Neuroprotective Mechanisms of Three Natural Antioxidants on a Rat Model of Parkinson’s Disease: A Comparative Study


 ,
,  ,
,  ,
,  ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
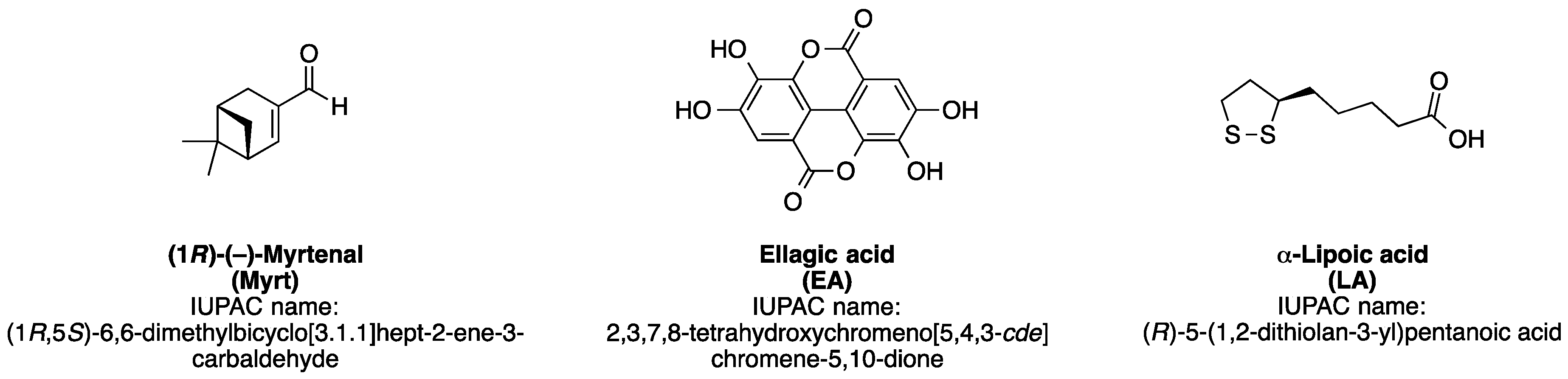
2.1. Materials
2.2. Animals
2.3. Experimental Protocol of the PD Model
2.4. Drug Treatment and Experimental Design
Animal Groups and Experimental Protocols
- SO (sham operated, 2 µL saline in striatum and treated daily with 0.5 mL/100 g saline, i.p.),
- 6-OHDA (2 µL/10 µg 6-OHDA in striatum and treated daily with 0.5 mL/100 g saline, i.p.),
- 6-OHDA + EA (lesioned and treated daily with 50 mg/kg EA, i.p.),
- 6-OHDA + LA (lesioned and treated daily with 50 mg/kg LA, i.p.),
- 6-OHDA + Myrt (lesioned and treated daily with 50 mg/kg Myrt, i.p.).
2.5. Behavioral Observations
2.5.1. Apomorphine-Induced Rotation Test (Apo)
2.5.2. Rotarod Test (RR)
2.5.3. Learning and Memory (Passive Avoidance Test)
2.6. Neurochemical Determination of Dopamine
2.7. Analytical Methods
2.8. Statistical Analysis
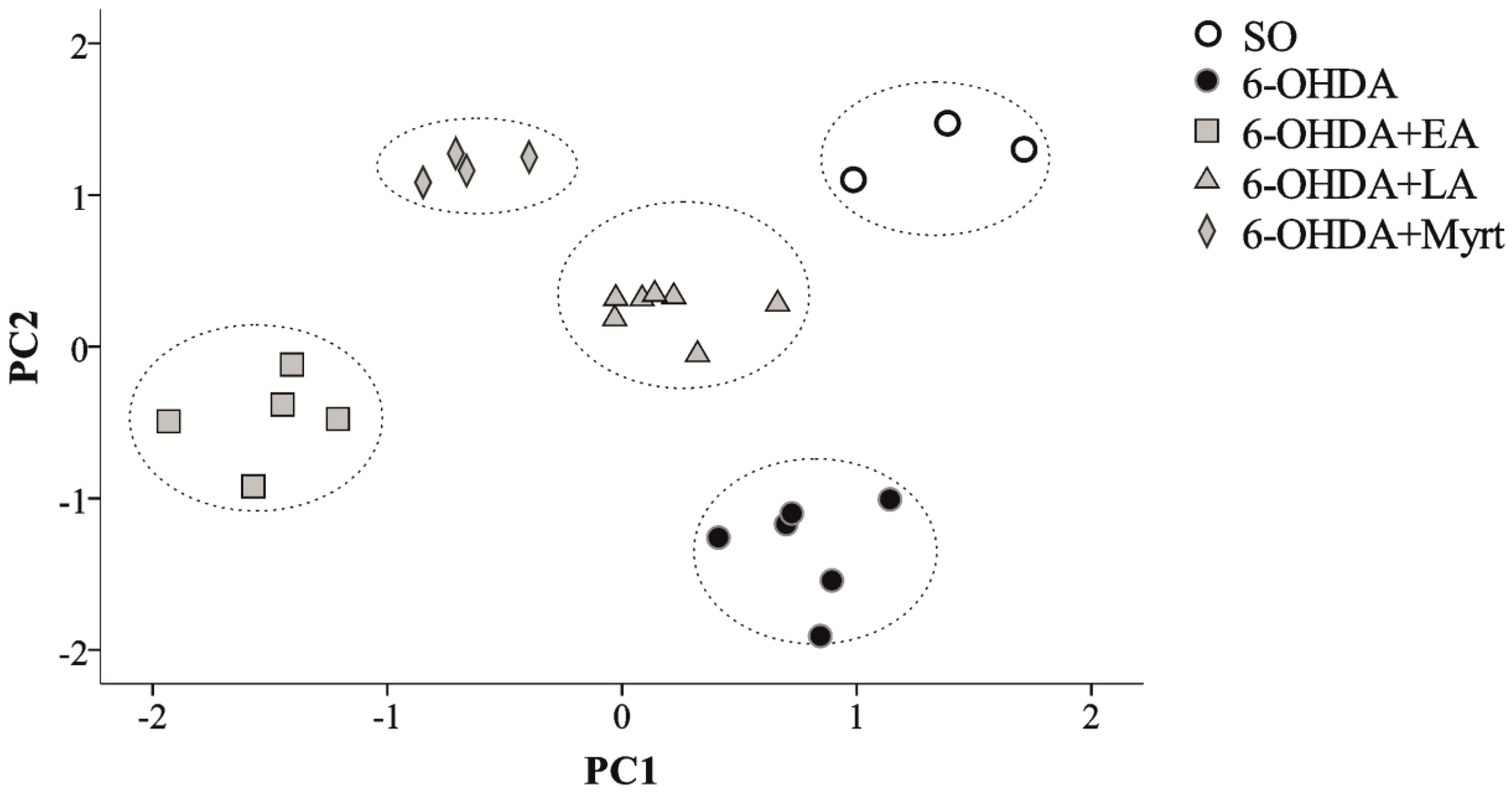
Principal Component Analysis
3. Results
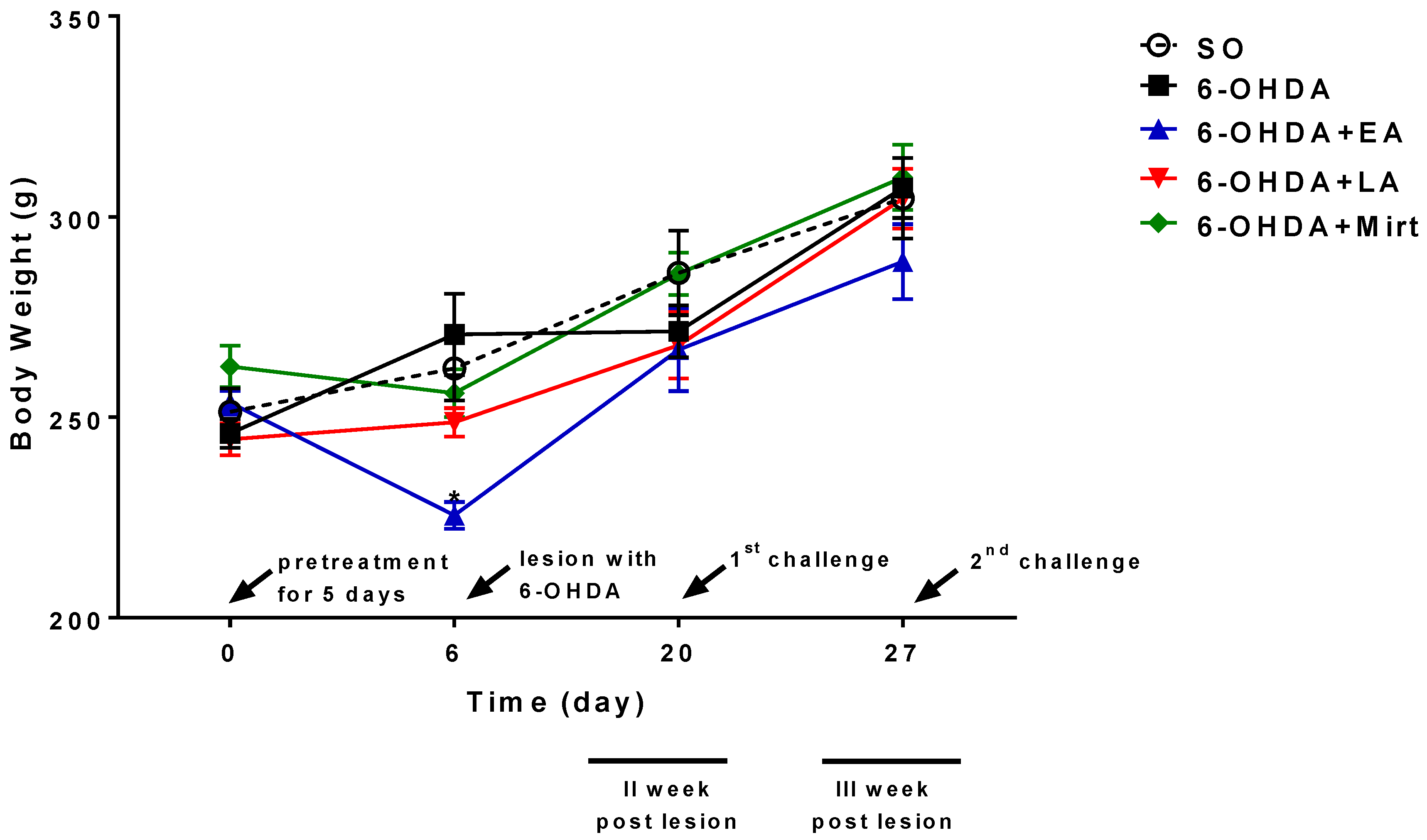
3.1. Effect of EA, LA, and Myrt on Body Weight of Wistar Rats
3.2. Behavioral Observation
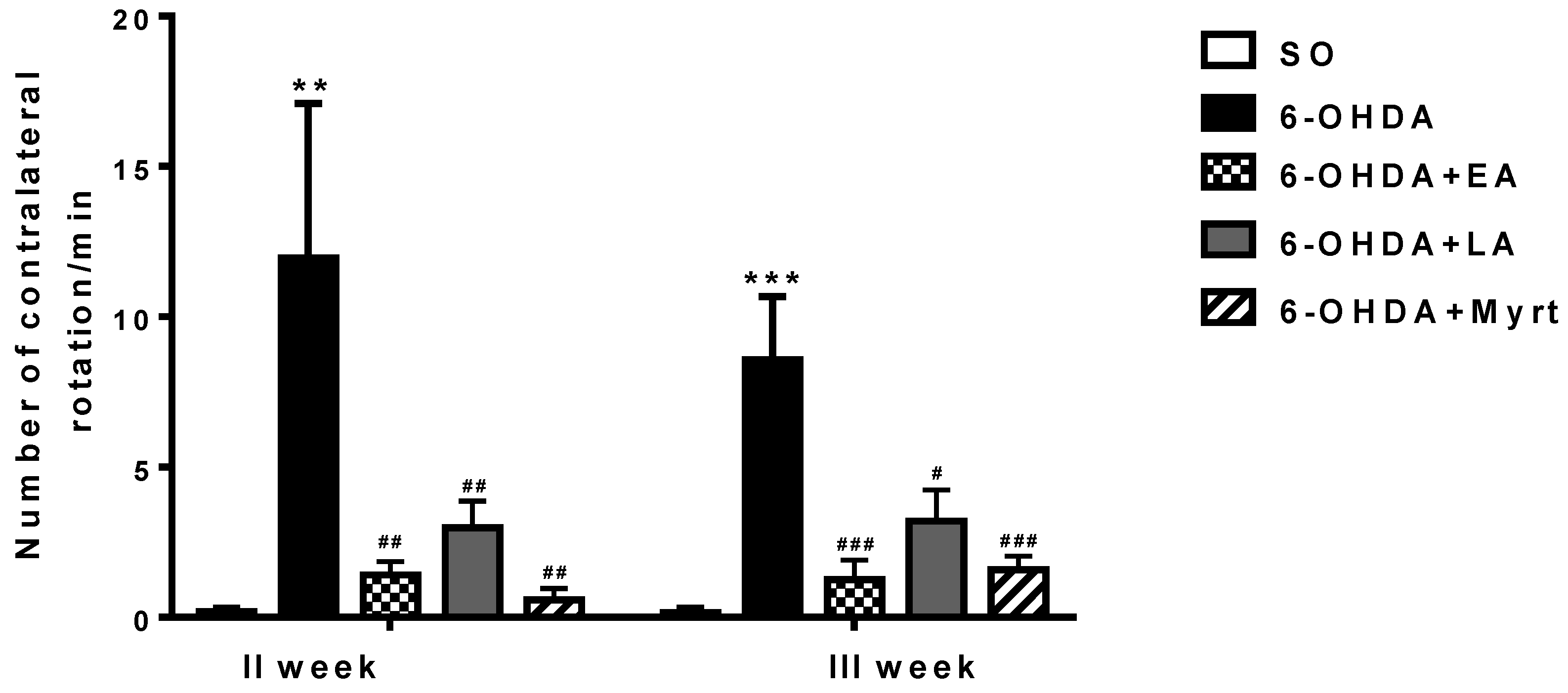
3.2.1. Apomorphine-Induced Rotations
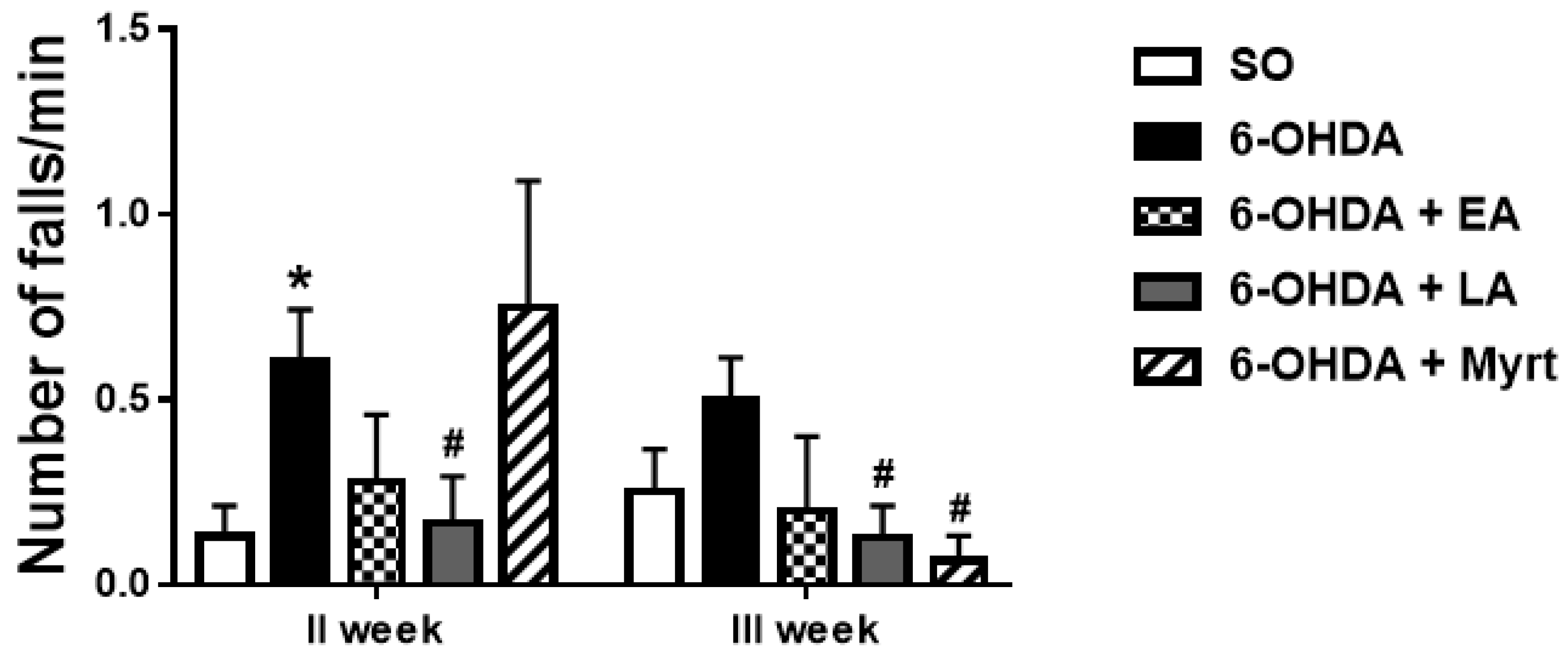
3.2.2. Neuromuscular Coordination (Rotarod Test)
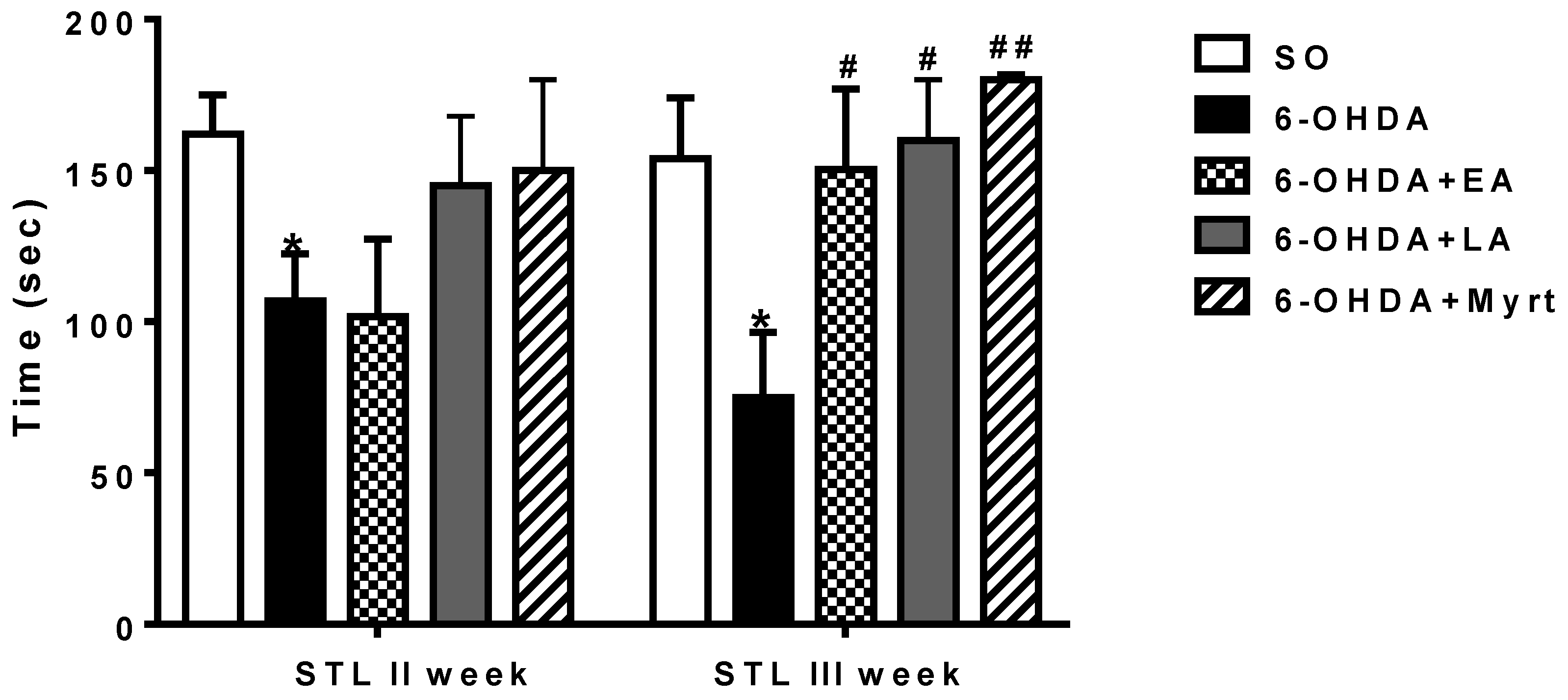
3.2.3. Passive Avoidance Test
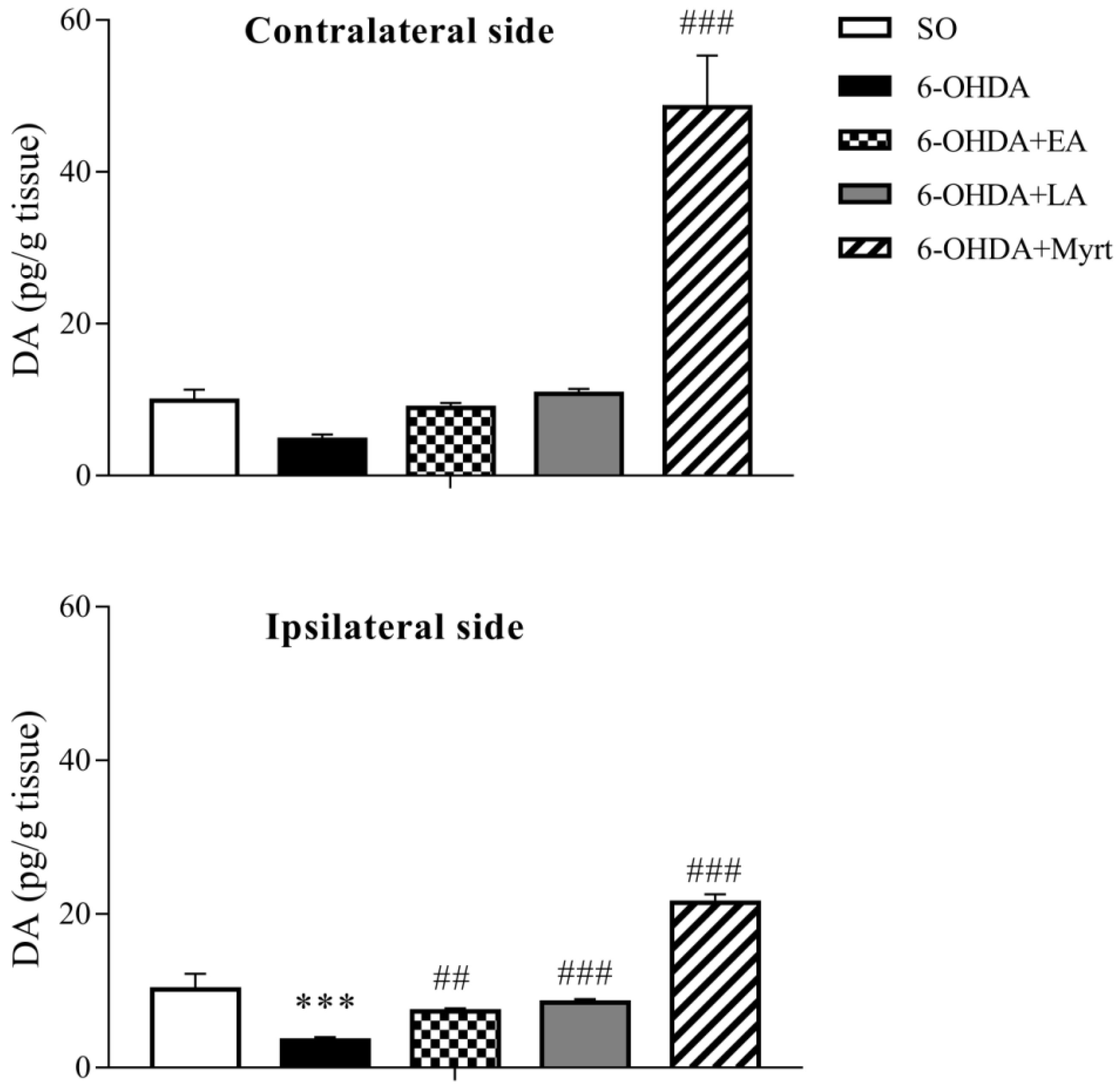
3.3. Modulation of Brain DA Levels by AOs Pretreatment
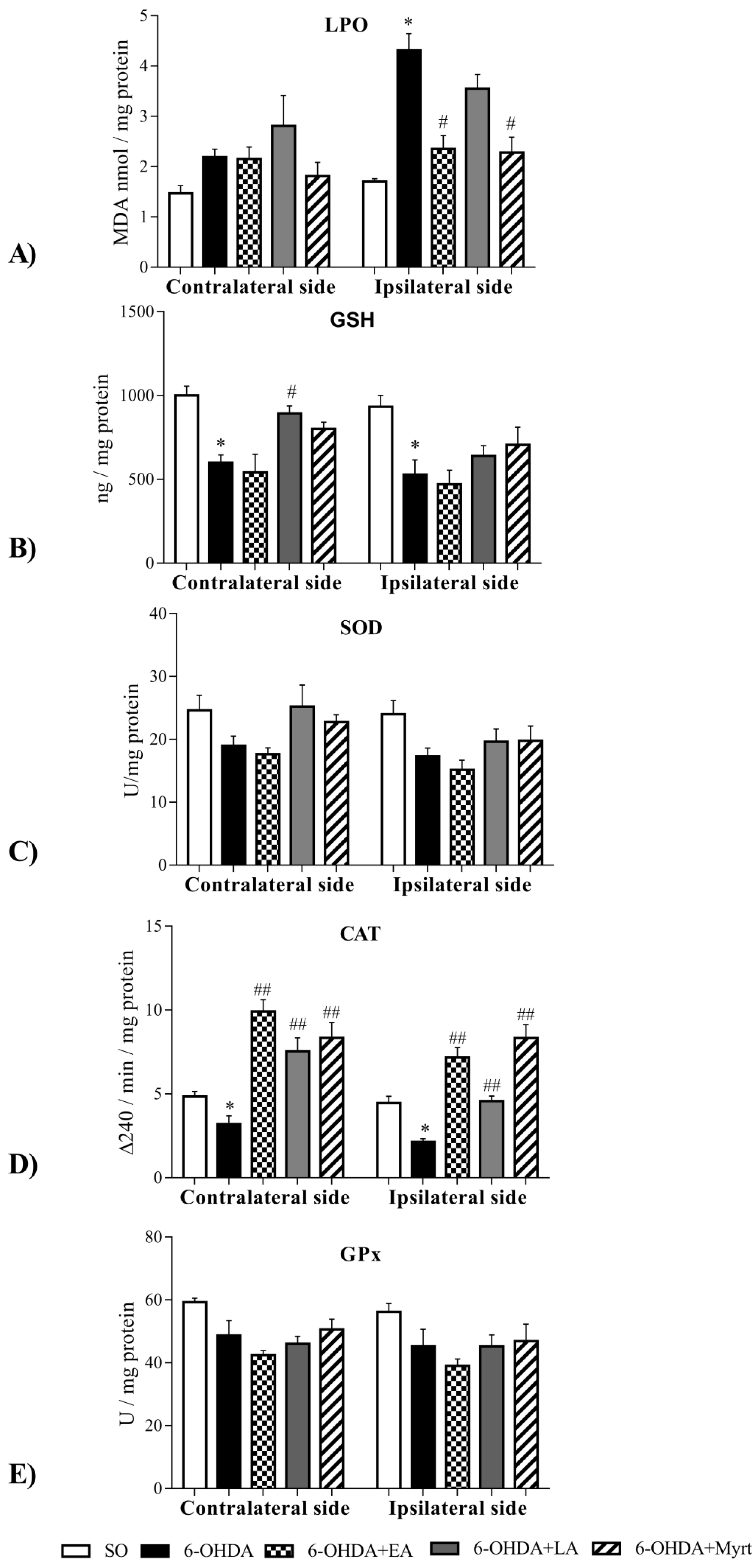
3.4. Effects of the Three Compounds on Parameters of Oxidative Stress in the Brain of PD Rats
3.4.1. Common Antioxidant Effects
3.4.2. Specific Antioxidant Effects
4. Discussion
- The three bio- AOs decreased LPO levels, ipsilateral in the brain and the strength of the effect diminished in the following sequence EA = Myrt > LA;
- LA and Myrt increased GSH brain levels. The effect of the dithiol compound was better and contralaterally located. The monoterpenoid Myrt enhanced GSH both ipsilateral and contralateral effects in the same way.
- (1)
- Significant PD recovery by the three AOs—there is clear evidence for neuroprotective AO effects and differentiation of the treated groups from PD rats.
- (2)
- There are close but yet distinguishable mechanisms of neuroprotection for each of three AOs: LA, EA, and Myrt.
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kalia, L.V.; Lang, A.E. Parkinson’s disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef]
- Michel, P.P.; Toulorge, D.; Guerreiro, S.; Hirsch, E.C. Specific needs of dopamine neurons for stimulation in order to survive: Implication for Parkinson disease. FASEB J. 2013, 27, 3414–3423. [Google Scholar] [CrossRef] [PubMed]
- Rodnitzky, R. Handbook of Therapy in Parkinson’s Disease. Neurology 1995, 45, 1794. [Google Scholar] [CrossRef]
- Adams, J.D., Jr.; Chang, M.L.; Klaidman, L. Parkinson’s disease-redox mechanisms. Curr. Med. Chem. 2001, 8, 809–814. [Google Scholar] [CrossRef]
- Beal, M.F. Does impairment of energy metabolism result in excitotoxic neuronal death in neurodegenerative illnesses. Ann. Neurol. 1992, 31, 119–130. [Google Scholar] [CrossRef]
- Sayre, L.M.; Smith, M.A.; Perry, G. Chemistry and biochemistry of oxidative stress in neurodegenerative diseases. Curr. Med. Chem. 2001, 8, 721–738. [Google Scholar] [CrossRef]
- Zeng, X.S.; Geng, W.S.; Jia, J.J.; Chen, L.; Zhang, P.P. Cellular and Molecular Basis of Neurodegeneration in Parkinson Disease. Front. Aging Neurosci. 2018, 10, 109. [Google Scholar] [CrossRef]
- Zeevalk, G.D.; Razmpour, R.; Bernard, L.P. Glutathione and Parkinson’s disease: Is this the elephant in the room? Biomed. Pharmacother. 2008, 62, 236–249. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Zhou, Z.Z.; Yuan, X.; Cheng, Y.F.; Bi, B.T.; Gong, M.F.; Chen, Y.P.; Xu, J.P. Chlorbipram: A novel PDE4 inhibitor with improved safety as a potential antidepressant and cognitive enhancer. Eur. J. Pharmacol. 2013, 721, 56–63. [Google Scholar] [CrossRef]
- Gazerani, P. Probiotics for Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 4121. [Google Scholar] [CrossRef]
- Yeng, A.W.K.; Georgieva, M.G.; Atanasov, A.G.; Tzvetkov, N.T. Monoamine oxidases (MAOs) as privileged molecular targets in neuroscience: Research literature analysis. Front. Mol. Neurosci. 2019, 20, 143. [Google Scholar] [CrossRef]
- Brown, M.K.; Evans, J.L.; Luo, Y. Beneficial effects of natural antioxidants EGCG and alpha-lipoic acid on life span and age-dependent behavioral declines in Caenorhabditis elegans. Pharmacol. Biochem. Behav. 2006, 85, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Lee, R.; Heber, D. Bioavailability of ellagic acid in human plasma after consumption of ellagitannins from pomegranate (Punica granatum L.) juice. Clin. Chim. Acta 2004, 348, 63–68. [Google Scholar] [CrossRef]
- Shinto, L.; Quinn, J.; Montine, T.; Dodge, H.H.; Woodward, W.; Baldauf-Wagner, S.; Waichunas, D.; Bumgarner, L.; Bourdette, D.; Silbert, L.; et al. A randomized placebo-controlled pilot trial of omega-3 fatty acids and alpha lipoic acid in Alzheimer’s disease. J. Alzheimer’s Dis. 2014, 38, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Khalili, M.; Eghtesadi, S.; Mirshafiey, A.; Eskandari, G.; Sanoobar, M.; Sahraian, M.A.; Motevalian, A.; Norouzi, A.; Moftakhar, S.; Azimi, A. Effect of lipoic acid consumption on oxidative stress among multiple sclerosis patients: A randomized controlled clinical trial. Nutr. Neurosci. 2014, 17, 16–20. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, S.; Wang, F.; Yu, H.; Li, X.; Dong, W.; Lin, R.; Liu, Q. Chronic administration of ellagic acid improved the cognition in middle-aged overweight men. Appl. Physiol. Nutr. Metab. 2017, 43, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Wei, J.; Yang, J.; Xu, J.; Pang, W.; Jiang, Y. Pomegranate juice is potentially better than apple juice in improving antioxidant function in elderly subjects. Nutr. Res. 2008, 28, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Parashar, A.; Badal, S. Pomegranate juice is potentially better than orange juice in improving antioxidant function in elderly subjects. Elixir Food Sci. 2011, 32, 2068–2074. [Google Scholar] [CrossRef] [PubMed]
- Daniel, E.M.; Krupnick, A.S.; Heur, Y.H.; Blinzler, J.A.; Nims, R.W.; Stoner, G.D. Extraction, stability, and quantitation of ellagic acid in various fruits and nuts. J. Food Compos. Anal. 1989, 2, 338–349. [Google Scholar] [CrossRef]
- Raudone, L.; Bobinaite, R.; Janulis, V.; Viskelis, P.; Trumbeckaite, S. Effects of raspberry fruit extracts and ellagic acid on respiratory burst in murine macrophages. Food Funct. 2014, 5, 1167–1174. [Google Scholar] [CrossRef]
- Zeb, A. A reversed phase HPLC-DAD method for the determination of phenolic compounds in plant leaves. Anal. Methods 2015, 7, 7753–7757. [Google Scholar] [CrossRef]
- Kabiraj, P.; Marin, J.E.; Varela-Ramirez, A.; Zubia, E.; Narayan, M. Ellagic acid mitigates SNOPDI induced aggregation of Parkinsonian biomarkers. ACS Chem. Neurosci. 2014, 5, 1209–1220. [Google Scholar] [CrossRef]
- Uzar, E.; Alp, H.; Cevik, M.U.; Firat, U.; Evliyaoglu, O.; Tufek, A.; Altun, Y. Ellagic acid attenuates oxidative stress on brain and sciatic nerve and improves histopathology of brain in streptozotocin-induced diabetic rats. Neurol. Sci. 2012, 33, 567–574. [Google Scholar] [CrossRef]
- Mansouri, M.T.; Farbood, Y.; Naghizadeh, B.; Shabani, S.; Mirshekar, M.A.; Sarkaki, A. Beneficial effects of ellagic acid against animal models of scopolamine- and diazepam induced cognitive impairments. Pharm. Biol. 2016, 54, 1947–1953. [Google Scholar] [CrossRef]
- Tancheva, L.P.; Popatanasov, A.B.; Dragomanova, S.T.; Tzvetanova, E.R.; Aleksandrova, S.M.; Alova, L.G.; Stefanova, S.M.; Kalfin, R.E. New mechanisms in preventive effect of ellagic acid on cognition in mice with Alzheimer’s disease type dementia. Bulg. Chem. Commun. 2018, 50, 20–24. [Google Scholar]
- Nejad, K.H.; Dianat, M.; Sarkaki, A.; Naseri, M.K.; Badavi, M.; Farbood, Y. Ellagic acid improves electrocardiogram waves and blood pressure against global cerebral ischemia rat experimental models. Electron. Physician 2015, 7, 1153–1162. [Google Scholar] [PubMed]
- Farbood, Y.; Sarkaki, A.; Dolatshahi, M.; Taqhi Mansouri, S.M.; Khodadadi, A. Ellagic Acid Protects the Brain Against 6-Hydroxydopamine Induced Neuroinflammation in a Rat Model of Parkinson’s Disease. Basic Clin. Neurosci. 2015, 6, 83–89. [Google Scholar] [PubMed]
- Baluchnejadmojarad, T.; Rabiee, N.; Zabihnejad, S.; Roghani, M. Ellagic acid exerts protective effect in intrastriatal 6-hydroxydopamine rat model of Parkinson’s disease: Possible involvement of ERbeta/Nrf2/HO-1 signaling. Brain Res. 2017, 1662, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Zhang, B.; Zhou, K.; Chen, M.; Wang, M.; Jia, Y.; Song, Y.; Li, Y.; Wen, A. Dietary ellagic acid improves oxidant-induced endothelial dysfunction and atherosclerosis: Role of Nrf2 activation. Int. J. Cardiol. 2014, 175, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Chatterjee, S.; Das, S.; Saha, A.; Chattopadhyay, S.; Bandyopadhyay, S.K. Ellagic acid facilitates indomethacin-induced gastric ulcer healing via COX-2 up-regulation. Acta Biochim. Biophys. Sin. (Shanghai) 2012, 44, 565–576. [Google Scholar] [CrossRef]
- Anderson, K.C.; Teuber, S.S. Ellagic acid and polyphenolics present in walnut kernels inhibit in vitro human peripheral blood mononuclear cell proliferation and alter cytokine production. Ann. N. Y. Acad. Sci. 2010, 1190, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Ahad, A.; Ganai, A.A.; Mujeeb, M.; Siddiqui, W.A. Ellagic acid, an NF-kappaB inhibitor, ameliorates renal function in experimental diabetic nephropathy. Chem. Biol. Interact. 2014, 219, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.C.; Huang, A.C.; Yu, C.S.; Lien, J.C.; Wu, S.H.; Huang, Y.P.; Huang, H.Y.; Kuo, J.H.; Liao, W.Y.; Yang, J.S.; et al. Ellagic acid induces apoptosis in TSGH8301 human bladder cancer cells through the endoplasmic reticulum stress- and mitochondria-dependent signaling pathways. Environ. Toxicol. 2014, 29, 1262–1274. [Google Scholar] [CrossRef] [PubMed]
- Anitha, P.; Priyadarsini, R.V.; Kavitha, K.; Thiyagarajan, P.; Nagini, S. Ellagic acid coordinately attenuates Wnt/beta-catenin and NF-kappaB signaling pathways to induce intrinsic apoptosis in an animal model of oral oncogenesis. Eur. J. Nutr. 2013, 52, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.S.; Vettakkorumakankav, N.N. Lipoic acid-requiring proteins: Recent advances. In Biothiols in Health and Disease; Packer, L., Cadenas, E., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1995; pp. 373–388. [Google Scholar]
- Rosenberg, E.H.R.; Culik, R. Effect of a-lipoic acid on vitamin C and vitamin E deficiencies. Arch. Biochem. Biophys. 1959, 80, 86–93. [Google Scholar] [CrossRef]
- Gegelashvili, G.; Dehnes, Y.; Danbolt, N.C.; Schousboe, A. The high-affinity glutamate transporters GLT1, GLAST, and EAAT4 are regulated via different signalling mechanisms. Neurochem. Int. 2000, 37, 163–170. [Google Scholar] [CrossRef]
- Karki, P.; Webb, A.; Smith, K.; Lee, K.; Son, D.S.; Lee, E. cAMP response element-binding protein (CREB) and nuclear factor κB mediate the tamoxifen-induced up-regulation of glutamate transporter 1 (GLT-1) in rat astrocytes. J. Biol. Chem. 2013, 288, 28975–28986. [Google Scholar] [CrossRef]
- Burgin, A.B.; Magnusson, O.T.; Singh, J.; Witte, P.; Starker, B.L.; Bjornsson, J.M.; Thorsteinsdottir, M.; Hrafnsdottir, S.; Hagen, T.; Kiselyov, A.S.; et al. Design of phosphodiesterase 4D (PDE4D) allosteric modulators for enhancing cognition with improved safety. Nat. Biotechnol. 2010, 28, 63–70. [Google Scholar] [CrossRef]
- Dragomanova, S.; Tancheva, L.P.; Georgieva, M.; Georgieva, A.; Dishovsky, C.; Stoeva, S.; Pavlov, S. Preventive effect of myrtenal and lipoic acid in combination on progression of Alzheimer’s disease. Eur. Neuropsychopharmacol. 2016, 26, S636. [Google Scholar] [CrossRef]
- Fleischhacker, W.W.; Hinterhuber, H.; Bauer, H.; Pflug, B.; Berner, P.; Simhandl, C.; Wolf, R.; Gerlach, W.; Jaklitsch, H.; Sastre-y-Hernandez, M.; et al. A multicenter double-blind study of three different doses of the new cAMP-phosphodiesterase inhibitor rolipram in patients with major depressive disorder. Neuropsychobiology 1992, 26, 59–64. [Google Scholar] [CrossRef]
- Saccomano, N.A.; Vinick, F.J.; Koe, B.K.; Kenneth, B.; Nielsen, J.A.; Whalen, W.M.; Meltz, M.; Phillips, D.; Thadieo, P.F.; Jung, S.; et al. Calcium-independent phospho-diesterase inhibitors as putative antidepressants: [3-(bicycloalkyloxy)-4-methoxyphenyl]-2-imidazolidinones. J. Med. Chem. 1991, 34, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Tzvetanova, E.R.; Georgieva, A.P.; Alexandrova, A.V.; Tancheva, L.P.; Lazarova, M.I.; Dragomanova, S.T.; Alova, L.G.; Stefanova, M.O.; Kalfin, R.E. Antioxidant mechanisms in neuroprotective action of lipoic acid on learning and memory of rats with experimental dementia. Bulg. Chem. Commun. 2018, 50, 52–57. [Google Scholar]
- Guo, J.; Lin, P.; Zhao, X.; Zhang, J.; Wei, X.; Wang, Q.; Wang, C. Etazolate abrogates the lipopolysaccharide (LPS)-induced downregulation of the cAMP/pCREB/BDNF signaling, neuroinflammatory response and depression-like behavior in mice. Neuroscience 2014, 28, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Halene, T.B.; Siegel, S.J. PDE inhibitors in psychiatry—Future options for dementia, depression and schizophrenia? Drug Discov. Today 2007, 12, 870–878. [Google Scholar] [CrossRef]
- Jindal, A.; Mahesh, R.; Bhatt, S. Type 4 phosphodiesterase enzyme inhibitor, rolipram rescues behavioral deficits in olfactory bulbectomy models of depression: Involvement of hypothalamic-pituitary-adrenal axis, cAMP signaling aspects and antioxidant defense system. Pharmacol. Biochem. Behav. 2015, 132, 20–32. [Google Scholar] [CrossRef]
- Mahar, I.; Bambico, F.R.; Mechawar, N.; Nobrega, J.N. Stress, serotonin, and hippocampal neurogenesis in relation to depression and antidepressant effects. Neurosci. Biobehav. Rev. 2014, 38, 173–192. [Google Scholar] [CrossRef]
- Wang, C.; Yang, X.M.; Zhuo, Y.Y.; Zhou, H.; Lin, H.B.; Cheng, Y.F.; Xu, J.P.; Zhang, H.T. The phosphodiesterase-4 inhibitor rolipram reverses Aβ-induced cognitive impairment and neuroinflammatory and apoptotic responses in rats. Int. J. Neuropsychopharmacol. 2012, 15, 749–766. [Google Scholar] [CrossRef]
- Moraghebi, F. Introduction of myrtenal as an inndicator component in essential oil of Cuminum cyminum Isfahan variety. J. Biodivers. Environ. Sci 2013, 3, 112–117. [Google Scholar]
- Smigielski, K.; Raj, A.; Krosowiak, K.; Gruska, R. Chemical composition of the essential oil of Lavandula angustifolia cultivated in poland. J. Essent. Oil Bear. 2009, 12, 338–347. [Google Scholar] [CrossRef]
- Hari Babu, L.; Perumal, S.; Balasubramanian, M.P. Myrtenal, a natural monoterpene, down-regulates TNF-α expression and suppresses carcinogen-in-duced hepatocellular carcinoma in rats. Mol. Cell. Biochem. 2012, 369, 183–193. [Google Scholar] [CrossRef]
- Begum, S.; Ali, M.; Gul, H.; Ahmad, W.; Alam, S.; Khan, M.; Khan, M.A.; Ahmad, M. In vitro enzyme inhibition activities of Myrtus communis L. Afr. J. Pharm. Pharmacol. 2012, 6, 1083–1087. [Google Scholar]
- Venkatachalam, S.; Boobathi, L.; Balasubramanian, M.P. Salubrious therapeutic efficacy of myrte¬nal on colon carcinoma induced by 1,2-dimethyl-hydrazine studied in experimental albino rats. Res. J. Pharmacol. Pharmacodyn. 2014, 6, 146–152. [Google Scholar]
- Gheibi, S.; Aboutaleb, N.; Khaksari, M.; Kalalian-Moghaddam, H.; Vakili, A.; Asadi, Y.; Mehrjerdi, F.Z.; Gheibi, A. Hydrogen sulfide protects the brain against ischemic reperfusion injury in a transient model of focal cerebral ischemia. J. Mol. Neurosci. 2014, 54, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press, Inc.: Cambridge, MA, USA, 1997. [Google Scholar]
- García, J.C.; Remires, D.; Leiva, A.; González, R. Depletion of brain glutathione potentiates the effect of 6-hydroxydopamine in a rat model of Parkinson’s disease. J. Mol. Neurosci. 2000, 14, 147–153. [Google Scholar] [CrossRef]
- Dragomanova, S.; Tancheva, L.; Georgieva, M.; Georgieva, A.; Dishovsky, C.; Kalfin, R.; Pehlivanov, I. P. 5. a. 005 Preventive effect of natural monoterpen Myrtenal on cognitive disorders in dement mice. Eur. Neuropsychopharmacol. 2015, 25, S578–S579. [Google Scholar] [CrossRef]
- Biewenga, G.P.; Haenen, G.R.; Bast, A. The pharmacology of the antioxidant lipoic acid. Gen. Pharmacol. 1997, 29, 315–331. [Google Scholar] [CrossRef]
- Kaufmann, D.; Dogra, A.K.; Wink, M. Myrtenal inhibits acetylcholinesterase, a known Alzheimer target. J. Pharm. Pharmacol. 2011, 63, 1368–1371. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef]
- Roghani, M.; Niknam, A.; Jalali-Nadoushan, M.R.; Kiasalari, Z.; Khalili, M.; Baluchnejadmojarad, T. Oral pelargonidin exerts dose-dependent neuroprotection in 6-hydroxydopamine rat model of hemi-parkinsonism. Brain Res. Bull. 2010, 82, 279–283. [Google Scholar] [CrossRef]
- Rozas, G.; Guerra, M.J.; Labandeira Garcia, J.L. An automated Rotarod method for quantitative drug-free evaluation of overall motor deficits in rat models of Parkinsonism. Brain Res. Protoc. 1997, 2, 75–84. [Google Scholar] [CrossRef]
- Jarvik, M.; Kopp, R. An improved one-trial passive avoidance learning situation. Psychol. Rep. 1967, 21, 221–224. [Google Scholar] [CrossRef]
- Jacobowitz, D.M.; Richardson, J.S. Method for the rapid determination of norepinephrine, dopamine, and serotonin in the same brain region. Pharmacol. Biochem. Behav. 1978, 8, 515–519. [Google Scholar] [CrossRef]
- Sauer, H.; Oertel, W.H. Progressive degeneration of nigrostriatal dopamine neurons following intrastriatal terminal lesions with 6-hydroxydopamine: A combined retrograde tracing and immunocytochemical study in the rat. Neuroscience 1994, 59, 401–415. [Google Scholar] [CrossRef]
- Przedborski, S.; Levivier, M.; Jiang, H.; Ferreira, M.; Jackson-Lewis, V.; Donaldson, D.; Togasaki, D.M. Dose-dependent lesions of the dopaminergic nigrostriatal pathway induced by intrastriatal injection of 6-hydroxydopamine. Neuroscience 1995, 67, 631–647. [Google Scholar] [CrossRef]
- Blandini, F.; Levandis, G.; Bazzini, E.; Nappi, G.; Armentero, M.T. Time-course of nigrostriatal damage, basal ganglia metabolic changes and behavioural alterations following intrastriatal injection of 6-hydroxydopamine in the rat: New clues from an old model. Eur. J. Neurosci. 2007, 25, 397–405. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randal, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Gower, J.C. Measures of similarity, dissimilarity, and distance. Encycl. Stat. Sci. 1985, 5, 397–405. [Google Scholar]
- Lombardo, S.D.; Presti, M.; Mangano, K.; Petralia, M.C.; Basile, M.S.; Libra, M.; Candido, S.; Fagone, P.; Mazzon, E.; Nicoletti, F.; et al. Prediction of PD-L1 expression in neuroblastoma via computational modeling. Brain Sci. 2019, 9, 221. [Google Scholar] [CrossRef]
- Presti, M.; Mazzon, E.; Basile, M.S.; Petralia, M.C.; Bramanti, A.; Colletti, G.; Bramanti, P.; Nicoletti, F.; Fagone, P. Overexpression of macrophage migration inhibitory factor and functionally-related genes, D-DT, CD74, CD44, CXCR2 and CXCR4, in glioblastoma. Oncol. Lett. 2018, 16, 2281–2286. [Google Scholar] [CrossRef]
- Mammana, S.; Bramanti, P.; Mazzon, E.; Cavalli, E.; Basile, M.S.; Fagone, P.; Petralia, M.C.; McCubrey, J.A.; Nicoletti, F.; Mangano, K. Preclinical evaluation of the PI3K/Akt/mTOR pathway in animal models of multilple sclerosis. Oncotarget 2018, 9, 8262–8277. [Google Scholar] [CrossRef]
- Fagone, P.; Mazzon, E.; Mammana, S.; Di Marco, R.; Spinasanta, F.; Basile, M.S.; Petralia, M.C.; Bramanti, P.; Nicoletti, F.; Mangano, K. Identification of CD4+ T cell biomarkers for predicting the response of patients with relapsing-remitting multiple sclerosis to natalizumab treatment. Mol. Med. Rep. 2019, 20, 678–684. [Google Scholar] [CrossRef]
- Nicoletti, F.; Mazzon, E.; Fagone, P.; Mangano, K.; Mammana, S.; Cavalli, E.; Basile, M.S.; Bramanti, P.; Scalabrino, G.; Lange, A.; et al. Prevention of clinical and histological signs of MOG-induced experimental allergic encephalomyelitis by prolonged treatment with recombinant human EGF. J. Neuroimmunol. 2019, 332, 224–232. [Google Scholar] [CrossRef]
- Ma, S.; Dai, Y. Principal component analysis based methods in bioinformatics studies. Brief. Bioinform. 2011, 12, 714–722. [Google Scholar] [CrossRef]
- Loeffler, D.A.; Klaver, A.C.; Coffey, M.P.; Aasly, J.O.; LeWitt, P.A. Increased Oxidative Stress Markers in Cerebrospinal Fluid from Healthy Subjects with Parkinson’s Disease-Associated LRRK2 Gene Mutations. Front. Aging Neurosci. 2017, 9, 89. [Google Scholar] [CrossRef]
- Toulorge, D.; Schapira, A.H.; Hajj, R. Molecular changes in the postmortem parkinsonian brain. J. Neurochem. 2016, 139, 27–58. [Google Scholar] [CrossRef]
- Cebrián, C.; Loike, J.D.; Sulzer, D. Neuroinflammation in Parkinson’s disease animal models: A cell stress response or a step in neurodegeneration? Curr. Top. Behav. Neurosci. 2015, 22, 237–270. [Google Scholar]
- Jenner, P. Altered mitochondrial function, iron metabolism and glutathione levels in Parkinson’s disease. Acta Neurol. Scand. Supp. 1993, 146, 6–13. [Google Scholar]
- Chandrasekhar, Y.; Kumar, G.P.; Ramya, E.M.; Anilakumar, K.R. Gallic Acid Protects 6-OHDA Induced Neurotoxicity by Attenuating Oxidative Stress in Human Dopaminergic Cell Line. Neurochem. Res. 2018, 43, 1150. [Google Scholar] [CrossRef]
- Doaee, P.; Rajaei, Z.; Roghani, M.; Alaei, H.; Kamalinejad, M. Effects of Boswellia serrata resin extract on motor dysfunction and brain oxidative stress in an experimental model of Parkinson’s disease. Avicenna J. Phytomed. 2019, 9, 281–290. [Google Scholar]
- Connell, B.J.; Khan, B.V.; Rajagopal, D.; Saleh, T.M. Novel Neurovascular Protective Agents: Effects of INV-155, INV-157, INV-159, and INV-161 versus Lipoic Acid and Captopril in a Rat Stroke Model. Cardiol. Res. Pract. 2012, 2012, 319230. [Google Scholar] [CrossRef]
- Fujita, H.; Shiosaka, M.; Ogino, T.; Okimura, Y.; Utsumi, T.; Sato, E.F.; Akagi, R.; Inoue, M.; Utsumi, K.; Sasaki, J. α-Lipoic acid suppresses 6-hydroxydopamine-induced ROS generation and apoptosis through the stimulation of glutathione synthesis but not by theexpression of heme oxygenase-1. Brain Res. 2008, 1206, 1–12. [Google Scholar] [CrossRef][Green Version]
- Sarkaki, A.; Farbood, Y.; Dolatshahi, M.; Mansouri, S.M.; Khodadadi, A. Neuroprotective Effects of Ellagic Acid in a Rat Model of Parkinson’s Disease. Acta Med. Iran. 2016, 54, 494–502. [Google Scholar]
- Dragomanova, S.; Tancheva, L.; Georgieva, M. A review: Biological activity of myrtenal and some myrtenal-containing medicinal plant essential oils. Scr. Sci. Pharm. 2018, 5, 22–33. [Google Scholar]
- Cohen, G.; Heikkila, R.E. The generation of hydrogen peroxide, superoxide radical, and hydroxyl radical by 6-hydroxydopamine, dialuric acid, and related cytotoxic agents. J. Biol. Chem. 1974, 249, 2447–2452. [Google Scholar]
- Kostrzewa, R.M.; Jacobowitz, D.M. Pharmacological actions of 6-hydroxydopamine. Pharmacol. Rev. 1974, 26, 199–288. [Google Scholar]
- Simola, N.; Morelli, M.; Carta, A.R. The 6-hydroxydopaminemodel of Parkinson’s disease. Neurotox. Res. 2007, 11, 151–167. [Google Scholar] [CrossRef]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.; Benabid, A.L.; Sadoul, R.; Verna, J.M. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef]
- Cadet, J.L.; Brannock, C. Free radicals and the pathobiology of brain dopamine systems. Neurochem. Int. 1998, 32, 117–131. [Google Scholar] [CrossRef]
- Glinka, Y.; Tipton, K.F.; Youdim, M.B. Nature of inhibition of mitochondrial respiratory complex I by 6-Hydroxydopamine. J. Neurochem. 1996, 66, 2004–2010. [Google Scholar] [CrossRef] [PubMed]
- Glinka, Y.; Gassen, M.; Youdim, M.B. Mechanism of 6-hydroxydopamine neurotoxicity. J. Neural Transm. 1997, 50, 55–66. [Google Scholar]
- Glinka, Y.; Youdim, M.B. Inhibition of mitochondrial complexes I and IV by 6-hydroxydopamine. Eur. J. Pharmacol. 1995, 292, 329–332. [Google Scholar] [CrossRef]
- Fricke, I.B.; Viel, T.; Worlitzer, M.M.; Collmann, F.M.; Vrachimis, A.; Faust, A.; Wachsmuth, L.; Faber, C.; Dollé, F.; Kuhlmann, M.T.; et al. 6-hydroxydopamine-induced Parkinson’s disease-like degeneration generates acute microgliosis and astrogliosis in the nigrostriatal system but no bioluminescence imaging-detectable alteration in adult neurogenesis. Eur. J. Neurosci. 2016, 43, 1352–1365. [Google Scholar] [CrossRef]
- Bove, J.; Perier, C. Neurotoxin-Based Models of Parkinson’s Disease. Neuroscience 2012, 211, 51–76. [Google Scholar] [CrossRef]
- Monville, C.; Torres, E.M.; Dunnett, S.B. Comparison of incremental and accelerating protocols of the rotarod test for the assessment of motor deficits in the 6-OHDA model. J. Neurosci. Methods 2006, 158, 219–223. [Google Scholar] [CrossRef]
- Ungerstedt, U. Postsynaptic supersensitivity after 6-hydroxy-dopamine induced degeneration of the nigro-striatal dopamine system. Acta Physiol. Scand. Suppl. 1971, 367, 69–93. [Google Scholar] [CrossRef]
- Jalali-Nadoushan, M.; Roghani, M. Alpha-lipoic acid protects against 6-hydroxydopamine-induced neurotoxicity in a rat model of hemi-parkinsonism. Brain Res. 2013, 1505, 68–74. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Development of Alzheimer-related neurofibrillary changes in the neocortex inversely recapitulates cortical myelogenesis. Acta Neuropathol. 1996, 92, 197–201. [Google Scholar] [CrossRef]
- Pinho, B.R.; Ferreres, F.; Valentão, P.; Andrade, P.B. Natural cholinesterase inhibitors. J. Pharm. Pharmacol. 2013, 65, 1681–1700. [Google Scholar] [CrossRef]
- Hald, A.; Lotharius, J. Oxidative stress and inflammation in Parkinson’s disease: Is there a causal link? Exp. Neurol. 2005, 193, 279–290. [Google Scholar] [CrossRef]
- Zhao, H.; Zhao, X.; Liu, L.; Zhang, H.; Xuan, M.; Guo, Z.; Wang, H.; Liu, C. Neurochemical effects of the R form of α-lipoic acid and its neuroprotective mechanism in cellular models of Parkinson’s disease. Int. J. Biochem. Cell B 2017, 87, 86–94. [Google Scholar] [CrossRef]
- Lazarova, M.; Popatanasov, A.; Klissurov, R.; Stoeva, S.; Pajpanova, T.; Kalfin, R.; Tancheva, L. Preventive effect of two new neurotensin analogues on Parkinson’s disease rat models. J. Mol. Neurosci. 2018, 66, 552–560. [Google Scholar] [CrossRef]
- Hritcu, L.; Ciobica, A.; Artenie, V. Effects of right-unilateral 6-hydroxydopamine infusion-induced memory impairment and oxidative stress: Relevance for Parkinson’s disease. Open Life Sci. 2008, 3, 250–257. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Rev. Mol. Nutt. Food Res. 2016, 61, 1500901. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters 1 | PC1 | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|
| DA-L | 0.60 | −0.34 | 0.64 | −0.01 | −0.18 |
| DA-D | 0.69 | −0.37 | 0.49 | 0.06 | −0.11 |
| LPO-L | −0.29 | −0.18 | 0.04 | 0.76 | 0.21 |
| LPO-D | −0.72 | 0.46 | 0.27 | 0.40 | 0.02 |
| GSH-L | 0.72 | 0.54 | −0.14 | 0.32 | 0.01 |
| GSH-D | 0.74 | 0.48 | −0.07 | −0.03 | 0.14 |
| CAT-L | 0.31 | −0.86 | −0.29 | 0.11 | −0.11 |
| CAT-D | 0.57 | −0.79 | 0.02 | −0.09 | −0.08 |
| SOD-D | 0.54 | 0.45 | −0.23 | 0.49 | −0.26 |
| SOD-L | 0.56 | 0.58 | −0.05 | 0.24 | −0.34 |
| GPx-L | 0.61 | 0.58 | 0.16 | −0.26 | 0.19 |
| GPx-D | 0.29 | 0.76 | −0.18 | −0.36 | −0.10 |
| Brain Weight | −0.35 | −0.32 | −0.67 | 0.15 | 0.11 |
| RR-Week 2 | 0.12 | −0.11 | 0.74 | 0.31 | 0.23 |
| RR-Week 3 | −0.52 | 0.31 | 0.18 | −0.36 | −0.07 |
| Apo-Week 2 | −0.58 | 0.38 | 0.18 | 0.15 | 0.19 |
| Apo-Week 3 | −0.68 | 0.40 | 0.32 | −0.01 | −0.32 |
| Component coefficients | 5.51 | 4.39 | 2.15 | 1.71 | 1.15 |
| Variance explained | 29.01 | 23.09 | 11.31 | 8.98 | 6.04 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tancheva, L.P.; Lazarova, M.I.; Alexandrova, A.V.; Dragomanova, S.T.; Nicoletti, F.; Tzvetanova, E.R.; Hodzhev, Y.K.; Kalfin, R.E.; Miteva, S.A.; Mazzon, E.; et al. Neuroprotective Mechanisms of Three Natural Antioxidants on a Rat Model of Parkinson’s Disease: A Comparative Study. Antioxidants 2020, 9, 49. https://doi.org/10.3390/antiox9010049
Tancheva LP, Lazarova MI, Alexandrova AV, Dragomanova ST, Nicoletti F, Tzvetanova ER, Hodzhev YK, Kalfin RE, Miteva SA, Mazzon E, et al. Neuroprotective Mechanisms of Three Natural Antioxidants on a Rat Model of Parkinson’s Disease: A Comparative Study. Antioxidants. 2020; 9(1):49. https://doi.org/10.3390/antiox9010049
Chicago/Turabian StyleTancheva, Lyubka P., Maria I. Lazarova, Albena V. Alexandrova, Stela T. Dragomanova, Ferdinando Nicoletti, Elina R. Tzvetanova, Yordan K. Hodzhev, Reni E. Kalfin, Simona A. Miteva, Emanuela Mazzon, and et al. 2020. "Neuroprotective Mechanisms of Three Natural Antioxidants on a Rat Model of Parkinson’s Disease: A Comparative Study" Antioxidants 9, no. 1: 49. https://doi.org/10.3390/antiox9010049
APA StyleTancheva, L. P., Lazarova, M. I., Alexandrova, A. V., Dragomanova, S. T., Nicoletti, F., Tzvetanova, E. R., Hodzhev, Y. K., Kalfin, R. E., Miteva, S. A., Mazzon, E., Tzvetkov, N. T., & Atanasov, A. G. (2020). Neuroprotective Mechanisms of Three Natural Antioxidants on a Rat Model of Parkinson’s Disease: A Comparative Study. Antioxidants, 9(1), 49. https://doi.org/10.3390/antiox9010049