Chronic Inhibition of Mitochondrial Dihydrolipoamide Dehydrogenase (DLDH) as an Approach to Managing Diabetic Oxidative Stress



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Oxidative Stress and Diabetes
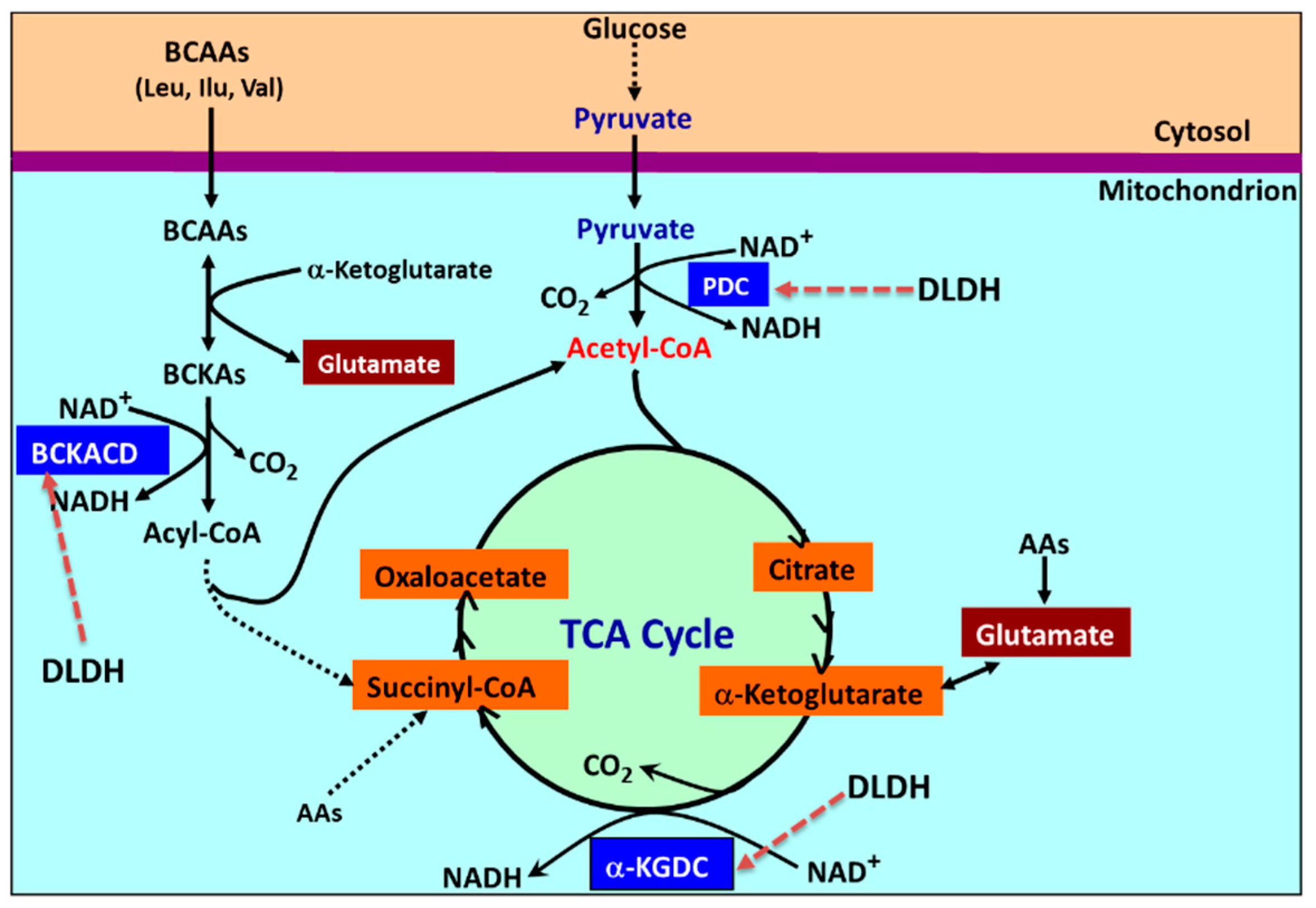
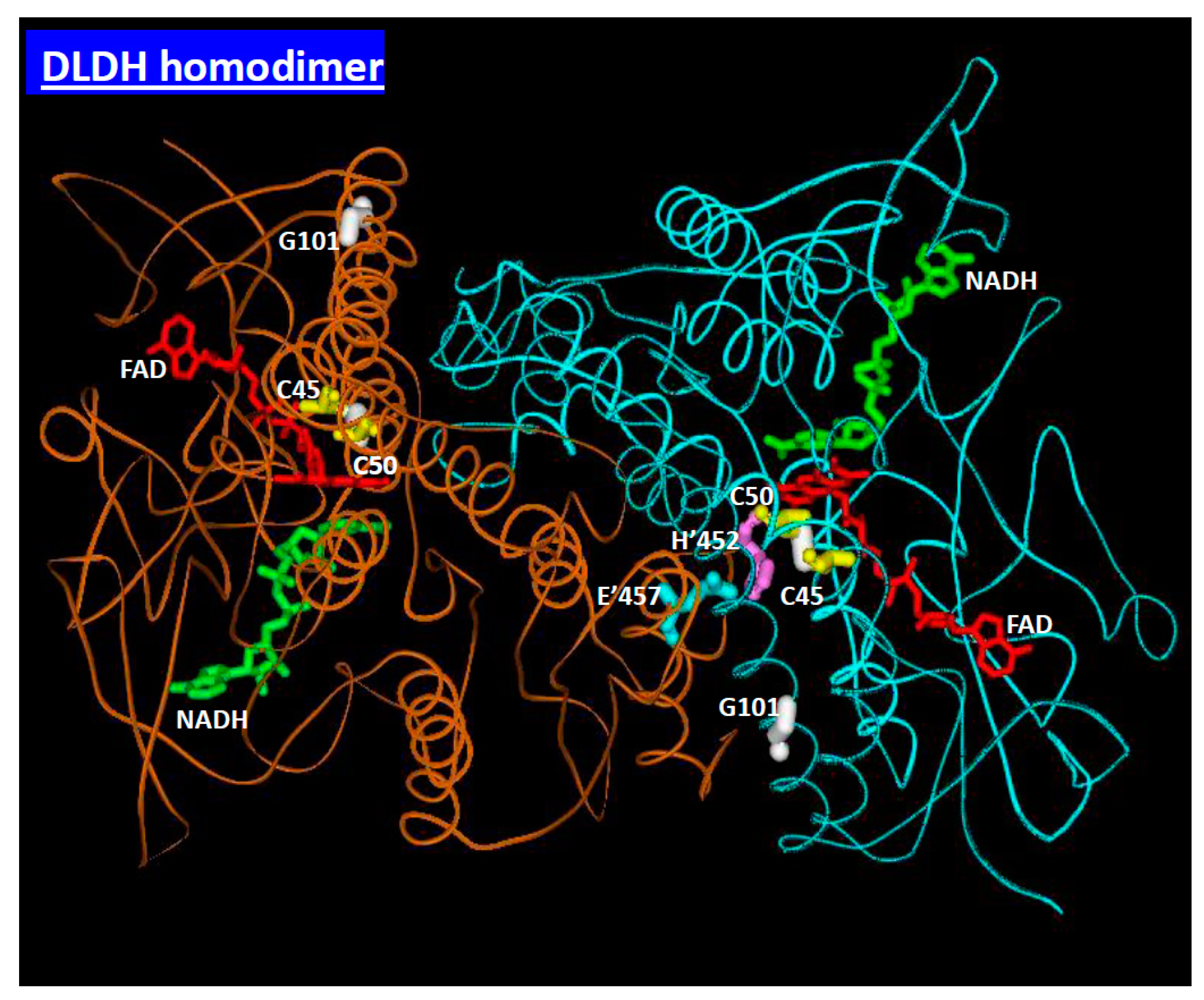
3. Mitochondrial Dihydrolipomide Dehydrogenase (DLDH)
4. Accumulation of Dihydrolipoamide
5. Accumulation of Pyruvate
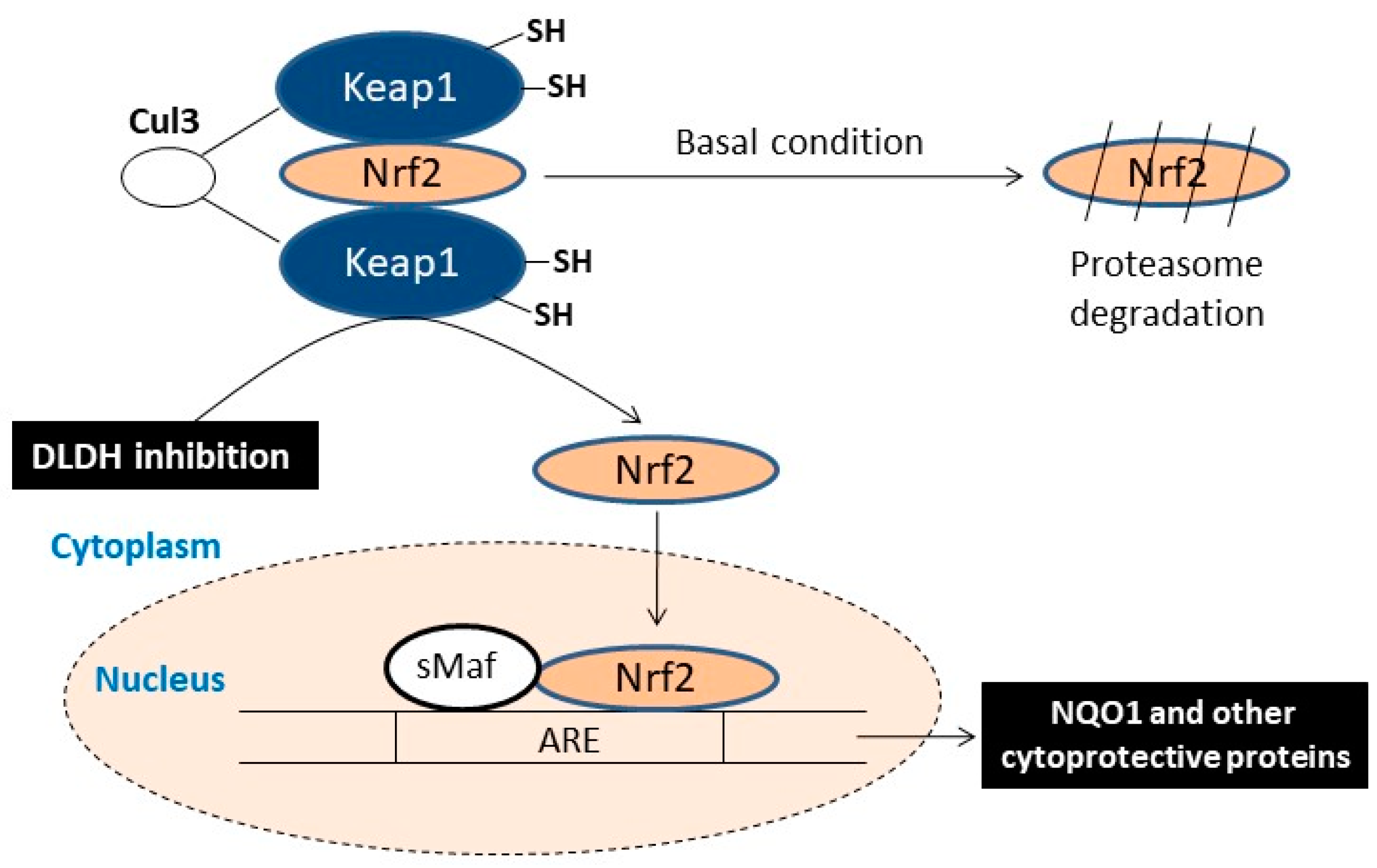
6. DLDH Chronic Inhibition and Activation of Nrf2 Signaling Pathway
7. Potential DLDH Inhibitors
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cernea, S.; Dobreanu, M. Diabetes and beta cell function: From mechanisms to evaluation and clinical implications. Biochem. Med. 2013, 23, 266–280. [Google Scholar] [CrossRef]
- Alzaid, A.; Rizza, R.A. Insulin resistance and its role in the pathogenesis of impaired glucose tolerance and non-insulin-dependent diabetes mellitus: Perspectives gained from in vivo studies. In Insulin Resistance; Moller, D.E., Ed.; John Wiley & Sons Ltd.: New York, NY, USA, 1993; pp. 143–186. [Google Scholar]
- Barnett, A.H. Type 2 Diabetes, 2rd ed.; Oxford University Press: Oxford, UK, 2012; p. 162. [Google Scholar]
- Tuch, B.; Dunlop, M.; Proietto, J. Diabetes Research: A Guide for Postgraduates; Harwood Academic Publishers: Reading, UK, 2000. [Google Scholar]
- Abdul-Ghani, M.A.; DeFronzo, R.A. Oxidative stress in type 2 diabetes. In Oxidative Stress in Aging; Miwa, S., Beckman, K.B., Muller, F.L., Eds.; Humana Press: New York, NY, USA, 2008; pp. 191–212. [Google Scholar]
- DeFronzo, R.A. Pathogenesis of type 2 diabetes mellitus. Med. Clin. N. Am. 2004, 88, 787–835. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jin, Z.; Zheng, H.; Yan, L.J. Sources and implications of nadh/nad (+) redox imbalance in diabetes and its complications. Diabetes Metab. Syndr. Obes. 2016, 9, 145–153. [Google Scholar]
- Yan, L.J. Pathogenesis of chronic hyperglycemia: From reductive stress to oxidative stress. J. Diabetes Res. 2014, 2014, 137919. [Google Scholar] [CrossRef] [PubMed]
- Rosen, P.; Nawroth, P.P.; King, G.; Moller, W.; Tritschler, H.J.; Packer, L. The role of oxidative stress in the onset and progression of diabetes and its complications: A summary of a congress series sponsored by unesco-mcbn, the american diabetes association and the german diabetes society. Diabetes Metab. Res. Rev. 2001, 17, 189–212. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Li, R.; Yan, L.J. Roles of pyruvate, nadh, and mitochondrial complex i in redox balance and imbalance in β cell function and dysfunction. J. Diabetes Res. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Ruskovska, T.; Bernlohr, D.A. Oxidative stress and protein carbonylation in adipose tissue—Implications for insulin resistance and diabetes mellitus. J. Proteom. 2013, 92, 323–334. [Google Scholar] [CrossRef]
- Robertson, R.P. Chronic oxidative stress as a central mechanism for glucose toxicity in pancreatic islet beta cells in diabetes. J. Biol. Chem. 2004, 279, 42351–42354. [Google Scholar] [CrossRef]
- Aragno, M.; Mastrocola, R. Dietary sugars and endogenous formation of advanced glycation endproducts: Emerging mechanisms of disease. Nutrients 2017, 9, 385. [Google Scholar] [CrossRef]
- Lovestone, S.; Smith, U. Advanced glycation end products, dementia, and diabetes. Proc. Natl. Acad. Sci. USA 2014, 111, 4743–4744. [Google Scholar] [CrossRef]
- Rosca, M.G.; Mustata, T.G.; Kinter, M.T.; Ozdemir, A.M.; Kern, T.S.; Szweda, L.I.; Brownlee, M.; Monnier, V.M.; Weiss, M.F. Glycation of mitochondrial proteins from diabetic rat kidney is associated with excess superoxide formation. Am. J. Physiol. Ren. Physiol. 2005, 289, F420–F430. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Jin, Z.; Yan, L.J. Redox imbalance and mitochondrial abnormalities in the diabetic lung. Redox Biol. 2017, 11, 51–59. [Google Scholar] [CrossRef]
- Beckman, K.B.; Ames, B.N. Oxidative decay of DNA. J. Biol. Chem. 1997, 272, 19633–19636. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.J.; Lodge, J.K.; Traber, M.G.; Packer, L. Apolipoprotein b carbonyl formation is enhanced by lipid peroxidation during copper-mediated oxidation of human low-density lipoproteins. Arch. Biochem. Biophys. 1997, 339, 165–171. [Google Scholar] [CrossRef]
- Yan, L.J. Analysis of oxidative modification of proteins. Curr. Protoc. Protein Sci. 2009, 56, 14.4.1–14.4.28. [Google Scholar]
- Yan, L.J.; Sohal, R.S. Gel electrophoretic quantitation of protein carbonyls derivatized with tritiated sodium borohydride. Anal. Biochem. 1998, 265, 176–182. [Google Scholar] [CrossRef]
- Pieper, A.A.; Brat, D.J.; Krug, D.K.; Watkins, C.C.; Gupta, A.; Blackshaw, S.; Verma, A.; Wang, Z.Q.; Snyder, S.H. Poly(adp-ribose) polymerase-deficient mice are protected from streptozotocin-induced diabetes. Proc. Natl. Acad. Sci. USA 1999, 96, 3059–3064. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Liaudet, L.; Soriano, F.G.; Mabley, J.G.; Szabo, E.; Szabo, C. The role of poly(adp-ribose) polymerase activation in the development of myocardial and endothelial dysfunction in diabetes. Diabetes 2002, 51, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.S.; Ho, E.C.; Lam, K.S.; Chung, S.K. Contribution of polyol pathway to diabetes-induced oxidative stress. J. Am. Soc. Nephrol. 2003, 14, S233–S236. [Google Scholar] [CrossRef]
- Li, Q.; Hwang, Y.C.; Ananthakrishnan, R.; Oates, P.J.; Guberski, D.; Ramasamy, R. Polyol pathway and modulation of ischemia-reperfusion injury in type 2 diabetic bbz rat hearts. Cardiovasc. Diabetol. 2008, 7, 33. [Google Scholar] [CrossRef]
- Oates, P.J. Polyol pathway and diabetic peripheral neuropathy. Int. Rev. Neurobiol. 2002, 50, 325–392. [Google Scholar] [PubMed]
- Yan, L.J. Redox imbalance stress in diabetes mellitus: Role of the polyol pathway. Anim. Model Exp. Med. 2018, 1, 7–13. [Google Scholar] [CrossRef]
- Hirschey, M.D.; Shimazu, T.; Jing, E.; Grueter, C.A.; Collins, A.M.; Aouizerat, B.; Stancakova, A.; Goetzman, E.; Lam, M.M.; Schwer, B.; et al. Sirt3 deficiency and mitochondrial protein hyperacetylation accelerate the development of the metabolic syndrome. Mol. Cell 2011, 44, 177–190. [Google Scholar] [CrossRef]
- Williams, C.H., Jr. Lipoamide dehydrogenase, glutathione reductase, thioredoxin reductase, and mercuric ion reductase-a family of flavoenzyme transhydrogenases. In Chemistry and Biochemistry of Flavoenzymes; Muller, F., Ed.; CRC Press: Boca Raton, FL, USA, 1992; Volume III, pp. 121–212. [Google Scholar]
- Yan, L.J.; Thangthaeng, N.; Forster, M.J. Changes in dihydrolipoamide dehydrogenase expression and activity during postnatal development and aging in the rat brain. Mech. Ageing Dev. 2008, 129, 282–290. [Google Scholar] [CrossRef]
- Yan, L.J.; Thangthaeng, N.; Sumien, N.; Forster, M.J. Serum dihydrolipoamide dehydrogenase is a labile enzyme. J. Biochem. Pharmacol. Res. 2013, 1, 30–42. [Google Scholar] [PubMed]
- Yan, L.J.; Yang, S.H.; Shu, H.; Prokai, L.; Forster, M.J. Histochemical staining and quantification of dihydrolipoamide dehydrogenase diaphorase activity using blue native page. Electrophoresis 2007, 28, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Babady, N.E.; Pang, Y.P.; Elpeleg, O.; Isaya, G. Cryptic proteolytic activity of dihydrolipoamide dehydrogenase. Proc. Natl. Acad. Sci. USA 2007, 104, 6158–6163. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, C.L.; Goncalves, R.L.; Hey-Mogensen, M.; Yadava, N.; Bunik, V.I.; Brand, M.D. The 2-oxoacid dehydrogenase complexes in mitochondria can produce superoxide/hydrogen peroxide at much higher rates than complex i. J. Biol. Chem. 2014. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Bjornstedt, M.; Nordman, T.; Eriksson, L.C.; Olsson, J.M. Reduction of ubiquinone by lipoamide dehydrogenase. An antioxidant regenerating pathway. Eur. J. Biochem. 2001, 268, 1486–1490. [Google Scholar] [CrossRef]
- Sreider, C.M.; Grinblat, L.; Stoppani, A.O. Catalysis of nitrofuran redox-cycling and superoxide anion production by heart lipoamide dehydrogenase. Biochem. Pharmacol. 1990, 40, 1849–1857. [Google Scholar] [CrossRef]
- Ambrus, A.; Torocsik, B.; Tretter, L.; Ozohanics, O.; Adam-Vizi, V. Stimulation of reactive oxygen species generation by disease-causing mutations of lipoamide dehydrogenase. Hum. Mol. Genet. 2011, 20, 2984–2995. [Google Scholar] [CrossRef]
- Chen, H.J.; Chen, Y.M.; Chang, C.M. Lipoyl dehydrogenase catalyzes reduction of nitrated DNA and protein adducts using dihydrolipoic acid or ubiquinol as the cofactor. Chem. Biol. Interact. 2002, 140, 199–213. [Google Scholar] [CrossRef]
- Yan, L.J.; Sumien, N.; Thangthaeng, N.; Forster, M.J. Reversible inactivation of dihydrolipoamide dehydrogenase by mitochondrial hydrogen peroxide. Free Radic. Res. 2013, 47, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.S.; Hong, Y.S. Lipoic acid as an antioxidant: The role of dihydrolipoamide dehydrogenase. In Free Radical and Antioxidant Protocols; Armstrong, D., Ed.; Humana Press: Totowa, NJ, USA, 1998; pp. 337–346. [Google Scholar]
- Jentoft, J.E.; Shoham, M.; Hurst, D.; Patel, M.S. A structural model for human dihydrolipoamide dehydrogenase. Proteins 1992, 14, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.C.; Kong, L.; Tan, T.W.; Ranganathan, S. Mpid-t: Database for sequence-structure-function information on t-cell receptor/peptide/mhc interactions. Appl. Bioinform. 2006, 5, 111–114. [Google Scholar] [CrossRef]
- Brautigam, C.A.; Chuang, J.L.; Tomchick, D.R.; Machius, M.; Chuang, D.T. Crystal structure of human dihydrolipoamide dehydrogenase: Nad+/nadh binding and the structural basis of disease-causing mutations. J. Mol. Biol. 2005, 350, 543–552. [Google Scholar] [CrossRef]
- Yan, L.J.; Liu, L.; Forster, M.J. Reversible inactivation of dihydrolipoamide dehydrogenase by angeli’s salt. Acta Biophys. Sin. (Sheng Wu Wu Li Hsueh Bao) 2012, 28, 341–350. [Google Scholar]
- Wu, J.; Li, R.; Li, W.; Ren, M.; Thangthaeng, N.; Sumien, N.; Liu, R.; Yang, S.; Simpkins, J.W.; Forster, M.J.; et al. Administration of 5-methoxyindole-2-carboxylic acid that potentially targets mitochondrial dihydrolipoamide dehydrogenase confers cerebral preconditioning against ischemic stroke injury. Free Radic. Biol. Med. 2017, 113, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.J.; Papa, S.; Bolanos, J.; Bruckdorfer, R.; Carlsen, H.; Elliott, R.M.; Flier, J.; Griffiths, H.R.; Heales, S.; Holst, B.; et al. Antioxidants, reactive oxygen and nitrogen species, gene induction and mitochondrial function. Mol. Asp. Med. 2002, 23, 209–285. [Google Scholar] [CrossRef]
- Son, S.M. Reactive oxygen and nitrogen species in pathogenesis of vascular complications of diabetes. Diabetes Metab. J. 2012, 36, 190–198. [Google Scholar] [CrossRef]
- Alzahrani, S.; Ebert, P.R. Oxygen and arsenite synergize phosphine toxicity by distinct mechanisms. Toxicol. Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Haramaki, N.; Han, D.; Handelman, G.J.; Tritschler, H.J.; Packer, L. Cytosolic and mitochondrial systems for nadh- and nadph-dependent reduction of alpha-lipoic acid. Free Radic. Biol. Med. 1997, 22, 535–542. [Google Scholar] [CrossRef]
- Bast, A.; Haenen, G.R. Lipoic acid: A multifunctional antioxidant. In Thiol Metabolism and Redox Regulation of Cellular Function; Pompella, A., Banhegyi, G., Wellman-Rousseau, M., Eds.; IOS Press: Amsterdam, The Netherlands, 2002; pp. 230–237. [Google Scholar]
- Packer, L.; Tritschler, H.J.; Wessel, K. Neuroprotection by the metabolic antioxidant alpha-lipoic acid. Free Radic. Biol. Med. 1997, 22, 359–378. [Google Scholar] [CrossRef]
- Panigrahi, M.; Sadguna, Y.; Shivakumar, B.R.; Kolluri, S.V.; Roy, S.; Packer, L.; Ravindranath, V. Alpha-lipoic acid protects against reperfusion injury following cerebral ischemia in rats. Brain Res. 1996, 717, 184–188. [Google Scholar] [CrossRef]
- Johnson, M.T.; Yang, H.S.; Magnuson, T.; Patel, M.S. Targeted disruption of the murine dihydrolipoamide dehydrogenase gene (dld) results in perigastrulation lethality. Proc. Natl. Acad. Sci. USA 1997, 94, 14512–14517. [Google Scholar] [CrossRef]
- Klivenyi, P.; Starkov, A.A.; Calingasan, N.Y.; Gardian, G.; Browne, S.E.; Yang, L.; Bubber, P.; Gibson, G.E.; Patel, M.S.; Beal, M.F. Mice deficient in dihydrolipoamide dehydrogenase show increased vulnerability to mptp, malonate and 3-nitropropionic acid neurotoxicity. J. Neurochem. 2004, 88, 1352–1360. [Google Scholar] [CrossRef]
- Ryou, M.G.; Liu, R.; Ren, M.; Sun, J.; Mallet, R.T.; Yang, S.H. Pyruvate protects the brain against ischemia-reperfusion injury by activating the erythropoietin signaling pathway. Stroke 2012, 43, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- Knott, E.M.; Sun, J.; Lei, Y.; Ryou, M.G.; Olivencia-Yurvati, A.H.; Mallet, R.T. Pyruvate mitigates oxidative stress during reperfusion of cardioplegia-arrested myocardium. Ann. Thorac. Surg. 2006, 81, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.B.; Barlow, M.A.; Yang, S.H.; Simpkins, J.W.; Mallet, R.T. Pyruvate enhances neurological recovery following cardiopulmonary arrest and resuscitation. Resuscitation 2008, 76, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Mallet, R.T.; Sun, J.; Knott, E.M.; Sharma, A.B.; Olivencia-Yurvati, A.H. Metabolic cardioprotection by pyruvate: Recent progress. Exp. Biol. Med. 2005, 230, 435–443. [Google Scholar] [CrossRef]
- Wang, X.; Perez, E.; Liu, R.; Yan, L.J.; Mallet, R.T.; Yang, S.H. Pyruvate protects mitochondria from oxidative stress in human neuroblastoma sk-n-sh cells. Brain Res. 2007, 1132, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jitrapakdee, S.; Vidal-Puig, A.; Wallace, J.C. Anaplerotic roles of pyruvate carboxylase in mammalian tissues. Cell. Mol. Life Sci. 2006, 63, 843–854. [Google Scholar] [CrossRef]
- Ying, W. Nad+/nadh and nadp+/nadph in cellular functions and cell death: Regulation and biological consequences. Antioxid. Redox Signal. 2008, 10, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, M.; De Groot, H. Nad(p)h, a directly operating antioxidant? FASEB J. 2001, 15, 1569–1574. [Google Scholar] [CrossRef]
- Yan, L.J.; Christians, E.S.; Liu, L.; Xiao, X.; Sohal, R.S.; Benjamin, I.J. Mouse heat shock transcription factor 1 deficiency alters cardiac redox homeostasis and increases mitochondrial oxidative damage. EMBO J. 2002, 21, 5164–5172. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.J.; Rajasekaran, N.S.; Sathyanarayanan, S.; Benjamin, I.J. Mouse hsf1 disruption perturbs redox state and increases mitochondrial oxidative stress in kidney. Antioxid. Redox Signal. 2005, 7, 465–471. [Google Scholar] [CrossRef]
- Wu, J.; Jin, Z.; Yang, X.; Yan, L.J. Post-ischemic administration of 5-methoxyindole-2-carboxylic acid at the onset of reperfusion affords neuroprotection against stroke injury by preserving mitochondrial function and attenuating oxidative stress. Biochem. Biophys. Res. Commun. 2018, 497, 444–450. [Google Scholar] [CrossRef]
- Sun, Z.; Park, S.Y.; Hwang, E.; Zhang, M.; Seo, S.A.; Lin, P.; Yi, T.H. Thymus vulgaris alleviates uvb irradiation induced skin damage via inhibition of mapk/ap-1 and activation of nrf2-are antioxidant system. J. Cell. Mol. Med. 2017, 21, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Du, J.; Hwang, E.; Yi, T.H. Paeonol extracted from paeonia suffruticosa andr. Ameliorated uvb-induced skin photoaging via dld/nrf2/are and mapk/ap-1 pathway. Phytother. Res. 2018, 32, 1741–1749. [Google Scholar] [CrossRef]
- Ahmad, W. Dihydrolipoamide dehydrogenase suppression induces human tau phosphorylation by increasing whole body glucose levels in a c. Elegans model of alzheimer’s disease. Exp. Brain Res. 2018, 236, 2857–2866. [Google Scholar] [CrossRef]
- Ahmad, W.; Ebert, P.R. 5-methoxyindole-2-carboxylic acid (mica) suppresses abeta-mediated pathology in C. elegans. Exp. Gerontol. 2018, 108, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.J. Reexploring 5-methoxyindole-2-carboxylic acid (mica) as a potential antidiabetic agent. Diabetes Metab. Syndr. Obes. 2018, 11, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Romoli, M.; Mazzocchetti, P.; D’Alonzo, R.; Siliquini, S.; Rinaldi, V.E.; Verrotti, A.; Calabresi, P.; Costa, C. Valproic acid and epilepsy: From molecular mechanisms to clinical evidences. Curr. Neuropharmacol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Luis, P.B.; Ruiter, J.P.; Aires, C.C.; Soveral, G.; Tavares de Almeida, I.; Duran, M.; Wanders, R.J.; Silva, M.F. Valproic acid metabolites inhibit dihydrolipoyl dehydrogenase activity leading to impaired 2-oxoglutarate-driven oxidative phosphorylation. Biochim. Biophys. Acta 2007, 1767, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, E.R.; Fischer, R.J.; Sugden, K.D.; Martin, B.D. Inhibition by methylated organo-arsenicals of the respiratory 2-oxo-acid dehydrogenases. J. Organomet. Chem. 2009, 694, 973–980. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Song, J.; Yan, L.-J. Chronic Inhibition of Mitochondrial Dihydrolipoamide Dehydrogenase (DLDH) as an Approach to Managing Diabetic Oxidative Stress. Antioxidants 2019, 8, 32. https://doi.org/10.3390/antiox8020032
Yang X, Song J, Yan L-J. Chronic Inhibition of Mitochondrial Dihydrolipoamide Dehydrogenase (DLDH) as an Approach to Managing Diabetic Oxidative Stress. Antioxidants. 2019; 8(2):32. https://doi.org/10.3390/antiox8020032
Chicago/Turabian StyleYang, Xiaojuan, Jing Song, and Liang-Jun Yan. 2019. "Chronic Inhibition of Mitochondrial Dihydrolipoamide Dehydrogenase (DLDH) as an Approach to Managing Diabetic Oxidative Stress" Antioxidants 8, no. 2: 32. https://doi.org/10.3390/antiox8020032
APA StyleYang, X., Song, J., & Yan, L.-J. (2019). Chronic Inhibition of Mitochondrial Dihydrolipoamide Dehydrogenase (DLDH) as an Approach to Managing Diabetic Oxidative Stress. Antioxidants, 8(2), 32. https://doi.org/10.3390/antiox8020032