.D.; Caude, B.D.; Yolande Sandrine, M.N.; Lohik, M.N.; Francine, M.M.; Larissa, D.T.; Mireille, K.P.; Pierre, K. Neuroprotective effects of the Anthocleista schweinfurthii Gilg. (loganiaceae) stem bark extract in postmenopause-like model of ovariectomized wistar rats. J. Complementary Integr. Med. 2019, 16. [Google Scholar] [CrossRef]
- El-Sayyad, H.I.H.; El-Shershaby, E.M.F.; El-Mansi, A.A. Anti-hypercholesterolemic impacts of barley and date palm fruits on the ovary of Wistar albino rats and their offspring. Reprod. Biol. 2018, 18, 236–251. [Google Scholar] [CrossRef]
- Dou, L. The effect of cinnamon on polycystic ovary syndrome in a mouse model. Reprod. Biol. Endocrinol. 2018, 16, 99. [Google Scholar] [CrossRef]
- Ngadjui, E.; Nkeng-Efouet, P.A.; Nguelefack, T.B.; Kamanyi, A.; Watcho, P. High fat diet-induced estrus cycle disruption: Effects of ficus asperifolia. J. Complementary Integr. Med. 2015, 12, 205–215. [Google Scholar] [CrossRef]
- Carbonel, A.A.F.; Lima, P.D.A.; Lim, J.J.; Fuchs, L.F.P.; Paiotti, A.P.R.; Sasso, G.R.d.S.; Simões, R.S.; Soares Junior, J.M.; Baracat, E.C.; Simoes, M.d.J.J. The effects of soybean isoflavones and 17β-estradiol in uterus and mammary glands of diabetic rat models. Gynecol. Endocrinol. 2018, 34, 314–319. [Google Scholar] [CrossRef]
- Ghanbari, E.; Khazaei, M.R.; Khazaei, M.; Nejati, V. Royal jelly promotes ovarian follicles growth and increases steroid hormones in immature rats. Int. J. Fertil. Steril. 2018, 11, 263–269. [Google Scholar]
- Colone, M.; Marelli, G.; Unfer, V.; Bozzuto, G.; Molinari, A.; Stringaro, A. Inositol activity in oligoasthenoteratospermia—An in vitro study. Eur. Rev. Med Pharmacol. Sci. 2010, 14, 891–896. [Google Scholar] [PubMed]
- Khaleghi, S.; Bakhtiari, M.; Asadmobini, A.; Esmaeili, F. Tribulus terrestris extract improves human sperm parameters in vitro. J. Evid. Based Complementary Altern. Med. 2017, 22, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Asadmobini, A.; Bakhtiari, M.; Khaleghi, S.; Esmaeili, F.; Mostafaei, A. The effect of tribulus terrestris extract on motility and viability of human sperms after cryopreservation. Cryobiology 2017, 75, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Du, B.; Jiang, X.; Ma, M.; Shi, L.; Zhang, Q.; Zhou, L. Effects of combining low-dose aspirin with a chinese patent medicine on follicular blood flow and pregnancy outcome. Mol. Med. Rep. 2014, 10, 2372–2376. [Google Scholar] [CrossRef] [PubMed]
- Kort, D.H.; Lobo, R.A. Preliminary evidence that cinnamon improves menstrual cyclicity in women with polycystic ovary syndrome: A randomized controlled trial. Am. J. Obstet. Gynecol. 2014, 211, e481–e487. [Google Scholar] [CrossRef]
- Swaroop, A.; Jaipuriar, A.S.; Gupta, S.K.; Bagchi, M.; Kumar, P.; Preuss, H.G.; Bagchi, D. Efficacy of a novel fenugreek seed extract (Trigonella foenum-graecum, furocyst) in polycystic ovary syndrome (pcos). Int. J. Med. Sci. 2015, 12, 825–831. [Google Scholar] [CrossRef]
- Najafizadeh, P.; Dehghani, F.; Shahin, M.P.; Taj, S.H. The effect of a hydro-alcoholic extract of olive fruit on reproductive argons in male sprague-dawley rat. Iran. J. Reprod. Med. 2013, 11, 293. [Google Scholar]
- Khaki, A.; Khaki, A.A.; Rajabzadeh, A. The effects of permethrin and antioxidant properties of Allium cepa (onion) on testicles parameters of male rats. Toxin Rev. 2017, 36, 1–6. [Google Scholar] [CrossRef]
- Adeoye, O.J.; Olayinka, O.A.; Bernard, S.A.; Sikiru, O.A.; Kofoworola, O.I.; Olufemi, O.J.; Adekunle, A.A. Testicular and spermatotoxic potential of methanol extract of red cultivar Allium cepa in rats. J. Complementary Integr. Med. 2018, 15. [Google Scholar] [CrossRef]
- Nath, S.; Choudhury, M.D.; Roychoudhury, S.; Talukdar, A.D.; Misro, M.M. Male contraceptive efficacy of Ricinus communis L. Extract. J. Ethnopharmacol. 2013, 149, 328–334. [Google Scholar] [CrossRef]
- Asuquo, O.R.; Ekanem, T.B.; Udoh, P.B.; Eluwa, M.A. Histomorphological study of the anti-fertility effect of Spondias mombin L. in adult male rats. IOSR J. Pharm. Biol. Sci. 2012, 3, 29–34. [Google Scholar] [CrossRef]
- Guo, J.; Huang, Y.; Lei, X.; Zhang, H.; Xiao, B.; Han, Z.; Liang, C.; Yang, W. Reproductive systemic toxicity and mechanism of glucosides of Tripterygium wilfordii hook. F.(gtw). Ann. Clin. Laboratory Sci. 2019, 49, 36–49. [Google Scholar]
- Zegers-Hochschild, F.; Adamson, G.D.; de Mouzon, J.; Ishihara, O.; Mansour, R.; Nygren, K.; Sullivan, E.; Van der Poel, S. The international committee for monitoring assisted reproductive technology (icmart) and the world health organization (who) revised glossary on art terminology, 2009. Hum. Reprod. 2009, 24, 2683–2687. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Gong, T.-T.; Jiang, Y.-T.; Zhang, S.; Zhao, Y.-H.; Wu, Q.-J. Global, regional, and national prevalence and disability-adjusted life-years for infertility in 195 countries and territories, 1990–2017: Results from a global burden of disease study, 2017. Aging (Albany NY) 2019, 11, 10952. [Google Scholar] [CrossRef] [PubMed]
- Cissen, M.; Bensdorp, A.; Cohlen, B.J.; Repping, S.; de Bruin, J.P.; van Wely, M. Assisted reproductive technologies for male subfertility. Cochrane Database Syst. Rev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Allen, V.M.; Wilson, R.D.; Cheung, A.; Blight, C.; Désilets, V.A.; Gagnon, A.; Langlois, S.F.; Summers, A.; Wyatt, P.; Claman, P. Pregnancy outcomes after assisted reproductive technology. J. Obstet. Gynaecol. Canada 2006, 28, 220–233. [Google Scholar] [CrossRef]
- Xu, X.; Yin, H.; Tang, D.; Zhang, L.; Gosden, R.G. Application of traditional chinese medicine in the treatment of infertility. Hum. Fertil. 2003, 6, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, F.; Nikzad, H.; Taherian, A.; Amini Mahabadi, J.; Salehi, M. Effects of herbal medicine on male infertility. Anat. Sci. J. 2013, 10, 3–16. [Google Scholar]
- Jiang, D.; Coscione, A.; Li, L.; Zeng, B.-Y. Effect of chinese herbal medicine on male infertility. In International Review of Neurobiology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 135, pp. 297–311. [Google Scholar]
- García-Rodríguez, A.; Gosálvez, J.; Agarwal, A.; Roy, R.; Johnston, S. DNA damage and repair in human reproductive cells. Int. J. Mol. Sci. 2018, 20, 31. [Google Scholar] [CrossRef]
- Zwart, A.D.; Urban, R.J.; Odell, W.D.; Veldhuis, J.D. Contrasts in the gonadotropin-releasing hormone dose-response relationships for luteinizing hormone, follicle-stimulating hormone and alpha-subunit release in young versus older men: Appraisal with high-specificity immunoradiometric assay and deconvolution analysis. Eur. J. Endocrinol. 1996, 135, 399–406. [Google Scholar]
- Johnsen, S.G. Maintenance of spermatogenesis induced by hmg treatment by means of continuous hcg treatment in hypogonadotrophic men. Acta Endocrinol. (Copenh.) 1978, 89, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, F.; Ye, L.; Zirkin, B.; Chen, H. Steroidogenesis in leydig cells: Effects of aging and environmental factors. Reproduction 2017, 154, R111–R122. [Google Scholar] [CrossRef] [PubMed]
- Midzak, A.; Zirkin, B.; Papadopoulos, V. Translocator protein: Pharmacology and steroidogenesis. Biochem. Soc. Trans. 2015, 43, 572–578. [Google Scholar] [CrossRef]
- Aghazadeh, Y.; Zirkin, B.R.; Papadopoulos, V. Pharmacological regulation of the cholesterol transport machinery in steroidogenic cells of the testis. Vitam. Horm. 2015, 98, 189–227. [Google Scholar]
- Singh, J.; O’Neill, C.; Handelsman, D.J. Induction of spermatogenesis by androgens in gonadotropin-deficient (hpg) mice. Endocrinology 1995, 136, 5311–5321. [Google Scholar] [CrossRef]
- Dohle, G.R.; Smit, M.; Weber, R.F.A. Androgens and male fertility. World J. Urol. 2003, 21, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.V.; Clapauch, R. Female infertility of endocrine origin. Arq. Bras. Endocrinol. Metabol. 2014, 58, 144–152. [Google Scholar] [CrossRef]
- de Ziegler, D.; Borghese, B.; Chapron, C. Endometriosis and infertility: Pathophysiology and management. Lancet 2010, 376, 730–738. [Google Scholar] [CrossRef]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Foster, W.G.; Neal, M.S.; Han, M.-S.; Dominguez, M.M. Environmental contaminants and human infertility: Hypothesis or cause for concern? J. Toxicol. Environ. Health B Crit. Rev. 2008, 11, 162–176. [Google Scholar] [CrossRef]
- Sifakis, S.; Androutsopoulos, V.P.; Tsatsakis, A.M.; Spandidos, D.A. Human exposure to endocrine disrupting chemicals: Effects on the male and female reproductive systems. Environ. Toxicol. Pharmacol. 2017, 51, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, M.; Cernera, G.; Giovannelli, P.; Galasso, G.; Bilancio, A.; Migliaccio, A.; Castoria, G. Recent advances on bisphenol-a and endocrine disruptor effects on human prostate cancer. Mol. Cell. Endocrinol. 2017, 457, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Dohle, G.R. Male infertility in cancer patients: Review of the literature. Int. J. Urol. 2010, 17, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Harkiss, D.; Buckingham, D. Relationship between iron-catalysed lipid peroxidation potential and human sperm function. J. Reprod. Fertil. 1993, 98, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Ohyashiki, T.; Ohtsuka, T.; Mohri, T. Increase of the molecular rigidity of the protein conformation in the intestinal brush-border membranes by lipid peroxidation. Biochim. Biophys. Acta 1988, 939, 383–392. [Google Scholar] [CrossRef]
- Ruder, E.H.; Hartman, T.J.; Goldman, M.B. Impact of oxidative stress on female fertility. Curr. Opin. Obstet. Gynecol. 2009, 21, 219–222. [Google Scholar] [CrossRef]
- Ruder, E.H.; Hartman, T.J.; Blumberg, J.; Goldman, M.B. Oxidative stress and antioxidants: Exposure and impact on female fertility. Hum. Reprod. Update 2008, 14, 345–357. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ros damage and regulating ros signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- La Vignera, S.; Calogero, A.E.; Condorelli, R.; Lanzafame, F.; Giammusso, B.; Vicari, E. Andrological characterization of the patient with diabetes mellitus. Minerva Endocrinol. 2009, 34, 1–9. [Google Scholar] [PubMed]
- La Vignera, S.; Condorelli, R.A.; Di Mauro, M.; Lo Presti, D.; Mongioì, L.M.; Russo, G.; Calogero, A.E. Reproductive function in male patients with type 1 diabetes mellitus. Andrology 2015, 3, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Miró, M.; Chillarón, J.J.; Pedro-Botet, J. Testosterone deficiency, metabolic syndrome and diabetes mellitus. Med. Clin. (Barc.) 2016, 146, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Abbasihormozi, S.H.; Babapour, V.; Kouhkan, A.; Niasari Naslji, A.; Afraz, K.; Zolfaghary, Z.; Shahverdi, A.H. Stress hormone and oxidative stress biomarkers link obesity and diabetes with reduced fertility potential. Cell J. 2019, 21, 307–313. [Google Scholar] [PubMed]
- Alves, M.G.; Martins, A.D.; Rato, L.; Moreira, P.I.; Socorro, S.; Oliveira, P.F. Molecular mechanisms beyond glucose transport in diabetes-related male infertility. Biochim. Biophys. Acta 2013, 1832, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Tavares, R.S.; Portela, J.M.D.; Sousa, M.I.; Mota, P.C.; Ramalho-Santos, J.; Amaral, S. High glucose levels affect spermatogenesis: An in vitro approach. Reprod. Fertil. Dev. 2017, 29, 1369–1378. [Google Scholar] [CrossRef]
- Mita, M.; Hall, P.F. Metabolism of round spermatids from rats: Lactate as the preferred substrate. Biol. Reprod. 1982, 26, 445–455. [Google Scholar] [CrossRef]
- Ding, G.-L.; Liu, Y.; Liu, M.-E.; Pan, J.-X.; Guo, M.-X.; Sheng, J.-Z.; Huang, H.-F. The effects of diabetes on male fertility and epigenetic regulation during spermatogenesis. Asian J. Androl. 2015, 17, 948–953. [Google Scholar]
- Oliveira, P.F.; Alves, M.G.; Rato, L.; Laurentino, S.; Silva, J.; Sá, R.; Barros, A.; Sousa, M.; Carvalho, R.A.; Cavaco, J.E.; et al. Effect of insulin deprivation on metabolism and metabolism-associated gene transcript levels of in vitro cultured human sertoli cells. Biochim. Biophys. Acta 2012, 1820, 84–89. [Google Scholar] [CrossRef]
- Riera, M.F.; Galardo, M.N.; Pellizzari, E.H.; Meroni, S.B.; Cigorraga, S.B. Molecular mechanisms involved in sertoli cell adaptation to glucose deprivation. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E907–E914. [Google Scholar] [CrossRef]
- Cooper, T.G.; Noonan, E.; Von Eckardstein, S.; Auger, J.; Baker, H.; Behre, H.M.; Haugen, T.B.; Kruger, T.; Wang, C.; Mbizvo, M.T. World health organization reference values for human semen characteristics. Hum. Reprod. Update 2010, 16, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Schloss, J.; Colosimo, M.; Vitetta, L. New insights into potential prevention and management options for chemotherapy-induced peripheral neuropathy. Asia Pac. J. Oncol. Nurs. 2016, 3, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Cascella, M.; Muzio, M.R. Potential application of the kampo medicine goshajinkigan for prevention of chemotherapy-induced peripheral neuropathy. J. Integr. Med. 2017, 15, 77–87. [Google Scholar] [CrossRef]
- Takahashi, K.; Nagahori, K.; Qu, N.; Kuramasu, M.; Hirayanagi, Y.; Hayashi, S.; Ogawa, Y.; Hatayama, N.; Terayama, H.; Suyama, K.; et al. The effectiveness of traditional japanese medicine goshajinkigan in irradiation-induced aspermatogenesis in mice. BMC Complementary Altern. Med. 2019, 19, 362. [Google Scholar] [CrossRef] [PubMed]
- Zang, Z.J.; Ji, S.Y.; Dong, W.; Zhang, Y.N.; Zhang, E.H.; Bin, Z. A herbal medicine, saikokaryukotsuboreito, improves serum testosterone levels and affects sexual behavior in old male mice. Aging Male 2015, 18, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, J.; Liu, X.; Wang, D.; Zheng, P.; Qi, A.; Yi, T.; Li, S. Jian-pi-yi-shen formula ameliorates chronic kidney disease: Involvement of mitochondrial quality control network. BMC Complementary Altern. Med. 2018, 18, 340. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, S.; Luo, L.; Liu, Y.; Li, E.; Zhu, Z.; Zhao, Z. Shengjing capsule improves spermatogenesis through upregulating integrin α6/β1 in the noa rats. Evid.-Based Complementary Altern. Med. 2019, 2019, 8494567. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, L.; Kang, R.; Li, F.; Li, E.; Zhang, T.; Luo, J.; Zhao, Z. Shengjing capsule improves erectile function through regulation of nitric oxide-induced relaxation in corpus cavernosum smooth muscle in a castrated rat model. Urology 2016, 91, 243.e7–243.e12. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, S.; Shangguan, J.; Pan, L.; Zhou, X.; Yaghmai, V.; Velichko, Y.; Hu, C.; Yang, J.; Zhang, Z. Mri assessment of associations between brown adipose tissue and cachexia in murine pancreatic ductal adenocarcinoma. Intern. Med. Open Access 2019, 9, 301. [Google Scholar] [CrossRef]
- Zhao, M.; Chan, C.P.S.; Cheung, C.W.C.; Alqawasmeh, O.; Wang, R.C.C.; Wu, J.C.Y.; Lin, Z.X.; Li, T.C.; Chung, J.P.W.; Mak, J.S.M.; et al. A double-blinded, randomized placebo-controlled trial on the effect of traditional chinese medicine formula wuzi yanzong pill on improving semen qualities in men with suboptimal parameters. Trials 2019, 20, 540. [Google Scholar] [CrossRef]
- Melo da Cunha, J.d.S.; Alfredo, T.M.; Dos Santos, J.M.; Alves Junior, V.V.; Rabelo, L.A.; Lima, E.S.; Boleti, A.P.d.A.; Carollo, C.A.; Dos Santos, E.L.; de Picoli Souza, K. Antioxidant, antihyperglycemic, and antidiabetic activity of Apis mellifera bee tea. PLoS ONE 2018, 13, e0197071. [Google Scholar] [CrossRef] [PubMed]
- Balen, A.H.; Morley, L.C.; Misso, M.; Franks, S.; Legro, R.S.; Wijeyaratne, C.N.; Stener-Victorin, E.; Fauser, B.C.; Norman, R.J.; Teede, H. The management of anovulatory infertility in women with polycystic ovary syndrome: An analysis of the evidence to support the development of global who guidance. Hum Reprod. Update 2016, 22, 687–708. [Google Scholar] [CrossRef] [PubMed]
- Lienou, L.; Telefo, P.; Njimou, J.; Nangue, C.; Bayala, B.; Goka, S.; Biapa, P.; Yemele, M.; Donfack, N.; Mbemya, J.J. Effect of the aqueous extract of Senecio biafrae (oliv. & hiern) J. Moore on some fertility parameters in immature female rat. J. Ethnopharmacol. 2015, 161, 156–162. [Google Scholar]
- Mishra, R.K.; Singh, S.; Singh, S.K. Natural products in regulation of male fertility. Indian J. Med. Res. 2018, 148, S107–S114. [Google Scholar] [PubMed]
- Bajaj, V.K.; Gupta, R.S. Review on research of suppression male fertility and male contraceptive drug development by natural products. Nat. Prod. Commun. 2013, 8, 1934578X1300800840. [Google Scholar] [CrossRef]
- Kumar, D.; Kumar, A.; Prakash, O. Potential antifertility agents from plants: A comprehensive review. J. Ethnopharmacol. 2012, 140, 1–32. [Google Scholar] [CrossRef]
- Kolangi, F.; Shafi, H.; Memariani, Z.; Kamalinejad, M.; Bioos, S.; Jorsaraei, S.G.A.; Bijani, A.; Shirafkan, H.; Mozaffarpur, S.A. Effect of Alpinia officinarum hance rhizome extract on spermatogram factors in men with idiopathic infertility: A prospective double-blinded randomised clinical trial. Andrologia 2019, 51, e13172. [Google Scholar] [CrossRef]
- Nasimi Doost Azgomi, R.; Nazemiyeh, H.; Sadeghi Bazargani, H.; Fazljou, S.M.B.; Nejatbakhsh, F.; Moini Jazani, A.; Ahmadi AsrBadr, Y.; Zomorrodi, A. Comparative evaluation of the effects of Withania somnifera with pentoxifylline on the sperm parameters in idiopathic male infertility: A triple-blind randomised clinical trial. Andrologia 2018, 50, e13041. [Google Scholar] [CrossRef]
- Wu, M.; Yin, Y.; Zhao, M.; Hu, L.; Chen, Q. The low expression of leukemia inhibitory factor in endometrium: Possible relevant to unexplained infertility with multiple implantation failures. Cytokine 2013, 62, 334–339. [Google Scholar] [CrossRef]
- Sharma, R.K.; Agarwal, A. Role of reactive oxygen species in male infertility. Urology 1996, 48, 835–850. [Google Scholar] [CrossRef]
- Kucera, M.; Stepan Ml, J.; Stelcl, M. Possibilities and real meaning of assessment of ovarian reserve. Ceska Gynekol. 2018, 83, 307–311. [Google Scholar] [PubMed]
- Gleicher, N.; Weghofer, A.; Barad, D.H. Discordances between follicle stimulating hormone (fsh) and anti-mullerian hormone (amh) in female infertility. Reprod. Biol. Endocrinol. 2010, 8, 64. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Compound/Extract | System | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|
| Plant | Date palm pollen extract | In vitro | Phoenix dactylifera Linn. | Sertoli cells, spermatogonial stem cells from mice | 0.06, 0.25, 0.62 mg/mL; 14 days | Increase of proliferation of spermatogonia | [40] | |
| Plant | 5H-purin-6-amine, Sedum sarmentosum extract | In vitro | Sedum sarmentosum | Spermatogonial stem cells C57BL. 6-TG-EGFP | 0.01, 0.1, 1, 10 mg/mL; 1 week | Increase of self-renewal in SSCs | ↑ PLZF, GFRα1, VASA, Lhx1 ↓ Pgk2 | [41] |
| Plant | Licorice extract | In vitro | Glycyrrhiza uralensis Fisch. | Testis tissue from C57BL/6N mice | 0.2, 2, 20 μmol/L; 72 h | Increase of proliferation of spermatogonia | ↑ PCNA, SCP3, Spo11 | [42] |
| Plant | Lycium barbarum polysaccharide | In vitro | Lycium barbarum Linn. | Leydig MLTC-1 | 50 μg/mL; 48 h | Increase of cell viability | ↑ Testosterone, ↓ p-PERK/PERK, p-elF2α/elF2α, ATF4/β-actin, apoptosis rate, LC3II/I, Atg5/β-actin | [43] |
| Plant | Morindae radix aqueous extract | In vitro | Morinda officinalis | TM3 cells, mouse Leydig cells | 10, 50, 100, 250 mg/mL; 24 h | Increase of testosterone production. Decrease of H2O2 induced cytotoxicity, and lipid peroxidation | ↑ SOD, CAT ↓ MDA | [44] |
| Plant | Taraxacum officinale aqueous extract | In vitro | Taraxacum officinale | TM3, ATCCNoCRL | 1, 10, 25, 50 mg/mL; 12, 48 h | Increase of the levels of steroidogenic enzymes | ↑ STAR, CYP11A1, CYP17A1 | [11] |
| Plant | Typha capensis rhizome extract F1 fraction | In vitro | Typha capensis (Rohrb.) N.E.Br. | TM3-Leydig cells | 10, 100 μg/mL; 96 h | ↑ Testosterone | [45] |
| Classification | Compound/Extract | System | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|
| Plant | Acacia hydaspica ethyl acetate extract | In vivo | Acacia hydaspica R. Parker | SD rats | 400 mg/kg; 21 days | Increase of seminiferous tubule diameter, area, epithelial height Decrease of width of tubular lumen, interstitial space, DNA damage | ↑ Testosterone, LH, FSH, SOD, POD, CAT, QR, GSH, GR, GST, GPx, γ-GT ↓ H2O2, NO, MDA | [46] |
| Plant | Achillea millefolium inflorescences ethanol extract | In vivo | Achillea millefolium Linn. | Wistar mice | 120 mg/kg; 48 days | Increase of sperm motility, capsule thickness, epithelial thickness, tubule differentiation index | ↑ SOD, LH ↓ LDH, NO, MDA | [47] |
| Plant | Aged garlic extract | In vivo | Allium sativum for. pekinense MAKINO | Wistar mice | 250 mg/kg; 14 days | Increase of testis weight, sperm count, motility, recovery of seminiferous tubules Decrease of the death of sperm, sperm abnormality | ↑ Testosterone, GSH, GSH-Px, CAT, SOD ↓ MDA | [10] |
| Plant | Angelica keiskei powder | In vivo | Angelica keiskei Koidz. | Self-breeding CD-1 mice | 57.5 mg/kg; 7 days | Increase of density of sperm, motility, motile sperm density, progressive sperm velocity, progressive sperm density Decrease of abnormal seminiferous tubules, DNA fragmentation | ↑ GSS, HO-1, Hspa11, Hspa2 Hsf1, Hsf2 | [1] |
| Plant | Banaba leaf and ginseng extract | In vivo | Lagerstroemia speciosa (L.) Pers. | Swiss mice | 150 mg/kg; 30 days | Increase of testis weight, epididymis weight, seminal vesicle weight, sperm density, sperm viability, progressive sperm motility Decrease of nonprogressive sperm motility, abnormal sperm morphology (head, tail, twisted body) | ↑ Testis glycogen, testis protein, testis fructose, seminal vesicle fructose, epididymal fructose, testis protein, seminal vesicle protein, epididymal protein ↓ Testis cholesterol | [37] |
| Plant | Bajijiasu | In vivo | Morinda officinalis F.C. |
kidney-yang-deficient Kunming mouse |
| Increase of the sexual behavior Decrease of DNA damage of sperm by H2O2 | ↑ Testosterone, ↓ Cortisol, SOD, GPx, CAT, MAD | [6] |
| Plant | Balanites aegyptiaca sapogenin extract | In vivo | Balanites aegyptiaca | Albino rats | 25, 50, 100 mg/kg; 70 days | Increase of semen quality | ↑ LH, Estradiol, Testosterone, Glucose ↓ FSH, Cholesterol, sAST, urea, creatinine | [48] |
| Plant | Bee bread | In vivo | Bee pollen | SD rats | 0.5 g/kg; 28 days | Increase in the weight of the prostate gland in adult rats | ↑ Arginine, L-carnitine, glutathione | [49] |
| Plant | Cistanche tubulosa extract | In vivo | Cistanche tubulosa (Schrenk) R. wight | SD rats | 200 mg/kg; 42 days | Increase of daily sperm production Decrease of abnormal morphology (isolated head, head without curvature), immobile sperm | ↑ LDH-x activity, testosterone, 3β-HSD, 17β-HSD, CYP17A1, CYP11A1, StAR | [50] |
| Plant | Crocus sativus L. aqueous extract | In vivo | Crocus sativus Linn. | Wistar rats | 100 mg/kg; one time/2 days; 16 days | Increase of mean sperm number | ↑ Testosterone ↓LPO | [27] |
| Plant | Curcumin | In vivo | Turmeric | SD rats | 100 mg/kg; 3 days | Decrease of the toxic effects of CdCl2 | ↑ GSH, CAT, GPx, SOD ↓ TBARS | [51] |
| Plant | Curcumin extract | In vivo | Curcuma longa Linn. | Bovine semen breeding bulls | 5, 10, 50, 100 µM/L; 24 h | Increase of spermatozoa activity and protection | ↓ Nitroblue-tetrazolium | [52] |
| Plant | Cymbopogon citrates aqueous extract | In vivo | Cymbopogon citratus | SD rats | 100 mg/kg; 30 days | Decrease of H2O2 induced reproductive system injury | ↑ GSH ↓ MAD | [4] |
| Plant | Date palm pollen extract | In vitro | Phoenix dactylifera Linn. | Sertoli cells, spermatogonial stem cells from mice | 0.06, 0.25, 0.62 mg/mL; 14 days | Increase of number of spermatogonial colony | [40] | |
| Plant | Date Palm Pollen extract | In vivo | Phoenix dactylifera Linn. | Wistar mice | 150 mg/kg; 56 days | Decrease of testicular dysfunction | ↑ LH, FSH, Testosterone, 3β-HSD, 17β-HSD Estradiol | [53] |
| Plant | Diallyl sulfide (DAS) | In vivo | Allium sativum Linn. | Swiss albino rats | 200 mg/kg; 49 days | Increase of sperm counts, weight of testis, epididymis, spermatogenesis | ↑ Testosterone, estradiol, CYP19, SOD, GSH ↓ MDA, NO | [54] |
| Plant | E. amoenum distillate | In vivo | Echium amoenum | Mus musculus mice | 150 ± 2.5, 75 ± 1.25 mL/kg; 3 weeks | Improvement of hormonal and sperm parameters | ↑ FSH, LH, Testosterone, Leydig cells | [34] |
| Plant | Echinacea purpurea extract | In vivo | Echinacea purpurea Linn. | Albino rats | 63 mg/kg; 4 weeks | Improvement of sperm parameters in the oxidative stress | ↑SOD, GST calcium ion ↓MDA, NO | [26] |
| Plant | Echinacoside | In vivo | Cistanche tubulosa Hook f. II. | Kunming mice | 5, 20, 80 mg/kg; 14 days | Increase of epididymal sperm count, sperm motility | ↑ LH, CYP11A1, CYP17A1, HSD3β1/2, HSD17β, StAR, LHβ, LHr, Gnrh 1, Gnrhr ↓ Hypothalamic AR in nuclei | [55] |
| Plant | Echinacoside | In vivo | Cistanche tubulosa (Schrenk) R. wight | SD rats | 6 mg/kg; 42 days | Increase of sperm number in testis, daily sperm production Decrease of abnormal morphology (isolated head, head without curvature), immobile sperm | ↑ LDH-x activity, testosterone, LH, FSH, 3β-HSD, 17β-HSD, CYP17A1, CYP11A1, StAR | [50] |
| Plant | Ethyl pyruvate | In vivo | NO | NMRI mice | 40 mg/kg; 35 days | Decrease of destructive effects of PHZ on sperm parameters, testosterone level, and lipid peroxidation | ↓ MDA | [29] |
| Plant | Eurycoma longifolia extract | In vivo | Eurycoma longifolia | SD rats | 8 mg/kg; 14 days | Increase of testicular function, spermatogenesis, sperm counts, and motility Decrease of the effects of an excessive estrogen state | ↓ Estrogen | [56] |
| Plant | Ginkgo biloba extract | In vivo | Ginkgo biloba Linn. | Wistar mice | 50 mg/kg; one time | Increase of seminiferous tubular diameter, primary spermatocyte number, round spermatid number, Leydig cell number | ↑ Testosterone, FSH ↓ Mitochondrial NAD, plasma TNF-α, plasma IL-1β | [57] |
| Plant | Ginger | In vivo | Zingiber officinale | Wistar rats | 50, 100 mg/kg; 20 days | Increase of sperm healthy parameters | ↑ TAC, LH, FSH ↓ MDA | [58] |
| Plant | Grape seed proanthocyanidin extract | In vivo | Vitis vinifera | Kunming mice | 100, 200, 400 mg/kg; 5 weeks | Decrease of oxidative stress damage in mice testis | ↑ Nrf2, GST, HO1, NQO1, SOD, T-AOC ↓ MDA, 8-OHdG | [59] |
| Plant | Ionidium suffruticosum methanol extract | In vivo | Ionidium suffruticosum (L.) Ging (Violaceae) | Albino rats | 250 mg/kg; 28 days | Increase of sperm count, cauda epididymis sperm motility, body weight, germinal epithelial cell mass, testis weight, sperm vitality Decrease of epidermal sperm agglutination, sperm morphology abnormality (detached tail, fusion of sperm, broken middle piece, detached tail, coiling of flagellum) | ↑ CAT, SOD ↓ MDA | [60] |
| Plant | Jurenia dolomiaea methanol extract | In vivo | Jurenia dolomiaea Boiss. | SD mice | 200, 400 mg/kg; 60 days | Increase of thickness in germinal layers | ↑ SOD, CAT, POD, testosterone, GSH, GST, GPx, GR, ↓ H2O2, TBARS, Nitrate | [61] |
| plant | KH-465 | In vivo | Epimedium koreanum Nakai, Angelica gigas Nakai | SD rats | 200, 400 mg/kg; 4 weeks | Increase of the sperm count and motility | ↑ LH, SOD ↓ 8-OHdG | [5] |
| Plant | Loranthus micranthus aqueous methanol extract | In vivo | Loranthus micranthus Linn. | Wistar rats | 100, 200 mg/kg; 14 days | Increase of testis weight, sperm motility, sperm viability, TSN, seminiferous tubule diameter, Leydig cells count Decrease of sperm abnormality | ↑ Testosterone, FSH, LH, 3β-HSD, 17β-HSD, SOD, CAT, GSH, GSH-Px, GST, Bcl-2 ↓ MDA, LPO | [62] |
| Plant | Lycium barbarum polysaccharide | In vivo | Lycium barbarum, Linn. | ICR mice | 20, 40 mg/kg; 62 days | Increase of testis weight, epididymis weight, testis organ coefficient, epididymis coefficient, sperm count, sperm viability, mating rate, fertility rate, recovery of spermatogonia, recovery of Sertoli cells | ↑ Testosterone, FSH, LH | [63] |
| Plant | Maca capsules | In vivo | Lepidium meyenii Walp. | Empire Breeders mice | 500, 1000 mg/kg; 28 days | Increase of sperm count, sperm motility, seminiferous tubule width, germinal cell layer thickness | ↑ Testosterone, GSH, CAT, SOD ↓ MDA | [64] |
| Plant | Maca extract | In vivo | Lepidium meyenii Walp. | BALB/c mice | 666 mg/kg; 35 days | Increase of sperm motility, sperm count | [65] | |
| Plant | Matricaria chamomilla hydroethanolic extract | In vivo | Matricaria chamomilla | Wistar rats | 200, 500 mg/kg; 30 days | Increase of testosterone, LH Enhancement of sperm counts, motility, viability | ↑ PI3k, Akt | [25] |
| Plant | Mentha spicata aqueous extract | In vivo | Mentha spicata | Albino mice | 40, 100, 400 mg/kg; 1 week | Decrease of ifosfamide induced chromosomal aberration in bone marrow cells of male albino mice | ↓ Ifosfamide | [36] |
| Plant | Carissa opaca leaves (MLC) methanolic extract | In vivo | Carissa opaca leaves | SD rats | 50, 100, 200 mg/kg; 8 weeks | Protective effect against CCl4-induced antioxidant and hormonal dysfunction | ↑ CAT, POD, SOD, GST, GPx, GR, GSH, QR ↓ Triglycerides, cholesterol, HDL, LDL, TBARS, H2O2 | [7] |
| Plant | Moringa oleifera Seed Aqueous Extract | In vivo | Moringa oleifera | Wistar strain albino rats |
| Enhancement of sexual behavior | [66] | |
| Plant | Moringa oleifera Lam. leaf powder | In vivo | Moringa oleifera Lam. | New Zealand White rabbits | 5, 10, 15 g/kg; 12 weeks | Increase of semen volume, sperm count, motility Decrease in abnormal morphology of sperm | ↑ FSH, LH | [17] |
| Plant | Moringa oleifera Lam. extract | In vivo | Moringa oleifera Lam. | SD rats | 400, 800 mg/kg; 2 weeks | Increase of germinal cell layer thickness, diameter of seminiferous tubules, testis weight index, testicular weight Decrease of perivascular fibrosis | ↑ SOD ↓ MDA, HSP70, | [67] |
| Plant |
|
|
| Wister albino rats |
| Increase of androgen, sperm count, and improvement of testicular structure Decrease of testicular lipid accumulation | ↑ Testosterone, DHEA, T/E2 ↓ Leptin, PRL, E2 | [23] |
| Plant | Naringenin | In vivo | Citrus species | SD rats | 40, 80 mg/kg; 10 weeks | Increase of progressive motility, seminiferous tubule lumen volume Decrease of seminiferous epithelium volume | [68] | |
| Plant | Nigella sativa Seed Powder | In vivo | Nigella sativa | Albino rats | 300 mg/kg; 45 days | Increase of testosterone levels | ↑ LH | [69] |
| Plant | Pedalium murex methanol fruit fraction | In vivo | Pedalium murex Linn. | Albino rats | 50, 10 mg/kg; 60 days | Increase of fertility, sperm motility, sperm density, spermatogenesis, germinal cell count, interstitial cell count, spermatid, preleptotene spermatocyte, fibroblast, mature Leydig cell | ↑ LH, FSH, testosterone, cholesterol, glycogen, sialic acid | [70] |
| Plant | Petasites japonicus MeOH extract | In vivo | Petasites japonicus | SSC/ C57BL/6 mice | 0.1, 1, 10 μg/mL; 7 days | Increase of spermatogonial stem cells | ↑ LHX1, GFRα1 | [71] |
| Plant | Phoenix dactylifera date palm pollen extract | In vivo | Phoenix dactylifera | SD rats | 30, 40, 120, 240 mg/kg; 35 days | Increase of sperm count, motility, morphology, DNA quality | ↑ Estrogen, testosterone | [72] |
| Plant | Phyllanthus emblica L. extract | In vivo | Phyllanthus emblica Linn. | SD rats | 50 mg/kg; 42 days | Increase in sperm concentration, testicular size Decrease in sperm head abnormality, acrosome-reacted sperm | ↑ Testosterone, StAR ↓ Corticosterone, MDA, | [73] |
| Plant | Pilea microphylla extract | In vivo | Pilea microphylla (L.) Liebm. | Wistar mice | 50 mg/kg; 10 weeks | Increase of left epididymal sperm count, motility, vitality, and morphology | [74] | |
| Plant | Pistia stratiotes Linn. ethanol extract | In vivo | Pistia stratiotes Linn. | Wistar strain albino mice | 100 mg/kg; 14 days | Increase of sperm motility Decrease of sperm abnormality | [75] | |
| Plant | Rosa damascene aqueous extract | In vivo | Rosa damascena Linn. | NMRI mice | 10, 20, 40 mg/kg; 40 days | Increase of sperm number, sperm motility, sperm viability, rate of normal sperm, testis weight, testis length, testis width, number of Leydig cells | ↑ Testosterone | [33] |
| Plant | Safed musli extract | In vivo | Chlorophytum borivilianum Santapau and Fernandes | Wistar mice | 125, 250 mg/kg; 52 days | Increase of sperm count, mount latency | [76] | |
| plant | Sanrego aqueous extract | In vivo | Lunasia amara | SD rats | 0, 30, 60, 90 mg/kg; 42 days | Increase of sperm number, sperm motility | [77] | |
| Plant | Shilajit water extract | In vivo | Shilajit | Swiss albino mice | 50, 100, 200 mg/kg; 35 days | Increase of weight of testis, epididymis, seminal vesicle, sperm production, sperm motility, sperm concentration, sialic acid concentration in epididymis, fructose concentration in seminal vesicle, libido, male fertility index Decrease of affected seminiferous tubules | ↑ Testosterone, 3β-HSD, 17β-HSD | [78] |
| Plant | Silymarin | In vivo | Silybummarianum seed | Wistar rats | 50, 100, 150 mg/kg; 28 days | Increase of spermatid and spermatozoid cells | ↑ FSH, GnRH, LH, testosterone, GPX, SOD, NE, serotonin, dopamine | [79] |
| Plant | Suruhan ethanol extract | In vivo | Peperomia pellucid L. Kunth | Albino mice | 56, 112, 168 mg/kg; 35 days | Increase of sperm counts, viability, motility and morphology recovery Decrease of blood glucose levels | ↓ Glucose | [80] |
| Plant | Tetracarpidium conophorum leaf extract | In vivo | Tetracarpidium conophorum (Mull. Arg.) Hutch and Dalziel | Wistar mice | 500, 1000 mg/kg; 21 days | Increase of testis weight, sperm concentration, sperm viability, sperm motility, normal chromatin integrity Decrease of total sperm abnormality (tailless head, headless tail, bent tail), abnormal chromatin integrity | ↑ G6PD activity, 3β-HSD, 17β-HSD, testicular glycogen, testicular Zn, Se content, epididymal Zn, Se content, FSH, LH, testosterone ↓ Testicular cholesterol | [81] |
| Plant | Walnut leaf extract | In vivo | Tetracarpidium conophorum (Müll.Arg.) Hutch. and Dalziel | Wistar albino mice | 500, 1000 mg/kg; 21 days | Increase of testis weight, epididymis weight, sperm count, sperm motility, curvilinear velocity, epididymal sperm viability Decrease of sperm abnormality (tailless head, headless tail, bent tail, coiled tail) | ↑ Testosterone, FSH, LH, 3β-HSD, 17β-HSD, G-6PDH, LDH, testicular Zn and Se content, epididymal Se content, testicular glycogen ↓ testicular cholesterol | [82] |
| Plant | Teucrium polium extract | In vivo | Teucrium polium Lam. | Wistar rats | 1 mL/kg; 10 weeks | Increase of sperm motility, sperm count Decrease of sperm abnormality | ↑ Testosterone, FSH, LH, GPx, CAT, SOD ↓ TBARS | [32] |
| Plant | Thymus algeriensis extract | In vivo | Thymus algeriensis Lam. (Boiss. et Reut.) | Wistar rats | 150 mg/kg; 2 weeks | Increase of sperm count, sperm viability, sperm motility, normal sperm morphology Decrease of sperm abnormality, DNA fragmentation | ↑ CAT, SOD, GPx, GSH, GST ↓ LPO | [83] |
| Plant | Tribulus terrestris dry extract | In vivo | Tribulus terrestris Linn. | Swiss albino mice | 11 mg/kg; 14 days | Increase of sperm motility | ↑ RS, TBRAS, SOD, CAT, GPx, GST, testosterone, 17β-HSD | [9] |
| Plant | WTEA | In vivo | White tea | Wistar rats | 1 g/100 mL; 2 months | Increase of sperm concentration, sperm viability, sperm motility Decrease of abnormal sperm morphology | ↑ Insulin sensitivity, glucose tolerance, FRAP ↓ Insulin resistance, testis carbonyl content, testicular antioxidant potential, testicular OS, TBARS | [84] |
| Plant | Xanthoangelol | In vivo | Angelica keiskei Koidz. | Self-breeding CD-1 mice | 3 mg/kg; 7 days | Increase of sperm motility, progressive sperm density, progressive sperm velocity Decrease of abnormal seminiferous tubules | ↑ GSS | [1] |
| Plant | Xanthosoma sagittifolium | In vivo | Xanthosoma sagittifolium Linn. | Wistar mice | 25, 50, 75, 100%, 100 mg/kg; 14 days | Increase of sperm count, sperm motility, sperm livability Decrease of luminal diameter | [85] | |
| Plant | Tetracarpidium conophorum leaf extraxct | In vivo | Tetracarpidium conophorum | Wistar mice | 50, 500, 1000 mg/kg; 21 days | Decrease of oxidative reproductive toxicity | ↑ MDA, GSH | [86] |
| Plant | Zingiber officinale aqueous extract | In vivo | Zingiber officinale | Broiler breeder | 5, 10 %; 64 weeks | Increase of spermatogenesis | ↑ FSH, Testosterone, LH ↓ MDA, TAC | [87] |
| Classification | Compound/Extract | System | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|
| Plant | Echinacea purpurea ethanol extract (encapsulated chitosan/silica nanoparticle) | In vitro and in vivo studies | Echinacea purpurea Linn. | (1) LC540 (2) SD rats | (1) 25 µg/mL; 24 h (2) 279, 465 mg/kg; 7 weeks | (2) Increase of seminiferous tubules diameter, germinal cell layer thickness, area of seminiferous tubules, area of seminiferous lumen, sperm motility, sperm DNA integrity Decrease of sperm abnormality | (1) ↓ NO (2) ↓ TNF-α, IL-1β | [88] |
| Plant | Echinacoside | In vitro and in vivo studies | Cistanche tubulosa (Schrenk) Hook. f. II. | (1) LC-540, TM3 (2) SD rats | (1) 5, 10 μM (2) 160, 320 mg/kg; 6 weeks | (1) Increase of cell viability (2) Increase of sperm number, sperm motility, seminiferous tubule thickness Decrease of sperm abnormality | (1) LC-540, TM3: ↓ Superoxide anion, LC-540: ↑ StAR, CYP11A1, CYP17A1, HSD17β3 ↓ RAGE, NF-κB, H2O2 (2) ↑ LH, KISS1, SIRT1, GPR54, SOCS -3, SOD, CAT ↓ NO, TNF-α, IL-6, superoxide, MDA | [22] |
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Insect | Drone milk | Apis mellifera | SD rats | 110 mg/kg; 5, 10 days | Increase of weight of androgen-sensitive organs (glans penis, seminal vesicle, muscles) | ↑ Testosterone, SLAP | [89] |
| Animal | Gelam Honey | Apis mellifera | SD rats | 1.0 mL/100 g; 60 days | Increase of fertility | ↑ Fructose | [21] |
| Insect | Hydroethanolic extract of Indian propolis | Apismellifera | Swiss albino mice | 400 mg/kg; 4 weeks | Increase of testis weight, sperm count, total motility, spermatozoa with normal head morphology, spermatozoa with normal DNA, number of tubules with complete spermatogenesis, diameter of seminiferous tubule, number of germ cells Decrease of sperm DNA damage, chromatin immaturity, apoptosis in spermatogonial germ cell | ↑ Testosterone, GSH, CAT ↓ MDA, RAD51 | [90] |
| Animal | Spermaurin | Scorpion Scorpio maurus palmatus | (1) Bovine sperm (2) Monkey sperm (3) Mouse oocytes | (1) dilution 1/20; 10 min (2) dilution 1/40; 10 min (3) dilution 1/40; 4 h | Improvement of sperm motility | [91] |
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Fungi | Antrodia cinnamomea ethanol extract | Antrodia cinnamomea Chang. | SD rats | 385, 770, 1540 mg/kg; 5 weeks | Increase of total sperm count, motility rate Decrease of abnormal sperm count, DNA damage in sperm | ↑ LH, testosterone, StAR, CYP11A1, 17β-HSD, SOD ↓ RAGE, GRP-78, H2O2, NO, MDA | [92] |
| Name of Formula | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|
| Gosha-jinki-gan decoction | Rehmanniae radix, Achyranthis radix, Corni officinalis Sieb., Dioscoreae opposita Thunb., Plantaginis semen, Alismatis orientale Juzepzuk., Poria Cocos Wolf., Moutan Radicis Cortex., Cinnamomum verum J.S. Presl., Aconitum napellus | C57BL/6J mice | 5.4%; 150 days | Increase of testis weight, epididymis spermatozoa count, fertility rate, normal seminiferous tubule appearance | ↓ Caspase-8, TLR2, TLR4, F4/80, Fas | [93] |
| MOTILIPERM | Morinda officinalis How, Allium cepa L., Cuscuta chinensis Lamark | SD mice | 100, 200 mg/kg; 4 weeks | Increase of left testis weight, sperm motility, sperm count, epididymis motility, vas deferens count, epididymis count, spermatogenic cell density Decrease of damage of seminiferous tubule | ↓ MDA, testosterone GRP-78, p-IRE1α, p-JNK | [94] |
| Saikokaryukotsuboreito | Bupleurum falcatum L., Pinellia ternata Breitenbach, Cinnamon verum J.Presl, Poria cocos Wolf, Scutellaria baicalensis Georgi, Zizyphus jujuba var. inermis, Crassostrea gigas., Panax ginseng C. A. Mey., Rinoceros spp., Rheum rhabarbarum. L., Zingiber officinale Roscoe | C57BL/6 mice | 300 mg/kg; 30 days | Increase of sperm number, sperm motility | ↑ Testosterone, intertesticular testosterone, SYCP3 | [95] |
| Shengjing capsule | Cornu cervi panto trichum, Cordyceps sinensis, Polygonatum kingianum Coll. Et Hermsl, Panax ginseng C. A. mey, Cuscuta chinesnsis Lam., Lycium barbarum L., Astragalus complanatus R. Brown, Epimedium wushanense TS Ying, Cortex eucommiae, Rosae laevigata Michx., Verbena officinalis L., Rubus chingii Hu, Curculigo orchioides Gaertner, Psoralea corylifolia L. | SPF Wistar mice | 0.45, 0.90, 1.80 g/kg; 56 days | Increase of sperm motility, normal sperm morphology, normal DNA fragmentation rate Decrease of abnormal sperm morphology, abnormal DNA fragmentation rate | ↑ GSH-PX, MDA, SOD, testosterone | [8] |
| Wuzi Yanzong pill | Lycium barbarum L., Cuscuta chinensis Lam., Rubus chingii Hu. Schizandra chinensis (Turcz.) Baill., Plantago asiatica L. | Kunming mice | 1.0 g/kg; 21 days | Increase of testis weight, sperm count, sperm motility | ↑ Testosterone, total antioxidant status, PCNA, ↓ MDA, total oxidant status, OSI | [30] |
| Yi Shen Jian Pi | Cuscuta chinensis, Lycium spp., Schisandra spp., Codonopsis pilosula, Astragalus membranaceus, Citrus reticulate, Bupleurum spp., Cimicifuga foetida, Ligusticum chuanxiong, Carthamus tinctorius | BALB/c mice | 1.35, 2.70 mg/kg; 4 weeks | Increase of sperm quality | [96] |
| Classification | Compound/Extract | System | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|
| Plant | Evodiae Fructus extract | In vitro | Evodia rutaecarpa Benth | CHO-K1, COV434 | 10, 50, 100, 300, 500 µg/mL; 24 h | Decrease of the ovotoxicity | ↑ mTOR, GSK-3β | [28] |
| Plant | Perilla frutescens | In vitro | Perilla frutescens var. acuta Kudo | Ishikawa, JAr | 50 μg/mL; 48 h | Decrease of implantation failure | ↑ LIF, Integrin β3, β5 | [15] |
| Classification | Compound/Extract | System | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|
| Plant | Anthocleista schweinfurthii aqueous extract | In vivo | Anthocleista schweinfurthii | Wistar rats | 200, 300, 400 mg/kg; 28 days | Decrease of oxidative stress in brain | ↑ Glutathione ↓ Malondialdehyde | [97] |
| Plant | Astragalus mongholicus extract | In vivo | Astragalus mongholicus | SPF/ICR mice | 5%; 56 days | Decrease of estrogen-dependent endometrial hyperplasia and ovarian dysfunction | ↑ PPARs, mDECR, estradiol | [35] |
| Plant | Barley Date palm fruits | In vivo | Hordeum vulgarePhoenix dactylifera | Rattus rattus | 10%; 120 days | Increase of the ovarian function and protection from high cholesterol diet | ↑ CAT, SOD, GST ↓ MDA | [98] |
| Plant | Cinnamon powder | In vivo | Cinnamomum verum | C57BL/6 mice | 10 mg/100 g; 20 days | Increase of insulin resistance | ↑ FSH, IGFBP-1 ↓ Insulin, IGF-1 | [99] |
| Plant | Cyperus rotundus Water extract | In vivo | Cyperus rotundus | C57BL/6 mice | 31.68 mg/kg; 7 days | Increase of cell adhesion and implantation of blastocysts | ↑ LIF, LIF-dependent integrin αV, β3, β5 | [12] |
| Plant | Eucalyptus robusta leaf extraxct | In vivo | Eucalyptus robusta | Wistar rats | 25 mg/kg; 5 days | Decrease of endometritis | ↑ COX-1, COX-2 ↓ TLR-4, TLR-9, Myleoperoxidase, iNOS, NO, SAA | [31] |
| Plant | Ficus asperifolia aqueous extract | In vivo | Ficus asperifolia (L) Hook. Ex Miq (Moraceae) | Wistar rats | 100 mg/kg once a day; 1 week (set I) or 4 weeks (set II) | Increase of conducive condition maintenance | ↑ HDL cholesterol ↓ Total plasma cholesterol, LDL cholesterol | [100] |
| Plant | Ficus vogelii aqueous extract | In vivo | Ficus vogelii | Wistar rats | 100, 300 mg/kg; 21 days | Decrease of lead reproductive toxicity | ↓ SOD | [13] |
| Plant | Milicia excelsa Aqueous extract | In vivo | Milicia excelsa | Wistar rats | 14, 7, 140 mg/kg; 7, 15 days | Decrease of the problems of amenorrhea | ↑ FSH, Estradiol | [18] |
| Plant | Schisandra chinensis extract | In vivo | Schisandra chinensis | SD rats | 200 mg/kg; 7 days | ↓ PRL | [101] | |
| Plant | Senecio biafrae (Oliv. and Hiern) J. Moore aqueous extract | In vivo | Senecio biafrae (Oliv. and Hiern) J. Moore | Albino Wistar rats | 8, 32, 64, 128 mg/kg; 20 days | Increase of the fertility parameters, uterine weight | ↑ FSH, LH, estradiol, progesterone | [20] |
| Plant | Soy isoflavones | In vivo | Glycine max | Rattus norvegicus albinus | 150 mg/kg; 30 days | Increase of insulin sensitivity | ↑ SLRP | [101] |
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Animal | Royal jelly | Apis mellifera | Wistar rats | 100, 200, 400 mg/kg; 14 days | Increase of ovarian hormones and folliculogenesis | ↑ FRAP, progesterone, estradiol ↓ NO | [102] |
| Gender | Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|---|
| Male | Plant | Bajijiasu | Morinda officinalis F.C. | Human sperm DNA | 0.50, 1.00, 2.00 mg/mL; 45 min | ↑ CAT, GSH-Px, SOD ↓ MDA | [6] | |
| Male | Plant | Inositol | Wholemeal cereals, citrus fruits, brewer’s yeast | Seminal fluid from healthy subjects and from subjects with oligoasthenoteratospermia | 2 mg/mL; 30 min, 1, 2 h | Increase of spermatozoa intermediate tract thickness Decrease of amorphous materials, damage in mitochondrial cristae | [103] | |
| Male | Animal | Spermaurin | Sorpio maurus plamatus | Human sperm | Dilution 1/40; 10 min | Increase of sperm motility | [91] | |
| Male | Plant | Tribulus terrestris extract | Tribulus terrestris Linn. | Human sperm | 40, 50 mg/mL; 120 min | Increase of total sperm motility, number of progressive motile spermatozoa, curvilinear velocity | [104] | |
| Male | Plant | Tribulus terrestris extract | Tribulus terrestris Linn. | Human sperm | 20, 40, 50 μg/mL; 120 min | Increase of sperm viability, progressive motile spermatozoa | [105] | |
| Male | Insect | Chilean propolis ethanol extract | Apis mellifera | Normozoospermic semen samples | 6, 12, 25 μg/mL; 1 h | Decrease of intracellular oxidants in spermatozoa and DNA damage | ↓ TBARS, LDH | [2] |
| Female | Plant | Low-dose aspirin in combination with Tiao Jing Cu Yun pills | Aspirin | PCOS patients | Enteric-coated aspirin tablets: 100 mg/day; until the day of ovulation | Increase of perifollicular artery blood flow, oocyte quality, rate of clinical pregnancy | ↑ FSH, estradiol, progesterone | [106] |
| Female | Plant | Cinnamon supplements | Cinnamomum cassia | 45 women with PCOS | 1.5 g/d; 6 months | Increase of menstrual cyclicity, insulin resistance | ↑ Insulin sensitivity, HOMA ↓ QUICK-I | [107] |
| Female | Plant | Fenugreek seed extract | Trigonella foenumgraecum L. | 50 premenopausal women diagnosed with PCOS | Two capsules of 500 mg; 90 days | Decrease of ovarian volume, the number of ovarian cysts | ↑ LH, FSH | [108] |
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Plant | Olea hydro-alcoholic extract | Olea europaea | SD rats | 50, 150, 450 mg/kg; 48 days | Decrease of weight of testicle, seminal vesicle, testosterone hormone, sperm count, motility | ↓ LH, 17 β-hydroxy steroid hydrogenase | [109] |
| Plant | Permethrin | Wistar rats | 35 mg/kg; 60 days | Decrease of Leydig cells | [110] | ||
| Plant | Red cultivar Allium cepa methanol extract | Allium cepa Linn. | Wistar rats | 100, 200, 40, 800, 1200 mg/kg; 14 days | Decrease of sperm concentration, sperm motility, sperm mass activity | [111] | |
| Plant | Ricinus communis L. aqueous extract | Ricinus communis L. | Human sperm | 100, 200, 300 mg/mL; 30 min | Decrease of sperm motility | ↓ NCD, HOS | [112] |
| Plant | Spondias Mombin L. ethanol extract | Spondias Mombin L. | Wistar strain rats | 250, 500 mg/kg; 8 weeks | Decrease of spermatogenesis process | ↓ Androgen | [113] |
| Plant | Tripterygium glycoside extract | Tripterygium wilfordii Hook. F. (GTW) | Female SD rats | 37.8, 94.5 mg/kg; 90 days | Decrease of metestrus phase | ↓ ER-α in hypothalamus | [114] |
| Male SD rats | 37.8, 94.5 mg/kg; 90 days | Increase of immature sperm, sperm abnormality rate | ↓ AR in testis and epididymis |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noh, S.; Go, A.; Kim, D.B.; Park, M.; Jeon, H.W.; Kim, B. Role of Antioxidant Natural Products in Management of Infertility: A Review of Their Medicinal Potential. Antioxidants 2020, 9, 957. https://doi.org/10.3390/antiox9100957
Noh S, Go A, Kim DB, Park M, Jeon HW, Kim B. Role of Antioxidant Natural Products in Management of Infertility: A Review of Their Medicinal Potential. Antioxidants. 2020; 9(10):957. https://doi.org/10.3390/antiox9100957
Chicago/Turabian StyleNoh, Seungjin, Ara Go, Da Bin Kim, Minjeong Park, Hee Won Jeon, and Bonglee Kim. 2020. "Role of Antioxidant Natural Products in Management of Infertility: A Review of Their Medicinal Potential" Antioxidants 9, no. 10: 957. https://doi.org/10.3390/antiox9100957
APA StyleNoh, S., Go, A., Kim, D. B., Park, M., Jeon, H. W., & Kim, B. (2020). Role of Antioxidant Natural Products in Management of Infertility: A Review of Their Medicinal Potential. Antioxidants, 9(10), 957. https://doi.org/10.3390/antiox9100957