Comparison of the Phenolic Profiles of Soaked and Germinated Peanut Cultivars via UPLC-QTOF-MS

 ,
,  , and
, and 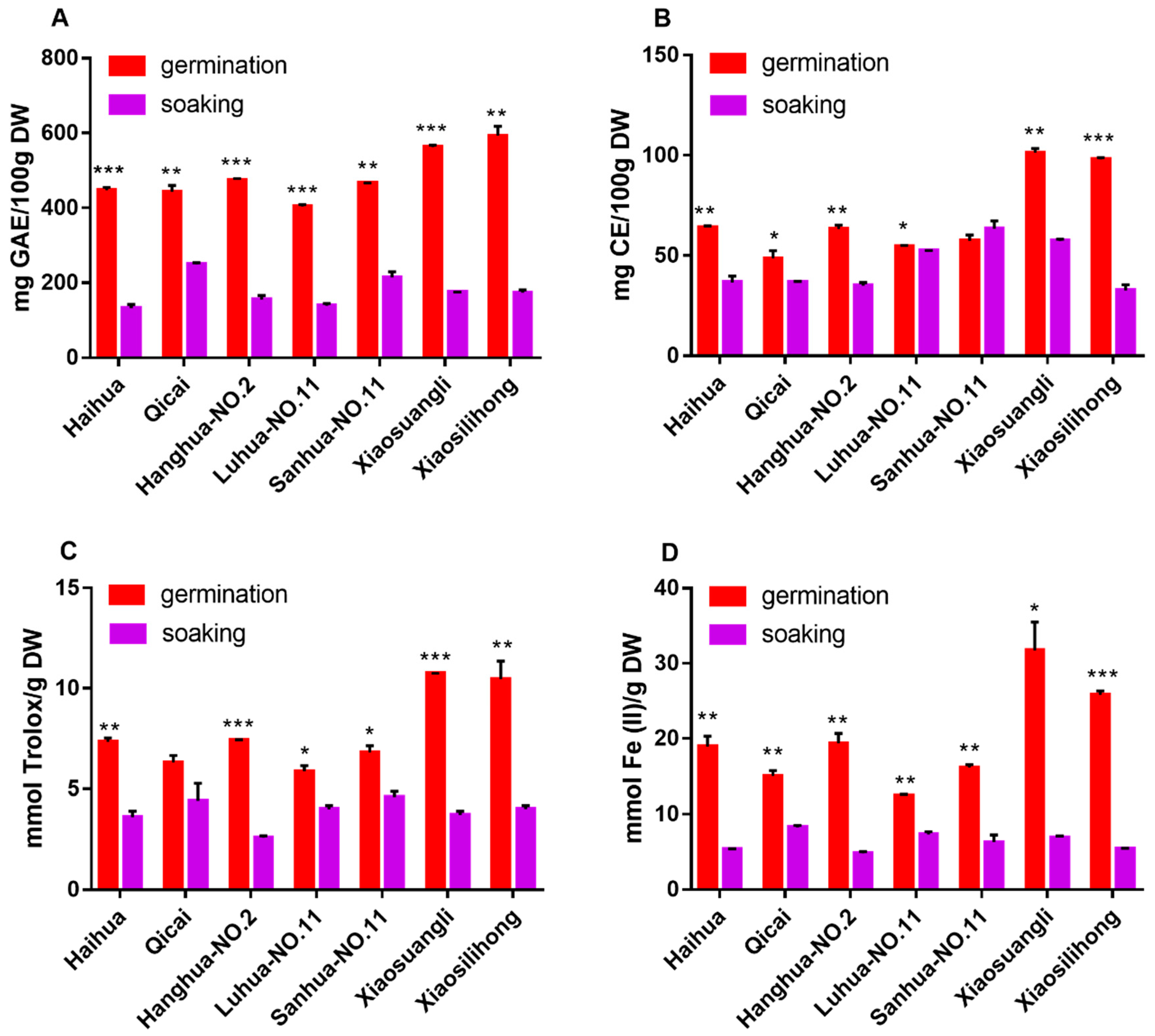{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Soaked and Germinated Peanuts
2.2. Extraction Procedure
2.3. Determination of Total Phenolic and Flavonoid Contents
2.4. Determination of Free Radical Scavenging Activity by 1,1-Diphenyl-2-Picrylhydrazyl (DPPH) Method
2.5. Ferric Ion Reducing Antioxidant Power (FRAP) Assay
2.6. UPLC-QTOF-MS Analysis
2.7. Statistical Analysis
3. Results and Discussion
3.1. Changes of TPC and TFC Induced by Germination
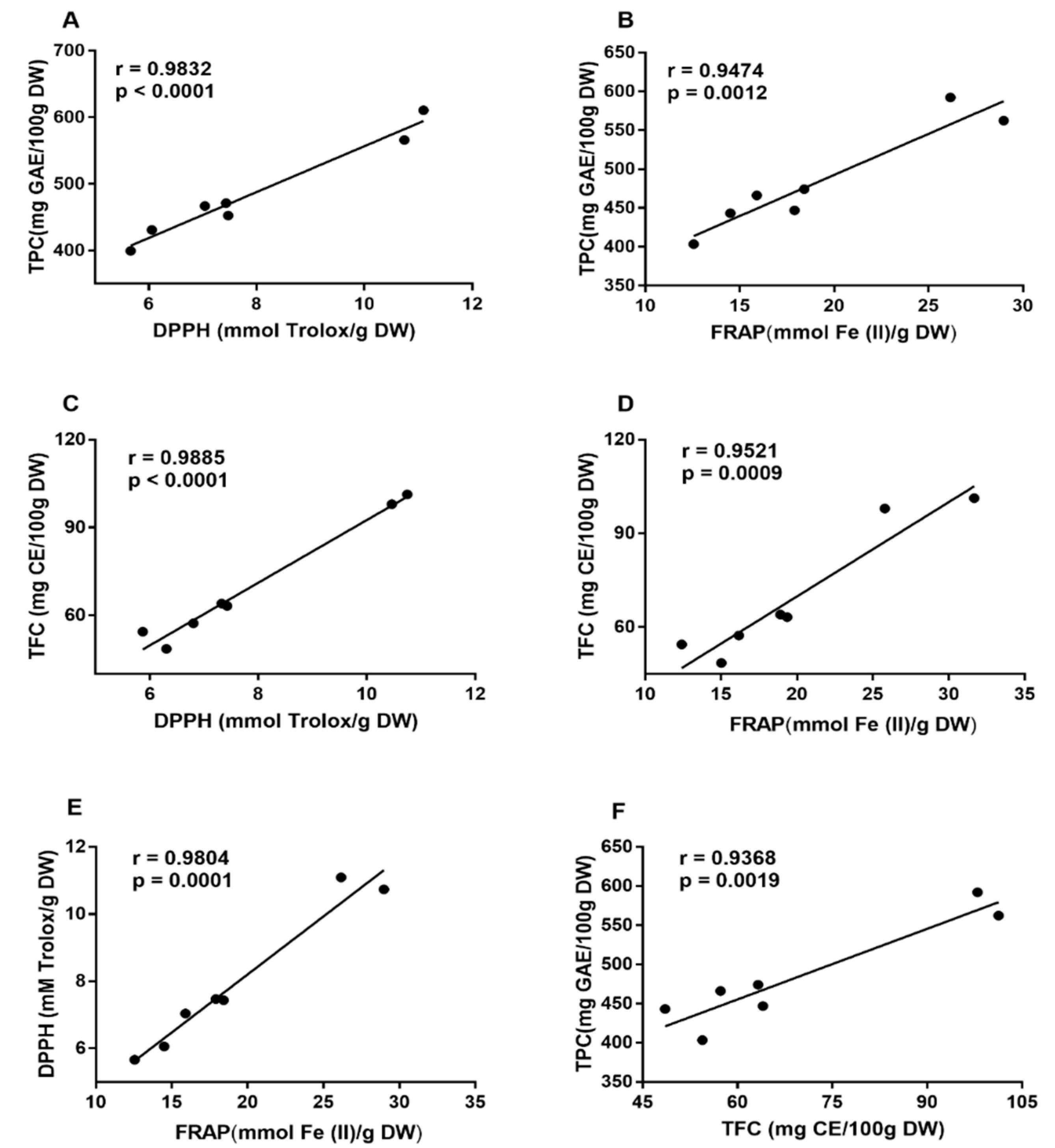
3.2. Changes of Antioxidant Activities Induced by Germination
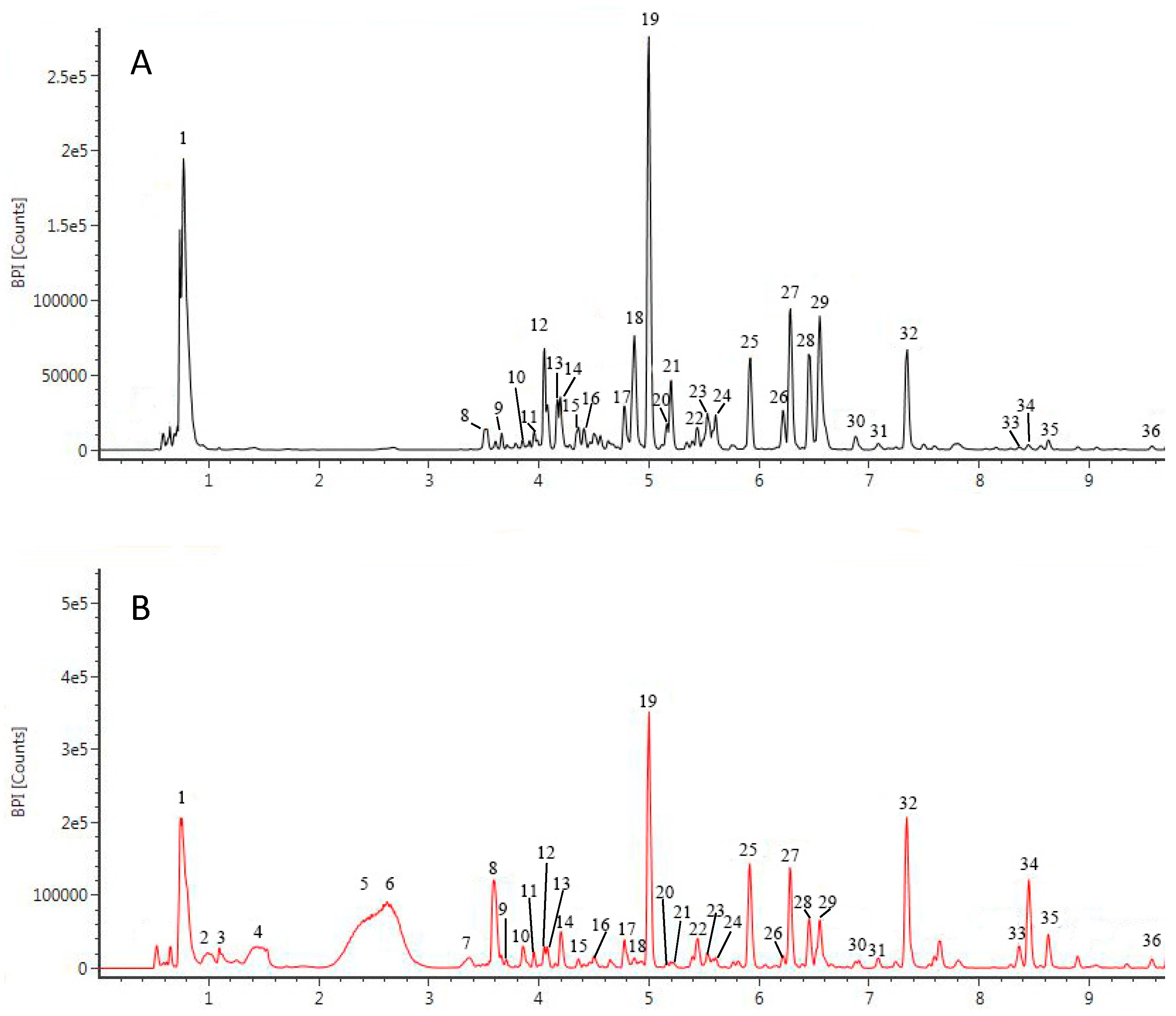
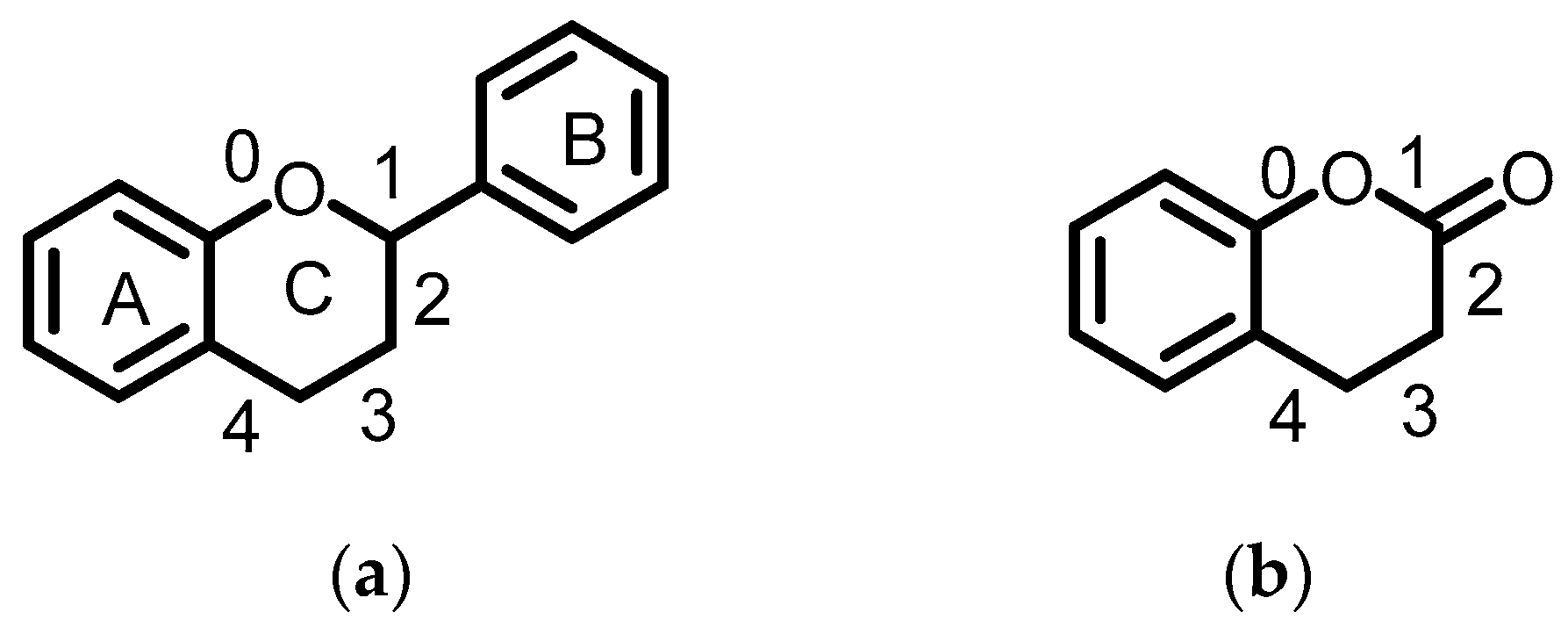
3.3. Characterization of the Phenolic Profile of Peanut Extracts by UPLC-QTOF/MS
3.3.1. Flavanols
3.3.2. Flavanones
3.3.3. Flavones
3.3.4. Flavonols
3.3.5. Isoflavonoids
3.3.6. Anthocyanins
3.3.7. Coumarins
3.3.8. Others
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gan, R.Y.; Lui, W.Y.; Wu, K.; Chan, C.L.; Dai, S.H.; Sui, Z.Q.; Corke, H. Bioactive compounds and bioactivities of germinated edible seeds and sprouts: An updated review. Trends Food Sci. Technol. 2017, 59, 1–14. [Google Scholar] [CrossRef]
- Luo, Y.W.; Xie, W.H. Effect of germination conditions on phytic acid and polyphenols of faba bean sprouts (Vicia faba L.). Legume Res. 2013, 36, 489–495. [Google Scholar]
- Aguilera, Y.; Liébana, R.; Herrera, T.; Rebollo-Hernanz, M.; Sanchez-Puelles, C.; Benitez, V.; Martín-Cabrejas, M.A. Effect of illumination on the content of melatonin, phenolic compounds, and antioxidant activity during germination of lentils (Lens culinaris L.) and kidney beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2014, 62, 10736–10743. [Google Scholar] [CrossRef]
- Tajoddin, M.; Manohar, S.; Lalitha, J. Effect of soaking and germination on polyphenol content and polyphenol oxidase activity of mung bean (Phaseolus aureus L.) cultivars differing in seed color. Int. J. Food Prop. 2014, 17, 782–790. [Google Scholar] [CrossRef]
- Lazo-Vélez, M.A.; Guardado-Félix, D.; Avilés-González, J.; Romo-López, I.; Serna-Saldívar, S.O. Effect of germination with sodium selenite on the isoflavones and cellular antioxidant activity of soybean (Glycine max). LWT Food Sci. Technol. 2018, 93, 64–70. [Google Scholar] [CrossRef]
- Wang, H.; Wang, J.H.; Guo, X.B.; Brennan, C.S.; Li, T.; Fu, X.; Chen, G.; Liu, R.H. Effect of germination on lignin biosynthesis, and antioxidant and antiproliferative activities in flaxseed (Linum usitatissimum L.). Food Chem. 2016, 205, 170–177. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Peanut (Arachis hypogaea L.): A prospective legume crop to offer multiple health benefits under changing climate. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1325–1338. [Google Scholar] [CrossRef]
- Adhikari, B.; Dhungana, S.K.; Ali, M.W.; Adhikari, A.; Kim, I.D.; Shin, D.H. Resveratrol, total phenolic and flavonoid contents, and antioxidant potential of seeds and sprouts of Korean peanuts. Food Sci. Biotechnol. 2018, 27, 1275–1284. [Google Scholar] [CrossRef]
- Bansode, R.R.; Randolph, P.; Ahmedna, M.; Williams, L.L.; Yu, J. Bioavailability and hypolipidemic effects of peanut skin polyphenols. J. Med. Food 2015, 18, 265–272. [Google Scholar] [CrossRef]
- de Camargo, A.C.; Regitano-d’Arce, M.A.B.; Rasera, G.B.; Canniatti-Brazaca, S.G.; do Prado-Silva, L.; Alvarenga, V.O.; Sant’Ana, A.S.; Shahidi, F. Phenolic acids and flavonoids of peanut by-products: Antioxidant capacity and antimicrobial effects. Food Chem. 2017, 237, 538–544. [Google Scholar] [CrossRef]
- Tsujita, T.; Shintani, T.; Sato, H. Preparation and characterization of peanut seed skin polyphenols. Food Chem. 2014, 151, 15–20. [Google Scholar] [CrossRef]
- Larrauri, M.; Zunino, M.P.; Zygadlo, J.A.; Grosso, N.R.; Nepote, V. Chemical characterization and antioxidant properties of fractions separated from extract of peanut skin derived from different industrial processes. Ind. Crops Prod. 2016, 94, 964–971. [Google Scholar] [CrossRef]
- Gan, R.Y.; Wang, M.F.; Lui, W.Y.; Wu, K.; Corke, H. Dynamic changes in phytochemical composition and antioxidant capacity in green and black mung bean (Vigna radiata) sprouts. Int. J. Food Sci. Technol. 2016, 51, 2090–2098. [Google Scholar] [CrossRef]
- Miron-Merida, V.A.; Yanez-Fernandez, J.; Montanez-Barragan, B.; Huerta, B.E.B. Valorization of coffee parchment waste (Coffea arabica) as a source of caffeine and phenolic compounds in antifungal gellan gum films. LWT Food Sci. Technol. 2019, 101, 167–174. [Google Scholar] [CrossRef]
- Gan, R.Y.; Xu, X.R.; Song, F.L.; Kuang, L.; Li, H.B. Antioxidant activity and total phenolic content of medicinal plants associated with prevention and treatment of cardiovascular and cerebrovascular diseases. J. Med. Plants Res. 2010, 4, 2438–2444. [Google Scholar] [CrossRef]
- Zhu, F.; Cai, Y.Z.; Yang, X.S.; Ke, J.X.; Corke, H. Anthocyanins, hydroxycinnamic acid derivatives, and antioxidant activity in roots of different Chinese purple-fleshed sweet potato genotypes. J. Agric. Food Chem. 2010, 58, 7588–7596. [Google Scholar] [CrossRef]
- Chuenchom, P.; Swatsitang, P.; Senawong, T.; Jogloy, S. Antioxidant capacity and phenolic content evaluation on peanut skins from 3 peanut types. Chiang Mai J. Sci. 2016, 43, 1177–1191. [Google Scholar]
- Limmongkon, A.; Nopprang, P.; Chaikeandee, P.; Somboon, T.; Wongshaya, P.; Pilaisangsuree, V. LC-MS/MS profiles and interrelationships between the anti-inflammatory activity, total phenolic content and antioxidant potential of Kalasin 2 cultivar peanut sprout crude extract. Food Chem. 2018, 239, 569–578. [Google Scholar] [CrossRef]
- Yu, L.L.; Haley, S.; Perret, J.; Harris, M.; Wilson, J.; Qian, M. Free radical scavenging properties of wheat extracts. J. Agric. Food Chem. 2002, 50, 1619–1624. [Google Scholar] [CrossRef]
- Fu, L.; Xu, B.T.; Xu, X.R.; Gan, R.Y.; Zhang, Y.; Xia, E.Q.; Li, H.B. Antioxidant capacities and total phenolic contents of 62 fruits. Food Chem. 2011, 129, 345–350. [Google Scholar] [CrossRef]
- Deng, G.F.; Lin, X.; Xu, X.R.; Gao, L.L.; Xie, J.F.; Li, H.B. Antioxidant capacities and total phenolic contents of 56 vegetables. J. Funct. Foods 2013, 5, 260–266. [Google Scholar] [CrossRef]
- Li, A.N.; Li, S.; Li, H.B.; Xu, D.P.; Xu, X.R.; Chen, F. Total phenolic contents and antioxidant capacities of 51 edible and wild flowers. J. Funct. Foods 2014, 6, 319–330. [Google Scholar] [CrossRef]
- Gunaratne, A.; Wu, K.; Li, D.Q.; Bentota, A.; Corke, H.; Cai, Y.Z. Antioxidant activity and nutritional quality of traditional red-grained rice varieties containing proanthocyanidins. Food Chem. 2013, 138, 1153–1161. [Google Scholar] [CrossRef]
- Shan, B.; Cai, Y.Z.; Sun, M.; Corke, H. Antioxidant capacity of 26 spice extracts and characterization of their phenolic constituents. J. Agric. Food Chem. 2005, 53, 7749–7759. [Google Scholar] [CrossRef]
- Gan, R.Y.; Kuang, L.; Xu, X.R.; Zhang, Y.; Xia, E.Q.; Song, F.L.; Li, H.B. Screening of natural antioxidants from traditional Chinese medicinal plants associated with treatment of rheumatic disease. Molecules 2010, 15, 5988–5997. [Google Scholar] [CrossRef]
- Ma, Y.Y.; Kosinska-Cagnazzo, A.; Kerr, W.L.; Amarowicz, R.; Swanson, R.B.; Pegg, R.B. Separation and characterization of soluble esterified and glycoside-bound phenolic compounds in dry-blanched peanut skins by liquid chromatography-electrospray ionization mass spectrometry. J. Agric. Food Chem. 2014, 62, 11488–11504. [Google Scholar] [CrossRef]
- Qiu, J.Y.; Chen, L.L.; Zhu, Q.J.; Wang, D.J.; Wang, W.L.; Sun, X.; Liu, X.Y.; Du, F.L. Screening natural antioxidants in peanut shell using DPPH-HPLC-DAD-TOF/MS methods. Food Chem. 2012, 135, 2366–2371. [Google Scholar] [CrossRef]
- Lopes, R.M.; Agostini-Costa, T.D.; Gimenes, M.A.; Silveira, D. Chemical composition and biological activities of arachis species. J. Agric. Food Chem. 2011, 59, 4321–4330. [Google Scholar] [CrossRef]
- Lee, J.H.; Baek, I.Y.; Ha, T.J.; Choung, M.G.; Ko, J.M.; Oh, S.K.; Kim, H.T.; Ryu, H.W.; Park, K.Y.; Park, K.H. Identification and characterization of phytochemicals from peanut (Arachis hypogaea L.) pods. Food Sci. Biotechnol. 2008, 17, 475–482. [Google Scholar]
- Tsamo, A.T.; Ndibewu, P.P.; Dakora, F.D. Phytochemical profile of seeds from 21 Bambara groundnut landraces via UPLC-qTOF-MS. Food Res. Int. 2018, 112, 160–168. [Google Scholar] [CrossRef]
- Frost, S.; Lerno, L.A.; Zweigenbaum, J.; Heymann, H.; Ebeler, S.E. Characterization of red wine proanthocyanidins using a putative proanthocyanidin database, amide hydrophilic interaction liquid chromatography (HILIC), and time-of-flight mass spectrometry. Molecules 2018, 23. [Google Scholar] [CrossRef]
- He, Y.J.; Cheng, P.; Wang, W.; Yan, S.; Tang, Q.; Liu, D.B.; Xie, H.Q. Rapid investigation and screening of bioactive components in simo decoction via LC-Q-TOF-MS and UF-HPLC-MD methods. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Shen, C.Y.; Jiang, J.G.; Huang, C.L.; Zhu, W.; Zheng, C.Y. Polyphenols from blossoms of Citrus aurantium L. var. amara Engl. show significant anti-complement and anti-inflammatory effects. J. Agric. Food Chem. 2017, 65, 9061–9068. [Google Scholar] [CrossRef]
- Shi, S.Y.; Zhao, Y.; Zhou, H.G.; Zhang, Y.P.; Jiang, X.Y.; Huang, K.L. Identification of antioxidants from Taraxacum mongolicum by high-performance liquid chromatography-diode array detection-radical-scavenging detection-electrospray ionization mass spectrometry and nuclear magnetic resonance experiments. J. Chromatogr. A 2008, 1209, 145–152. [Google Scholar] [CrossRef]
- Kim, B.; Woo, S.; Kim, M.J.; Kwon, S.W.; Lee, J.; Sung, S.H.; Koh, H.J. Identification and quantification of flavonoids in yellow grain mutant of rice (Oryza sativa L.). Food Chem. 2018, 241, 154–162. [Google Scholar] [CrossRef]
- Garcia-Cayuela, T.; Gomez-Maqueo, A.; Guajardo-Flores, D.; Welti-Chanes, J.; Cano, M.P. Characterization and quantification of individual betalain and phenolic compounds in Mexican and Spanish prickly pear (Opuntia ficus-indica L. Mill) tissues: A comparative study. J. Food Comp. Anal. 2019, 76, 1–13. [Google Scholar] [CrossRef]
- Yao, Y.F.; Lin, C.Z.; Liu, F.L.; Zhang, R.J.; Zhang, Q.Y.; Huang, T.; Zou, Y.S.; Wang, M.Q.; Zhu, C.C. Identification and pharmacokinetic studies on complanatuside and its major metabolites in rats by UHPLC-Q-TOF-MS/MS and LC-MS/MS. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Bicha, S.; Benmekhebi, L.; Boubekri, N.; Khellaf, R.; Brouard, I.; Zama, D.; Benayache, S.; Benayache, F. Compositional study, antibacterial and antioxidant potential of Lepidium draba L. (Brascicaceae). Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 283–287. [Google Scholar]
- Sun, M.J.; Yin, Y.W.; Wei, J.; Chen, X.P.; Ouyang, H.Z.; Chang, Y.X.; Gao, X.M.; He, J. Development and validation of a HPLC-MS/MS method for simultaneous determination of twelve bioactive compounds in epimedium: Application to a pharmacokinetic study in rats. Molecules 2018, 23. [Google Scholar] [CrossRef]
- Zheng, Y.; Xu, X.L.; Yuan, F.; Yao, M.Y.; Ji, S.L.; Huang, Z.Q.; Zhang, F. Simultaneous analysis of simple coumarins and furocoumarines in cigarettes by solid-phase extraction with gas chromatography-mass spectrometry. J. AOAC Int. 2017, 100, 1559–1564. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Sun, W.B.; Yang, Z.; Liang, Y.; Zhou, W.; Tang, L. Hemostatic chemical constituents from natural medicine Toddalia asiatica root bark by LC-ESI Q-TOF MS. Chem. Cent. J. 2017, 11, 55. [Google Scholar] [CrossRef]
- Rojas-Garbanzo, C.; Winter, J.; Montero, M.L.; Zimmermann, B.F.; Schieber, A. Characterization of phytochemicals in Costa Rican guava (Psidium friedrichsthalianum-Nied.) fruit and stability of main compounds during juice processing–(U)HPLC-DAD-ESI-TQD-MSn. J. Food Comp. Anal. 2019, 75, 26–42. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.-Q.; Cheng, L.; Long, Z.-Y.; Li, H.-B.; Gunaratne, A.; Gan, R.-Y.; Corke, H. Comparison of the Phenolic Profiles of Soaked and Germinated Peanut Cultivars via UPLC-QTOF-MS. Antioxidants 2019, 8, 47. https://doi.org/10.3390/antiox8020047
Yang Q-Q, Cheng L, Long Z-Y, Li H-B, Gunaratne A, Gan R-Y, Corke H. Comparison of the Phenolic Profiles of Soaked and Germinated Peanut Cultivars via UPLC-QTOF-MS. Antioxidants. 2019; 8(2):47. https://doi.org/10.3390/antiox8020047
Chicago/Turabian StyleYang, Qiong-Qiong, Lin Cheng, Zhi-Yuan Long, Hua-Bin Li, Anil Gunaratne, Ren-You Gan, and Harold Corke. 2019. "Comparison of the Phenolic Profiles of Soaked and Germinated Peanut Cultivars via UPLC-QTOF-MS" Antioxidants 8, no. 2: 47. https://doi.org/10.3390/antiox8020047
APA StyleYang, Q.-Q., Cheng, L., Long, Z.-Y., Li, H.-B., Gunaratne, A., Gan, R.-Y., & Corke, H. (2019). Comparison of the Phenolic Profiles of Soaked and Germinated Peanut Cultivars via UPLC-QTOF-MS. Antioxidants, 8(2), 47. https://doi.org/10.3390/antiox8020047