Neopterin, Inflammation, and Oxidative Stress: What Could We Be Missing?



{kind=link}
Abstract
:1. Introduction to Neopterin and 7,8-Dihydroneopterin
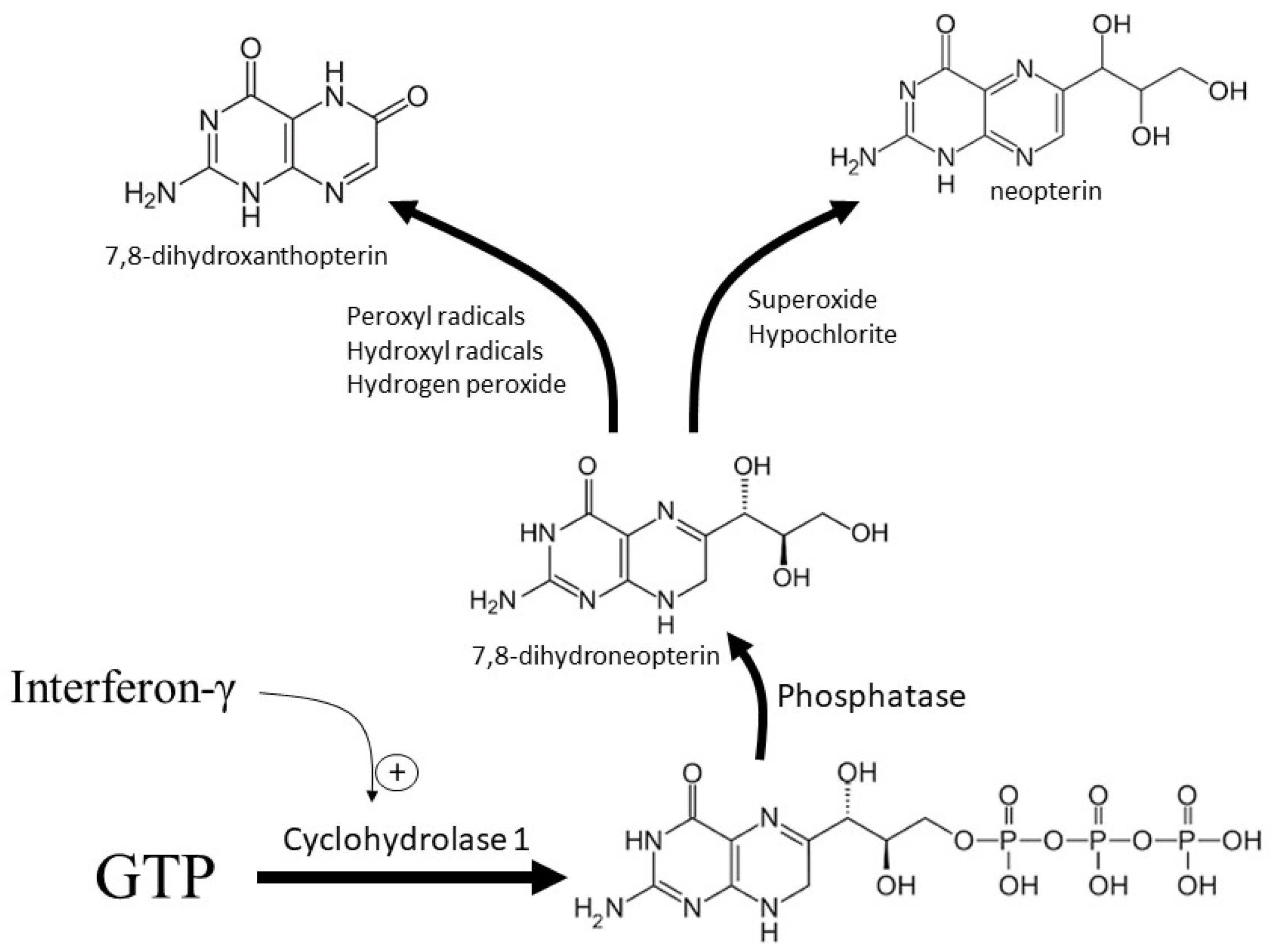
2. Synthesis within Macrophages
3. Biological Activity of 7,8-Dihydroneopterin
4. Measurement of Neopterin and Total Neopterin
5. Clinical Effectiveness of Neopterin and Total Neopterin
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Pedersen, E.R.; Midttun, O.; Ueland, P.M.; Schartum-Hansen, H.; Seifert, R.; Igland, J.; Nordrehaug, J.E.; Ebbing, M.; Svingen, G.; Bleie, O.; et al. Systemic Markers of Interferon-gamma-Mediated Immune Activation and Long-Term Prognosis in Patients With Stable Coronary Artery Disease. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Melichar, B.; Spisarova, M.; Bartouskova, M.; Krcmova, L.K.; Javorska, L.; Studentova, H. Neopterin as a biomarker of immune response in cancer patients. Ann. Transl. Med. 2017, 5, 280. [Google Scholar] [CrossRef] [PubMed]
- Baydar, T.; Yuksel, O.; Sahin, T.T.; Dikmen, K.; Girgin, G.; Sipahi, H.; Kurukahvecioglu, O.; Bostanci, H.; Sare, M. Neopterin as a prognostic biomarker in intensive care unit patients. J. Crit. Care 2009, 24, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M. Neopterin in Diagnosis and Monitoring of Infectious Diseases. J. Biomark. 2013, 2013, 196432. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.S.; Anzaldi, M.; Fiore, V.; Candido, S.; di Marco, R.; Mangano, K.; Neri, S. Neopterin: A potential marker in chronic peripheral arterial disease. Mol. Med. Rep. 2013, 7, 1855–1858. [Google Scholar] [CrossRef] [PubMed]
- Parrak, V.; Secnik, P.; Skrakova, M. Neopterin Screening of Blood Donations. Pteridines 2006, 17, 105–106. [Google Scholar] [CrossRef]
- Lindsay, A.; Lewis, J.; Scarrott, C.; Draper, N.; Gieseg, S.P. Changes in acute biochemical markers of inflammatory and structural stress in rugby union. J. Sports Sci. 2014, 33, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.; Carr, S.; Cross, S.; Petersen, C.; Lewis, J.G.; Gieseg, S.P. The physiological response to cold-water immersion following a mixed martial arts training session. Appl. Physiol. Nutr. Metab. 2017, 42, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.; Janmale, T.; Draper, N.; Gieseg, S.P. Measurement of changes in urinary neopterin and total neopterin in body builders using SCX HPLC. Pteridines 2014, 25, 53–62. [Google Scholar] [CrossRef]
- Huber, C.; Fuchs, D.; Hausen, A.; Margreiter, R.; Reibnegger, G.; Spielberger, M.; Wachter, H. Pteridines as a new marker to detect human T cells activated by allogeneic or modified self major histocompatibility complex (MHC) determinants. J. Immunol. 1983, 130, 1047–1050. [Google Scholar] [PubMed]
- Huber, C.; Batchelor, J.R.; Fuchs, D.; Hausen, A.; Lang, A.; Niederwieser, D.; Reibnegger, G.; Swetly, P.; Troppmair, J.; Wachter, H. Immune response associated production of neopterin- Release from macrophages primarily under control of interferon-gamma. J. Exp. Med. 1984, 160, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, T.; Nixon, J.C. Analysis of reduced forms of biopterin in biological tissues and fluids. Anal. Biochem. 1980, 102, 176–188. [Google Scholar] [CrossRef]
- Flavall, E.A.; Crone, E.M.; Moore, G.A.; Gieseg, S.P. Dissociation of neopterin and 7,8-dihydroneopterin from plasma components before HPLC analysis. J. Chromatogr. B 2008, 863, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Westermann, J.; Thiemann, F.; Gerstner, L.; Tatzber, F.; Kozak, I.; Bertsch, T.; Kruger, C. Evaluation of a new simple and rapid enzyme-linked immunosorbent assay kit for neopterin determination. Clin. Chem. Lab. Med. 2000, 38, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Estelberger, W.; Weiss, G.; Petek, W.; Paletta, B.; Wachter, H.; Reibnegger, G. Determination of Renal Clearance of Neopterin by a Pharmacokinetic Approach. FEBS Lett. 1993, 329, 13–16. [Google Scholar] [CrossRef]
- Fuchs, D.; Milstien, S.; Kramer, A.; Reibnegger, G.; Werner, E.R.; Goedert, J.J.; Kaufman, S.; Wachter, H. Urinary neopterin concentrations vs total neopterins for clinical utility. Clin. Chem. 1989, 35, 2305–2307. [Google Scholar] [PubMed]
- Shi, Z.G.; Wu, Y.B.; Luo, Y.B.; Feng, Y.Q. Analysis of Pterins in Urine by HILIC. Chromatographia 2010, 71, 761–768. [Google Scholar] [CrossRef]
- Schoedon, G.; Troppmair, J.; Fontana, A.; Huber, C.; Curtius, H.C.; Niederwieser, A. Biosynthesis and metabolism of pterins in peripheral-blood mononuclear-cells and leukemia lines of man and mouse. Eur. J. Biochem. 1987, 166, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Troppmair, J.; Nachbaur, K.; Herold, M.; Aulitzky, W.; Tilg, H.; Gastl, G.; Bieling, P.; Kotlan, B.; Flener, R.; Mull, B.; et al. In vitro and in vivo studies on the induction of neopterin biosynthesis by cytokines, alloantigens and lipopolysaccharides (LPS). Clin. Exp. Immunol. 1988, 74, 392–397. [Google Scholar] [PubMed]
- Muller, M.M.; Curtis, H.; Herold, M.; Huber, C.H. Neopterin in clinical practice. Clin. Chim. Acta 1991, 201, 1–16. [Google Scholar] [CrossRef]
- Werner, E.R.; Werner-Felmayer, G.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Yim, J.J.; Pfleiderer, W.; Wachter, H. Tetrahydrobiopterin biosynthetic activities in human macrophages, fibroblasts, THP-1 and T 24 cells. J. Biol. Chem. 1990, 265, 3189–3192. [Google Scholar] [PubMed]
- Yang, Y.T.; Whiteman, M.; Gieseg, S.P. Intracellular glutathione protects human monocyte-derived macrophages from hypochlorite damage. Life Sci. 2012, 90, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Reibnegger, G.; Wachter, H.; Esterbauer, H. 7,8-Dihydroneopterin inhibits low density lipoprotein oxidation in vitro. Evidence that this macrophage secreted pteridine is an antioxidant. Free Radic. Res. 1995, 23, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Duggan, S.; Rait, C.; Platt, A. Protein and thiol oxidation in cells exposed to peroxyl radicals, is inhibited by the macrophage synthesised pterin 7,8-dihydroneopterin. Biochim. Biophys. Acta 2002, 1591, 139–145. [Google Scholar]
- Duggan, S.; Rait, C.; Gebicki, J.M.; Gieseg, S.P. Inhibition of protein oxidation by the macrophage synthesised antioxidant 7,8-dihydroneopterin. Redox Rep. 2001, 6, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Pearson, J.; Firth, C.A. Protein hydroperoxides are a major product of low density lipoprotein oxidation during copper, peroxyl radical and macrophage-mediated oxidation. Free Radic. Res. 2003, 37, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Cato, S. Inhibition of THP-1 cell-mediated low-density lipoprotein oxidation by the macrophage-synthesised pterin, 7,8-dihydroneopterin. Redox Rep. 2003, 8, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Katouah, H.; Chen, A.; Othman, I.; Gieseg, S.P. Oxidised low density lipoprotein causes human macrophage cell death through oxidant generation and inhibition of key catabolic enzymes. Int. J. Biochem. Cell Biol. 2015, 67, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Shchepetkina, A.A.; Hock, B.D.; Miller, A.; Kennedy, M.A.; Gieseg, S.P. Effect of 7,8-dihydroneopterin mediated CD36 down regulation and oxidant scavenging on oxidised low-density lipoprotein induced cell death in human macrophages. Int. J. Biochem. Cell Biol. 2017, 87, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Baird, S.K.; Reid, L.; Hampton, M.; Gieseg, S.P. OxLDL induced cell death is inhibited by the macrophage synthesised pterin, 7,8-dihydroneopterin, in U937 cells but not THP-1 cells. Biochim. Biophys. Acta 2005, 1745, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Maghzal, G.; Glubb, D. Inhibition of haemolysis by the macrophage synthesized antioxidant, 7,8-dihydroneopterin. Redox Rep. 2000, 5, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Widner, B.; Mayr, C.; Wirleitner, B.; Fuchs, D. Oxidation of 7,8-Dihydroneopterin by hypochlorous acid yields neopterin. Biochem. Biophys. Res. Commun. 2000, 275, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Oettl, K.; Dikalov, S.; Freisleben, H.J.; Mlekusch, W.; Reibnegger, G. Spin trapping study of antioxidant properties of neopterin and 7,8-dihydroneopterin. Biochem. Biophys. Res. Commmun. 1997, 234, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Glubb, D.; Maghzal, G. Protection of erythrocytes by the macrophage synthesized antioxidant 7,8 dihydroneopterin. Free Radic. Res. 2001, 34, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.; Healy, J.; Mills, W.; Lewis, J.; Gill, N.; Draper, N.; Gieseg, S.P. Impact induced muscle damage and urinary pterins in professional rugby: 7,8-dihydroneopterin oxidation by myoglobin. Scand. J. Med. Sci. Sports 2016, 26, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S.; Nomura, T.; Icho, T.; Kajiwara, Y.; Kitabatake, K.; Kubota, K. Inhibitory effects of neopterin on NADPH-dependent superoxide-generating oxidase of rat peritoneal macrophages. FEBS Lett. 1993, 329, 125–128. [Google Scholar] [CrossRef]
- Oettl, K.; Reibnegger, G. Pteridine derivatives as modulators of oxidative stress. Curr. Drug Metab. 2002, 3, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Crone, E.M.; Flavall, E.A.; Amit, Z. Potential to inhibit growth of atherosclerotic plaque development through modulation of macrophage neopterin/7,8-dihydroneopterin synthesis. Br. J. Pharmacol. 2008, 153, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Gieseg, S.P.; Amit, Z.; Yang, Y.T.; Shchepetkina, A.; Katouah, H. Oxidant production, oxLDL uptake, and CD36 levels in human monocyte derived macrophages are down regulated by the macrophage generated antioxidant 7,8-dihydroneopterin. Antioxid. Redox Signal. 2010, 13, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Schobersberger, W.; Hoffmann, G.; Grote, J.; Wachter, H.; Fuchs, D. Induction of inducible nitric oxide synthase expression by neopterin in vascular smooth muscle cells. FEBS Lett. 1995, 377, 461–464. [Google Scholar] [PubMed] [Green Version]
- Rieder, J.; Amann, A.; Schloesser, M.; Czechowski, M.; Seibel, M.; Marth, C.; Hoffmann, G. Suppressive Effects of Neopterin on Inducible Nitric Oxide Synthase Gene Expression in Ovarian Carcinoma Cells in vitro. Pteridines 2001, 12, 140–146. [Google Scholar] [CrossRef]
- Schobersterger, W.; Hoffmann, G.; Hobisch-Hagen, P.; Bock, G.; Volkl, H.; Baier-Bitterlich, G.; Wirleitner, B.; Wachter, H.; Fuchs, D. Neopterin and 7,8-dihydroneopterin induce apoptosis in the rat alveolar epithelial cell line L2. FEBS Lett. 1996, 397, 263–268. [Google Scholar] [CrossRef] [Green Version]
- Wirleitner, B.; Baier-Bitterlich, G.; Boeck, G.; Widner, B.; Fuchs, D. 7,8-dihydroneopterin-induced apoptosis in Jurkat T lymphocytes: A comparison with anti-Fas-and hydrogen peroxide-mediated cell death. Biochem. Pharmacol. 1998, 56, 1181–1187. [Google Scholar] [CrossRef]
- Spottl, N.; Wirleitner, B.; Boeck, G.; Widner, B.; Fuchs, D.; Baier-Bitterlich, G. Reduced pteridine derivatives induce apoptosis in human neuronal NT2/HNT cells. Immunobiology 2000, 201, 478–491. [Google Scholar] [CrossRef]
- Enzinger, C.; Wirleitner, B.; Spottl, N.; Bock, G.; Fuchs, D.; Baier-Bitterlich, G. Reduced pteridine derivatives induce apoptosis in PC12 cells. Neurochem. Int. 2002, 41, 71–78. [Google Scholar] [CrossRef]
- Enzinger, C.; Wirleitner, B.; Lutz, C.; Bock, G.; Tomaselli, B.; Baier, G.; Fuchs, D.; Baier-Bitterlich, G. 7,8-Dihydroneopterin induces apoptosis of Jurkat T-lymphocytes via a Bcl-2-sensitive pathway. Eur. J. Cell Biol. 2002, 81, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, A.; Goto, M. Neopterin—Isolation from human urine. J. Biochem. 1967, 61, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Wachter, H.; Hausen, A.; Grassmayr, K. Increased urinary-excretion of neopterin in patients with malignant-tumors and with virus diseases. Hoppe-Seylers Z. Physiol. Chem. 1979, 360, 1957–1960. [Google Scholar] [PubMed]
- Neumann, M.C.; Sprenger, H.; Grebe, S.O.; Gemsa, D.; Reibnegger, G.; Lange, H.; Muller, T.F. Neopterin, serum amyloid A, and cytokine monitoring after renal transplantation. Pteridines 1998, 9, 113–121. [Google Scholar] [CrossRef]
- Hyland, K. Estimation of tetrahydro, dihydro and fully oxidised pterins by high-performance liquid chromatography using sequential electrochemical and fluorometric detection. J. Chromatogr. B Biomed. Sci. Appl. 1985, 343, 35–41. [Google Scholar] [CrossRef]
- Lunte, C.E.; Kissinger, P.T. Determination of pterins by liquid chromatography/electrochemistry. Methods Enzolmol. 1986, 122, 300–309. [Google Scholar]
- Werner, E.R.; Bichler, A.; Daxenbichler, G.; Fuchs, D.; Fuith, L.C.; Hausen, A.; Hetzel, H.; Reibnegger, G.; Wachter, H. Determination of neopterin in serum and urine. Clin. Chem. 1987, 33, 62–66. [Google Scholar] [PubMed]
- Sattler, W.; Leblhuber, F.; Walli, J.; Widner, B.; Fuchs, D. Cerebrospinal fluid levels of alpha-tocopherol and neopterin in patients with dementia. Pteridines 1999, 10, 220–224. [Google Scholar] [CrossRef]
- Firth, C.A.; Laing, A.D.; Baird, S.K.; Pearson, J.; Gieseg, S.P. Inflammatory sites as a source of plasma neopterin: Measurement of high levels of neopterin and markers of oxidative stress in pus drained from human abscesses. Clin. Biochem. 2008, 41, 1078–1083. [Google Scholar] [CrossRef] [PubMed]
- Allegri, G.; Netto, H.; Gomes, L.; de Oliveira, M.L.C.; Scalco, F.B.; Neto, F.R.D. Determination of six pterins in urine by LC-MS/MS. Bioanalysis 2012, 4, 1739–1746. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.R.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Wachter, H. Simultaneous determination of neopterin and creatine in serum with solid phase extraction and on line elution liquid chromatography. Clin. Chem. 1987, 33, 2028–2033. [Google Scholar] [PubMed]
- Dantola, M.L.; Vignoni, M.; Capparelli, A.L.; Lorente, C.; Thomas, A.H. Stability of 7,8-dihydropterins in air-equilibrated aqueous solutions. Helv. Chim. Acta 2008, 91, 411–425. [Google Scholar] [CrossRef]
- Ziegler, I. Synthesis and interferon-gamma controlled release of pteridines during activation of human peripheral blood mononuclear cells. Biochem. Biophys. Res. Commun. 1985, 132, 404–411. [Google Scholar] [CrossRef]
- Burton, C.; Shi, H.L.; Ma, Y.F. Daily variation and effect of dietary folate on urinary pteridines. Metabolomics 2016, 12, 10. [Google Scholar] [CrossRef]
- Giron, A.J.; Martin-Tornero, E.; Sanchez, M.C.H.; Meras, I.D.; Mansilla, A.E. A simple HPLC-ESI-MS method for the direct determination of ten pteridinic biomarkers in human urine. Talanta 2012, 101, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, D.; Hausen, A.; Reibnegger, G.; Werner, E.R.; Dierich, M.P.; Wachter, H. Neopterin as a marker for activated cell-mediated immunity:application in HIV infection. Immunol. Today 1988, 9, 150–155. [Google Scholar] [CrossRef]
- Fuchs, D.; Hausen, A.; Kofler, M.; Kosanowski, H.; Reibnegger, G.; Wachter, H. Neopterin as an index of immune response in patients with tuberculosis. Lung 1984, 162, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Maerkeralzer, G.; Diemer, O.; Strumper, R.; Rohe, M. Neopterin Production in Inflamed Knee Joints—High-Levels in Synovial-Fluids. Rheumatol. Int. 1986, 6, 151–154. [Google Scholar] [CrossRef]
- Yokoyama, K.; Tajima, M.; Yoshida, H.; Nakayama, M.; Tokutome, G.; Sakagami, H.; Hosoya, T. Plasma pteridine concentrations in patients with chronic renal failure. Nephrol. Dialysis Transplant. 2002, 17, 1032–1036. [Google Scholar] [CrossRef] [Green Version]
- Mildvan, D.; Spritzler, J.; Grossberg, S.E.; Fahey, J.L.; Johnston, D.M.; Schock, B.R.; Kagan, J. Serum neopterin, an immune activation marker, independently predicts disease progression in advanced HIV-1 infection. Clin. Infect. Dis. 2005, 40, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.D.; Khanna, S.; Kaminski, H.J.; Rao, J.S.; Merriam, A.P.; Richmonds, C.R.; Leahy, P.; Li, J.; Guo, W.; Andrade, F.H. A chronic inflammatory response dominates the skeletal muscle molecular signature in dystrophin-deficient mdx mice. Hum. Mol. Genet. 2002, 11, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, A.; Schmiechen, A.; Chamberlain, C.M.; Ervasti, J.M.; Lowe, D.A. Neopterin/7,8-dihydroneopterin is elevated in Duchenne muscular dystrophy patients and protects mdx skeletal muscle function, Experimental physiology. Exp. Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.; Costello, J.T. Realising the Potential of Urine and Saliva as Diagnostic Tools in Sport and Exercise Medicine. Sports Med. 2017, 47, 11–31. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Geiger, D.; Schauer, M.; Gatterer, H.; Burtscher, M.; Fuchs, D. Effects of Exhaustive Aerobic Exercise on Tryptophan-Kynurenine Metabolism in Trained Athletes. PLoS ONE 2016, 11, e0153617. [Google Scholar] [CrossRef] [PubMed]
- Moser, B.; Schroecksnadel, K.; Hörtnagl, H.; Rieder, J.; Fuchs, D.; Gottardis, M. Influence of Extreme Long Endurance Sports Activity on Neopterin Excretion. Pteridines 2008, 19, 114. [Google Scholar] [CrossRef]
- Lindsay, A.; Othman, M.I.; Prebble, H.; Davies, S.; Gieseg, S.P. Repetitive cryotherapy attenuates the in vitro and in vivo mononuclear cell activation response. Exp. Physiol. 2016, 101, 851–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, A.; Carr, S.; Othmana, M.I.I.; Marks, E.; Davies, S.; Petersen, C.; Draper, N.; Gieseg, S.P. The physiological and mononuclear cell activation response to cryotherapy following a mixed martial arts contest: A pilot study. Pteridines 2015, 26, 143–151. [Google Scholar] [CrossRef]
- Lindsay, A.; Lewis, J.G.; Scarrott, C.; Gill, N.; Gieseg, S.P.; Draper, N. Assessing the Effectiveness of Selected Biomarkers in the Acute and Cumulative Physiological Stress Response in Professional Rugby Union through Non-invasive Assessment. Int. J. Sports Med. 2015, 36, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.; Lewis, J.; Gill, N.; Gieseg, S.P.; Draper, N. Immunity, inflammatory and psychophysiological stress response during a competition of professional rugby union. Pteridines 2015, 26, 153–160. [Google Scholar] [CrossRef]
- Jakeman, P.M.; Weller, A.; Warrington, G. Cellular immune activity in response to increased training of elite oarsmen prior to Olympic competition. J. Sports Sci. 1995, 13, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Gunga, H.C.; Machotta, A.; Schobersberger, W.; Mittermayr, M.; Kirsch, K.; Koralewski, E.; Rocker, L. Neopterin, IgG, IgA, IgM, and Plasma Volume Changes During Long-Distance Running. Pteridines 2002, 13, 15–20. [Google Scholar] [CrossRef]
- De Lucas, R.D.; Caputo, F.; de Souza, K.M.; Sigwalt, A.R.; Ghisoni, K.; Silveira, P.C.L.; Remor, A.P.; Scheffer, D.D.; Guglielmo, L.G.; Latini, A. Increased platelet oxidative metabolism, blood oxidative stress and neopterin levels after ultra-endurance exercise. J. Sports Sci. 2014, 32, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.; Draper, N.; Lewis, J.; Gieseg, S.P.; Gill, N. Positional demands of professional rugby. Eur. J. Sport Sci. 2015, 15, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, A.; Lewis, J.; Gill, N.; Gieseg, S.P.; Draper, N. Effect of varied recovery interventions on markers of psychophysiological stress in professional rugby union. Eur. J. Sport Sci. 2015, 15, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Janmale, T.; Genet, R.; Crone, E.; Flavall, E.; Firth, C.; Pirker, J.; Roake, J.A.; Gieseg, S.P. Neopterin and 7,8-dihydroneopterin are generated within atherosclerotic plaque. Pteridines 2015, 26, 93–103. [Google Scholar] [CrossRef]
- Kettle, A.J.; Winterbourn, C.C. Myeloperoxidase: A key regulator of neutrophil oxidant production. Redox Rep. 1997, 3, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, S.; Okada, Y.; Sukhova, G.K.; Virmani, R.; Heinecke, J.W.; Libby, P. Macrophage myeloperoxidase regulation by granulocyte macrophage colony-stimulating factor in human atherosclerosis and implications in acute coronary syndromes. Am. J. Pathol. 2001, 158, 879–891. [Google Scholar] [CrossRef]
- Van Dyke, B.R.; Saltman, P. Hemoglobin: A mechanism for the generation of hydroxyl radicals. Free Radic. Biol. Med. 1996, 20, 985–989. [Google Scholar] [CrossRef]
- Giulivi, C.; Cadenas, E. Heme protein radicals: Formation, fate, and biological consequences. Free Radic. Biol. Med. 1998, 24, 269–279. [Google Scholar] [CrossRef]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gieseg, S.P.; Baxter-Parker, G.; Lindsay, A. Neopterin, Inflammation, and Oxidative Stress: What Could We Be Missing? Antioxidants 2018, 7, 80. https://doi.org/10.3390/antiox7070080
Gieseg SP, Baxter-Parker G, Lindsay A. Neopterin, Inflammation, and Oxidative Stress: What Could We Be Missing? Antioxidants. 2018; 7(7):80. https://doi.org/10.3390/antiox7070080
Chicago/Turabian StyleGieseg, Steven P., Gregory Baxter-Parker, and Angus Lindsay. 2018. "Neopterin, Inflammation, and Oxidative Stress: What Could We Be Missing?" Antioxidants 7, no. 7: 80. https://doi.org/10.3390/antiox7070080
APA StyleGieseg, S. P., Baxter-Parker, G., & Lindsay, A. (2018). Neopterin, Inflammation, and Oxidative Stress: What Could We Be Missing? Antioxidants, 7(7), 80. https://doi.org/10.3390/antiox7070080