Vitreous Antioxidants, Degeneration, and Vitreo-Retinopathy: Exploring the Links



Abstract
1. Introduction
Ocular Antioxidants—Protection against Oxidative Damage and Disease
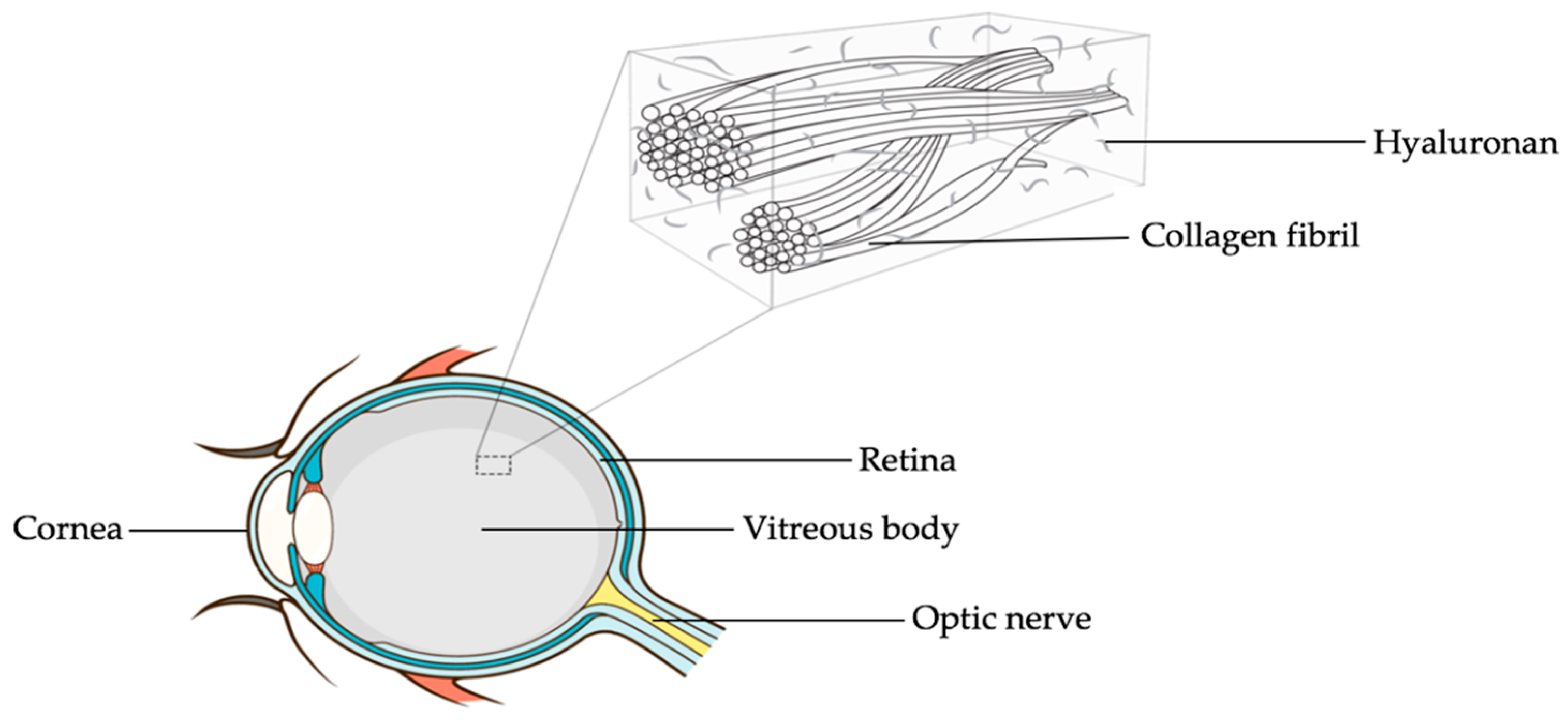
2. The Vitreous Body
A Look at the Vitreous Body and Not just through It
3. Vitreous Degeneration
3.1. Vitreous Liquefaction
3.1.1. Oxidative-Stress-Induced Liquefaction
3.1.2. Enzymatic Liquefaction
3.2. Posterior Vitreous Detachment
4. Vitreous Antioxidant Profile in Health and Disease
4.1. Non-Enzymatic Vitreous Antioxidants
4.1.1. Vitamins
4.1.2. Proteins and Free Amino Acids
4.1.3. Trace Elements: Two Trace Elements Detected in Human Vitreous Are Selenium and Zinc
4.1.4. Uric Acid
4.2. Enzymatic Vitreous Antioxidants
5. Conclusions and Future Strategies
Author Contributions
Funding
Conflicts of Interest
References
- Kolb, H. How the retina works: Much of the construction of an image takes place in the retina itself through the use of specialized neural circuits. Am. Sci. 2003, 91, 28–35. [Google Scholar] [CrossRef]
- Sacca, S.C.; Roszkowska, A.M.; Izzotti, A. Environmental light and endogenous antioxidants as the main determinants of non-cancer ocular diseases. Mutat. Res. 2013, 752, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Riley, P.A. Free radicals in biology: Oxidative stress and the effects of ionizing radiation. Int. J. Radiat. Biol. 1994, 65, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, H.; Halliwell, B. Damage to DNA by reactive oxygen and nitrogen species: Role in inflammatory disease and progression to cancer. Biochem. J. 1996, 313, 17. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M. The definition and measurement of antioxidants in biological systems. Free Radic. Biol. Med. 1995, 18, 125–126. [Google Scholar] [CrossRef]
- Sies, H. Introductory Remarks in Oxidative Stress; Academic Press: London, UK, 1985. [Google Scholar]
- Lazo-de-la-Vega, M.-L.; Fernández-Mejía, C. Oxidative stress in diabetes mellitus and the role of vitamins with antioxidant actions. In Oxidative Stress and Chronic Degenerative Diseases—A Role for Antioxidants; IntechOpen: London, UK, 2013. [Google Scholar]
- Suzuki, M.; Tsujikawa, M.; Itabe, H.; Du, Z.-J.; Xie, P.; Matsumura, N.; Fu, X.; Zhang, R.; Sonoda, K.-H.; Egashira, K. Chronic photo-oxidative stress and subsequent MCP-1 activation as causative factors for age-related macular degeneration. J. Cell. Sci. 2012, 125, 2407–2415. [Google Scholar] [CrossRef]
- Jurkunas, U.V.; Bitar, M.S.; Funaki, T.; Azizi, B. Evidence of oxidative stress in the pathogenesis of fuchs endothelial corneal dystrophy. Am. J. Path. 2010, 177, 2278–2289. [Google Scholar] [CrossRef]
- Halliwell, B.; Murcia, M.A.; Chirico, S.; Aruoma, O.I. Free radicals and antioxidants in food and in vivo: What they do and how they work. Crit. Rev. Food Sci. Nutr. 1995, 35, 7–20. [Google Scholar] [CrossRef]
- Ratnam, D.V.; Ankola, D.D.; Bhardwaj, V.; Sahana, D.K.; Kumar, M.N.V.R. Role of antioxidants in prophylaxis and therapy: A pharmaceutical perspective. J. Control. Release 2006, 113, 189–207. [Google Scholar] [CrossRef]
- Akuffo, K.O.; Beatty, S.; Peto, T.; Stack, J.; Stringham, J.; Kelly, D.; Leung, I.; Corcoran, L.; Nolan, J.M. The Impact of Supplemental Antioxidants on Visual Function in Nonadvanced Age-Related Macular Degeneration: A Head-to-Head Randomized Clinical Trial. Investig. Ophthalmol. Vis. Sci. 2017, 58, 5347–5360. [Google Scholar] [CrossRef]
- World Health Organization. Declaration of Alma-Ata: International Conference on Primary Health Care, Alma-Ata, USSR, 6–12 September 1978; World Health Organization: Geneva, Switzerland, 1978. [Google Scholar]
- Sebag, J. The diagnosis of health. Prev. Med. 1979, 8, 76–88. [Google Scholar] [CrossRef]
- di Sarsina, P.R.; Alivia, M.; Guadagni, P. Widening the paradigm in medicine and health: Person-centred medicine as the common ground of traditional, complementary, alternative and non-conventional medicine. In Healthcare Overview; Springer: Berlin/Heidelberg, Germany, 2012; pp. 335–353. [Google Scholar]
- Rugnetta, M.; Kramer, W. Paving the Way for Personalized Medicine: Facilitating Inter-Agency Coordination for the Integration. Sci. Prog. 2009. Available online: https://www.scienceprogress.org/wp-content/uploads/2009/09/personalized_medicine.pdf (accessed on 16 August 2019).
- Cabrera, M.P.; Chihuailaf, R.H. Antioxidants and the integrity of ocular tissues. Vet. Med. Int. 2011, 2011, 905153. [Google Scholar] [CrossRef] [PubMed]
- Rose, R.C.; Gogia, R.; Richer, S.P. Properties of electrochemically active components in mammalian vitreous humor. Exp. Eye Res. 1997, 64, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Bishop, P.N. Structural macromolecules and supramolecular organisation of the vitreous gel. Prog. Retin. Eye Res. 2000, 19, 323–344. [Google Scholar] [CrossRef]
- Foulds, W.S. Is your vitreous really necessary? Eye 1987, 1, 641. [Google Scholar] [CrossRef]
- Sebag, J. Vitreous: In Health and Disease; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Kokavec, J.; Min, S.H.; Tan, M.H.; Gilhotra, J.S.; Newland, H.S.; Durkin, S.R.; Grigg, J.; Casson, R.J. Biochemical analysis of the living human vitreous. Clin. Exp. Ophthalmol. 2016, 44, 597–609. [Google Scholar] [CrossRef]
- Bishop, P. The biochemical structure of mammalian vitreous. Eye 1996, 10, 664. [Google Scholar] [CrossRef]
- Milston, R.; Madigan, M.C.; Sebag, J. Vitreous floaters: Etiology, diagnostics, and management. Surv. Ophthalmol. 2016, 61, 211–227. [Google Scholar] [CrossRef]
- Sebag, J. Macromolecular structure of the corpus vitreus. Prog. Polym. Sci. 1998, 23, 415–446. [Google Scholar] [CrossRef]
- Kita, T.; Sakamoto, T.; Ishibashi, T., II. D. Hyalocytes: Essential Vitreous Cells in Vitreoretinal Health and Disease. In Vitreous; Springer: Berlin/Heidelberg, Germany, 2014; pp. 151–164. [Google Scholar]
- Shui, Y.-B.; Holekamp, N.M.; Kramer, B.C.; Crowley, J.R.; Wilkins, M.A.; Chu, F.; Malone, P.E.; Mangers, S.J.; Hou, J.H.; Siegfried, C.J. The gel state of the vitreous and ascorbate-dependent oxygen consumption: Relationship to the etiology of nuclear cataracts. Arch. Ophthalmol. 2009, 127, 475–482. [Google Scholar] [CrossRef]
- Sa, A.; Elawadi, A.I. Liquefactionof the Vitreous Humor floaters is a Risk Factor for Lens Opacity and Retinal Dysfunction. J. Am. Sci. 2011, 7, 927–936. [Google Scholar]
- Bailey, A.J. Structure, function and ageing of the collagens of the eye. Eye 1987, 1, 175. [Google Scholar] [CrossRef] [PubMed]
- Cunha-Vaz, J.G. The blood–retinal barriers system. Basic concepts and clinical evaluation. Exp. Eye Res. 2004, 78, 715–721. [Google Scholar] [CrossRef]
- Lee, B.; Litt, M.; Buchsbaum, G. Rheology of the vitreous body. Part I: Viscoelasticity of human vitreous. Biorheology 1992, 29, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Walker, F.; Patrick, R.S. Constituent monosaccharides and hexosamine concentration of normal human vitreous humour. Exp. Eye Res. 1967, 6, 227–232. [Google Scholar] [CrossRef]
- Balazs, E.A. Physiology of the vitreous body. In Importance of the Vitreous Body in Retina Surgery with Special Emphasis on Reoperations; Schepens, C.L., Ed.; C.V. Mosby Co.: St. Louis, MO, USA, 1960; pp. 29–57. [Google Scholar]
- Balazs, E.A.; Denlinger, J.L. Aging Changes in the Vitreous. In Aging and Human Visual Function; Alan R. Liss: New York, NY, USA, 1982; Volume 45. [Google Scholar]
- Hultsch, E. The vitreous of the baby owl monkey. A model for rapid and complete gel-liquefaction. Dev. Ophthalmol. 1981, 2, 1–7. [Google Scholar]
- Balazs, E.A. Therapeutic use of hyaluronan. Struct. Chem. 2009, 20, 341–349. [Google Scholar] [CrossRef]
- Balazs, E.A. Amino-sugar-containing macromolecules in the tissues of the eye and ear. In The Amino Sugars, The Chemistry and Biology of Compounds Containing Amino Sugars; Balazs, E.A., Jeanloz, R.W., Eds.; Academic: New York, NY, USA, 1965; Volume IIA, pp. 401–460. [Google Scholar]
- Filas, B.A.; Zhang, Q.; Okamoto, R.J.; Shui, Y.-B.; Beebe, D.C. Enzymatic degradation identifies components responsible for the structural properties of the vitreous body. Investig. Ophthalmol. Vis. Sci. 2014, 55, 55–63. [Google Scholar] [CrossRef]
- Sebag, J.; Yee, K.M.P. Vitreous: From biochemistry to clinical relevance. In Duane’s Foundations of Clinical Ophthalmology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 1–67. [Google Scholar]
- Nguyen, B.Q.; Fife, R.S. Vitreous contains a cartilage-related protein. Exp. Eye Res. 1986, 43, 375–382. [Google Scholar] [CrossRef]
- Sebag, J. Structure, function, and age-related changes of the human vitreous. In The Vitreous and Vitreoretinal Interface; Springer: Berlin/Heidelberg, Germany, 1987; pp. 37–57. [Google Scholar]
- Tozer, K.; Johnson, M.W.; Sebag, J., II. C. Vitreous aging and posterior vitreous detachment. In Vitreous; Springer: Berlin/Heidelberg, Germany, 2014; pp. 131–150. [Google Scholar]
- Foos, R.Y.; Wheeler, N.C. Vitreoretinal juncture: Synchysis senilis and posterior vitreous detachment. Ophthalmology 1982, 89, 1502–1512. [Google Scholar] [CrossRef]
- Worst, J.G.F. Cisternal systems of the full developed vitreous body in the young adult. Trans. Ophthalmol. Soc. UK 1977, 97, 550–554. [Google Scholar] [PubMed]
- Los, L.I.; van der Worp, R.J.; van Luyn, M.J.A.; Hooymans, J.M.M. Age-related liquefaction of the human vitreous body: LM and TEM evaluation of the role of proteoglycans and collagen. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2828–2833. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.W. Perifoveal vitreous detachment and its macular complications. Trans. Am. Ophthalmol. Soc. 2005, 103, 537. [Google Scholar] [CrossRef] [PubMed]
- Sebag, J. Posterior Vitreous Detachment. Ophthalmology 2018, 125, 1384–1385. [Google Scholar] [CrossRef]
- Balazs, E.A. Functional anatomy of the vitreous. In Ocular Anatomy, Embryology and Teratology; Jakobiec, F.A., Ed.; Harper and Row: New York, NY, USA, 1982; pp. 425–440. [Google Scholar]
- Mamou, J.; Wa, C.A.; Yee, K.M.P.; Silverman, R.H.; Ketterling, J.A.; Sadun, A.A.; Sebag, J. Ultrasound-based quantification of vitreous floaters correlates with contrast sensitivity and quality of life. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1611–1617. [Google Scholar] [CrossRef]
- Nguyen, J.H.; Nguyen-Cuu, J.; Yu, F.; Yee, K.M.; Mamou, J.; Silverman, R.H.; Ketterling, J.; Sebag, J. Assessment of Vitreous Structure and Visual Function after Neodymium: Yttrium–Aluminum–Garnet Laser Vitreolysis. Ophthalmology 2019, 126, 1517–1526. [Google Scholar] [CrossRef]
- Nuzzi, R.; Marchese, A.; Gulino, G.R.; Versino, E.; Ghigo, D. Influence of posterior vitreous detachment and type of intraocular lens on lipid peroxidation in the human vitreous. Mol. Vis. 2015, 21, 1106. [Google Scholar]
- Vaughan-Thomas, A.; Gilbert, S.J.; Duance, V.C. Elevated levels of proteolytic enzymes in the aging human vitreous. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3299–3304. [Google Scholar]
- Berra, A.; Ferreira, S.; Stanga, P.; Llesuy, S. Age-related antioxidant capacity of the vitreous and its possible relationship with simultaneous changes in photoreceptors, retinal pigment epithelium and Bruchs’ membrane in human donors’ eyes. Arch. Gerontol. Geriatr. 2002, 34, 371–377. [Google Scholar] [CrossRef]
- Wickham, L.; Karim, A.-K.; Elgohary, M.; Ballagan, K.; Moss, R.; Firth, M.; Firth, G.; Thompson, G. A comparison of water soluble antioxidants in human vitreous humour in retinal ischaemia. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1351. [Google Scholar]
- Mancino, R.; Di Pierro, D.; Varesi, C.; Cerulli, A.; Feraco, A.; Cedrone, C.; Pinazo-Duran, M.D.; Coletta, M.; Nucci, C. Lipid peroxidation and total antioxidant capacity in vitreous, aqueous humor, and blood samples from patients with diabetic retinopathy. Mol. Vis. 2011, 17, 1298. [Google Scholar]
- Harocopos, G.J.; Shui, Y.-B.; McKinnon, M.; Holekamp, N.M.; Gordon, M.O.; Beebe, D.C. Importance of vitreous liquefaction in age-related cataract. Investig. Ophthalmol. Vis. Sci. 2004, 45, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Ueno, N. Changes in vitreous structure caused by oxygen free radicals. Nippon Ganka Gakkai Zasshi 1995, 99, 1342–1360. [Google Scholar] [PubMed]
- Akiba, J.; Ueno, N.; Chakrabarti, B. Mechanisms of photo-induced vitreous liquefaction. Curr. Eye Res. 1994, 13, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Ueno, N.; Sebag, J.; Hirokawa, H.; Chakrabarti, B. Effects of visible-light irradiation on vitreous structure in the presence of a photosensitizer. Exp. Eye Res. 1987, 44, 863–870. [Google Scholar]
- Akiba, J. Photodynamically induced vitreous liquefaction in vivo. Nippon Ganka Gakkai Zasshi 1992, 96, 731–736. [Google Scholar]
- Matsumura, G.; Herp, A.; Pigman, W. Depolymerization of hyaluronic acid by autoxidants and radiations. Radiat. Res. 1966, 28, 735–752. [Google Scholar] [CrossRef]
- Deguine, V.; Menasche, M.; Ferrari, P.; Fraisse, L.; Pouliquen, Y.; Robert, L. Free radical depolymerization of hyaluronan by Maillard reaction products: Role in liquefaction of aging vitreous. Int. J. Biol. Macromol. 1998, 22, 17–22. [Google Scholar] [CrossRef]
- Góes, R.M.; Nader, H.B.; Porcionatto, M.A.; Haddad, A.; Laicine, E.M. Chondroitin sulfate proteoglycans are structural renewable constituents of the rabbit vitreous body. Curr. Eye Res. 2005, 30, 405–413. [Google Scholar] [CrossRef]
- Bishop, P.N.; Holmes, D.F.; Kadler, K.E.; McLeod, D.; Bos, K.J. Age-related changes on the surface of vitreous collagen fibrils. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1041–1046. [Google Scholar] [CrossRef]
- Rangchian, A.; Francone, A.; Farajzadeh, M.; Hosseini, H.; Connelly, K.; Hubschman, J.P.; Kavehpour, P. Effects of Collagenase type II on Vitreous Humor, an in-situ Rheological Study. J. Biomech. Eng. 2019, 141, 081007. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Chen, Y.-S.; Xu, Q.; Hanes, J.; Rupenthal, I.D. Effects of enzymatic degradation on dynamic mechanical properties of the vitreous and intravitreal nanoparticle mobility. Eur. J. Pharm. Sci. 2018, 118, 124–133. [Google Scholar] [CrossRef] [PubMed]
- van Deemter, M.; Kuijer, R.; Pas, H.H.; van der Worp, R.J.; Hooymans, J.M.M.; Los, L.I. Trypsin-mediated enzymatic degradation of type II collagen in the human vitreous. Mol. Vis. 2013, 19, 1591. [Google Scholar] [PubMed]
- Wang, Z.-L.; Zhang, X.; Xu, X.; Sun, X.-D.; Wang, F. PVD following plasmin but not hyaluronidase: Implications for combination pharmacologic vitreolysis therapy. Retina 2005, 25, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Hikichi, T.; Kado, M.; Yoshida, A. Intravitreal injection of hyaluronidase cannot induce posterior vitreous detachment in the rabbit. Retina (Phila. Pa) 2000, 20, 195–198. [Google Scholar] [CrossRef]
- Napolitano, F.; Di Iorio, V.; Di Iorio, G.; Melone, M.A.B.; Gianfrancesco, F.; Simonelli, F.; Esposito, T.; Testa, F.; Sampaolo, S. Early posterior vitreous detachment is associated with LAMA5 dominant mutation. Ophthalmic Genet. 2019, 40, 39–42. [Google Scholar] [CrossRef]
- Chuo, J.Y.; Lee, T.Y.Y.; Hollands, H.; Morris, A.H.; Reyes, R.C.; Rossiter, J.D.; Meredith, S.P.; Maberley, D.A.L. Risk factors for posterior vitreous detachment: A case-control study. Am. J. Ophthalmol. 2006, 142, 931–937. [Google Scholar] [CrossRef]
- Akiba, J. Prevalence of posterior vitreous detachment in high myopia. Ophthalmology 1993, 100, 1384–1388. [Google Scholar] [CrossRef]
- Morita, H.; Funata, M.; Tokoro, T. A clinical study of the development of posterior vitreous detachment in high myopia. Retina (Phila. Pa) 1995, 15, 117–124. [Google Scholar] [CrossRef]
- Sebag, J. Anomalous posterior vitreous detachment: A unifying concept in vitreo-retinal disease. Graefe’s Arch. Clin. Exp. Ophthalmol. 2004, 242, 690–698. [Google Scholar] [CrossRef]
- Sebag, J. Vitreous Anatomy, Aging, and Anomalous Posterior Vitreous Detachment. In Encyclopedia of the Eye; Darlene, D.A., Joseph, B.C., Reza, D., Eds.; Elsevier: Oxford, UK, 2010; Volume 4, pp. 307–315. [Google Scholar]
- Sebag, J. Diabetic vitreopathy. Ophthalmology 1996, 103, 205–206. [Google Scholar] [CrossRef]
- Snead, M.P.; Yates, J.R.W. Clinical and molecular genetics of Stickler syndrome. J. Med. Genet. 1999, 36, 353–359. [Google Scholar] [PubMed]
- Badrinath, S.S.; Gopal, L.; Sharma, T.; Parikh, S.; Shanmugam, M.P.; Bhende, P.; Biswas, J. Vitreoschisis in Eales’ disease: Pathogenic role and significance in surgery. Retina (Phila. Pa) 1999, 19, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Gale, J.; Aiello, L.P.; Sebag, J. IE Diabetic Vitreopathy. In Vitreous; Springer: Berlin/Heidelberg, Germany, 2014; pp. 57–79. [Google Scholar]
- Sebag, J.; Niemeyer, M.; Koss, M.J., III. B. Anomalous Posterior Vitreous Detachment and Vitreoschisis. In Vitreous; Springer: Berlin/Heidelberg, Germany, 2014; pp. 241–263. [Google Scholar]
- Sebag, J. Vitreous: The resplendent enigma. Br. J. Ophthalmol. 2009, 93, 989–991. [Google Scholar] [CrossRef] [PubMed]
- Sebag, J. Vitreoschisis. Graefe’s Arch. Clin. Exp. Ophthalmol. 2008, 246, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Dunker, S.; Sadun, A.A.; Sebag, J. Neuron specific enolase in retinal detachment. Curr. Eye Res. 2001, 23, 382–385. [Google Scholar] [CrossRef] [PubMed]
- Ko, G.J.; Chu, Q.; Garg, S.; Sanchez, R.; Sadun, A.A.; Sebag, J.; Lam, T.T. Neuron-specific enolase in cell damage after retinal ischemia-reperfusion injury in rats. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3614. [Google Scholar]
- Jalkh, A.; Takahashi, M.; Topilow, H.W.; Trempe, C.L.; McMeel, J.W. Prognostic value of vitreous findings in diabetic retinopathy. Arch. Ophthalmol. 1982, 100, 432–434. [Google Scholar] [CrossRef]
- Nasrallah, F.P.; Jalkh, A.E.; Van Coppenolle, F.; Kado, M.; Trempe, C.L.; McMeel, J.W.; Schepens, C.L. The role of the vitreous in diabetic macular edema. Ophthalmology 1988, 95, 1335–1339. [Google Scholar] [CrossRef]
- Nasrallah, F.P.; Van De Velde, F.; Jalkh, A.E.; Trempe, C.L.; McMeel, J.W.; Schepens, C.L. Importance of the vitreous in young diabetics with macular edema. Ophthalmology 1989, 96, 1511–1517. [Google Scholar] [CrossRef]
- Krebs, I.; Brannath, W.; Glittenberg, C.; Zeiler, F.; Sebag, J.; Binder, S. Posterior vitreomacular adhesion: A potential risk factor for exudative age-related macular degeneration? Am. J. Ophthalmol. 2007, 144, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Robison, C.D.; Krebs, I.; Binder, S.; Barbazetto, I.A.; Kotsolis, A.I.; Yannuzzi, L.A.; Sadun, A.A.; Sebag, J. Vitreomacular adhesion in active and end-stage age-related macular degeneration. Am. J. Ophthalmol. 2009, 148, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Krebs, I.; Glittenberg, C.; Zeiler, F.; Binder, S. Spectral domain optical coherence tomography for higher precision in the evaluation of vitreoretinal adhesions in exudative age-related macular degeneration. Br. J. Ophthalmol. 2011, 95, 1415–1418. [Google Scholar] [CrossRef] [PubMed]
- Sebag, J. Vitreous in AMD therapy–the medium is the message (Guest Editorial). Retina 2015, 35, 1715–1718. [Google Scholar] [CrossRef]
- McMeel, J.W.; Pavan, P.R.; Avila, M.P.; Jalkh, A.E. The diabetic vitreous. Int. Ophthalmol. Clin. 1984, 24, 31–45. [Google Scholar]
- Crouch, R.; Priest, D.G.; Duke, E.J. Superoxide dismutase activities of bovine ocular tissues. Exp. Eye Res. 1978, 27, 503–509. [Google Scholar] [CrossRef]
- Ohia, S.E.; Opere, C.A.; LeDay, A.M. Pharmacological consequences of oxidative stress in ocular tissues. Mutat. Res. 2005, 579, 22–36. [Google Scholar] [CrossRef]
- Mirończuk-Chodakowska, I.; Witkowska, A.M.; Zujko, M.E. Endogenous non-enzymatic antioxidants in the human body. Adv. Med. Sci. 2018, 63, 68–78. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Burns, J.J. Introduction: Overview of ascorbic acid metabolism. Ann. N. Y. Acad. Sci. 1975, 258, 5–6. [Google Scholar] [CrossRef]
- Socci, R.; Delamere, N. Characteristics of ascorbic acid uptake by the iris-ciliary body. In Proceedings of ARVO Abstracts. Investig. Ophthalmol. Vis. Sci. 1987, 28, 74. [Google Scholar]
- Duarte, T.L.; Lunec, J. Review: When is an antioxidant not an antioxidant? A review of novel actions and reactions of vitamin C. Free Radic. Res. 2005, 39, 671–686. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Ghim, W.; Oh, S.; Kim, Y.; Park, U.C.; Kang, J.; Yu, H.G. Association of vitreous vitamin C depletion with diabetic macular ischemia in proliferative diabetic retinopathy. PLoS ONE 2019, 14, e0218433. [Google Scholar] [CrossRef] [PubMed]
- Hanashima, C.; Namiki, H. Reduced viability of vascular endothelial cells by high concentration of ascorbic acid in vitreous humor. Cell. Biol. Int. 1999, 23, 287–298. [Google Scholar] [CrossRef]
- Sommer, F.; Kobuch, K.; Brandl, F.; Wild, B.; Framme, C.; Weiser, B.; Tessmar, J.; Gabel, V.P.; Blunk, T.; Goepferich, A. Ascorbic acid modulates proliferation and extracellular matrix accumulation of hyalocytes. Tissue Eng. 2007, 13, 1281–1289. [Google Scholar] [CrossRef]
- Traber, M.G.; Stevens, J.F. Vitamins C and E: Beneficial effects from a mechanistic perspective. Free Radic. Biol. Med. 2011, 51, 1000–1013. [Google Scholar] [CrossRef]
- Philpot, F.J.; Pirie, A. Riboflavin and riboflavin adenine dinucleotide in ox ocular tissues. Biochem. J. 1943, 37, 250. [Google Scholar] [CrossRef]
- Long, C. The Biochemists’ Handbook; E. & F. N. Spon Ltd.: London, UK, 1961; p. 1092. [Google Scholar]
- Ashoori, M.; Saedisomeolia, A. Riboflavin (vitamin B2) and oxidative stress: A review. Br. J. Nutr. 2014, 111, 1985–1991. [Google Scholar] [CrossRef]
- Saedisomeolia, A.; Ashoori, M. Riboflavin in human health: A review of current evidences. In Advances in Food and Nutrition Research; Elsevier: Oxford, UK, 2018; Volume 83, pp. 57–81. [Google Scholar]
- Toyosaki, T. Antioxidant effect of riboflavin in enzymic lipid peroxidation. J. Agric. Food Chem. 1992, 40, 1727–1730. [Google Scholar] [CrossRef]
- Matsuki, T.; Yubisui, T.; Tomoda, A.; Yoneyama, Y.; Takeshita, M.; Hirano, M.; Kobayashi, K.; Tani, Y. Acceleration of methaemoglobin reduction by riboflavin in human erythrocytes. Br. J. Haematol. 1978, 39, 523–528. [Google Scholar] [CrossRef]
- Feng, X.; Hultquist, D.E. Coupling of dihydroriboflavin oxidation to the formation of the higher valence states of hemeproteins. Biochem. Biophys. Res. Commun. 1991, 181, 197–203. [Google Scholar] [CrossRef][Green Version]
- Wood, A.W.; Sayer, J.M.; Newmark, H.L.; Yagi, H.; Michaud, D.P.; Jerina, D.M.; Conney, A.H. Mechanism of the inhibition of mutagenicity of a benzo [a] pyrene 7,8-diol 9,10-epoxide by riboflavin 5′-phosphate. Proc. Natl. Acad. Sci. USA 1982, 79, 5122–5126. [Google Scholar] [CrossRef] [PubMed]
- Yee, K.M.P.; Feener, E.P.; Madigan, M.; Jackson, N.J.; Gao, B.-B.; Ross-Cisneros, F.N.; Provis, J.; Aiello, L.P.; Sadun, A.A.; Sebag, J. Proteomic analysis of embryonic and young human vitreous. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7036–7042. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Minamoto, A.; Yamashita, H.; Takamura, H.; Miyamoto-Myoken, Y.; Yoshizato, K.; Nabetani, T.; Tsugita, A.; Mishima, H.K. Proteome analysis of human vitreous proteins. Mol. Cell. Proteomics 2003, 2, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Skeie, J.M.; Roybal, C.N.; Mahajan, V.B. Proteomic insight into the molecular function of the vitreous. PLoS ONE 2015, 10, e0127567. [Google Scholar] [CrossRef] [PubMed]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Golbidi, S.; Laher, I. Antioxidant therapy in human endocrine disorders. Med. Sci. Monit. 2010, 16, RA9–RA24. [Google Scholar]
- Cicik, E.; Tekin, H.; Akar, S.; Ekmekçi, Ö.B.; Donma, O.; Koldaş, L.; Özkan, Ş. Interleukin-8, nitric oxide and glutathione status in proliferative vitreoretinopathy and proliferative diabetic retinopathy. Ophthalmic Res. 2003, 35, 251–255. [Google Scholar] [CrossRef]
- Golbidi, S.; Badran, M.; Laher, I. Antioxidant and anti-inflammatory effects of exercise in diabetic patients. Exp. Diabetes Res. 2012, 2012, 941868. [Google Scholar] [CrossRef]
- Sunitha, K.; Suresh, P.; Santhosh, M.S.; Hemshekhar, M.; Thushara, R.M.; Marathe, G.K.; Thirunavukkarasu, C.; Kemparaju, K.; Kumar, M.S.; Girish, K.S. Inhibition of hyaluronidase by N-acetyl cysteine and glutathione: Role of thiol group in hyaluronan protection. Int. J. Biol. Macromol. 2013, 55, 39–46. [Google Scholar] [CrossRef]
- Hrabárová, E.; Valachová, K.; Rapta, P.; Soltés, L. An alternative standard for Trolox-equivalent antioxidant-capacity estimation based on thiol antioxidants. Comparative 2,2′-azinobis[3-ethylbenzothiazoline-6-sulfonic acid] decolorization and rotational viscometry study regarding hyaluronan degradation. Chem. Biodivers. 2010, 7, 2191–2200. [Google Scholar] [CrossRef] [PubMed]
- Sulochana, K.N.; Biswas, J.; Ramakrishnan, S. Eales’ disease: Increased oxidation and peroxidation products of membrane constituents chiefly lipids and decreased antioxidant enzymes and reduced glutathione in vitreous. Curr. Eye Res. 1999, 19, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Géhl, Z.; Bakondi, E.; Resch, M.D.; Hegedűs, C.; Kovács, K.; Lakatos, P.; Szabó, A.; Nagy, Z.; Virág, L. Diabetes-induced oxidative stress in the vitreous humor. Redox Biol. 2016, 9, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Sebag, J. Abnormalities of human vitreous structure in diabetes. Graefes Arch. Clin. Exp. Ophthalmol. 1993, 231, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Tinggi, U. Determination of selenium in meat products by hydride generation atomic absorption spectrophotometry. J. AOAC Int. 1999, 82, 364–367. [Google Scholar] [PubMed]
- Heinämäki, A.A.; Muhonen, A.S.H.; Piha, R.S. Taurine and other free amino acids in the retina, vitreous, lens, irisciliary body, and cornea of the rat eye. Neurochem. Res. 1986, 11, 535–542. [Google Scholar] [CrossRef]
- Froger, N.; Moutsimilli, L.; Cadetti, L.; Jammoul, F.; Wang, Q.-P.; Fan, Y.; Gaucher, D.; Rosolen, S.G.; Neveux, N.; Cynober, L. Taurine: The comeback of a neutraceutical in the prevention of retinal degenerations. Prog. Retin. Eye Res. 2014, 41, 44–63. [Google Scholar] [CrossRef]
- Wright, C.E.; Tallan, H.H.; Lin, Y.Y.; Gaull, G.E. Taurine: Biological update. Annu. Rev. Biochem. 1986, 55, 427–453. [Google Scholar] [CrossRef]
- Pasantes-amorales, H.; Venture, N.B.; Wioland, N.; Mandel, P. Effect of intravitreal injections of taurine and GABA on chicken electroretinogram. Int. J. Neurosci. 1973, 5, 235–241. [Google Scholar] [CrossRef]
- Ripps, H.; Shen, W. Taurine: A “very essential” amino acid. Mol. Vis. 2012, 18, 2673. [Google Scholar]
- Schmidt, S.Y.; Berson, E.L.; Hayes, K.C. Retinal degeneration in cats fed casein. I. Taurine deficiency. Investig. Ophthalmol. Vis. Sci. 1976, 15, 47–52. [Google Scholar]
- Horwitz, J. Alpha-crystallin can function as a molecular chaperone. Proc. Natl. Acad. Sci. USA 1992, 89, 10449–10453. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.M.; Yang, Y.S. A potential role of crystallin in the vitreous bodies of rats after ischemia-reperfusion injury. Korean J. Ophthalmol. 2012, 26, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.W.; Sauter, J.L.; Johnson, P.K.; Chen, C.-D.; Olsen, T.W. Identification and localization of major soluble vitreous proteins in human ocular tissue. Am. J. Ophthalmol. 2004, 137, 655–661. [Google Scholar] [PubMed]
- Wang, H.; Feng, L.; Hu, J.W.; Xie, C.L.; Wang, F. Characterisation of the vitreous proteome in proliferative diabetic retinopathy. Proteome Sci. 2012, 10, 15. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.J.; Rho, J.H.; Yoon, Y.G.; Yoo, S.H.; Jeong, N.Y.; Ryu, W.Y.; Ahn, H.B.; Park, W.C.; Rho, S.H.; Yoon, H.S. Cytoplasmic and nuclear anti-apoptotic roles of αB-crystallin in retinal pigment epithelial cells. PLoS ONE 2012, 7, e45754. [Google Scholar] [CrossRef]
- Bounous, G.; Sukkar, S.; Molson, J.H. The antioxidant system. Anticancer Res. 2003, 23, 1411–1416. [Google Scholar]
- Gülçin, I. Comparison of in vitro antioxidant and antiradical activities of l-tyrosine and l-Dopa. Amino Acids 2007, 32, 431. [Google Scholar] [CrossRef]
- Shih, V.E. Amino acid analysis. In Physician’s Guide to the Laboratory Diagnosis of Metabolic Diseases; Springer: Berlin/Heidelberg, Germany, 2003; pp. 11–26. [Google Scholar]
- Stillwell, W. Chapter 14—Membrane Biogenesis: Fatty Acids. In An Introduction to Biological Membranes, 2nd ed.; Stillwell, W., Ed.; Elsevier: Oxford, UK, 2016; pp. 315–329. [Google Scholar]
- Angi, M.; Kalirai, H.; Coupland, S.E.; Damato, B.E.; Semeraro, F.; Romano, M.R. Proteomic analyses of the vitreous humour. Mediators Inflamm. 2012, 2012, 148039. [Google Scholar] [CrossRef]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef]
- Brock, J.H.; Mainou-Fowler, T. The role of iron and transferrin in lymphocyte transformation. Immunol. Today 1983, 4, 347–351. [Google Scholar] [CrossRef]
- Phillips, J.L. Specific binding of zinc transferrin to human lymphocytes. Biochem. Biophys. Res. Commun. 1976, 72, 634–639. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Iron and free radical reactions: Two aspects of antioxidant protection. Trends Biochem. Sci. 1986, 11, 372–375. [Google Scholar] [CrossRef]
- Wong, R.W.; Chimene, R.D.; Hahn, P.; Green, W.R.; Dunaief, J.L. Iron toxicity as a potential factor in AMD. Retina 2007, 27, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, M.; West, K.; Crabb, J.W.; Kinoshita, S.; Kamei, M. Proteomic analysis of vitreous from diabetic macular edema. Exp. Eye Res. 2005, 81, 176–182. [Google Scholar] [CrossRef]
- Skeie, J.M.; Mahajan, V.B. Proteomic interactions in the mouse vitreous-retina complex. PLoS ONE 2013, 8, e82140. [Google Scholar] [CrossRef]
- Yamagishi, S.; Nakamura, K.; Inoue, H.; Takeuchi, M. Met72Thr polymorphism of pigment epithelium-derived factor gene and susceptibility to age-related macular degeneration. Med. Hypotheses 2005, 64, 1202–1204. [Google Scholar] [CrossRef]
- Burk, R.F. Selenium, an antioxidant nutrient. Nutr. Clin. Care 2002, 5, 75–79. [Google Scholar] [CrossRef]
- Tinggi, U.; Reilly, C.; Patterson, C.M. Determination of selenium in foodstuffs using spectrofluorometry and hydride generation atomic absorption spectrometry. J. Food Compos. Anal. 1992, 5, 269–280. [Google Scholar] [CrossRef]
- Tinggi, U. Selenium: Its role as antioxidant in human health. Environ. Health Prev. Med. 2008, 13, 102. [Google Scholar] [CrossRef]
- Karcioglu, Z.A. Zinc in the eye. Surv. Ophthalmol. 1982, 27, 114–122. [Google Scholar] [CrossRef]
- Konerirajapuram, N.S.; Coral, K.; Punitham, R.; Sharma, T.; Kasinathan, N.; Sivaramakrishnan, R. Trace elements iron, copper and zinc in vitreous of patients with various vitreoretinal diseases. Indian J. Ophthalmol. 2004, 52, 145–148. [Google Scholar] [PubMed]
- Koumantakis, E.; Alexiou, D.; Grimanis, A.; Kaskarelis, D.; Bouzas, A. Zinc, cobalt and selenium concentrations in the premature and full-term newborn eye. Ophthalmologica 1983, 186, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kondoh, M. Recent studies on metallothionein: Protection against toxicity of heavy metals and oxygen free radicals. Tohoku J. Exp. Med. 2002, 196, 9–22. [Google Scholar] [CrossRef]
- Krizova, L.; Kalousova, M.; Kubena, A.; Benakova, H.; Zima, T.; Kovarik, Z.; Kalvoda, J.; Kalvodova, B. Increased uric acid and glucose concentrations in vitreous and serum of patients with diabetic macular oedema. Ophthalmic Res. 2011, 46, 73–79. [Google Scholar] [CrossRef]
- Hayden, M.R.; Tyagi, S.C. Uric acid: A new look at an old risk marker for cardiovascular disease, metabolic syndrome, and type 2 diabetes mellitus: The urate redox shuttle. Nutr. Metab. 2004, 1, 10. [Google Scholar] [CrossRef]
- Maples, K.R.; Mason, R.P. Free radical metabolite of uric acid. J. Biol. Chem. 1988, 263, 1709–1712. [Google Scholar]
- Sevanian, A.; Davies, K.J.; Hochstein, P. Serum urate as an antioxidant for ascorbic acid. Am. J. Clin. Nutr. 1991, 54, 1129S–1134S. [Google Scholar] [CrossRef]
- Sebag, J. The vitreous. In Adler’s Physiology of the Eye, 9th ed.; Hart, W.M.J., Ed.; Mosby Inc.: St. Louis, MO, USA, 1992; pp. 268–347. [Google Scholar]
- Krizova, L.; Kalousova, M.; Kubena, A.A.; Chrapek, O.; Chrapkova, B.; Sin, M.; Zima, T. Correlation of vitreous vascular endothelial growth factor and uric acid concentration using optical coherence tomography in diabetic macular edema. J. Ophthalmol. 2015, 2015, 478509. [Google Scholar] [CrossRef]
- McGahan, M.C. Ascorbic acid levels in aqueous and vitreous humors of the rabbit: Effects of inflammation and ceruloplasmin. Exp. Eye Res. 1985, 41, 291–298. [Google Scholar] [CrossRef]
- Diederen, R.M.H.; La Heij, E.C.; Deutz, N.E.P.; Kijlstra, A.; Kessels, A.G.H.; van Eijk, H.M.H.; Liem, A.T.A.; Dieudonné, S.; Hendrikse, F. Increased glutamate levels in the vitreous of patients with retinal detachment. Exp. Eye Res. 2006, 83, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Mayer, U. Comparative investigations of catalase activity in different ocular tissues of cattle and man. Graefe. Arch. Klin. Exp. Ophthal. 1980, 213, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Behndig, A.; Svensson, B.; Marklund, S.L.; Karlsson, K. Superoxide dismutase isoenzymes in the human eye. Investig. Ophthalmol. Vis. Sci. 1998, 39, 471–475. [Google Scholar]
- Weisiger, R.A.; Fridovich, I. Mitochondrial superoxide simutase. Site of synthesis and intramitochondrial localization. J. Biol. Chem. 1973, 248, 4793–4796. [Google Scholar]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar]
- Marklund, S.L. Human copper-containing superoxide dismutase of high molecular weight. Proc. Natl. Acad. Sci. USA 1982, 79, 7634–7638. [Google Scholar] [CrossRef]
- Wert, K.J.; Velez, G.; Cross, M.R.; Wagner, B.A.; Teoh-Fitzgerald, M.L.; Buettner, G.R.; McAnany, J.J.; Olivier, A.; Tsang, S.H.; Harper, M.M.; et al. Extracellular superoxide dismutase (SOD3) regulates oxidative stress at the vitreoretinal interface. Free Radic. Biol. Med. 2018, 124, 408–419. [Google Scholar] [CrossRef]
- De Vega, R.G.; Fernández-Sánchez, M.L.; Iglesias, H.G.; Prados, M.C.; Sanz-Medel, A. Quantitative selenium speciation by HPLC-ICP-MS (IDA) and simultaneous activity measurements in human vitreous humor. Anal. Bioanal. Chem. 2015, 407, 2405–2413. [Google Scholar] [CrossRef]
- Herbette, S.; Roeckel-Drevet, P.; Drevet, J.R. Seleno-independent glutathione peroxidases: More than simple antioxidant scavengers. FEBS J. 2007, 274, 2163–2180. [Google Scholar] [CrossRef]
- Bhuyan, K.C.; Bhuyan, D.K. Catalase in ocular tissue and its intracellular distribution in corneal epithelium. Am. J. Ophthalmol. 1970, 69, 147–153. [Google Scholar] [CrossRef]
- Barracchini, A.; Franceschini, N.; Filippello, M.; Pantaleoni, P.; Di, A.G.; Amicosante, G.; Pantaleoni, G. Leukocyanidines and collagenases: In vitro enzyme inhibition activity. Clin. Ter. 1999, 150, 275–278. [Google Scholar] [PubMed]
- Mosca, M.; Ambrosone, L.; Semeraro, F.; Casamassima, D.; Vizzarri, F.; Costagliola, C. Ocular tissues and fluids oxidative stress in hares fed on verbascoside supplement. Int. J. Food Sci. Nutr. 2014, 65, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Liao, S.; Mi, H.; Guo, C.; Qi, D.; Li, F.; Zhang, C.; Yang, Z. Hesperidin prevents retinal and plasma abnormalities in streptozotocin-induced diabetic rats. Molecules 2012, 17, 12868–12881. [Google Scholar] [CrossRef] [PubMed]
- Pradeep, K.; Ko, K.C.; Choi, M.H.; Kang, J.A.; Chung, Y.J.; Park, S.H. Protective effect of hesperidin, a citrus flavanoglycone, against γ-radiation-induced tissue damage in Sprague–Dawley rats. J. Med. Food 2012, 15, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Urios, P.; Grigorova-Borsos, A.-M.; Sternberg, M. Flavonoids inhibit the formation of the cross-linking AGE pentosidine in collagen incubated with glucose, according to their structure. Eur. J. Nutr. 2007, 46, 139. [Google Scholar] [CrossRef]
- Sulochana, K.N.; Ramprasad, S.; Coral, K.; Lakshmi, S.; Punitham, R.; Narayanasamy, A.; Ramakrishnan, S. Glycation and glycoxidation studies in vitro on isolated human vitreous collagen. Med. Sci. Monit. 2003, 9, BR220–BR224. [Google Scholar]
- Murthy, K.R.; Goel, R.; Subbannayya, Y.; Jacob, H.K.C.; Murthy, P.R.; Manda, S.S.; Patil, A.H.; Sharma, R.; Sahasrabuddhe, N.A.; Parashar, A. Proteomic analysis of human vitreous humor. Clin. Proteom. 2014, 11, 29. [Google Scholar] [CrossRef]
- Łukasik, M.; Małkowska, A.; Cieślak, P.A.; Sołtyszewski, I.; Szutowski, M. Postmortem Vitreous Humor Analysis for Xenobiotics and their Metabolites. Law Forensic Sci. 2018, 15, 1–8. [Google Scholar]
- Occhiutto, M.L.; Freitas, F.R.; Maranhao, R.C.; Costa, V.P. Breakdown of the blood-ocular barrier as a strategy for the systemic use of nanosystems. Pharmaceutics 2012, 4, 252–275. [Google Scholar] [CrossRef]
- Honkanen, R.A.; Baruah, S.; Zimmerman, M.B.; Khanna, C.L.; Weaver, Y.K.; Narkiewicz, J.; Waziri, R.; Gehrs, K.M.; Weingeist, T.A.; Boldt, H.C. Vitreous amino acid concentrations in patients with glaucoma undergoing vitrectomy. Arch. Ophthalmol. 2003, 121, 183–188. [Google Scholar] [CrossRef]
- Ishikawa, S.; Nakazawa, M.; Ishikawa, A.; Ishiguro, S.-I.; Tamai, M. Alteration of glutamine concentration in the vitreous humor in patients with proliferative vitreoretinopathy. Curr. Eye Res. 1995, 14, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.; Varma, L.T.; Yadav, K. Drug Delivery to Posterior Segment of the Eye: Conventional Delivery Strategies, Their Barriers, and Restrictions. In Drug Delivery for the Retina and Posterior Segment Disease; Springer: Berlin/Heidelberg, Germany, 2018; pp. 51–67. [Google Scholar]
- Institute of Medicine. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids; The National Academies Press: Washington, DC, USA, 2000. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Antioxidant | Author | Human Vitreous * | Animal Vitreous | Vitreous from Diseased Eye |
|---|---|---|---|---|
| Ascorbic Acid | Duarte & Lunec [99] McGahan [162] Park et al. [100] | 2 mmol/L 172.7 µg/mL | 0.43 mmol/kg—rabbit | 19.1 µg/mL—PDR |
| Riboflavin | Philpot & Pirie [104] Long [105] | 0.8 µg/ 100 mL—ox 8.0 µg/L—bovine | ||
| Glutathione | Sulochana et al. [121] Cicik et al. [117] Géhl et al. [122] | 0.26 mmol/L 2.35 µmol/µg protein | 2.8 µg/mg protein—ED 17.7 µg/mg protein—DVH 0.58 µmol/L—PDR 15.7 µmol/L—PVR 4.54 µmol/µg protein—PDR | |
| Taurine | Diederen et al. [163] Heinämäki et al. [125] | 22.6 µM | 1.72 µmol/mL | 26.0 µM—RRD 28.1 µM—PDR |
| Uric acid | Sebag [160] Krizova et al. [156,161] | 156–170 µmol/L | 170 µM—bovine | 52–70 µmol/L—DMO |
| Tyrosine | Shih [138] | 91 µmol/L | ||
| Transferrin | Kokavec [22] | 0.0878 g/L | ||
| Selenium | Kokavec [22] | 0.1035µmol/L | ||
| Zinc | Kokavec [22] | 1.95µmol/L | ||
| Superoxide dismutase | Sulochana et al. [121] | 0.9 IU/mg protein—ED 22.1 IU/mg protein—DVH | ||
| Glutathione peroxidase | Sulochana et al. [121] | 0.61 µmol of GSH utilized/mg protein/min—ED 0.49 µmol of GSH utilized/mg protein/min—DVH | ||
| Catalase | Mayer [164] | 58 µL O2/mg protein | ||
| PEDF | Ouchi et al. [146] | 0.83 µg/mL | 2.03 µg/mL—DME |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ankamah, E.; Sebag, J.; Ng, E.; Nolan, J.M. Vitreous Antioxidants, Degeneration, and Vitreo-Retinopathy: Exploring the Links. Antioxidants 2020, 9, 7. https://doi.org/10.3390/antiox9010007
Ankamah E, Sebag J, Ng E, Nolan JM. Vitreous Antioxidants, Degeneration, and Vitreo-Retinopathy: Exploring the Links. Antioxidants. 2020; 9(1):7. https://doi.org/10.3390/antiox9010007
Chicago/Turabian StyleAnkamah, Emmanuel, J. Sebag, Eugene Ng, and John M. Nolan. 2020. "Vitreous Antioxidants, Degeneration, and Vitreo-Retinopathy: Exploring the Links" Antioxidants 9, no. 1: 7. https://doi.org/10.3390/antiox9010007
APA StyleAnkamah, E., Sebag, J., Ng, E., & Nolan, J. M. (2020). Vitreous Antioxidants, Degeneration, and Vitreo-Retinopathy: Exploring the Links. Antioxidants, 9(1), 7. https://doi.org/10.3390/antiox9010007