3,3′-Diindolylmethane Promotes BDNF and Antioxidant Enzyme Formation via TrkB/Akt Pathway Activation for Neuroprotection against Oxidative Stress-Induced Apoptosis in Hippocampal Neuronal Cells

 , ,
, , 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Behavior Tests
2.3.1. Morris Water Maze Test
2.3.2. Passive Avoidance Test
2.4. Analysis of Antioxidant or Neurobiological Biomarkers in Brain Tissues
2.4.1. Determination of Lipid Peroxidation
2.4.2. Measurement of Glutathione
2.4.3. Activities of Glutathione Reductase and Glutathione Peroxidase
2.4.4. Activities of Acetylcholinesterase and Choline Acetyltransferase
2.5. Hematoxylin and Eosin Staining Assay
2.6. Cell Culture
2.7. Cell Viability Assay
2.8. Measurement of Intracellular ROS Level
2.9. Extraction of Nuclear and Cytosolic Protein
2.10. Immunoblot Analysis
2.11. Statistical Analysis
3. Results
3.1. Neuroprotective Effect of 3,3′-Diindolylmethane on Glutamate-Treated HT-22 Cells
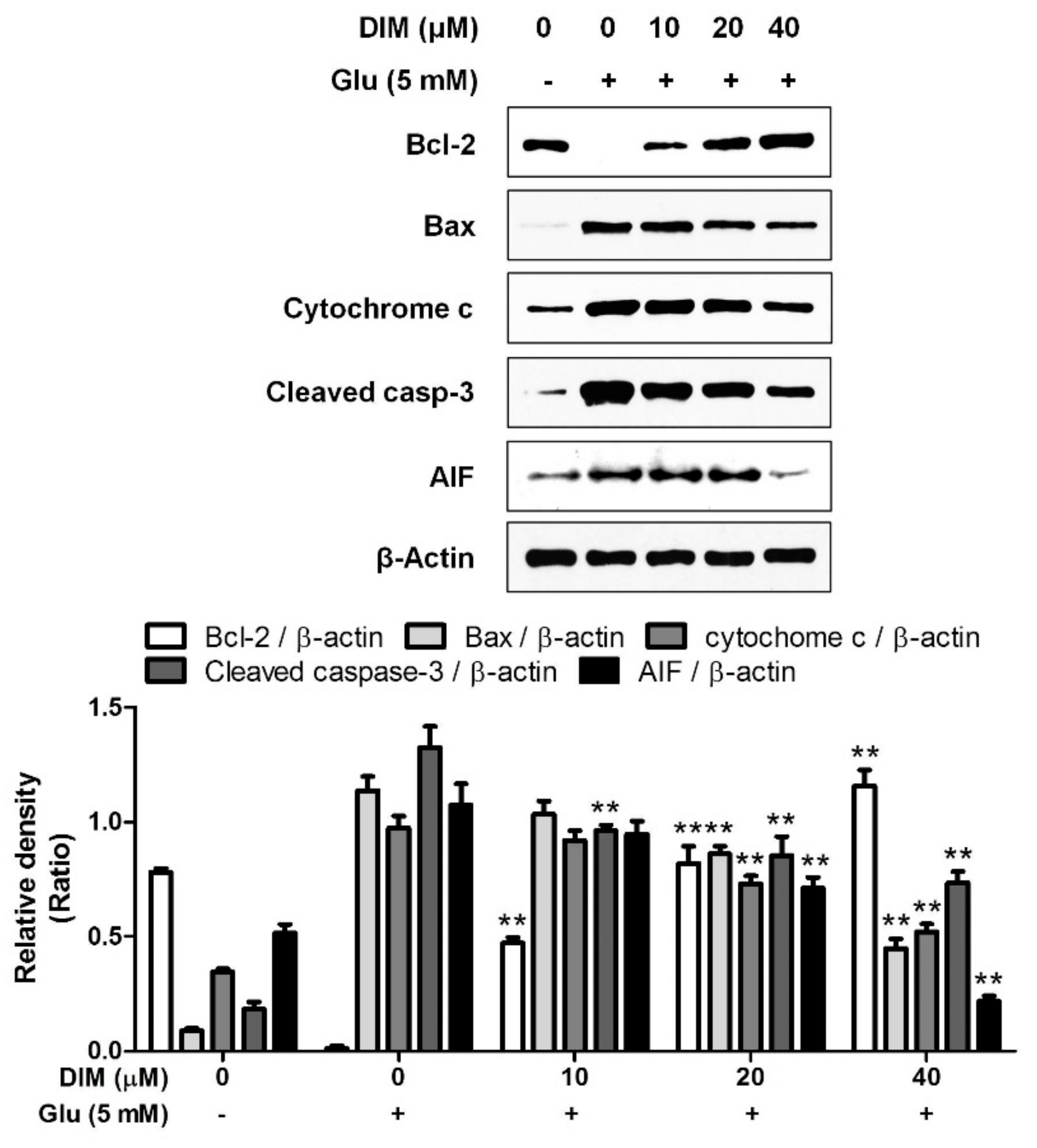
3.2. Inhibitory Effect of 3,3′-Diindolylmethane on Oxidative Stress-Induced Apoptosis
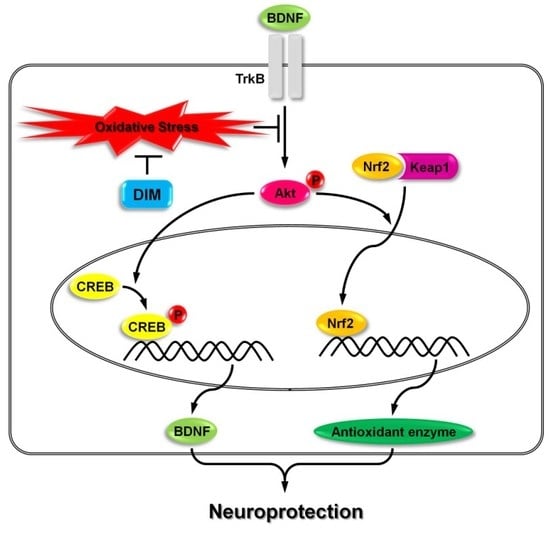
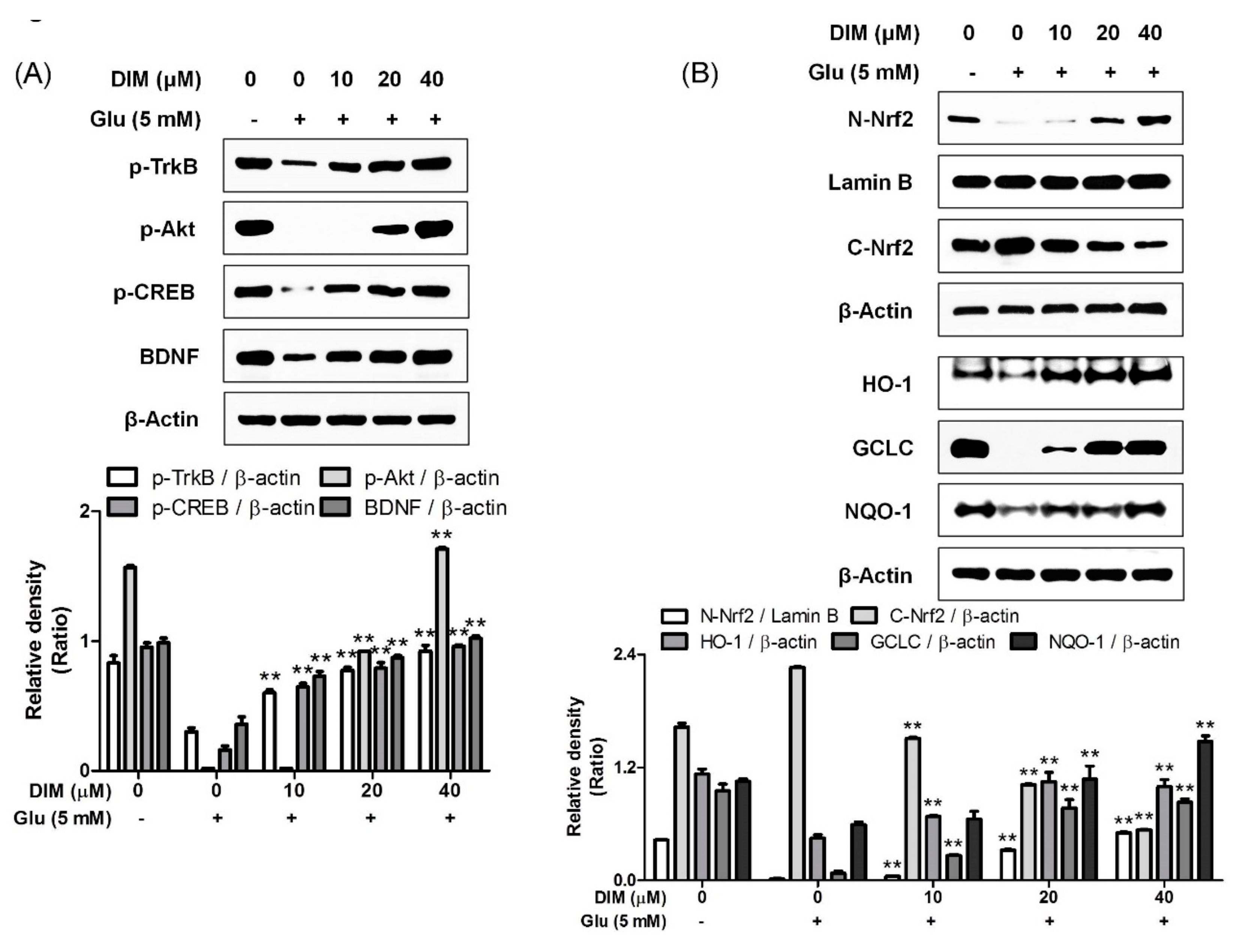
3.3. Activatory Effect of 3,3′-Diindolylmethane on Both TrkB/CREB/BDNF Pathway and Akt/Nrf2/ARE Pathway
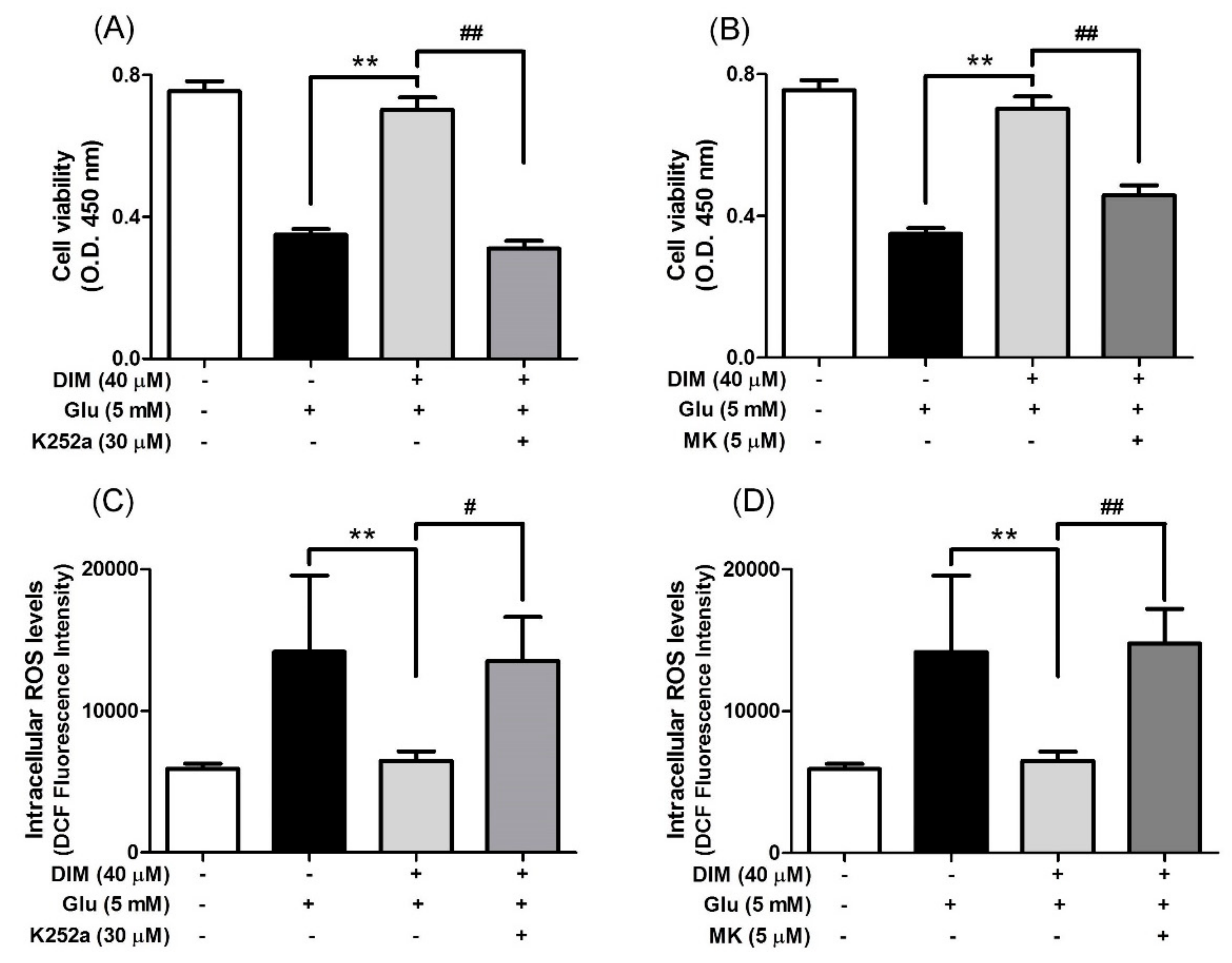
3.4. Suppressive Effects of K252a and MK-2206 on Neuroprotective Action of 3,3′-Diindolylmethane
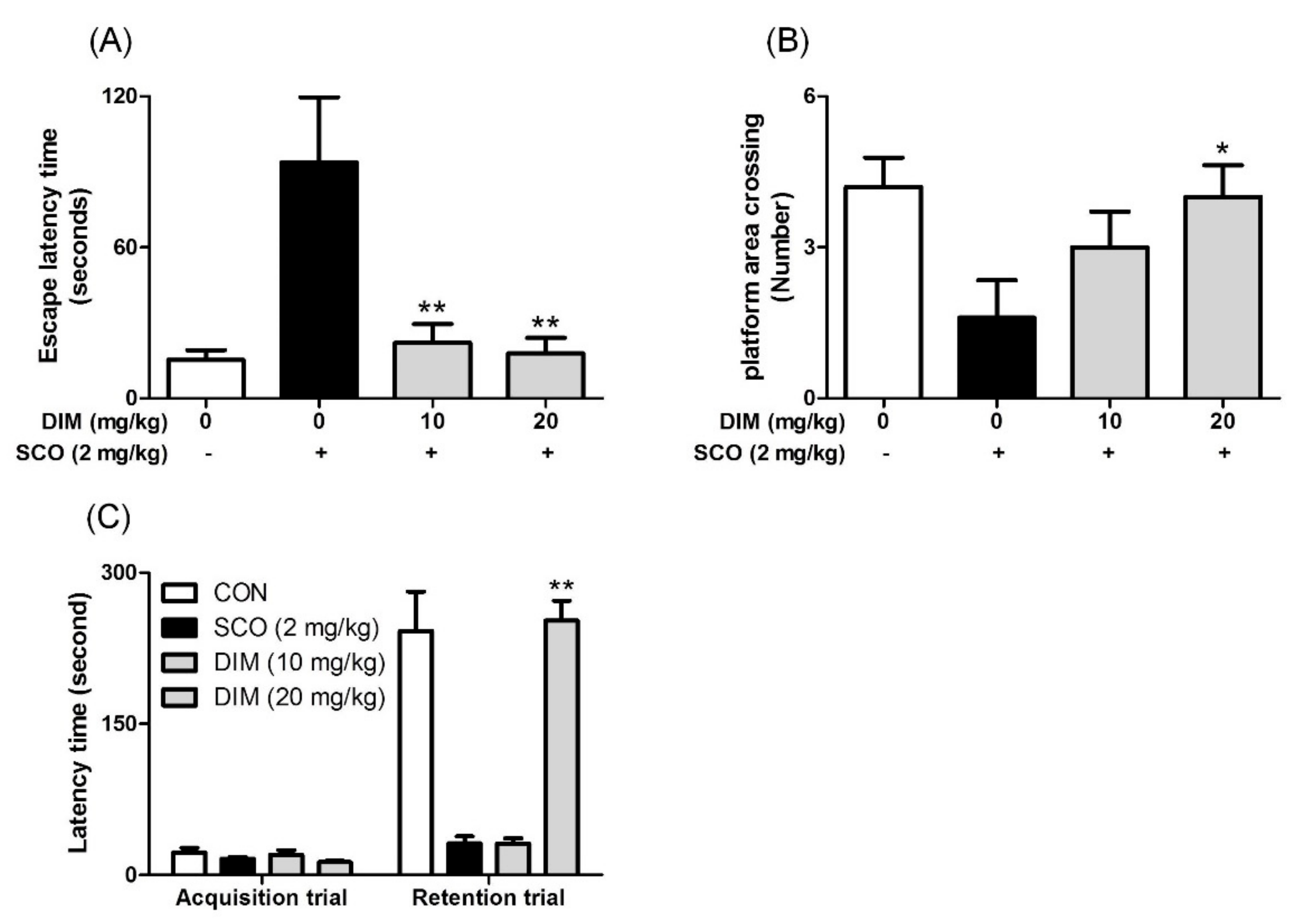
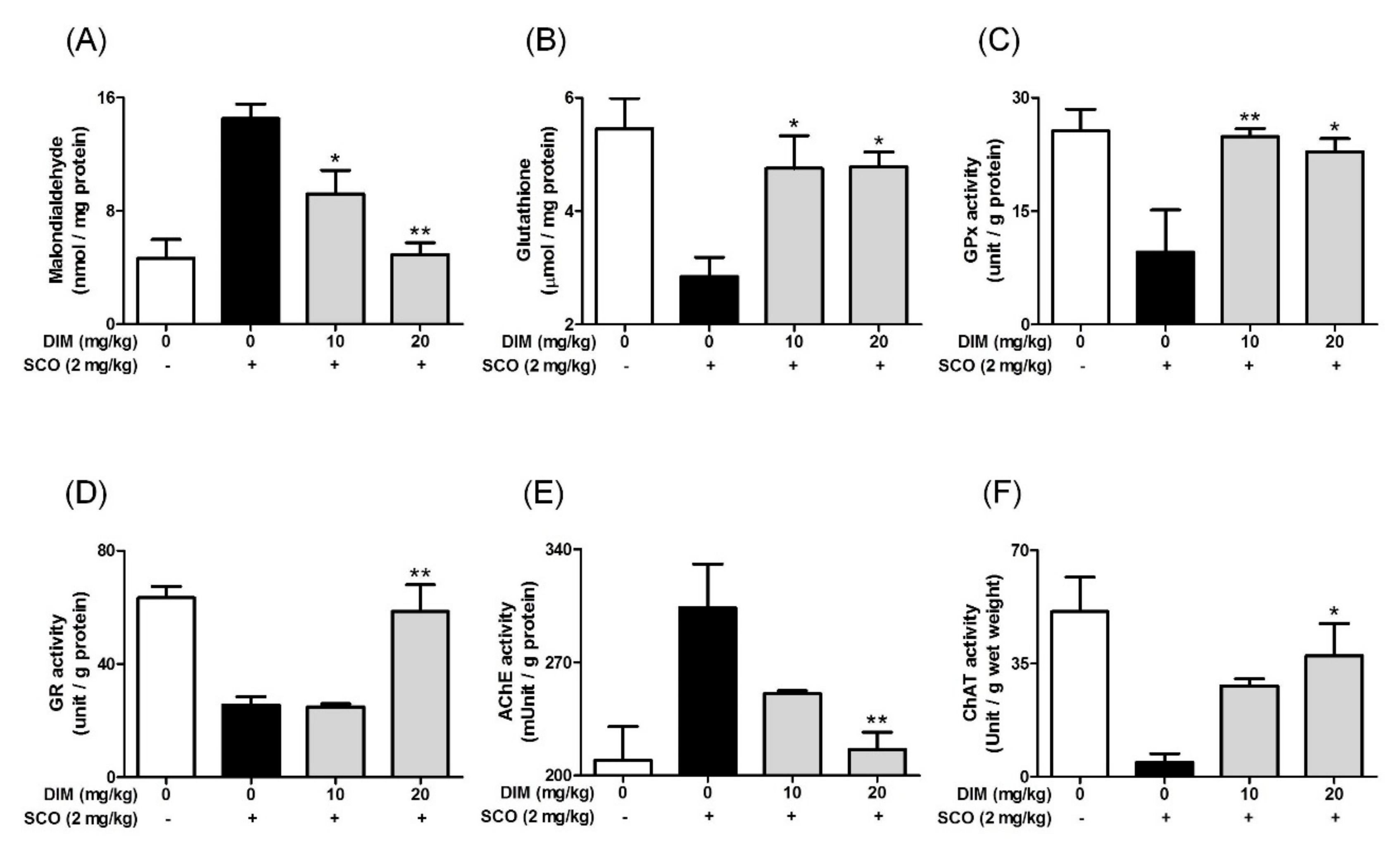
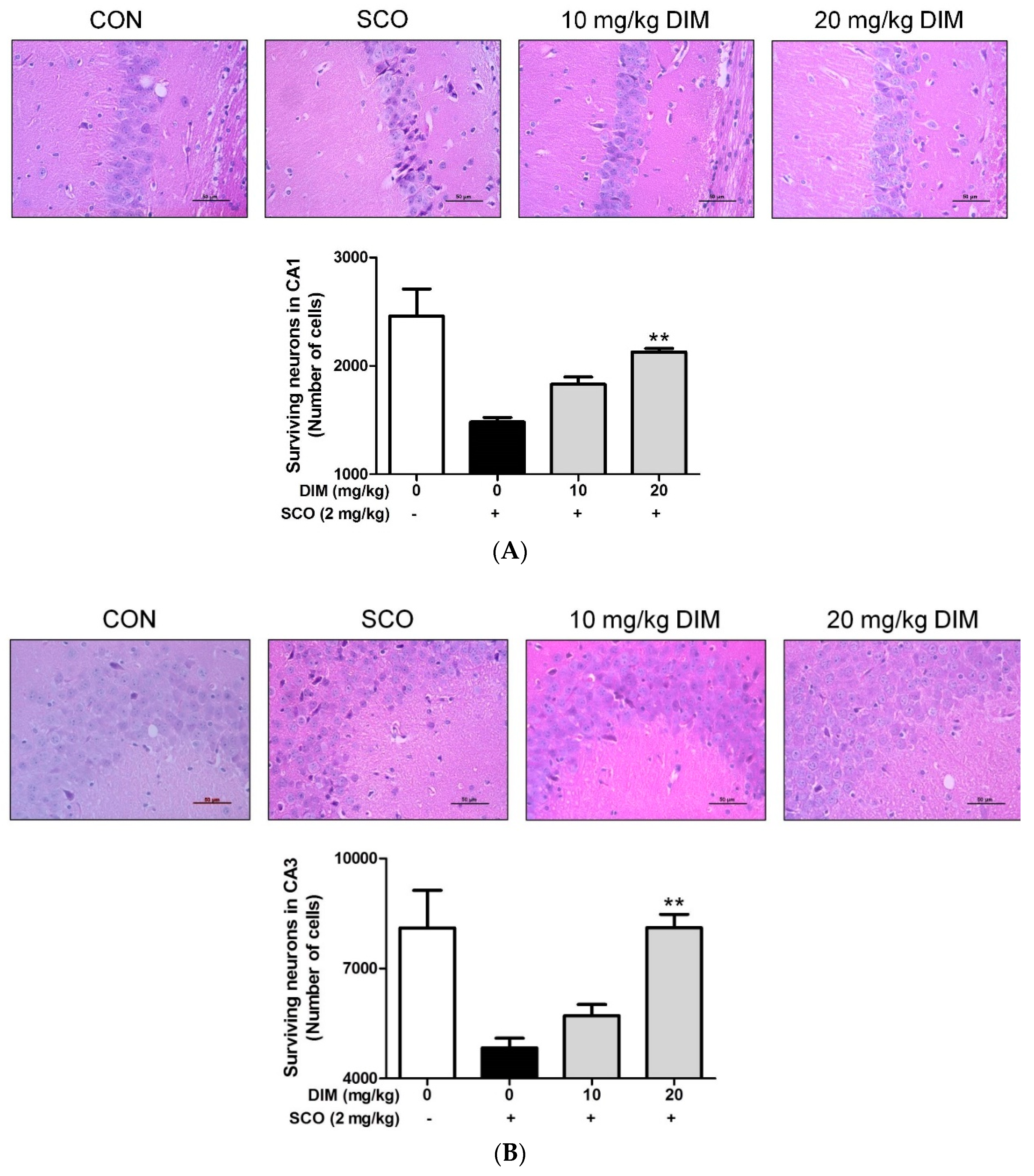
3.5. Improving Effect of 3,3′-Diindolylmethane on Scopolamine-Induced Memory Impairment in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- McNaughton, S.A.; Marks, G.C. Development of a food composition database for the estimation of dietary intakes of glucosinolates, the biologically active constituents of cruciferous vegetables. Br. J. Nutr. 2003, 90, 687–697. [Google Scholar] [CrossRef]
- Higdon, J.V.; Delage, B.; Williams, D.E.; Dashwood, R.H. Cruciferous vegetables and human cancer risk: Epidemiologic evidence and mechanistic basis. Pharmacol. Res. 2007, 55, 224–236. [Google Scholar] [CrossRef]
- Zhang, W.W.; Feng, Z.; Narod, S.A. Multiple therapeutic and preventive effects of 3,3’-diindolylmethane on cancers including prostate cancer and high grade prostatic intraepithelial neoplasia. J. Biomed. Res. 2014, 28, 339–348. [Google Scholar]
- Fuentes, F.; Paredes-Gonzalez, X.; Kong, A.N. Dietary glucosinolates sulforaphane, phenethyl isothiocyanate, indole-3-carbinol/3,3’-diindolylmethane: Anti-oxidative stress/inflammation, nrf2, epigenetics/epigenomics and in vivo cancer chemopreventive efficacy. Curr. Pharmacol. Rep. 2015, 1, 179–196. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Ahmad, A.; Bao, B.; Sarkar, F.H. Antioxidant function of isoflavone and 3,3’-diindolylmethane: Are they important for cancer prevention and therapy? Antioxid. Redox Signal. 2013, 19, 139–150. [Google Scholar] [CrossRef]
- Wang, S.Q.; Cheng, L.S.; Liu, Y.; Wang, J.Y.; Jiang, W. Indole-3-carbinol (i3c) and its major derivatives: Their pharmacokinetics and important roles in hepatic protection. Curr. Drug Metab. 2016, 17, 401–409. [Google Scholar] [CrossRef]
- Rzemieniec, J.; Litwa, E.; Wnuk, A.; Lason, W.; Krzeptowski, W.; Kajta, M. Selective aryl hydrocarbon receptor modulator 3,3’-diindolylmethane impairs ahr and arnt signaling and protects mouse neuronal cells against hypoxia. Mol. Neurobiol. 2016, 53, 5591–5606. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Kim, J.; Kim, J.; Lee, S.; Choi, B.R.; Han, J.S.; Lee, K.W.; Lee, H.J. 3,3′-diindolylmethane inhibits lipopolysaccharide-induced microglial hyperactivation and attenuates brain inflammation. Toxicol. Sci. Off. J. Society Toxicol. 2014, 137, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Reed, G.A.; Sunega, J.M.; Sullivan, D.K.; Gray, J.C.; Mayo, M.S.; Crowell, J.A.; Hurwitz, A. Single-dose pharmacokinetics and tolerability of absorption-enhanced 3,3′-diindolylmethane in healthy subjects. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2008, 17, 2619–2624. [Google Scholar] [CrossRef] [PubMed]
- Tarafdar, A.; Pula, G. The role of nadph oxidases and oxidative stress in neurodegenerative disorders. Int. J. Mol. Sci. 2018, 19, 3824. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-M.; Lee, B.D.; Lee, S.J.; Ma, J.Y.; Kim, M.R. Anti-apoptotic effect of n-palmitoyl serotonin on glutamate-mediated apoptosis through secretion of bdnf and activation of trkb/creb pathway in ht-22 cells. Eur. J. Lipid Sci. Technol. 2018, 120, 1700397. [Google Scholar] [CrossRef]
- Yoo, J.M.; Lee, B.D.; Sok, D.E.; Ma, J.Y.; Kim, M.R. Neuroprotective action of n-acetyl serotonin in oxidative stress-induced apoptosis through the activation of both trkb/creb/bdnf pathway and akt/nrf2/antioxidant enzyme in neuronal cells. Redox Biol. 2017, 11, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Zafra, F.; Lindholm, D.; Castren, E.; Hartikka, J.; Thoenen, H. Regulation of brain-derived neurotrophic factor and nerve growth factor mrna in primary cultures of hippocampal neurons and astrocytes. J. Neurosci. Off. J. Soc. Neurosci. 1992, 12, 4793–4799. [Google Scholar] [CrossRef]
- Miwa, T.; Furukawa, S.; Nakajima, K.; Furukawa, Y.; Kohsaka, S. Lipopolysaccharide enhances synthesis of brain-derived neurotrophic factor in cultured rat microglia. J. Neurosci. Res. 1997, 50, 1023–1029. [Google Scholar] [CrossRef]
- Conner, J.M.; Lauterborn, J.C.; Yan, Q.; Gall, C.M.; Varon, S. Distribution of brain-derived neurotrophic factor (bdnf) protein and mrna in the normal adult rat cns: Evidence for anterograde axonal transport. J. Neurosci. Off. J. Soc. Neurosci. 1997, 17, 2295–2313. [Google Scholar] [CrossRef]
- Rossler, O.G.; Giehl, K.M.; Thiel, G. Neuroprotection of immortalized hippocampal neurones by brain-derived neurotrophic factor and raf-1 protein kinase: Role of extracellular signal-regulated protein kinase and phosphatidylinositol 3-kinase. J. Neurochem. 2004, 88, 1240–1252. [Google Scholar] [CrossRef]
- Vilar, M.; Mira, H. Regulation of neurogenesis by neurotrophins during adulthood: Expected and unexpected roles. Front. Neurosci. 2016, 10, 26. [Google Scholar] [CrossRef]
- Almeida, R.D.; Manadas, B.J.; Melo, C.V.; Gomes, J.R.; Mendes, C.S.; Graos, M.M.; Carvalho, R.F.; Carvalho, A.P.; Duarte, C.B. Neuroprotection by bdnf against glutamate-induced apoptotic cell death is mediated by erk and pi3-kinase pathways. Cell Death Differ. 2005, 12, 1329–1343. [Google Scholar] [CrossRef]
- Min, A.Y.; Doo, C.N.; Son, E.J.; Sung, N.Y.; Lee, K.J.; Sok, D.E.; Kim, M.R. N-palmitoyl serotonin alleviates scopolamine-induced memory impairment via regulation of cholinergic and antioxidant systems, and expression of bdnf and p-creb in mice. Chem.-Biol. Interact. 2015, 242, 153–162. [Google Scholar] [CrossRef]
- Habtemariam, S. Natural products in alzheimer’s disease therapy: Would old therapeutic approaches fix the broken promise of modern medicines? Molecules 2019, 24, 1519. [Google Scholar] [CrossRef]
- Schwartz, J.N.; Daniels, C.A.; Shivers, J.C.; Klintworth, G.K. Experimental cytomegalovirus ophthalmitis. Am. J. Pathol. 1974, 77, 477–492. [Google Scholar] [PubMed]
- Jeon, H.L.; Yoo, J.M.; Lee, B.D.; Lee, S.J.; Sohn, E.J.; Kim, M.R. Anti-inflammatory and antioxidant actions of n-arachidonoyl serotonin in raw264.7 cells. Pharmacology 2016, 97, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.M.; Park, K.I.; Ma, J.Y. Anticolitic effect of viscum coloratum through suppression of mast cell activation. Am. J. Chin. Med. 2019, 47, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.C.; Yoo, J.M.; Sok, D.E.; Kim, M.R. Neuroprotective effect of n-acyl 5-hydroxytryptamines on glutamate-induced cytotoxicity in ht-22 cells. Neurochem. Res. 2014, 39, 2440–2451. [Google Scholar] [CrossRef]
- Anderton, M.J.; Manson, M.M.; Verschoyle, R.; Gescher, A.; Steward, W.P.; Williams, M.L.; Mager, D.E. Physiological modeling of formulated and crystalline 3,3′-diindolylmethane pharmacokinetics following oral administration in mice. Drug Metab. Dispos. Biol. Fate Chem. 2004, 32, 632–638. [Google Scholar] [CrossRef]
- Yu, A.R.; Jeong, Y.J.; Hwang, C.Y.; Yoon, K.S.; Choe, W.; Ha, J.; Kim, S.S.; Pak, Y.K.; Yeo, E.J.; Kang, I. Alpha-naphthoflavone induces apoptosis through endoplasmic reticulum stress via c-src-, ros-, mapks-, and arylhydrocarbon receptor-dependent pathways in ht22 hippocampal neuronal cells. Neurotoxicology 2019, 71, 39–51. [Google Scholar] [CrossRef]
- Guo, W.; Ji, Y.; Wang, S.; Sun, Y.; Lu, B. Neuronal activity alters bdnf-trkb signaling kinetics and downstream functions. J. Cell Sci. 2014, 127, 2249–2260. [Google Scholar] [CrossRef]
- Cunha, C.; Brambilla, R.; Thomas, K.L. A simple role for bdnf in learning and memory? Front. Mol. Neurosci. 2010, 3, 1. [Google Scholar] [CrossRef]
- Saw, C.L.; Cintron, M.; Wu, T.Y.; Guo, Y.; Huang, Y.; Jeong, W.S.; Kong, A.N. Pharmacodynamics of dietary phytochemical indoles i3c and dim: Induction of nrf2-mediated phase ii drug metabolizing and antioxidant genes and synergism with isothiocyanates. Biopharm. Drug Dispos. 2011, 32, 289–300. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.D.; Yoo, J.-M.; Baek, S.Y.; Li, F.Y.; Sok, D.-E.; Kim, M.R. 3,3′-Diindolylmethane Promotes BDNF and Antioxidant Enzyme Formation via TrkB/Akt Pathway Activation for Neuroprotection against Oxidative Stress-Induced Apoptosis in Hippocampal Neuronal Cells. Antioxidants 2020, 9, 3. https://doi.org/10.3390/antiox9010003
Lee BD, Yoo J-M, Baek SY, Li FY, Sok D-E, Kim MR. 3,3′-Diindolylmethane Promotes BDNF and Antioxidant Enzyme Formation via TrkB/Akt Pathway Activation for Neuroprotection against Oxidative Stress-Induced Apoptosis in Hippocampal Neuronal Cells. Antioxidants. 2020; 9(1):3. https://doi.org/10.3390/antiox9010003
Chicago/Turabian StyleLee, Bo Dam, Jae-Myung Yoo, Seong Yeon Baek, Fu Yi Li, Dai-Eun Sok, and Mee Ree Kim. 2020. "3,3′-Diindolylmethane Promotes BDNF and Antioxidant Enzyme Formation via TrkB/Akt Pathway Activation for Neuroprotection against Oxidative Stress-Induced Apoptosis in Hippocampal Neuronal Cells" Antioxidants 9, no. 1: 3. https://doi.org/10.3390/antiox9010003
APA StyleLee, B. D., Yoo, J.-M., Baek, S. Y., Li, F. Y., Sok, D.-E., & Kim, M. R. (2020). 3,3′-Diindolylmethane Promotes BDNF and Antioxidant Enzyme Formation via TrkB/Akt Pathway Activation for Neuroprotection against Oxidative Stress-Induced Apoptosis in Hippocampal Neuronal Cells. Antioxidants, 9(1), 3. https://doi.org/10.3390/antiox9010003