Anti-Inflammatory, Antioxidant, and Microbiota-Modulating Effects of Camellia Oil from Camellia brevistyla on Acetic Acid-Induced Colitis in Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Edible Oils
2.2. Chemical Features and Antioxidant Activity of Oil
2.3. In Vitro Gastric and Small Intestinal Digestion
2.4. In Vitro Fermentation of Human Intestinal Microbiota
2.5. Animal Grouping and Treatment Procedures
2.6. Bacterial DNA Purification and 16S rRNA Sequencing
2.7. Histopathology
2.8. Preparation of Colon Homogenate and Total Protein Concentration
2.9. Measurement of IL-6, IL-1ß, and TNF-α Levels
2.10. Estimation of Superoxide Dismutase (SOD), Glutathione (GSH), Malondialdehyde–Thiobarbituric Acid Reactive Substances (MDA–TBARSs), and Myeloperoxidase (MPO)
2.11. Statistical Analysis
3. Results
3.1. Chemical Compositions and Antioxidant Activity of PCO and TCCO
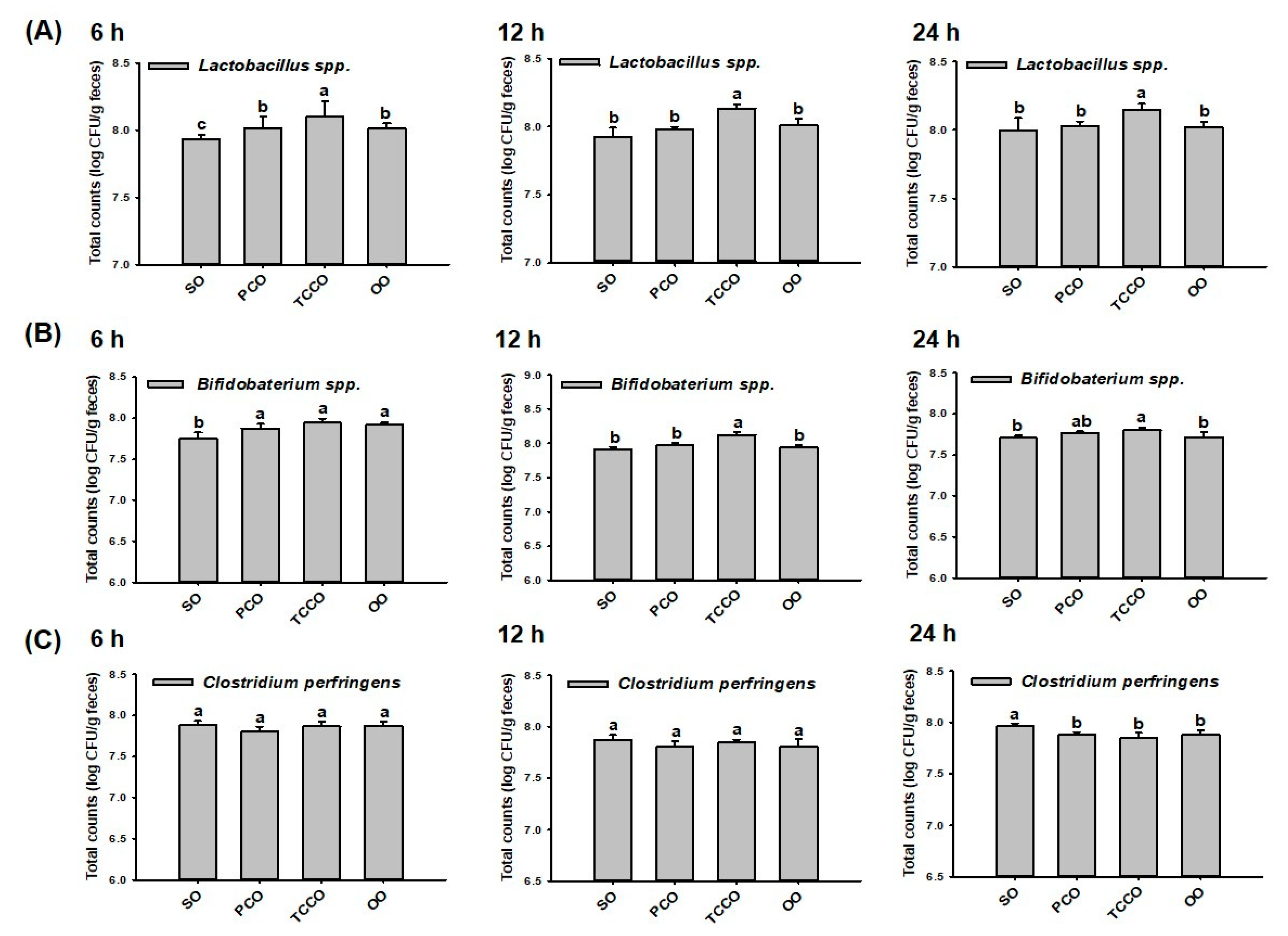
3.2. Effects of PCO and TCCO on the Bacterial Proliferation In Vitro
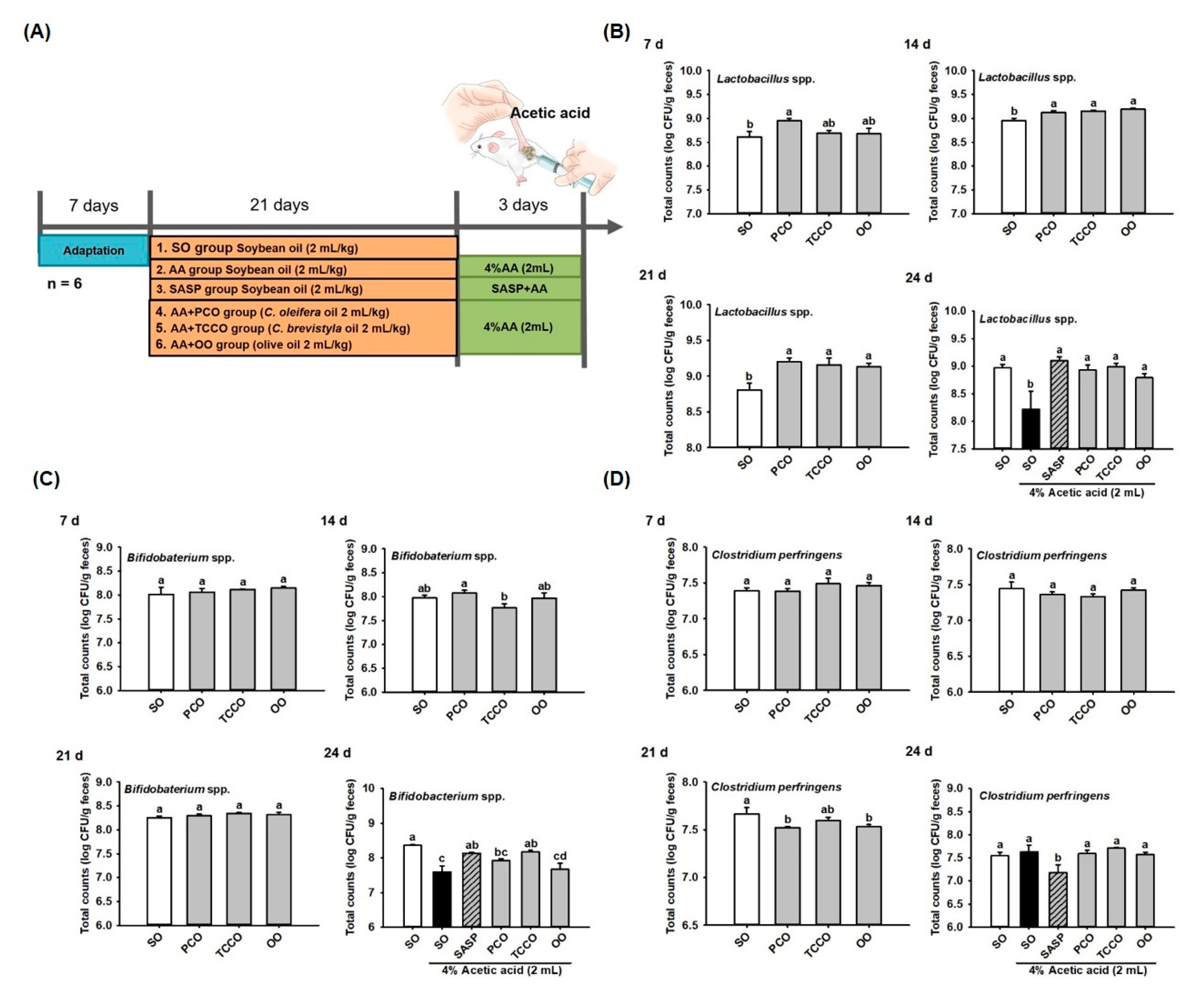
3.3. Effect of Camellia Oils on the Growth of Gut Microbiota in Rats
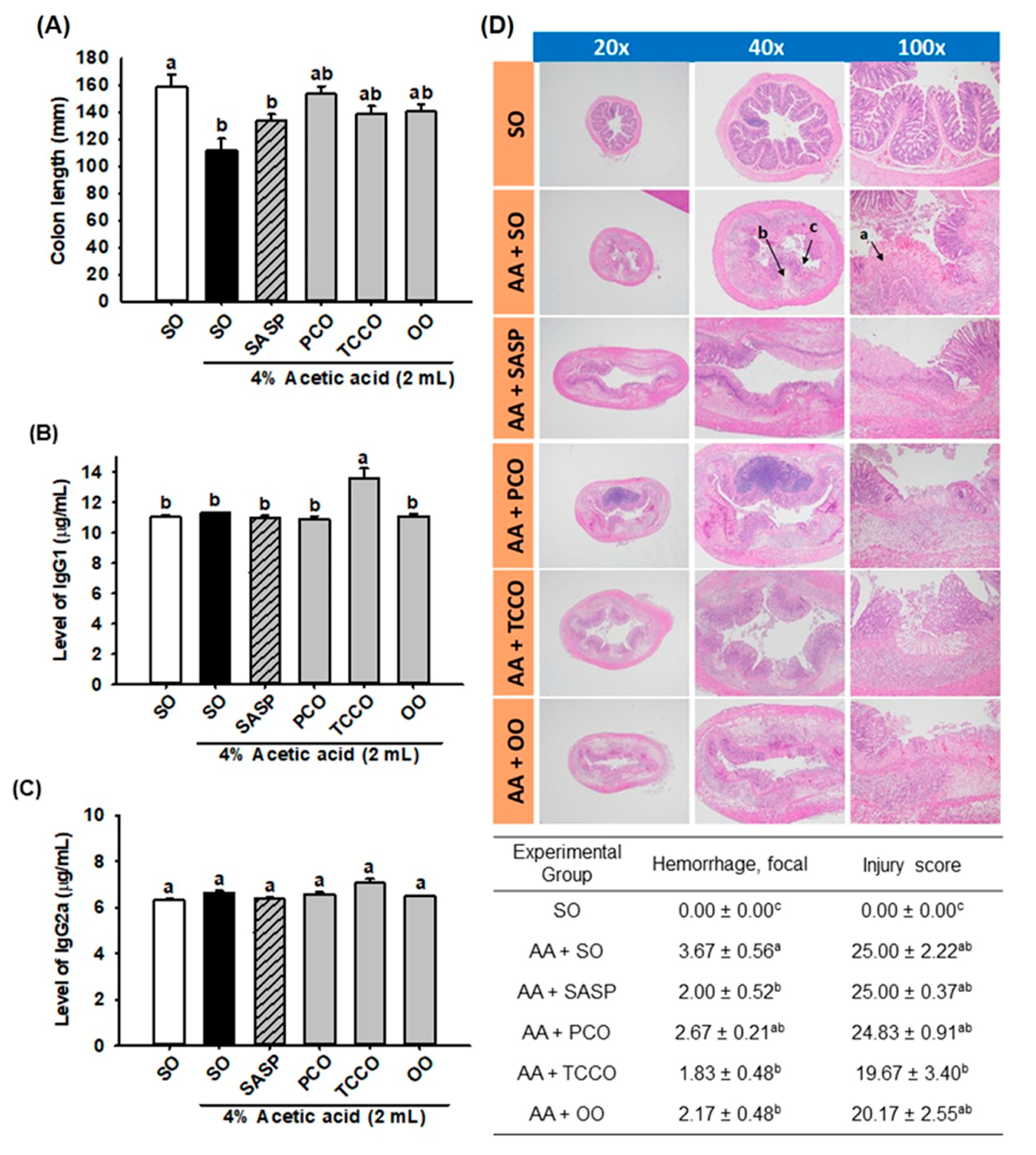
3.4. Effects of Camellia Oils on the Colon Length and the Levels of IgG1 and IgG2a
3.5. Effects of Camellia Oils on the Histological Changes in a Rat Model of AA-Induced Colitis
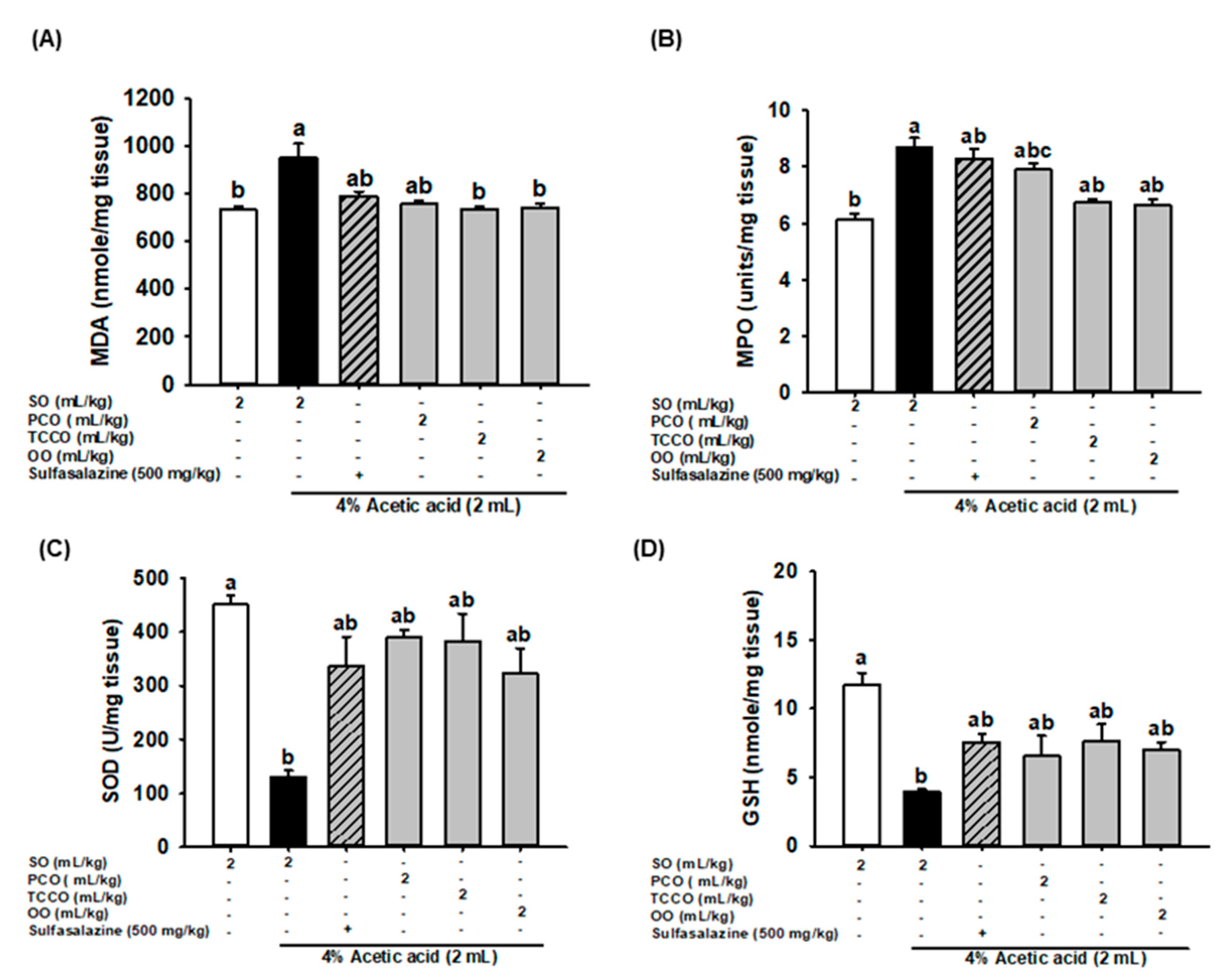
3.6. Effects of Camellia Oils on Colonic MDA, MPO, SOD, and GSH Secretion in AA-Induced Colitis in Rats
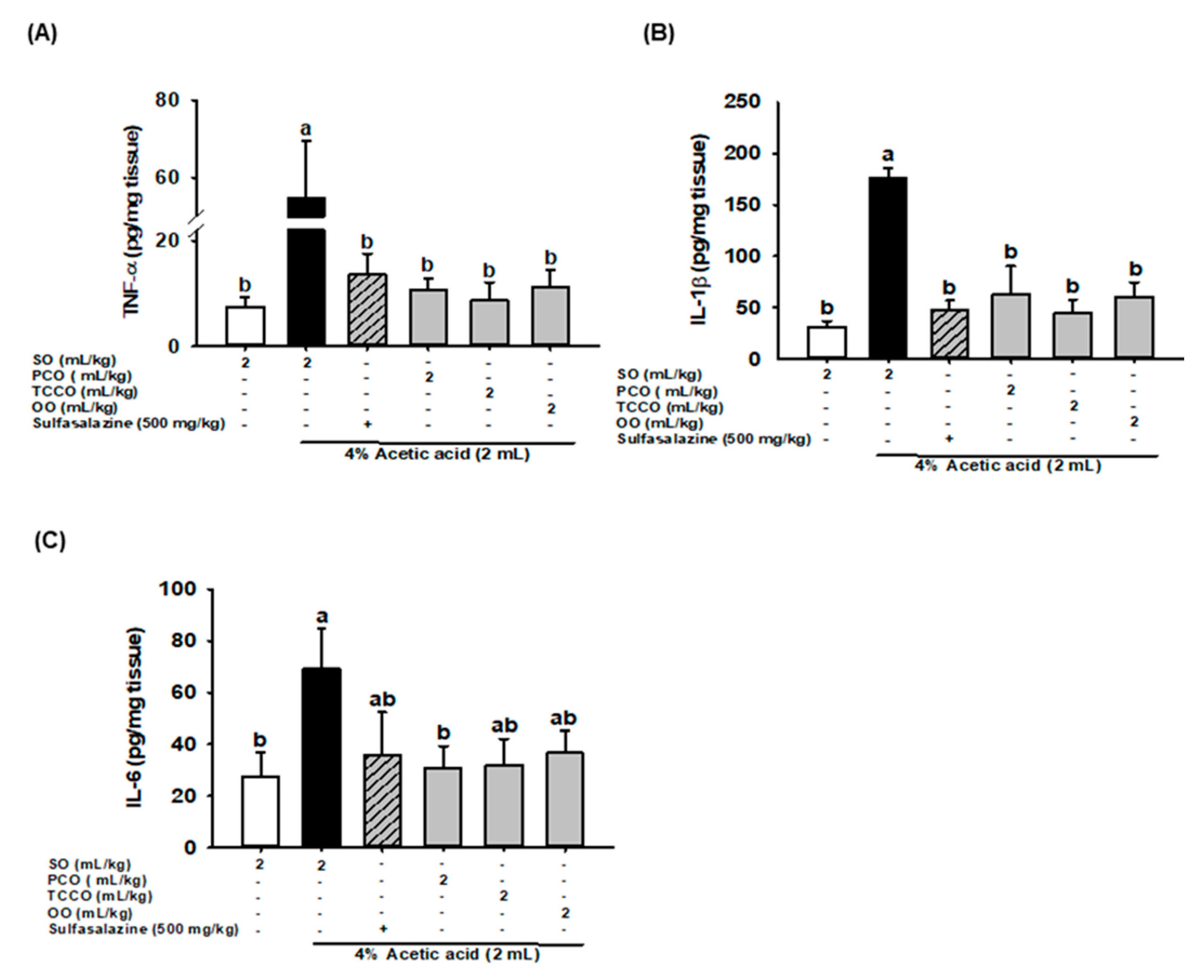
3.7. Effects of Camellia Oils on Colonic IL-6, IL-1β, and TNF-α production in AA-Induced Colitis
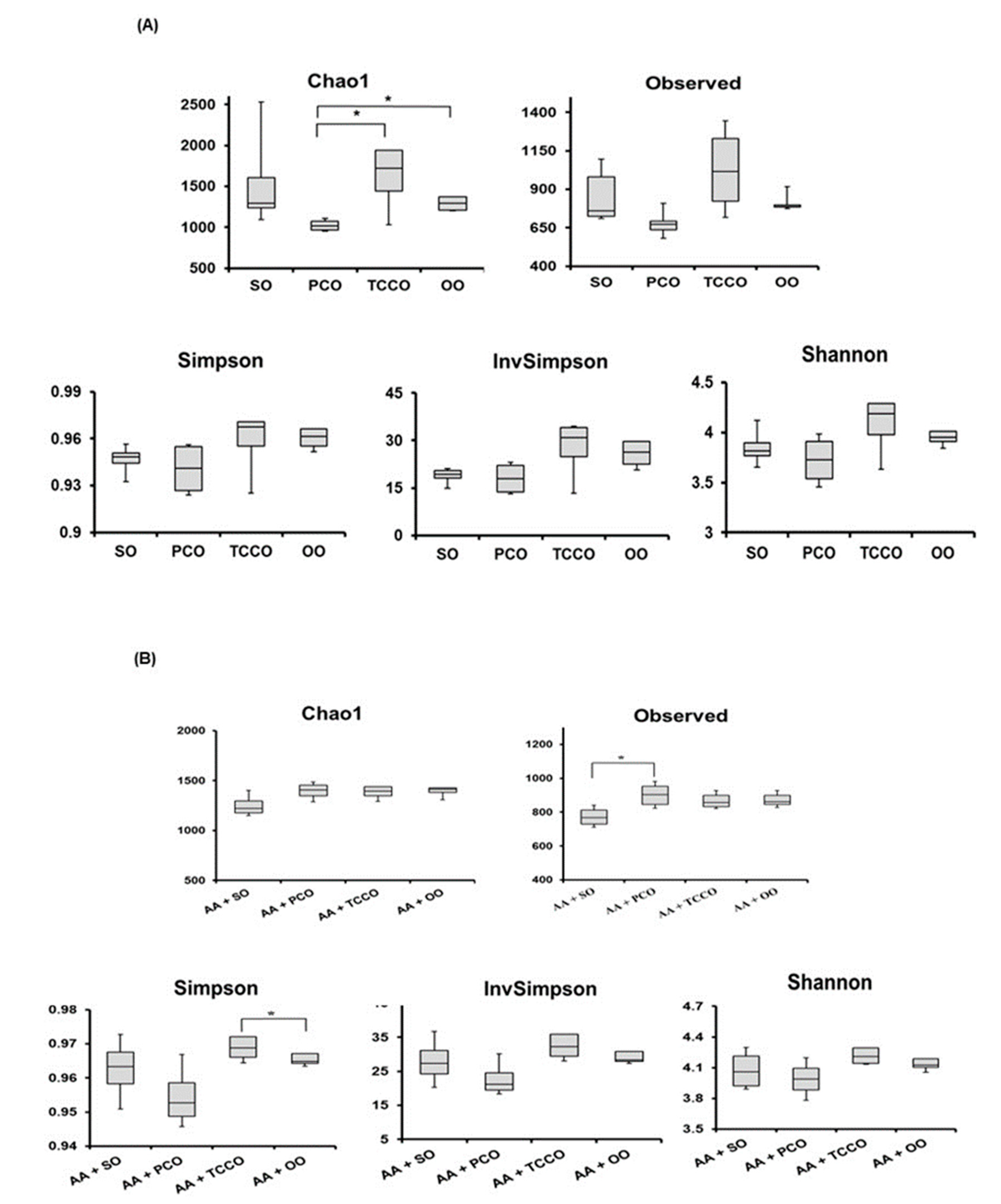
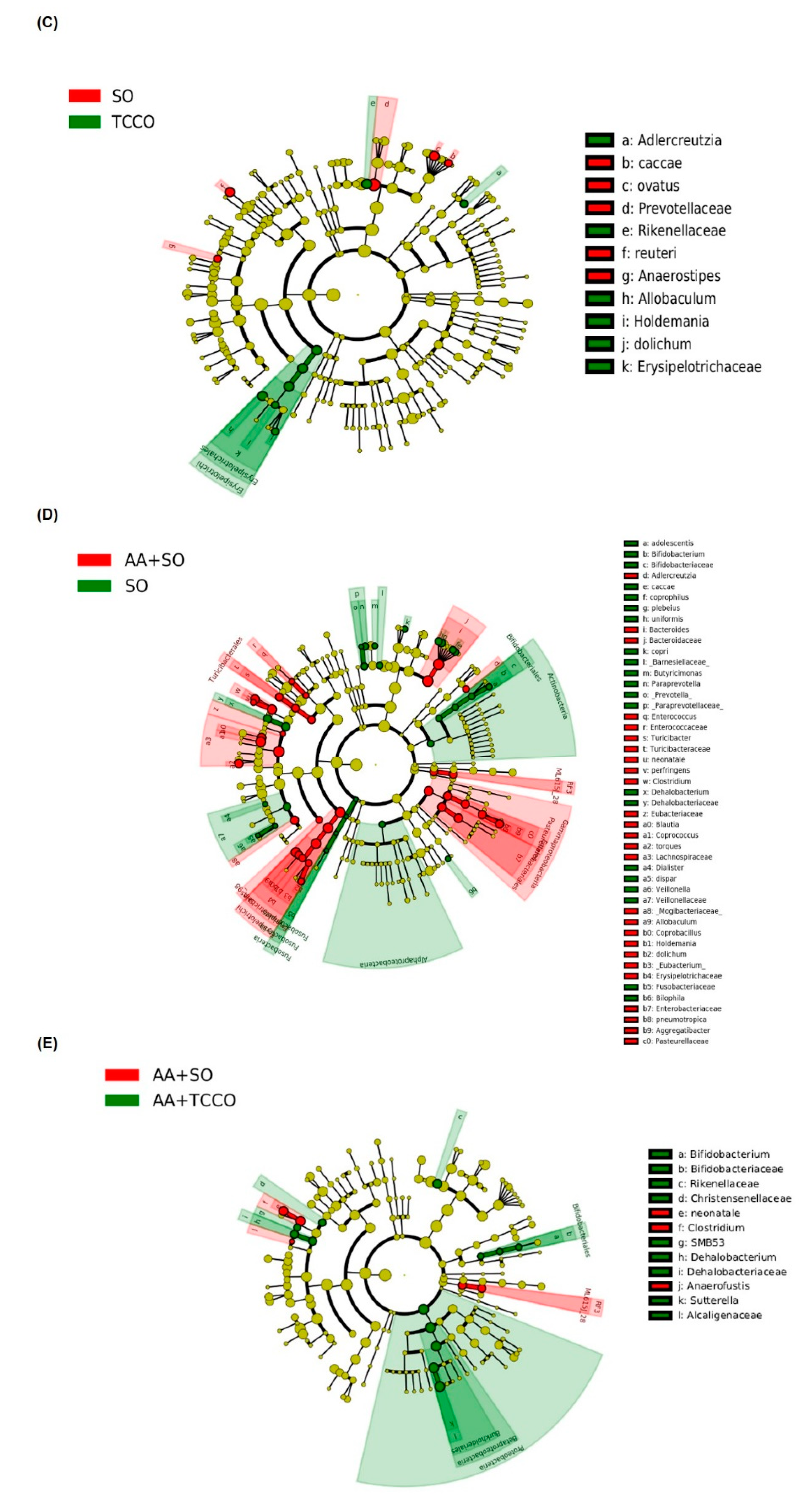
3.8. Effects of Camellia Oils on the Gut Microbiota of Rats
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AA | Acetic acid |
| CD | Crohn’s disease |
| CFU | Colony-forming unit |
| GSH | Glutathione |
| IBD | Inflammatory bowel disease |
| IBS | Irritable bowel syndrome |
| LDA | Linear discriminant analysis |
| LEfSe | LDA effect size |
| MDA | Malondialdehyde |
| MPO | Myeloperoxidase |
| NSAIDs | Non-steroidal anti-inflammatory drugs |
| OO | Olive oil |
| ORAC | Oxygen radical absorbance capacity |
| OUT | Operational taxonomic units |
| PCO | Camellia oil from Camellia oleifera |
| ROS | Reactive oxygen species |
| SASP | Sulfasalazine |
| SO | Soybean oil |
| SOD | Superoxide dismutase |
| TCCO | Camellia oil from Camellia brevistyla |
| TEAC | Trolox equivalent antioxidant capacity |
| UC | Ulcerative colitis |
References
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2018, 390, 2769–2778. [Google Scholar] [CrossRef]
- Moura, F.A.; de Andrade, K.Q.; Dos Santos, J.C.F.; Araújo, O.R.P.; Goulart, M.O.F. Antioxidant therapy for treatment of inflammatory bowel disease: Does it work? Redox. Biol. 2015, 6, 617–639. [Google Scholar] [CrossRef]
- Rodino-Janeiro, B.K.; Martinez, C.; Fortea, M.; Lobo, B.; Pigrau, M.; Nieto, A.; Gonzalez-Castro, A.M.; Salvo-Romero, E.; Guagnozzi, D.; Pardo-Camacho, C.; et al. Decreased TESK1-mediated cofilin 1 phosphorylation in the jejunum of IBS-D patients may explain increased female predisposition to epithelial dysfunction. Sci. Rep. 2018, 8, 2255. [Google Scholar] [CrossRef]
- Andoh, A.; Yagi, Y.; Shioya, M.; Nishida, A.; Tsujikawa, T.; Fujiyama, Y. Mucosal cytokine network in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 5154–5161. [Google Scholar] [CrossRef]
- Sartor, R.B. Mechanisms of disease: Pathogenesis of Crohn’s disease and ulcerative colitis. Nat. Rev. Gastroenterol. Hepatol. 2006, 3, 390. [Google Scholar] [CrossRef] [PubMed]
- Neish, A.S. Microbes in gastrointestinal health and disease. Gastroenterology 2009, 136, 65–80. [Google Scholar] [CrossRef]
- Lee, C.P.; Yen, G.C. Antioxidant activity and bioactive compounds of tea seed (Camellia oleifera Abel.) oil. J. Agric. Food Chem. 2006, 54, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.P.; Shih, P.H.; Hsu, C.L.; Yen, G.C. Hepatoprotection of tea seed oil (Camellia oleifera Abel.) against CCl4-induced oxidative damage in rats. Food Chem. Toxicol. 2007, 45, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.T.; Wu, S.L.; Ho, C.Y.; Huang, S.M.; Cheng, C.L.; Yen, G.C. Beneficial effects of Camellia Oil (Camellia oleifera Abel.) on ketoprofen-induced gastrointestinal mucosal damage through upregulation of HO-1 and VEGF. J. Agric. Food Chem. 2014, 62, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Tu, P.S.; Tung, Y.T.; Lee, W.T.; Yen, G.C. Protective Effect of Camellia Oil (Camellia oleifera Abel.) against Ethanol-Induced Acute Oxidative Injury of the Gastric Mucosa in Mice. J. Agric. Food Chem. 2017, 65, 4932–4941. [Google Scholar] [CrossRef]
- Lee, W.T.; Tung, Y.T.; Wu, C.C.; Tu, P.S.; Yen, G.C. Camellia Oil (Camellia oleifera Abel.) Modifies the Composition of Gut Microbiota and Alleviates Acetic Acid-Induced Colitis in Rats. J. Agric. Food Chem. 2018, 66, 7384–7392. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Chen, C.S.; Chang, W.T.; Wu, M.F.; Cheng, F.T.; Shiau, D.K.; Hsu, C.L. Antioxidant activity and anticancer effect of ethanolic and aqueous extracts of the roots of Ficus beecheyana and their phenolic components. J. Food Drug Anal. 2018, 26, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Flakelar, C.L.; Prenzler, P.D.; Luckett, D.J.; Howitt, J.A.; Doran, G. A rapid method for the simultaneous quantification of the major tocopherols, carotenoids, free and esterified sterols in canola (Brassica napus) oil using normal phase liquid chromatography. Food Chem. 2017, 214, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xie, M.; Wan, P.; Chen, D.; Ye, H.; Chen, L.; Zeng, X.; Liu, Z. Digestion under saliva, simulated gastric and small intestinal conditions and fermentation in vitro by human intestinal microbiota of polysaccharides from Fuzhuan brick tea. Food Chem. 2018, 244, 331–339. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Randhawa, P.K.; Singh, K.; Singh, N.; Jaggi, A.S. A review on chemical-induced inflammatory bowel disease models in rodents. Korean J. Physiol. Pharmacol. 2014, 18, 279–288. [Google Scholar] [CrossRef]
- Horton, N.; Wu, X.; Philpott, J.; Garber, A.; Achkar, J.P.; Brzezinski, A.; Lashner, B.A.; Shen, B. Impact of Low Immunoglobulin G Levels on Disease Outcomes in Patients with Inflammatory Bowel Diseases. Dig. Dis. Sci. 2016, 61, 3270–3277. [Google Scholar] [CrossRef]
- Closa, D.; Folch-Puy, E. Oxygen free radicals and the systemic inflammatory response. IUBMB Life 2004, 56, 185–191. [Google Scholar] [CrossRef]
- Ye, Y.; Xing, H.T.; Guo, Y. Hypolipidemic effect of a novel biflavonoid from shells of Camellia oleifera (Abel.). Indian J. Exp. Biol. 2013, 51, 458–463. [Google Scholar] [PubMed]
- Xiao, X.; He, L.; Chen, Y.; Wu, L.; Wang, L.; Liu, Z. Anti-inflammatory and antioxidative effects of Camellia oleifera Abel components. Future Med. Chem. 2017, 9, 2069–2079. [Google Scholar] [CrossRef] [PubMed]
- Dicksved, J.; Schreiber, O.; Willing, B.; Petersson, J.; Rang, S.; Phillipson, M.; Holm, L.; Roos, S. Lactobacillus reuteri maintains a functional mucosal barrier during DSS treatment despite mucus layer dysfunction. PLoS ONE 2012, 7, e46399. [Google Scholar] [CrossRef] [PubMed]
- Tzounis, X.; Vulevic, J.; Kuhnle, G.G.; George, T.; Leonczak, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Flavanol monomer-induced changes to the human faecal microflora. Br. J. Nutr. 2008, 99, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Manichanh, C.; Borruel, N.; Casellas, F.; Guarner, F. The gut microbiota in IBD. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.H.; Duan, J.A.; Jiang, S.; Feng, N.N.; Qiu, W.Q.; Ling, Y. Polysaccharides from Chrysanthemum morifolium Ramat ameliorate colitis rats by modulating the intestinal microbiota community. Oncotarget 2017, 8, 80790–80803. [Google Scholar] [CrossRef] [PubMed]
- Darnaud, M.; Dos Santos, A.; Gonzalez, P.; Augui, S.; Lacoste, C.; Desterke, C.; De Hertogh, G.; Valentino, E.; Braun, E.; Zheng, J.; et al. Enteric Delivery of Regenerating Family Member 3 alpha Alters the Intestinal Microbiota and Controls Inflammation in Mice with Colitis. Gastroenterology 2018, 154, 1009–1023. [Google Scholar] [CrossRef]
- Hansen, A.K.; Hansen, C.H.; Krych, L.; Nielsen, D.S. Impact of the gut microbiota on rodent models of human disease. World J. Gastroenterol. 2014, 20, 17727–17736. [Google Scholar] [CrossRef]
- Kaakoush, N.O. Insights into the Role of Erysipelotrichaceae in the Human Host. Front. Cell. Infect. Microbiol. 2015, 5, 84. [Google Scholar] [CrossRef]
- Glavan, T.W.; Gaulke, C.A.; Santos Rocha, C.; Sankaran-Walters, S.; Hirao, L.A.; Raffatellu, M.; Jiang, G.; Baumler, A.J.; Goulart, L.R.; Dandekar, S. Gut immune dysfunction through impaired innate pattern recognition receptor expression and gut microbiota dysbiosis in chronic SIV infection. Mucosal Immunol. 2016, 9, 677–688. [Google Scholar] [CrossRef]
- Hiippala, K.; Kainulainen, V.; Kalliomaki, M.; Arkkila, P.; Satokari, R. Mucosal Prevalence and Interactions with the Epithelium Indicate Commensalism of Sutterella spp. Front. Microbiol. 2016, 7, 1706. [Google Scholar] [CrossRef] [PubMed]
- Roze, J.C.; Ancel, P.Y.; Lepage, P.; Martin-Marchand, L.; Al Nabhani, Z.; Delannoy, J.; Picaud, J.C.; Lapillonne, A.; Aires, J.; Durox, M.; et al. Nutritional strategies and gut microbiota composition as risk factors for necrotizing enterocolitis in very-preterm infants. Am. J. Clin. Nutr. 2017, 106, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Liu, S.; Zhou, Y.; Yao, Z.; Zhang, D.; Cao, S.; Wei, Z.; Tan, B.; Li, Y.; Lian, Z.; et al. Evolutionary biologic changes of gut microbiota in an ‘adenoma-carcinoma sequence’ mouse colorectal cancer model induced by 1,2-Dimethylhydrazine. Oncotarget 2017, 8, 444–457. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.-C.; Tung, Y.-T.; Chen, S.-Y.; Lee, W.-T.; Lin, H.-T.; Yen, G.-C. Anti-Inflammatory, Antioxidant, and Microbiota-Modulating Effects of Camellia Oil from Camellia brevistyla on Acetic Acid-Induced Colitis in Rats. Antioxidants 2020, 9, 58. https://doi.org/10.3390/antiox9010058
Wu C-C, Tung Y-T, Chen S-Y, Lee W-T, Lin H-T, Yen G-C. Anti-Inflammatory, Antioxidant, and Microbiota-Modulating Effects of Camellia Oil from Camellia brevistyla on Acetic Acid-Induced Colitis in Rats. Antioxidants. 2020; 9(1):58. https://doi.org/10.3390/antiox9010058
Chicago/Turabian StyleWu, Chun-Ching, Yu-Tang Tung, Sheng-Yi Chen, Wei-Ting Lee, Hsin-Tang Lin, and Gow-Chin Yen. 2020. "Anti-Inflammatory, Antioxidant, and Microbiota-Modulating Effects of Camellia Oil from Camellia brevistyla on Acetic Acid-Induced Colitis in Rats" Antioxidants 9, no. 1: 58. https://doi.org/10.3390/antiox9010058
APA StyleWu, C.-C., Tung, Y.-T., Chen, S.-Y., Lee, W.-T., Lin, H.-T., & Yen, G.-C. (2020). Anti-Inflammatory, Antioxidant, and Microbiota-Modulating Effects of Camellia Oil from Camellia brevistyla on Acetic Acid-Induced Colitis in Rats. Antioxidants, 9(1), 58. https://doi.org/10.3390/antiox9010058