Morphological Pathways of Mitochondrial Division

{kind=link}
{kind=link}
{kind=link}
Abstract
:Omne granulum e granuloAltmann, 1890 [1]
1. Early Recognition of Mitochondrial Division
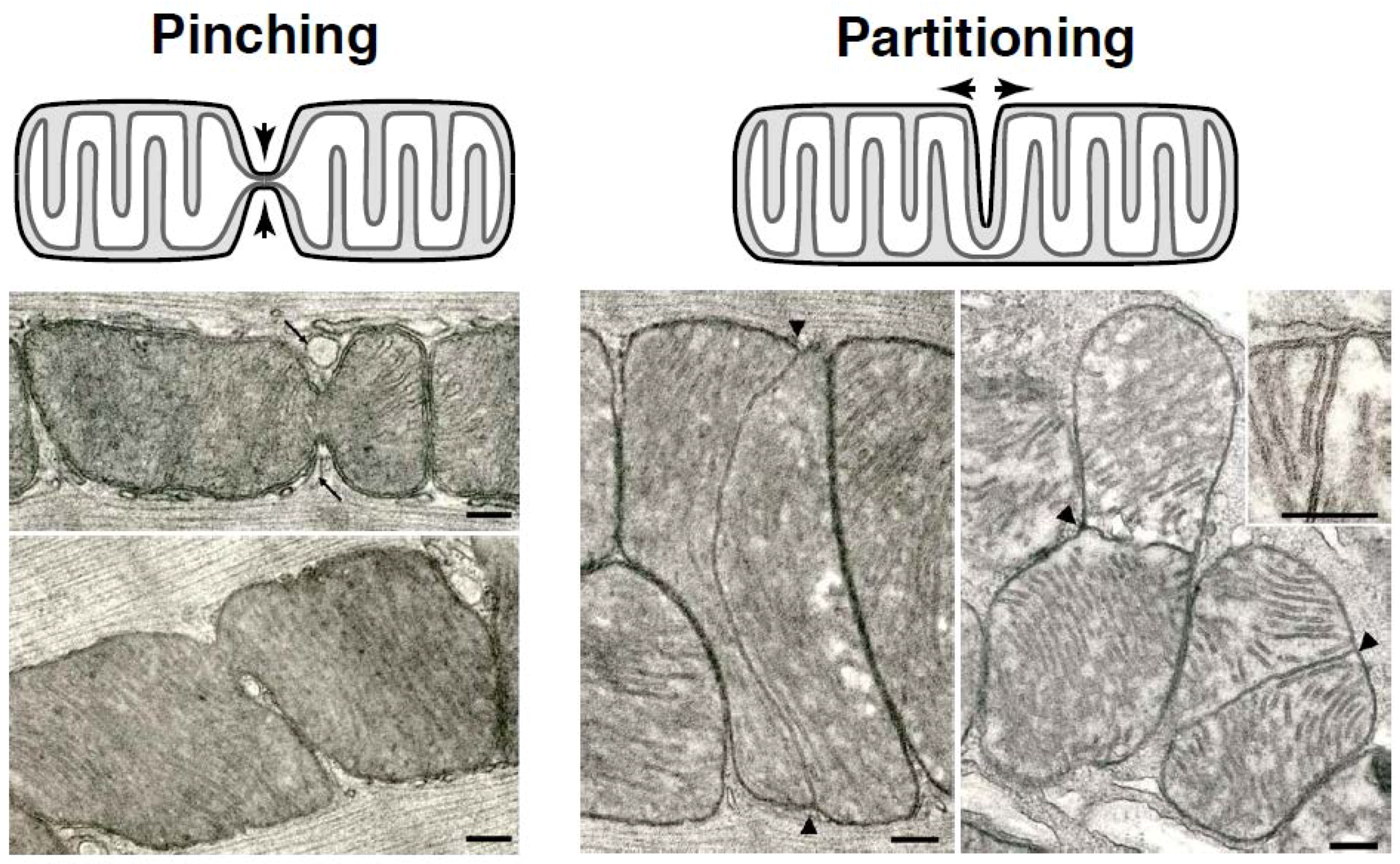
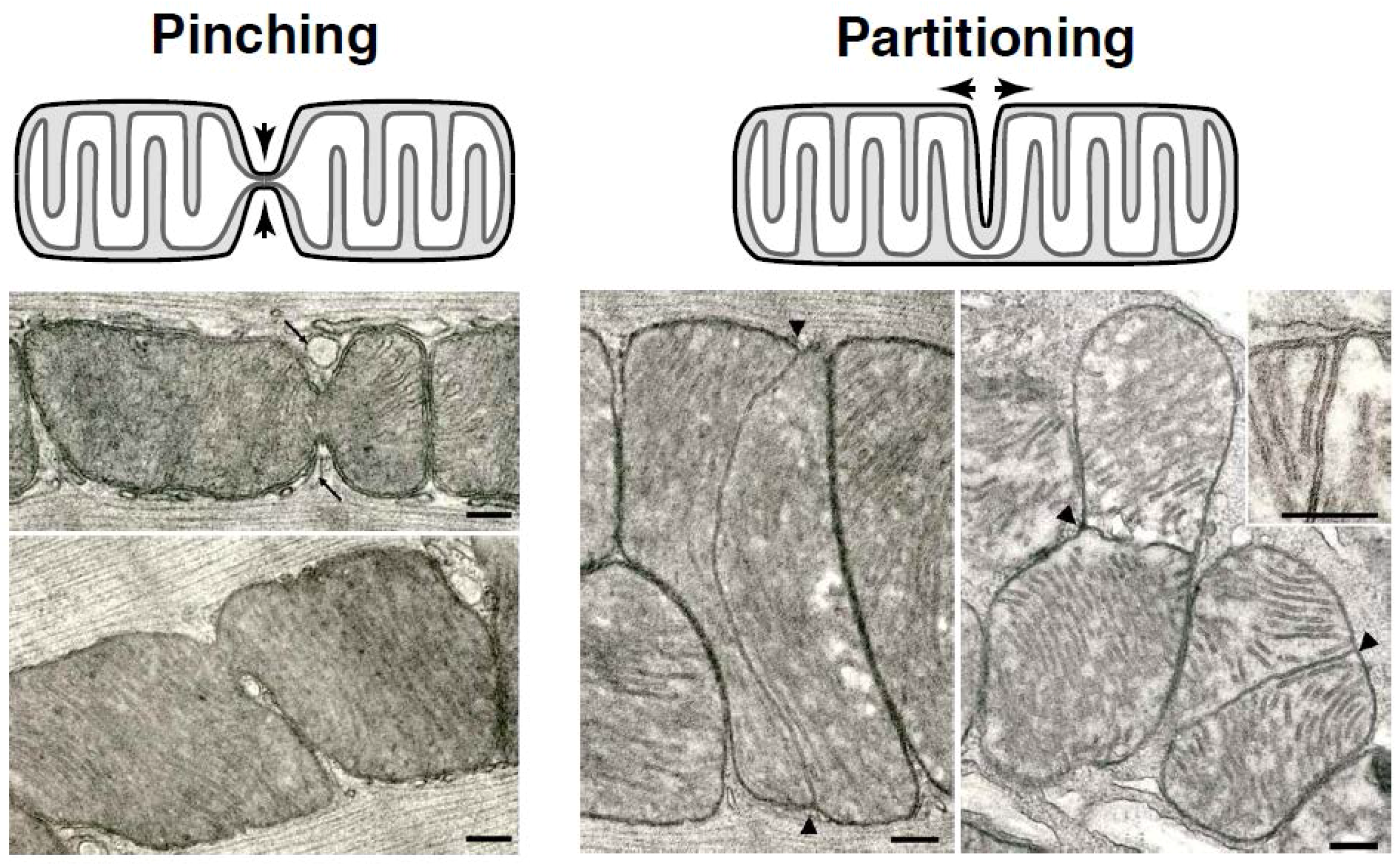
1.1. Mitochondrial Partitioning Is Observed
1.2. Mitochondrial Pinching—A Distinct Fission Pathway
1.3. Alternative Fission Pathways
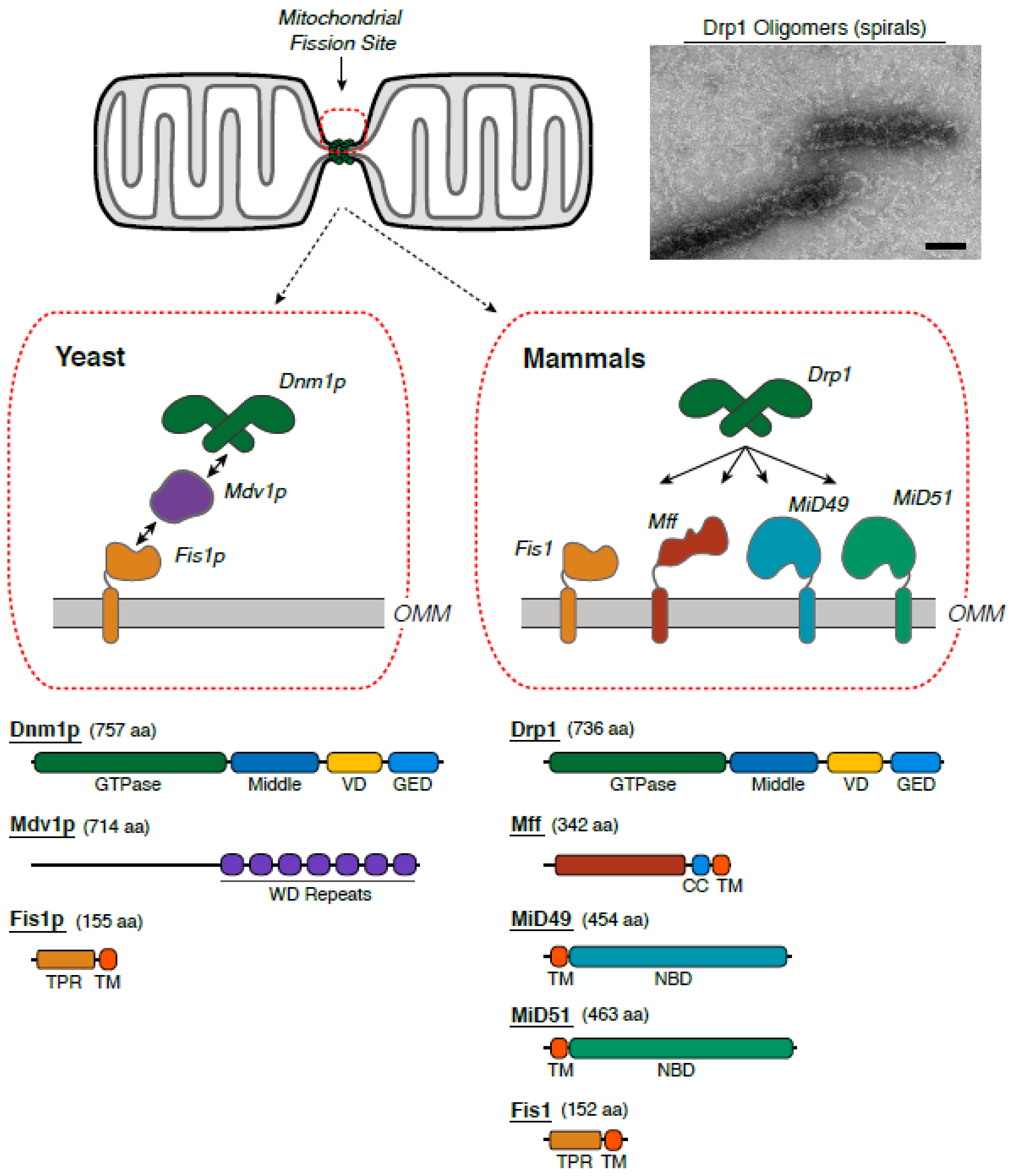
2. The Molecular Ensemble Driving Mitochondrial Division
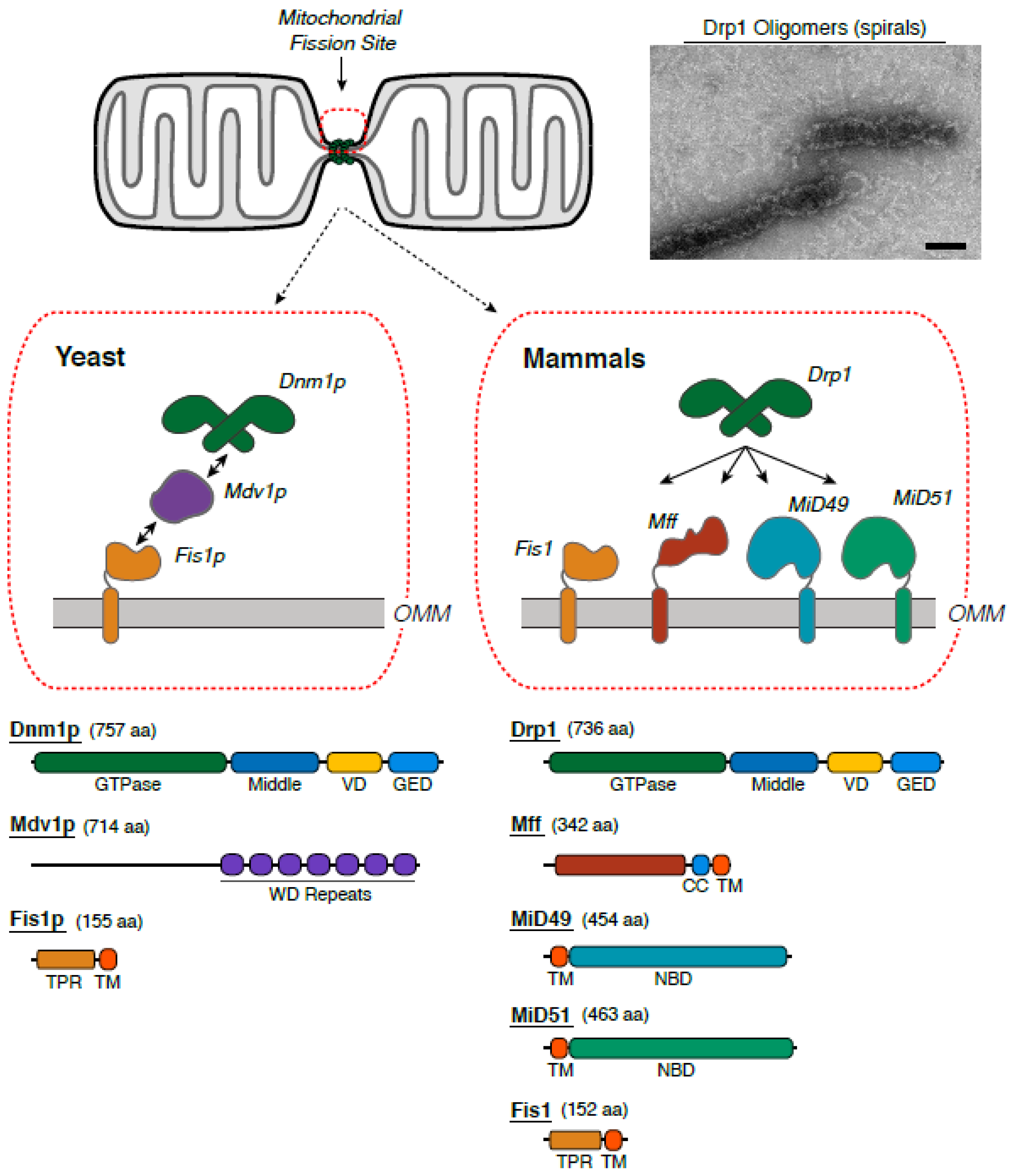
2.1. The Identification of Protein Machineries
2.2. Assorted Factors Regulate the Fission Machinery
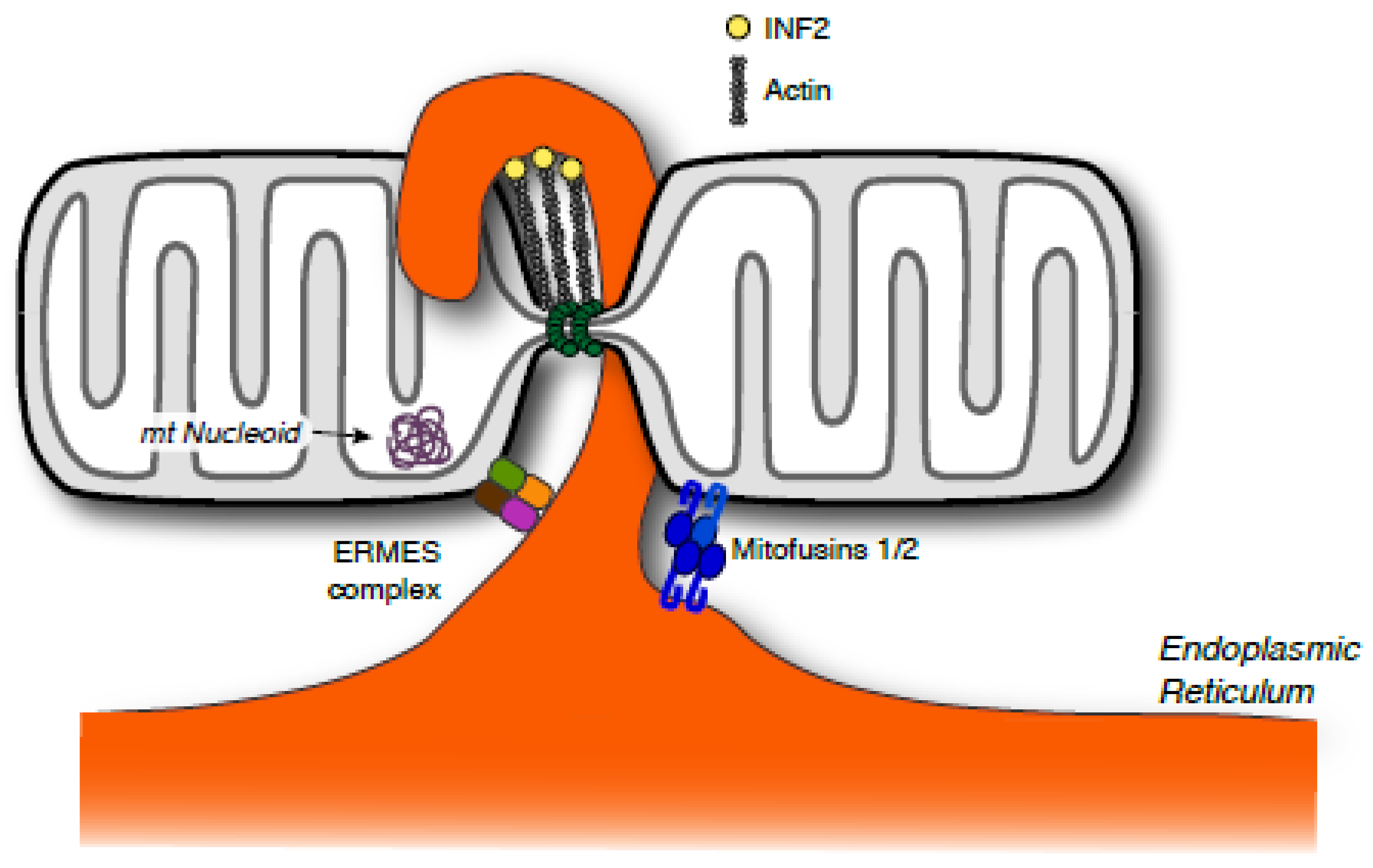
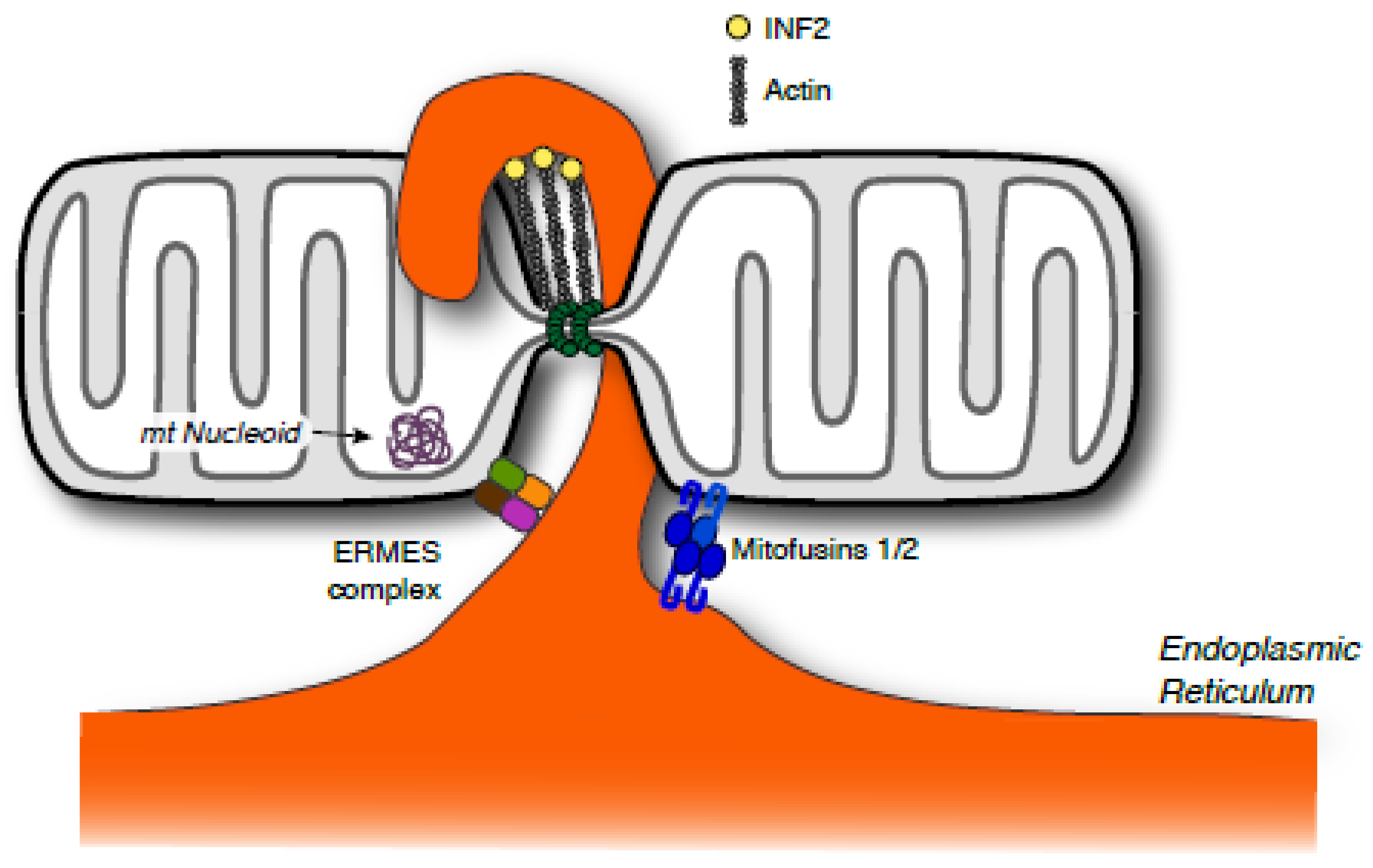
2.3. Inter-Organelle Connections Come to the Fore
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Altmann, R. Die Elementarorganismen Und Ihre Beziehungen Zu Den Zellen; The Elementary Organisms and Their Relationship to the Cell; Veit & Co.: Leipzig, Germany, 1890; p. 155. [Google Scholar]
- Siekevitz, P. Powerhouse of the Cell. Sci. Am. 1957, 197, 131–144. [Google Scholar] [CrossRef]
- Novikoff, A.B. Mitochondria (Chondriosomes). Cell 1961, II, 299–421. [Google Scholar]
- Lehninger, A.L. The Mitochondrion. Molecular Basis of Structure and Function; W. A. Benjamin, Inc.: New York, NY, USA; Amsterdam, The Netherlands, 1964. [Google Scholar]
- Nass, M.M.; Nass, S. Intramitochondrial Fibers with DNA Charachteristics. I. Fixation and Electron Staining Reactions. J. Cell Biol. 1963, 19, 593–611. [Google Scholar] [CrossRef] [PubMed]
- Rabinowitz, M.; Sinclair, J.; Desalle, L.; Haselkorn, R.; Swift, H.H. Isolation of Deoxyribonucleic Acid from Mitochondria of Chick Embryo Heart and Liver. Proc. Natl. Acad. Sci. USA 1965, 53, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Luck, D.F. Formation of Mitochondria in Neurospora Crassa. A Study Based on Mitochondrial Density Changes. J. Cell Biol. 1965, 24, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Luck, D.J. Formation of Mitochondria in Neurospora Crassa. A Quantitative Radioautographic Study. J. Cell Biol. 1963, 16, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.A.; Rustad, R.C. The Distribution of DNA among Dividing Mitochondria of Tetrahymena Pyriformis. J. Cell Biol. 1968, 37, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Tandler, B.; Erlandson, R.A.; Smith, A.L.; Wynder, E.L. Riboflavin and Mouse Hepatic Cell Structure and Function. II. Division of Mitochondria during Recovery from Simple Deficiency. J. Cell Biol. 1969, 41, 477–493. [Google Scholar] [CrossRef] [PubMed]
- Tandler, B.; Erlandson, R.A.; Wynder, E.L. Riboflavin and Mouse Hepatic Cell Structure and Function. I. Ultrastructural Alterations in Simple Deficiency. Am. J. Pathol. 1968, 52, 69–96. [Google Scholar] [PubMed]
- Tandler, B.; Hoppel, C.L. Ultrastructural Effects of Dietary Galactoflavin on Mouse Hepatocytes. Exp. Mol. Pathol. 1974, 21, 88–101. [Google Scholar] [CrossRef]
- Larsen, W.J. Genesis of Mitochondria in Insect Fat Body. J. Cell Biol. 1970, 47, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, D.W. The Cell, 2nd ed.; WB Saunders Company: Philadelphia, PA, USA, 1981. [Google Scholar]
- Tandler, B.; Hoppel, C.L. Studies on Giant Mitochondria. Ann. N. Y. Acad. Sci. 1986, 488, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Tandler, B.; Hoppel, C.L. Possible Division of Cardiac Mitochondria. Anat. Rec. 1972, 173, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, H.; Tandler, B.; Hoppel, C.L. Mitochondrial Division in Rat Cardiomyocytes: An Electron Microscope Study. Anat. Rec. 2012, 295, 1455–1461. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, D.M.; Jennings, D. Role of Copper in Mitochondrial Biogenesis via Interaction with ATP Synthase and Cytochrome C Oxidase. J. Bioenerg. Biomembr. 2002, 34, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K. Giant Hepatic Mitochondria: Production in Mice Fed with Cuprizone. Science 1969, 163, 81–82. [Google Scholar] [CrossRef] [PubMed]
- Tandler, B.; Hoppel, C.L. Division of Giant Mitochondria during Recovery from Cuprizone Intoxication. J. Cell Biol. 1973, 56, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Bowen, R.H. Studies on Insect Spermatogenesis: I. The History of the Cytoplasmic Components of the Sperm in Hemiptera. Biol. Bull. 1920, 39, 316–362. [Google Scholar] [CrossRef]
- Wilson, E.B. The Cell in Heredity and Development; Macmillan: New York, NY, USA, 1925. [Google Scholar]
- Wilson, E.B. The Distribution of the Chondriosomes to the Spermatozoa in Scorpions. Proc. Natl. Acad. Sci. USA 1916, 2, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Bereiter-Hahn, J. Intracellular Motility of Mitochondria: Role of the Inner Compartment in Migration and Shape Changes of Mitochondria in XTH-Cells. J. Cell Sci. 1978, 30, 99–115. [Google Scholar] [PubMed]
- Otsuga, D.; Keegan, B.R.; Brisch, E.; Thatcher, J.W.; Hermann, G.J.; Bleazard, W.; Shaw, J.M. The Dynamin-Related GTPase, Dnm1p, Controls Mitochondrial Morphology in Yeast. J. Cell Biol. 1998, 143, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Sesaki, H.; Jensen, R.E. Division Versus Fusion: Dnm1p and Fzo1p Antagonistically Regulate Mitochondrial Shape. J. Cell Biol. 1999, 147, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Heymann, J.A.; Hinshaw, J.E. Dynamins at a Glance. J. Cell Sci. 2009, 122, 3427–3431. [Google Scholar] [CrossRef] [PubMed]
- Praefcke, G.J.; Mcmahon, H.T. The Dynamin Superfamily: Universal Membrane Tubulation and Fission Molecules? Nat. Rev. Mol. Cell Biol. 2004, 5, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Legesse-Miller, A.; Massol, R.H.; Kirchhausen, T. Constriction and Dnm1p Recruitment are Distinct Processes in Mitochondrial Fission. Mol. Biol. Cell 2003, 14, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Bleazard, W.; Mccaffery, J.M.; King, E.J.; Bale, S.; Mozdy, A.; Tieu, Q.; Nunnari, J.; Shaw, J.M. The Dynamin-Related GTPase Dnm1 Regulates Mitochondrial Fission in Yeast. Nat. Cell Biol. 1999, 1, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Ingerman, E.; Perkins, E.M.; Marino, M.; Mears, J.A.; Mccaffery, J.M.; Hinshaw, J.E.; Nunnari, J. Dnm1 Forms Spirals That are Structurally Tailored to Fit Mitochondria. J. Cell Biol. 2005, 170, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Mears, J.A.; Lackner, L.L.; Fang, S.; Ingerman, E.; Nunnari, J.; Hinshaw, J.E. Conformational Changes in Dnm1 Support a Contractile Mechanism for Mitochondrial Fission. Nat. Struct. Mol. Biol. 2011, 18, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Mozdy, A.D.; Mccaffery, J.M.; Shaw, J.M. Dnm1p Gtpase-Mediated Mitochondrial Fission is a Multi-Step Process Requiring the Novel Integral Membrane Component Fis1p. J. Cell Biol. 2000, 151, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Tieu, Q.; Okreglak, V.; Naylor, K.; Nunnari, J. The WD Repeat Protein, Mdv1p, Functions as a Molecular Adaptor by Interacting with Dnm1p and Fis1p during Mitochondrial Fission. J. Cell Biol. 2002, 158, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Naylor, K.; Ingerman, E.; Okreglak, V.; Marino, M.; Hinshaw, J.E.; Nunnari, J. Mdv1 Interacts with Assembled Dnm1 to Promote Mitochondrial Division. J. Biol Chem. 2006, 281, 2177–2183. [Google Scholar] [CrossRef] [PubMed]
- Lackner, L.L.; Horner, J.S.; Nunnari, J. Mechanistic Analysis of a Dynamin Effector. Science 2009, 325, 874–877. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.; Shurland, D.L.; Ryazantsev, S.N.; Van Der Bliek, A.M. A Human Dynamin-Related Protein Controls the Distribution of Mitochondria. J. Cell Biol. 1998, 143, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, E.; Griparic, L.; Shurland, D.L.; Van Der Bliek, A.M. Dynamin-Related Protein Drp1 is Required for Mitochondrial Division in Mammalian Cells. Mol. Biol. Cell 2001, 12, 2245–2256. [Google Scholar] [CrossRef] [PubMed]
- James, D.I.; Parone, P.A.; Mattenberger, Y.; Martinou, J.C. hFis1, a Novel Component of the Mammalian Mitochondrial Fission Machinery. J. Biol. Chem. 2003, 278, 36373–36379. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; Krueger, E.W.; Oswald, B.J.; Mcniven, M.A. The Mitochondrial Protein hFis1 Regulates Mitochondrial Fission in Mammalian Cells through an Interaction with the Dynamin-Like Protein DLP1. Mol. Cell Biol. 2003, 23, 5409–5420. [Google Scholar] [CrossRef] [PubMed]
- Gandre-Babbe, S.; Van Der Bliek, A.M. The Novel Tail-Anchored Membrane Protein Mff Controls Mitochondrial and Peroxisomal Fission in Mammalian Cells. Mol. Biol. Cell 2008, 19, 2402–2412. [Google Scholar] [CrossRef] [PubMed]
- Otera, H.; Wang, C.; Cleland, M.M.; Setoguchi, K.; Yokota, S.; Youle, R.J.; Mihara, K. Mff is an Essential Factor for Mitochondrial Recruitment of Drp1 during Mitochondrial Fission in Mammalian Cells. J. Cell Biol. 2010, 191, 1141–1158. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.S.; Osellame, L.D.; Laine, D.; Koutsopoulos, O.S.; Frazier, A.E.; Ryan, M.T. MiD49 and MiD51, New Components of the Mitochondrial Fission Machinery. EMBO Rep. 2011, 12, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, T.; Jin, S.; Wang, X.; Qu, M.; Uhlen, P.; Tomilin, N.; Shupliakov, O.; Lendahl, U.; Nister, M. Human MIEF1 Recruits Drp1 to Mitochondrial Outer Membranes and Promotes Mitochondrial Fusion Rather than Fission. EMBO J. 2011, 30, 2762–2778. [Google Scholar] [CrossRef] [PubMed]
- Kashatus, J.A.; Nascimento, A.; Myers, L.J.; Sher, A.; Byrne, F.L.; Hoehn, K.L.; Counter, C.M.; Kashatus, D.F. Erk2 Phosphorylation of Drp1 Promotes Mitochondrial Fission and MAPK-Driven Tumor Growth. Mol. Cell 2015, 57, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Marsboom, G.; Toth, P.T.; Ryan, J.J.; Hong, Z.; Wu, X.; Fang, Y.H.; Thenappan, T.; Piao, L.; Zhang, H.J.; Pogoriler, J.; et al. Dynamin-Related Protein 1-Mediated Mitochondrial Mitotic Fission Permits Hyperproliferation of Vascular Smooth Muscle Cells and Offers a Novel Therapeutic Target in Pulmonary Hypertension. Circ. Res. 2012, 110, 1484–1497. [Google Scholar] [CrossRef] [PubMed]
- Mitra, K.; Wunder, C.; Roysam, B.; Lin, G.; Lippincott-Schwartz, J. A Hyperfused Mitochondrial State Achieved at G1-S Regulates Cyclin E Buildup and Entry into S Phase. Proc. Natl. Acad. Sci. USA 2009, 106, 11960–11965. [Google Scholar] [CrossRef] [PubMed]
- Sastre-Serra, J.; Nadal-Serrano, M.; Pons, D.G.; Roca, P.; Oliver, J. Mitochondrial Dynamics is Affected by 17beta-Estradiol in the MCF-7 Breast Cancer Cell Line. Effects on Fusion and Fission Related Genes. Int. J. Biochem. Cell Biol. 2012, 44, 1901–1905. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Wu, Q.; Horbinski, C.M.; Flavahan, W.A.; Yang, K.; Zhou, W.; Dombrowski, S.M.; Huang, Z.; Fang, X.; Shi, Y.; et al. Mitochondrial Control by DRP1 in Brain Tumor Initiating Cells. Nat. Neurosci. 2015, 18, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Gaume, B.; Bergmann-Leitner, E.S.; Leitner, W.W.; Robert, E.G.; Catez, F.; Smith, C.L.; Youle, R.J. The Role of Dynamin-Related Protein 1, a Mediator of Mitochondrial Fission, in Apoptosis. Dev. Cell 2001, 1, 515–525. [Google Scholar] [CrossRef]
- Fannjiang, Y.; Cheng, W.C.; Lee, S.J.; Qi, B.; Pevsner, J.; Mccaffery, J.M.; Hill, R.B.; Basanez, G.; Hardwick, J.M. Mitochondrial Fission Proteins Regulate Programmed Cell Death in Yeast. Genes Dev. 2004, 18, 2785–2797. [Google Scholar] [CrossRef] [PubMed]
- Arnoult, D.; Rismanchi, N.; Grodet, A.; Roberts, R.G.; Seeburg, D.P.; Estaquier, J.; Sheng, M.; Blackstone, C. Bax/Bak-Dependent Release of DDP/TIMM8a Promotes Drp1-Mediated Mitochondrial Fission and Mitoptosis during Programmed Cell Death. Curr. Biol. 2005, 15, 2112–2118. [Google Scholar] [CrossRef] [PubMed]
- Estaquier, J.; Arnoult, D. Inhibiting Drp1-Mediated Mitochondrial Fission Selectively Prevents the Release of Cytochrome C during Apoptosis. Cell Death Differ. 2007, 14, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.C.; Teng, X.; Park, H.K.; Tucker, C.M.; Dunham, M.J.; Hardwick, J.M. Fis1 Deficiency Selects for Compensatory Mutations Responsible for Cell Death and Growth Control Defects. Cell Death Differ. 2008, 15, 1838–1846. [Google Scholar] [CrossRef] [PubMed]
- Cereghetti, G.M.; Stangherlin, A.; Martins de Brito, O.; Chang, C.R.; Blackstone, C.; Bernardi, P.; Scorrano, L. Dephosphorylation by Calcineurin Regulates Translocation of Drp1 to Mitochondria. Proc. Natl. Acad. Sci. USA 2008, 105, 15803–15808. [Google Scholar] [CrossRef] [PubMed]
- Cribbs, J.T.; Strack, S. Reversible Phosphorylation of Drp1 by Cyclic Amp-Dependent Protein Kinase and Calcineurin Regulates Mitochondrial Fission and Cell Death. EMBO Rep. 2007, 8, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Saotome, M.; Safiulina, D.; Szabadkai, G.; Das, S.; Fransson, A.; Aspenstrom, P.; Rizzuto, R.; Hajnoczky, G. Bidirectional Ca2+-Dependent Control of Mitochondrial Dynamics by the Miro GTPase. Proc. Natl. Acad. Sci. USA 2008, 105, 20728–20733. [Google Scholar] [CrossRef] [PubMed]
- Slupe, A.M.; Merrill, R.A.; Flippo, K.H.; Lobas, M.A.; Houtman, J.C.; Strack, S. A Calcineurin Docking Motif (LXVP) in Dynamin-Related Protein 1 Contributes to Mitochondrial Fragmentation and Ischemic Neuronal Injury. J. Biol. Chem. 2013, 288, 12353–12365. [Google Scholar] [CrossRef] [PubMed]
- Strack, S.; Wilson, T.J.; Cribbs, J.T. Cyclin-Dependent Kinases Regulate Splice-Specific Targeting of Dynamin-Related Protein 1 to Microtubules. J. Cell Biol. 2013, 201, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Disatnik, M.H.; Shen, N.; Sobel, R.A.; Mochly-Rosen, D. Aberrant Mitochondrial Fission in Neurons Induced by Protein Kinase C{delta} Under Oxidative Stress Conditions in Vivo. Mol. Biol. Cell 2011, 22, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Kutty, S.; Toth, P.T.; Marsboom, G.; Hammel, J.M.; Chamberlain, C.; Ryan, J.J.; Zhang, H.J.; Sharp, W.W.; Morrow, E.; et al. Role of Dynamin-Related Protein 1 (Drp1)-Mediated Mitochondrial Fission in Oxygen Sensing and Constriction of the Ductus Arteriosus. Circ. Res. 2013, 112, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.; Leon, M.; Ponsoda, X.; Sendra, R.; Bort, R.; Ferrer-Lorente, R.; Raya, A.; Lopez-Garcia, C.; Torres, J. Early ERK1/2 Activation Promotes Drp1-Dependent Mitochondrial Fission Necessary for Cell Reprogramming. Nat. Commun. 2016, 7, 11124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.S.; Lin, S.C. AMPK Promotes Autophagy by Facilitating Mitochondrial Fission. Cell Metab. 2016, 23, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Toyama, E.Q.; Herzig, S.; Courchet, J.; Lewis, T.L.J.; Loson, O.C.; Hellberg, K.; Young, N.P.; Chen, H.; Polleux, F.; Chan, D.C.; et al. Metabolism. AMP-Activated Protein Kinase Mediates Mitochondrial Fission in Response to Energy Stress. Science 2016, 351, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Francy, C.A.; Alvarez, F.J.; Zhou, L.; Ramachandran, R.; Mears, J.A. The Mechanoenzymatic Core of Dynamin-Related Protein 1 Comprises the Minimal Machinery Required for Membrane Constriction. J. Biol. Chem. 2015, 290, 11692–11703. [Google Scholar] [CrossRef] [PubMed]
- Bustillo-Zabalbeitia, I.; Montessuit, S.; Raemy, E.; Basanez, G.; Terrones, O.; Martinou, J.C. Specific Interaction with Cardiolipin Triggers Functional Activation of Dynamin-Related Protein 1. PLoS ONE 2014, 9, e102738. [Google Scholar] [CrossRef] [PubMed]
- Stepanyants, N.; Macdonald, P.J.; Francy, C.A.; Mears, J.A.; Qi, X.; Ramachandran, R. Cardiolipin’s Propensity for Phase Transition and Its Reorganization by Dynamin-Related Protein 1 form a Basis for Mitochondrial Membrane Fission. Mol. Biol. Cell 2015, 26, 3104–3116. [Google Scholar] [CrossRef] [PubMed]
- Clinton, R.W.; Francy, C.A.; Ramachandran, R.; Qi, X.; Mears, J.A. Dynamin-Related Protein 1 Oligomerization in Solution Impairs Functional Interactions with Membrane-Anchored Mitochondrial Fission Factor. J. Biol. Chem. 2016, 291, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Itoh, K.; Yamada, T.; Cerveny, K.L.; Suzuki, T.L.; Macdonald, P.; Frohman, M.A.; Ramachandran, R.; Iijima, M.; Sesaki, H. Coincident Phosphatidic Acid Interaction Restrains Drp1 in Mitochondrial Division. Mol. Cell 2016, 63, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, P.J.; Stepanyants, N.; Mehrotra, N.; Mears, J.A.; Qi, X.; Sesaki, H.; Ramachandran, R. A Dimeric Equilibrium Intermediate Nucleates Drp1 Reassembly on Mitochondrial Membranes for Fission. Mol. Biol. Cell 2014, 25, 1905–1915. [Google Scholar] [CrossRef] [PubMed]
- Frohlich, C.; Grabiger, S.; Schwefel, D.; Faelber, K.; Rosenbaum, E.; Mears, J.; Rocks, O.; Daumke, O. Structural Insights into Oligomerization and Mitochondrial Remodelling of Dynamin 1-Like Protein. EMBO J. 2013, 32, 1280–1292. [Google Scholar] [CrossRef] [PubMed]
- Koirala, S.; Guo, Q.; Kalia, R.; Bui, H.T.; Eckert, D.M.; Frost, A.; Shaw, J.M. Interchangeable Adaptors Regulate Mitochondrial Dynamin Assembly for Membrane Scission. Proc. Natl. Acad. Sci. USA 2013, 110, E1342–E1351. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Chen, J.; Petrilli, A.; Liot, G.; Klinglmayr, E.; Zhou, Y.; Poquiz, P.; Tjong, J.; Pouladi, M.A.; Hayden, M.R.; et al. Mutant Huntingtin Binds the Mitochondrial Fission Gtpase Dynamin-Related Protein-1 and Increases Its Enzymatic Activity. Nat. Med. 2011, 17, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Yoon, Y.; Pitts, K.R.; Mcniven, M.A. Mammalian Dynamin-Like Protein DLP1 Tubulates Membranes. Mol. Biol. Cell 2001, 12, 2894–2905. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, P.J.; Francy, C.A.; Stepanyants, N.; Lehman, L.; Baglio, A.; Mears, J.A.; Qi, X.; Ramachandran, R. Distinct Splice Variants of Dynamin-Related Protein 1 Differentially Utilize Mitochondrial Fission Factor as an Effector of Cooperative GTPase Activity. J. Biol. Chem. 2016, 291, 493–507. [Google Scholar] [CrossRef] [PubMed]
- Francy, C.A.; Clinton, R.W.; Fröhlich, C.; Murphy, C.; Mears, J.A. Cryo-EM Studies of Drp1 Reveal Cardiolipin Interactions That Activate the Helical Oligomer. Sci. Rep. 2017, 7, 10744. [Google Scholar] [CrossRef] [PubMed]
- De Brito, O.M.; Scorrano, L. Mitofusin 2 Tethers Endoplasmic Reticulum to Mitochondria. Nature 2008, 456, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Rizzuto, R.; Hajnoczky, G.; Su, T.P. Mam: More than Just a Housekeeper. Trends Cell Biol. 2009, 19, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Mironov, S.L.; Symonchuk, N. ER Vesicles and Mitochondria Move and Communicate at Synapses. J. Cell Sci. 2006, 119, 4926–4934. [Google Scholar] [CrossRef] [PubMed]
- Shiao, Y.J.; Lupo, G.; Vance, J.E. Evidence That Phosphatidylserine is Imported into Mitochondria via a Mitochondria-Associated Membrane and That the Majority of Mitochondrial Phosphatidylethanolamine is Derived from Decarboxylation of Phosphatidylserine. J. Biol. Chem. 1995, 270, 11190–11198. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.R.; Lackner, L.L.; West, M.; Dibenedetto, J.R.; Nunnari, J.; Voeltz, G.K. ER Tubules Mark Sites of Mitochondrial Division. Science 2011, 334, 358–362. [Google Scholar] [CrossRef] [PubMed]
- Korobova, F.; Ramabhadran, V.; Higgs, H.N. An Actin-Dependent Step in Mitochondrial Fission Mediated by the ER-Associated Formin INF2. Science 2013, 339, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Kornmann, B.; Currie, E.; Collins, S.R.; Schuldiner, M.; Nunnari, J.; Weissman, J.S.; Walter, P. An ER-Mitochondria Tethering Complex Revealed by a Synthetic Biology Screen. Science 2009, 325, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Toulmay, A.; Prinz, W.A. A Conserved Membrane-Binding Domain Targets Proteins to Organelle Contact Sites. J. Cell Sci. 2012, 125, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Voss, C.; Lahiri, S.; Young, B.P.; Loewen, C.J.; Prinz, W.A. ER-Shaping Proteins Facilitate Lipid Exchange between the ER and Mitochondria in S. cerevisiae. J. Cell Sci. 2012, 125, 4791–4799. [Google Scholar] [CrossRef] [PubMed]
- Murley, A.; Lackner, L.L.; Osman, C.; West, M.; Voeltz, G.K.; Walter, P.; Nunnari, J. ER-Associated Mitochondrial Division Links the Distribution of Mitochondria and Mitochondrial DNA in Yeast. eLife 2013, 2, e00422. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, E.S.; Higgs, H.N. INF2 is a WASP Homology 2 Motif-Containing Formin That Severs Actin Filaments and Accelerates both Polymerization and Depolymerization. J. Biol. Chem. 2006, 281, 26754–26767. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Westrate, L.M.; Wu, H.; Page, C.; Voeltz, G.K. Multiple Dynamin Family Members Collaborate to Drive Mitochondrial Division. Nature 2016, 540, 139–143. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tandler, B.; Hoppel, C.L.; Mears, J.A. Morphological Pathways of Mitochondrial Division. Antioxidants 2018, 7, 30. https://doi.org/10.3390/antiox7020030
Tandler B, Hoppel CL, Mears JA. Morphological Pathways of Mitochondrial Division. Antioxidants. 2018; 7(2):30. https://doi.org/10.3390/antiox7020030
Chicago/Turabian StyleTandler, Bernard, Charles L. Hoppel, and Jason A. Mears. 2018. "Morphological Pathways of Mitochondrial Division" Antioxidants 7, no. 2: 30. https://doi.org/10.3390/antiox7020030
APA StyleTandler, B., Hoppel, C. L., & Mears, J. A. (2018). Morphological Pathways of Mitochondrial Division. Antioxidants, 7(2), 30. https://doi.org/10.3390/antiox7020030