Antioxidant Protection from UV- and Light-Stress Related to Carotenoid Structures

Abstract
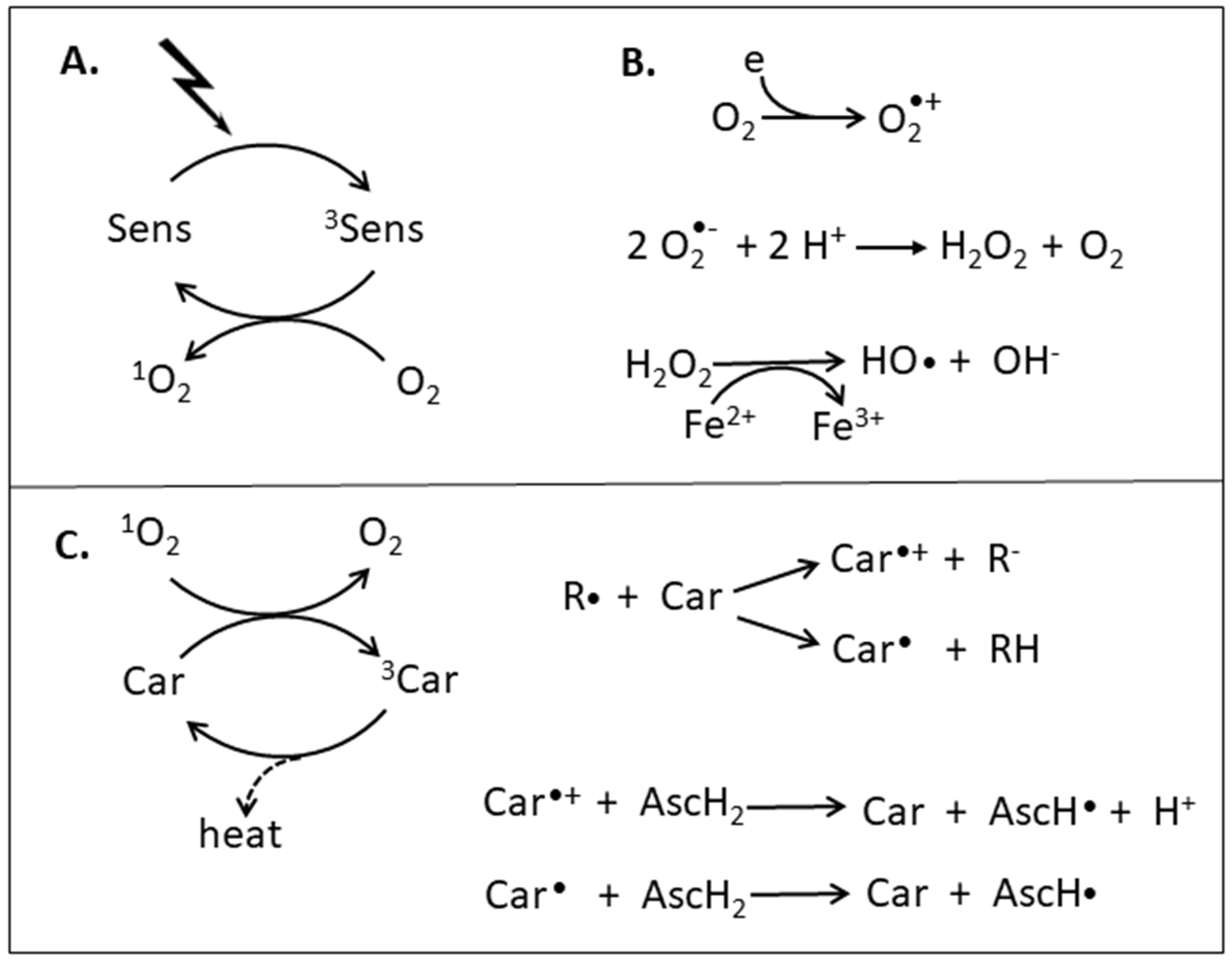
:1. Oxidants and Antioxidants
2. In Vitro Antioxidant Assays for Different Carotenoids
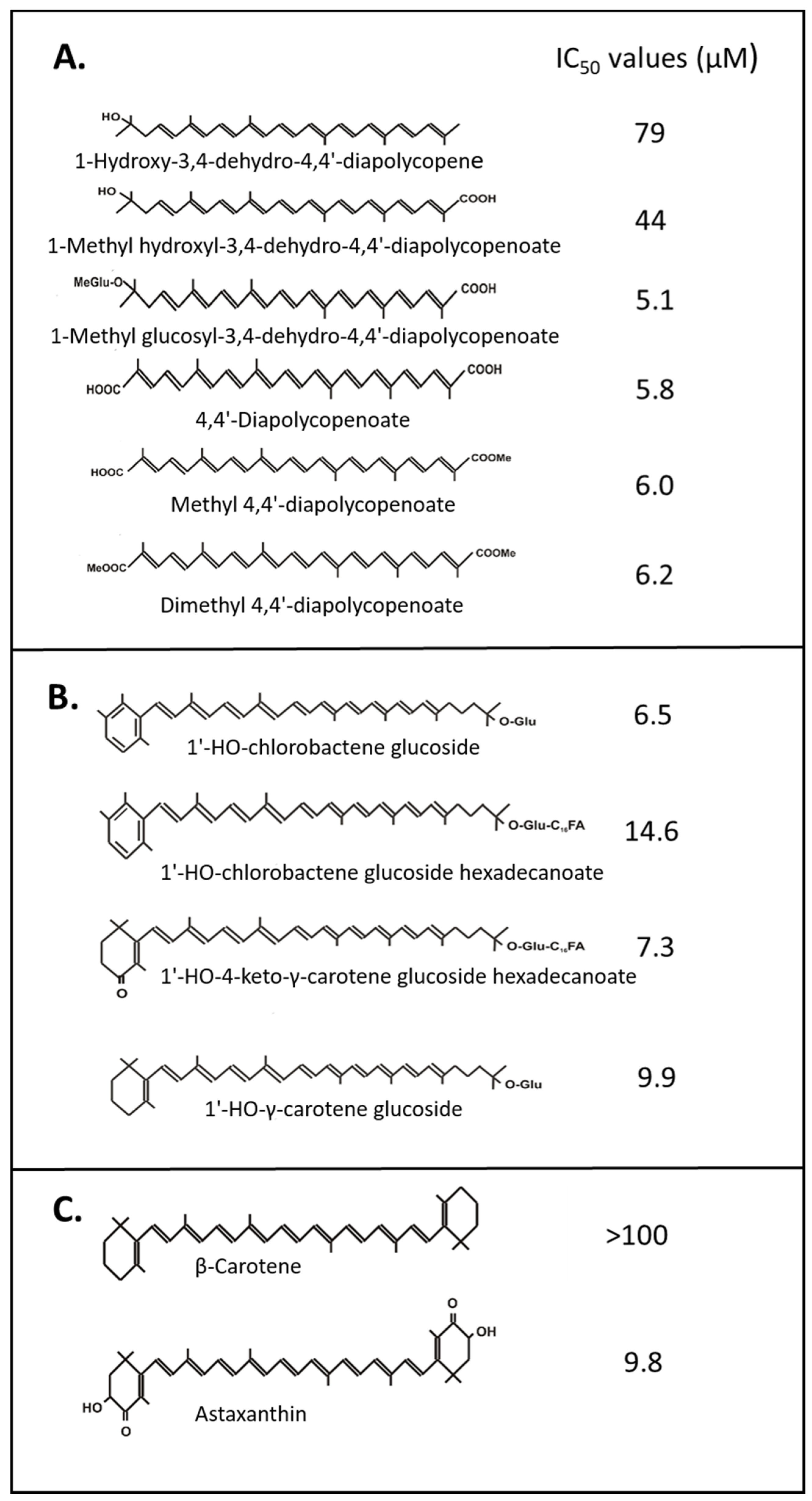
2.1. O2 Quenching by Carotenoids
2.2. Radical Scavenging by Carotenoids
3. Protection in Living Organisms by Carotenoids against High Light and UV Radiation
3.1. Heterotrophic Organisms
3.2. Photosynthetic Organisms
4. Conclusion on Carotenoids as Antioxidants
Funding
Conflicts of Interest
References
- Santos, A.L.; Oliveira, V.; Baptista, I.; Henriques, I.; Gomes, N.C.; Almeida, A.; Correia, A.; Cunha, A. Wavelength dependence of biological damage induced by UV radiation on bacteria. Arch. Microbiol. 2013, 195, 63–74. [Google Scholar] [CrossRef]
- Edge, R.; McGarvey, D.J.; Truscott, T.G. The carotenoids as antioxidants—A review. J. Photochem. Photobiol. B Biol. 1997, 41, 189–200. [Google Scholar] [CrossRef]
- Sies, H. Strategies of antioxidant defense. Eur. J. Biochem. 1993, 215, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Ogilby, P.R. Singlet oxygen: There is still something new under the sun, and it is better than ever. Photochem. Photobiol. Sci. 2010, 9, 1543–1560. [Google Scholar] [CrossRef] [PubMed]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Marklund, S. Spectrophotometric study of spontaneous disproportionation of superoxide anion radical and sensitive direct assay for superoxide dismutase. J. Biol. Chem. 1976, 251, 7504–7507. [Google Scholar]
- Kehrer, J.P. The Haber–Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Mortensen, A.; Skibsted, L.H.; Willnow, A.; Everett, S.A. Kinetics of photobleaching of β-carotene in chloroform and formation of transient carotenoid species absorbing in the near infra-red. Free Rad. Res. 1998, 28, 69–80. [Google Scholar] [CrossRef]
- Liebler, D.C.; McClure, T.D. Antioxidant reactions of β-carotene: Identification of carotenoid-radical adducts. Chem. Res. Toxicol. 1996, 9, 8–11. [Google Scholar] [CrossRef]
- Woodall, A.A.; Lee, S.W.; Weesie, R.J.; Jackson, M.J.; Britton, G. Oxidation of carotenoids by free radicals: Relationship between structure and reactivity. Biochim. Biophys. Acta 1997, 1336, 33–42. [Google Scholar] [CrossRef]
- Burke, M.; Edge, R.; Land, E.J.; McGarvey, D.J.; Truscott, T.G. One-electron reduction potentials of dietary carotenoid radical cations in aqueous micellar environments. FEBS Lett. 2001, 500, 132–136. [Google Scholar] [CrossRef]
- Böhm, F.; Edge, R.; Truscott, T.G. Interactions of dietary carotenoids with activated (singlet) oxygen and free radicals: Potential effects for human health. Mol. Nutr. Food Res. 2012, 56, 205–216. [Google Scholar] [CrossRef] [PubMed]
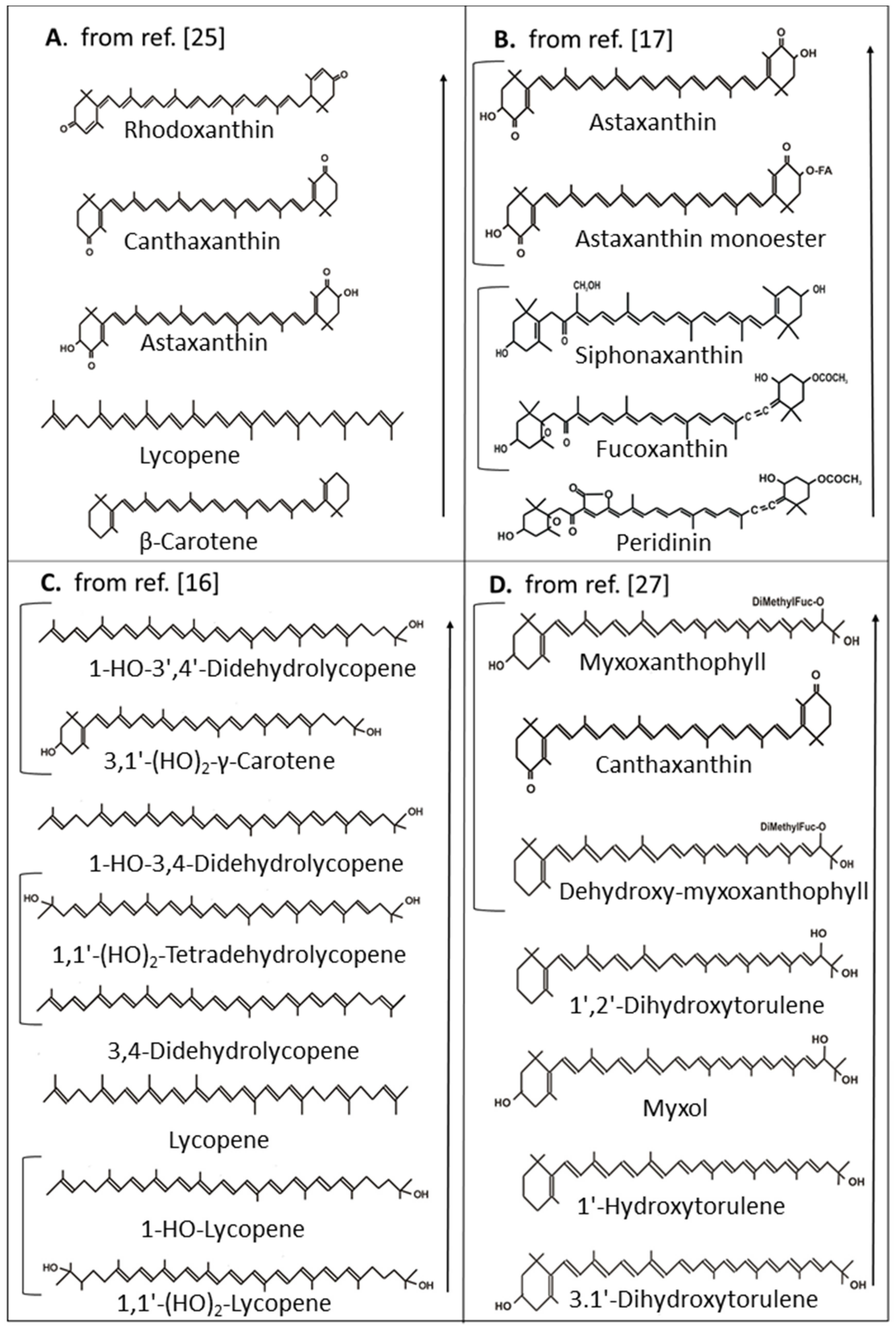
- Krinsky, N.I.; Yeum, K.J. Carotenoid-radical interactions. Biochem. Biophys. Res. Commun. 2003, 305, 754–760. [Google Scholar] [CrossRef]
- Yeum, K.-J.; Aldini, G.; Russel, R.R.; Krinsky, N.I. Antioxidant/pro-oxidant actions of carotenoids. In Carotenoids Nutrition and Health; Birkhäuser Verlag: Basel, Switzerland, 2009; Volume 5, pp. 235–268. [Google Scholar]
- Britton, G.; Liaan-Jensen, S.; Pfander, H. Carotenoid Handbook; Birkhäuser Verlag: Basel, Switzerland, 2004. [Google Scholar]
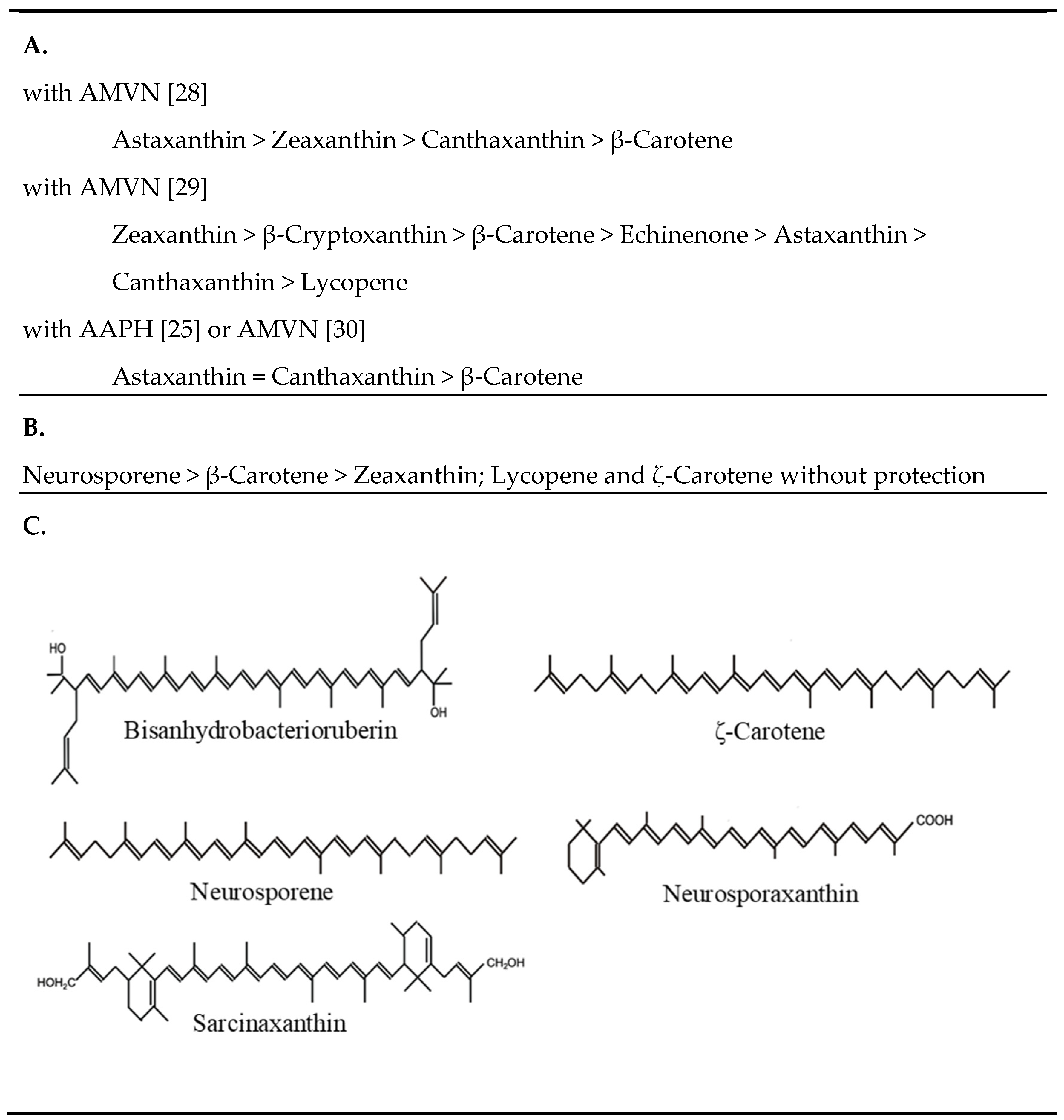
- Albrecht, M.; Takaichi, S.; Steiger, S.; Wang, Z.-Y.; Sandmann, G. Novel hydroxycarotenoids with improved antioxidative properties produced by gene combination in Escherichia coli. Nat. Biotechnol. 2000, 18, 843–846. [Google Scholar] [CrossRef] [PubMed]
- Dambeck, M.; Sandmann, G. Antioxidative activities of algal keto carotenoids acting as antioxidative protectants in the chloroplast. Photochem. Photobiol. 2014, 90, 814–819. [Google Scholar] [CrossRef]
- Hirayama, O.; Nakamura, K.; Hamada, S.; Kobayashi, Y. Singlet oxygen quenching ability of naturally occurring carotenoids. Lipids 1994, 29, 149–150. [Google Scholar] [CrossRef]
- Kobayashi, M.; Sakamoto, Y. Singlet oxygen quenching ability of astaxanthin esters from the green alga Haematococcus pluvialis. Biotechnol. Lett. 1999, 21, 265–269. [Google Scholar] [CrossRef]
- Osawa, A.; Iki, K.; Sandmann, G.; Shindo, K. Isolation and identification of 4,4′-diapolycopene-4,4′-dioic acid produced by Bacillus firmus GB1 and its singlet oxygen quenching activity. J. Oleo Sci. 2013, 62, 955–960. [Google Scholar] [CrossRef]
- Osawa, A.; Ishii, Y.; Sasamura, N.; Morita, M.; Köcher, S.; Müller, V.; Sandmann, G.; Shindo, K. Hydroxy-3,4-dehydro-apo-8′-lycopene and methyl hydroxy-3,4-dehydro-apo-8′-lycopenoate, novel C (30) carotenoids produced by a mutant of marine bacterium Halobacillus halophilus. J. Antibiot. 2010, 63, 291–295, reprinted in J. Antibiot. 2014, 67, 733–735. [Google Scholar] [CrossRef]
- Shindo, K.; Endo, M.; Miyake, Y.; Wakasugi, K.; Morritt, D.; Bramley, P.M.; Fraser, P.D.; Kasai, H.; Misawa, N. Methyl glucosyl-3,4-dehydro-apo-8′-lycopenoate, a novel antioxidative glyco-C (30)-carotenoic acid produced by a marine bacterium Planococcus maritimus. J. Antibiot. 2008, 61, 729–735, reprinted in J. Antibiot. 2014, 67, 731–733. [Google Scholar] [CrossRef]
- Shindo, K.; Asagi, E.; Sano, A.; Hotta, E.; Minemura, N.; Mikami, K.; Tamesada, E.; Misawa, N.; Maoka, T. Diapolycopenedioic acid xylosyl esters A, B, and C, novel antioxidative glyco-C30-carotenoic acids produced by a new marine bacterium Rubritalea squalenifaciens. J. Antibiot. 2008, 61, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Osawa, A.; Kasahara, A.; Mastuoka, S.; Gassel, S.; Sandmann, G.; Shindo, K. Isolation of a novel carotenoid, OH-chlorobactene glucoside hexadecanoate, and related rare carotenoids from Rhodococcus sp. CIP and their antioxidative activities. Biosci. Biotechnol. Biochem. 2011, 75, 2142–2147. [Google Scholar] [CrossRef] [PubMed]
- Beutner, S.; Bloedorn, B.; Frixel, S.; Hernandez Blanco, I.; Hoffmann, T.; Martin, H.-D.; Mayer, B.; Noack, P.; Ruck, C.; Schmidt, M.; et al. assessment of antioxidant properties of natural colorants and phytochemicals: Carotenoids, flavonoids, phenols and indigoids. The role of β-carotene in antioxidant functions. J. Sci. Food Agric. 2001, 81, 559–568. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Gildenhoff, N. Isolation von Myxoxanthophyll und Seinen Derivaten und ihre Bedeutung beim Schutz vor Photooxidation. Master’s Thesis, Goethe University, Frankfurt, Germany, 2005. (Unpublished results). [Google Scholar]
- Lim, B.P.; Nagao, A.; Terao, J.; Tanaka, K.; Suzuki, T.; Takama, K. Antioxidant activity of xanthophylls on peroxyl radical-mediated phospholipid peroxidation. Biochim. Biophys. Acta 1992, 1126, 178–184. [Google Scholar] [PubMed]
- Woodall, A.A.; Britton, G.; Jackson, M.J. Carotenoids and protection of phospholipids in solution or in liposomes against oxidation by peroxyl radicals: Relationship between carotenoid structure and protective ability. Biochim. Biophys. Acta 1997, 1336, 575–586. [Google Scholar] [CrossRef]
- Palozza, P.; Krinsky, N.I. Astaxanthin and canthaxanthin are potent antioxidants in a membrane model. Arch. Biochem. Biophys. 1992, 297, 291–295. [Google Scholar] [CrossRef]
- Zalazar, L.; Pagola, P.; Miro, M.V.; Churio, M.S.; Cerletti, M.; Martınez, C.; Iniesta-Cuerda, M.; Soler, A.J.; Cesari, A.; De Castro, R. Bacterioruberin extracts from a genetically modified hyperpigmented Haloferax volcanii strain: Antioxidant activity and bioactive properties on sperm cells. J. Appl. Microbiol. 2019, 126, 796–810. [Google Scholar] [CrossRef]
- Krinsky, N.I. Non-photosynthetic functions of carotenoids. Phil. Trans. R. Soc. Lond. B. 1978, 248, 581–590. [Google Scholar] [CrossRef]
- Palozza, P.; Krinsky, N.I. Antioxidant effect of carotenoids in vivo and in vitro: An overview. Methods Enzymol. 1992, 213, 403–420. [Google Scholar]
- Liaaen-Jensen, S.; Weeks, O.B.; Strang, R.H.; Thirkell, D. Identity of the C-50-carotenoid dehydrogenans-P439 and sarcinaxanthin. Nature 1967, 214, 379–380. [Google Scholar] [CrossRef] [PubMed]
- Mathews-Roth, M.M.; Krinsky, N.I. Sudies on the protective function of the carotenoid pigments of Sarcina lutea. Photochem. Photobiol. 1970, 11, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Mathews, M.M.; Sistrom, W.R. The function of carotenoid pigments of Sarcina lutea. Arch. Microbiol. 1960, 35, 139–146. [Google Scholar] [CrossRef]
- Norgard, S.; Aasen, A.J.; Liaaen-Jensen, S. Bacterial carotenoids. XXXII. C50-Carotenoids. 6. Carotenoids from Corynebacterium poinsettiae including for new C50-diols. Acta Chem. Scand. 1970, 24, 2183–2197. [Google Scholar] [CrossRef] [PubMed]
- Kunisawa, R.; Stanier, R.Y. Studies on the role of carotenoid pigments in a chemoheterotrophic bacterium Corynebacterium poinsettiae. Arch. Microbiol. 1958, 31, 146–156. [Google Scholar] [CrossRef]
- Steiger, S.; Perez-Fons, L.; Fraser, P.D.; Sandmann, G. Biosynthesis of a novel C30 carotenoid in Bacillus firmus isolates. J. Appl. Microbiol. 2011, 13, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Garber, E.D.; Baird, M.L.; Chapman, D.J. Genetics of Ustilago violacea I: Carotenoid mutants and carotenogenesis. Botanical Gazette 1975, 136, 341–346. [Google Scholar] [CrossRef]
- Will III, O.H.; Jankowski, P.; Kowacs, A.; Rossing, W.; Schneider, P.; Newland, N.A. A comparison of photo-killing among carotene and cytochrome c accumulating strains of the smut fungus Ustilaga violacea at specific wavelengths from 400 to 600 nm. Photochem. Photobiol. 1987, 45, 609–615. [Google Scholar] [CrossRef]
- Rau, W. Mechanism of photoregulation of carotenoid biosynthesis in plants. Pure Appl. Chem. 1985, 57, 777–784. [Google Scholar] [CrossRef]
- Blanc, P.L.; Tuveson, R.W.; Sargent, M.L. Inactivation of carotenoid-producing and albino strains of Neurospora crassa by visible light, blacklight, and ultraviolet radiation. J. Bacteriol. 1976, 125, 616–625. [Google Scholar] [PubMed]
- Tuveson, R.W.; Larson, R.A.; Kagan, J. Role of cloned carotenoid genes expressed in Escherichia coli in protecting against inactivation by near-UV light and specific phototoxic molecules. J. Bacteriol. 1988, 170, 4675–4680. [Google Scholar] [CrossRef] [PubMed]
- Kagan, J.; Bazin, M.; Santus, R. Photosensitization with alpha-terthienyl: The formation of superoxide ion in aqueous media. J. Photochem. Photobiol. B. 1989, 3, 165–174. [Google Scholar] [CrossRef]
- Sandmann, G.; Kuhn, S.; Böger, P. Evaluation of structurally different carotenoids in Escherichia coli transformants as protectants against UV-B radiation. Appl. Environ. Microbiol. 1998, 64, 1972–1974. [Google Scholar] [PubMed]
- Bohn, T. Carotenoids and markers of oxidative stress in human observational studies and intervention trials: Implications for chronic diseases. Antioxidants 2019, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Mathews, M.M. Protective effect of β-carotene against lethal photosensitization by hæmatoporphyrin. Nature 1964, 203, 1092. [Google Scholar] [CrossRef] [PubMed]
- Mathews, M.M. Carotenoids in erythropoietic protoporphyria and other photosensitivity diseases. Ann. N. Y. Acad. Sci. 1993, 691, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Lotan, S.B.; Vitt, K.; Scholz, P.; Keck, C.M.; Meinke, M.C. ROS production and glutathione response in keratinocytes after application of β-carotene and VIS/NIR irradiation. Chem. Biol. Interact. 2018, 280, 1–7. [Google Scholar]
- Russel, R.M. The enigma of β-carotene in carcinogenesis: What can be learned from animal studies? J. Nutr. 2004, 134, 262S–268S. [Google Scholar] [CrossRef] [PubMed]
- Telfer, A.; Pascal, A.; Gall, A. Carotenoids in photosynthesis. In Carotenoids Natural Functions; Birkhäuser Verlag: Basel, Switzerland, 2008; Volume 4, pp. 266–306. [Google Scholar]
- Glaser, A.N. Light harvesting by phycobilisomes. Ann. Rev. Biophys. Biophys. Chem. 1985, 14, 47–77. [Google Scholar] [CrossRef]
- Schäfer, L.; Vioque, A.; Sandmann, G. Functional in situ evaluation of photosynthesis-protecting carotenoids in mutants of the cyanobacterium Synechocystis PCC6803. J. Photochem. Photobiol. B Biol. 2005, 78, 195–201. [Google Scholar] [CrossRef]
- Götz, T.; Windhövel, U.; Böger, P.; Sandmann, G. Protection of Photosynthesis against Ultraviolet-B Radiation by Carotenoids in Transformants of the CyanobacteriumSynechococcus PCC7942. Plant Physiol. 1999, 120, 599–604. [Google Scholar]
- Albrecht, M.; Steiger, S.; Sandmann, G. Expression of a ketolase gene mediates the synthesis of canthaxanthin in Synechococcus leading to tolerance against photoinhibition, pigment degradation and UV-B sensitivity of photosynthesis. Photochem. Photobiol. 2001, 73, 551–555. [Google Scholar] [CrossRef]
- Schöpf, L.; Mautz, J.; Sandmann, G. Multiple ketolases involved in light regulation of canthaxanthin biosynthesis in Nostoc punctiforme PCC 73102. Planta 2013, 237, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Steiger, S.; Schäfer, L.; Sandmann, G. High-light upregulation of carotenoids and their antioxidative properties in the cyanobacterium Synechocystis PCC 6803. J. Photochem. Photobiol. B Biol. 1999, 521, 14–18. [Google Scholar] [CrossRef]
- Götz, T.; Sandmann, G.; Römer, S. Expression of a bacterial carotene hydroxylase gene (crtZ) enhances UV tolerance in tobacco. Plant Mol. Biol. 2002, 50, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M. Carotenoids as membrane stabilizers in chloroplasts. Trend Plant Sci. 1998, 3, 147–151. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transformants | Carotenoid (mg/g dw) | Photosynthesis (% of Untreated Control) | |
|---|---|---|---|
| High Light * | UV Treatment ** | ||
| non-transgenic | Zeax 1.0 | 66 | 32 |
| CrtB | Zeax 1.1 | -- | 37 |
| Psy | Zeax 1.7 | -- | 58 |
| CrtZ | Zeax 1.9 | 82 | 60 |
| Bkt *** | Canth 1.6+ Zeax 0.7 | 95 | -- |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandmann, G. Antioxidant Protection from UV- and Light-Stress Related to Carotenoid Structures. Antioxidants 2019, 8, 219. https://doi.org/10.3390/antiox8070219
Sandmann G. Antioxidant Protection from UV- and Light-Stress Related to Carotenoid Structures. Antioxidants. 2019; 8(7):219. https://doi.org/10.3390/antiox8070219
Chicago/Turabian StyleSandmann, Gerhard. 2019. "Antioxidant Protection from UV- and Light-Stress Related to Carotenoid Structures" Antioxidants 8, no. 7: 219. https://doi.org/10.3390/antiox8070219
APA StyleSandmann, G. (2019). Antioxidant Protection from UV- and Light-Stress Related to Carotenoid Structures. Antioxidants, 8(7), 219. https://doi.org/10.3390/antiox8070219