Abstract
Green tea catechins (GTCs) are a family of chemically related compounds usually classified as antioxidant molecules. Epidemiological evidences, supported by interventional studies, highlighted a more than promising role for GTCs in human prostate cancer (PCa) chemoprevention. In the last decades, many efforts have been made to gain new insights into the mechanism of action of GTCs. Now it is clear that GTCs’ anticancer action can no longer be simplistically limited to their direct antioxidant/pro-oxidant properties. Recent contributions to the advancement of knowledge in this field have shown that GTCs specifically interact with cellular targets, including cell surface receptors, lipid rafts, and endoplasmic reticulum, modulate gene expression through direct effect on transcription factors or indirect epigenetic mechanisms, and interfere with intracellular proteostasis at various levels. Many of the effects observed in vitro are dose and cell context dependent and take place at concentrations that cannot be achieved in vivo. Poor intestinal absorption together with an extensive systemic and enteric metabolism influence GTCs’ bioavailability through still poorly understood mechanisms. Recent efforts to develop delivery systems that increase GTCs’ overall bioavailability, by means of biopolymeric nanoparticles, represent the main way to translate preclinical results in a real clinical scenario for PCa chemoprevention.
1. Introduction
Prostate cancer (PCa) is the second most common malignancy and a major cause of cancer deaths in men [1,2]. The increasing trend in PCa incidence reported over the last two decades, as well as differences in the geographic distribution of this value among different countries, show a good parallelism with differences in the use of the Prostate Specific Antigen (PSA) assay for PCa diagnosis. The highest number of screenings occurs in Western countries, where the highest incidence is found [3]. In contrast, the incidence of PCa is much lower in Asia and Africa, where PSA screening has not been widely adopted until now. Apart from differences caused by a scarce diffusion of the PSA test in some countries, which may lead to a reduced detection of latent PCa, different dietary habits and lifestyles may independently contribute to the highest risk of developing PCa in elder men who live in Western countries. In support of this concept, we have to keep in mind that there was already more than a 50-fold difference in international PCa incidence rates across Eastern and Western countries in 1980, before the PSA test was introduced [4]. To further emphasize the role of lifestyle in PCa development, migratory studies have found that Asian men who relocate to the United States and adopt a western lifestyle have a significantly higher risk to develop PCa than their native Asian counterparts [5,6]. PCa is an age related disease, highly heterogeneous and largely incurable at metastatic stages. The vast majority of prostate adenocarcinomas are characterized by a long latency period, which is generally clinically silent. Accordingly, only a few patients that receive a diagnosis of PCa will experience invasive and eventually metastatic cancer and will die of the cancer. The prognosis of high grade PCa is often unfavorable, and the surgical management of the localized disease is related to adverse events that negatively impact on the patients’ quality of life. For these reasons prevention may be the best approach to reduce PCa morbidity and mortality at the present time [7].
Among many the dietary agents investigated for chemopreventive properties against PCa, green tea derived polyphenols (GTPs) have received much attention [8]. In many European countries, the consumption of tea is occasional, whereas in Asian countries tea is extensively consumed as a beverage and has been associated with a reduced risk of advanced PCa [9].
Green tea is sourced from Camellia sinensis (L.) O. Kuntze (Theaceae), the same plant from which black and oolong teas are derived. The various tea types are classified on the basis of their manufacturing processes that influence taste, colour, and polyphenolic content composition. During green tea production, the endogenous oxidase enzymes in tealeaves are heating inactivated so that GTPs are preserved [10]. Flavonoids are the major active polyphenolic components of dried green tea extracts, in which flavan-3-ols account for more than 10% by weight [11]. Healthy properties of green tea have been associated with a family of flavan-3-ols called catechins. In decreasing order of abundance, the main green tea catechins (GTCs) are epigallocatechin gallate (EGCG), epigallocatechin (EGC), epicatechin gallate (ECG), epicatechin (EC), gallocatechin (GC), and catechin (C) (Figure 1).
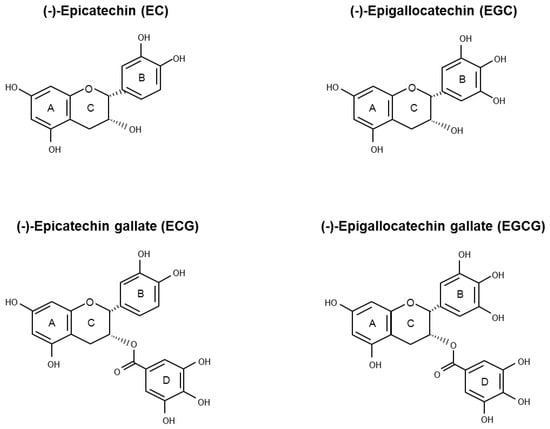
Figure 1.
Chemical structure of the four main green tea catechins (GTCs): (−)-epicatechin (EC), (−)-epigallocatechin (EGC), (−)-epicatechin gallate (ECG), and (−)-epigallocatechin gallate (EGCG).
EGCG accounts for about 40% of the total catechin content in green tea leaves and is the focus of much of the research on the health beneficial properties of green tea and green tea derived preparations. EGCG has significant growth inhibitory properties in PCa cells, with an observed half maximal inhibitory concentration (IC50) ranging from 40 μM to 80 μM, depending on the cell line used, the length of treatment, and the administration protocol [8].
It is noteworthy that the cytostatic action of GTCs is cancer specific, since EGCG is capable of inducing growth arrest both in SV-40 immortalized prostate epithelial cells and in PCa cells at concentrations that do not cause any toxic effect in normal human prostate epithelial cells [12,13,14].
The inhibition of PCa by GTCs and the prevention of tumor progression to the metastatic stage have been consistently documented in animal models that spontaneously develop PCa by many independent research groups [12,15,16]. Similar results have been documented also in men bearing precancerous lesions and achieving standardized doses of GTCs for one year [17,18]. The main obstacle to fully translate the more than promising results observed in preclinical models to large clinical trials is the identification of a reliable mechanism of action in vivo. Due to the very low bioavailability of dietary GTPs and standardized GTCs preparation, the mean plasmatic concentration reached by EGCG is between 50 to 100 times lower than the concentration used in vitro [19].
Recent works focus their attention on molecular or cellular mechanisms that are specifically altered during the process of cell transformation and that might be the in vivo target of GTCs, even at nanomolar concentrations.
This review will start discussing the antioxidant/pro-oxidant properties of GTCs, then will discuss recent experimental achievements on specific mechanisms of action that involve direct interaction with surface receptors and relevant intracellular targets, and then will move to describe observational and interventional studies that investigated the effectiveness and safety of GTCs in a real clinical setting (Figure 2). Attention will be paid to GTCs metabolism and bioavailability and, finally, to recent advances in the development of nanodelivery systems that improve GTCs bioactivity through increased stability, improved absorption, and reduced excretion.
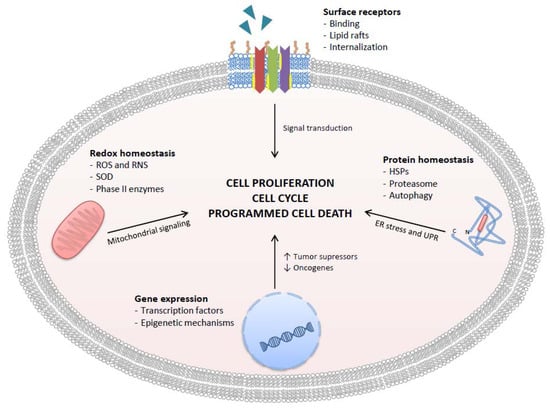
Figure 2.
Targets of GTCs in human PCa cells. GTCs inhibit cancer cell proliferation, cause cell cycle arrest, and promote programmed cell death by four main mechanisms: inhibition of ligand-receptor complex formation, redox homeostasis alteration, gene expression control, and protein homeostasis alteration. ROS = reactive oxygen species; RNS = reactive nitrogen species; SOD = superoxide dismutase; HSPs = heat shock proteins.
2. Effects of GTCs on Cellular Redox Homeostasis and Antioxidant Signaling
In both physiological and pathological conditions, reactive oxygen and nitrogen species (ROS and RNS, respectively) are produced. These reactive species are essential for cell survival and are involved in normal processes such as adaptation to hypoxia, regulation of autophagy, immunity, differentiation, and longevity [20]. A moderate ROS increase triggers the hormesis effect, an adaptive paradigmatic response to reactive species that helps cells to cope with oxidative stress through the activation of specific redox-cellular signaling pathways [21,22,23]. When the production of ROS and NOS overwhelms the cell capacity to scavenge them, oxidative stress occurs, leading to damage to biological molecules, loss of function, and eventually cell death.
The redox homeostasis imbalance is considered to be an important mechanism in promoting different diseases, including cancer onset and progression. Chronic exposure to elevated ROS concentration has been associated with PCa development [24]. Different studies have revealed that oxidative stress markers are higher in PCa cells (both in cell lines and human samples) in comparison to their non-pathological counterparts [24,25]. The cellular redox imbalance cooperates with other factors associated with PCa etiogenesis such as aging, androgen receptor activity, hormonal imbalance, and chronic inflammation. Different detoxification enzymes and antioxidant molecules are involved in maintaining the redox balance inside the cell. One of the most important mechanisms involved in superoxide detoxification involves the enzymes superoxide dismutases (SODs) that produce H2O2, which is in turn switched-off by catalase or glutathione peroxidase activity. Along with enzymatic activities, different endogenous compounds like glutathione, plasma protein thiols, and iron-binding proteins are needed to maintain the oxidative balance. Dietary components, like vitamins and polyphenols also contribute to maintaining intracellular redox homeostasis [26,27]. An increasing number of studies have demonstrated that GTPs are strong antioxidant compounds [21,28]; nonetheless GTPs are also able to exert pro-oxidant effects. The antioxidant/pro-oxidant behavior of GTPs greatly depends on many experimental conditions, such as the final concentration of use, temperature, pH, presence of metal ions, and differences in culture media composition [21,29,30,31]. All these variables make it very difficult to compare results, which are sometimes contradictory, published by various authors so far.
2.1. Effects of GTCs on Intracellular ROS Production
GTCs backbone (Figure 1) is characterized by the presence of two benzene rings (A- and B-ring) with two or more hydroxyl groups attached and a dihydropyran heterocycle (C-ring). The conjugation of hydroxyl groups on the C-ring with gallic acid (D-ring) leads to the formation of EGCG and ECG. The antioxidant properties of the GTCs are due to the presence of both the phenolic groups and the galloyl moiety, which allow electron delocalization and free radical quenching ability. According to their structural features, EGCG and ECG are, among the four most abundant GTCs, the strongest antioxidants [27,32]. However, GTCs may undergo auto-oxidation reactions, leading to the formation of catechin dimers and free radicals that may be responsible for the pro-oxidant action observed in vitro after polyphenol administration. The level of generated H2O2 is positively correlated with exposure time and the polyphenol concentration used. Incubation of 100 μM EGCG or EGC in different cell culture media for 1 h determined the generation of H2O2, ranging between 50.7 ÷ 71.6 µM and 98.1 ÷ 115 µM, respectively. However, at the concentration of 10 μM (a value that is about 10-fold higher than the values achievable in human plasma after ingestion of 800 mg of GTCs), the amount of generated H2O2 is very low [33]. Of note, the administration of EGCG up to 50 µM in different cell culture media in the presence of CHO cells exerts mild effects on cell viability, indicating that living cells possess efficient defense systems against H2O2 produced by GTCs in aqueous solutions [34]. Transition metals are able to initiate the catechins’ auto-oxidation reaction, also promoting the formation of catechins-metal ion complexes. Depending on the experimental model, these mechanisms have been correlated both to an antioxidant or a pro-oxidant behavior of GTCs [21,28]. Zinc is required for normal prostate metabolism due to its ability to inhibit aconitase activity, determining the excretion of citrate into seminal fluid. The ability of Zn2+ to accumulate in the prostate gland is lost during PCa progression [35]. Experimental data have shown that, in PCa cell lines, a mixture of Zn2+ and EGCG enhances both the growth inhibitory effect and the free radical scavenging ability with respect to EGCG alone. Moreover, Zn2+ enhances EGCG effects in terms of reduction of mitochondrial membrane potential and caspase-9 activation [36,37,38]. In PC-3 ML and in IBC-10a primary prostate cell lines, the administration of EGCG (up to 60 µM) determines a reduction of mitochondrial membrane potential and integrity, followed by apoptosis without increase in ROS production [39]. Chung and colleagues have observed that the incubation of DU145 cells with green tea extract (50 μg/mL) or pure green tea compounds (100 μM) for 48 h induces apoptosis through a peroxide and superoxide anions level increase and mitochondrial depolarization [40]. Kanwal and colleagues treated LNCaP cells with EGCG (20 µM) or Polyphenon E® (10 µg/mL), a standardized GTCs-enriched mixture that is caffeine free, containing 80% to 98% total catechins by weight. The authors have demonstrated that the 72 h-treatments make cells more resistant to both H2O2-induced oxidative stress and DNA damage [41].
We studied the effects of increasing doses of Polyphenon E® on ROS production and cell viability in PCa cell lines. We reported that concentrations of Polyphenon E® yielding a significant increase in ROS production (5 ÷ 20 µg/mL and 5 ÷ 50 µg/mL for PNT1a and PC3, respectively) displayed marginal effects on cell proliferation. A similar effect on ROS production was obtained by treating cells with H2O2 (10 µM and 50 µM for PNT1a and PC3, respectively). Higher concentrations of Polyphenon E® that caused a significant reduction of cell proliferation (25 ÷ 35 µg/mL and 75 ÷ 145 µg/mL for PNT1a and PC3, respectively) produced an amount of ROS below that of untreated cells or cells given H2O2. There is not good correlation between ROS induction and cell death in PCa cells following Polyphenon E® treatment. Therefore, it is very unlikely ROS production plays a direct role in GTCs induced cell death. Nonetheless, this process is strictly associated with the induction of endoplasmic reticulum stress, a mechanism that impacts on protein homeostasis, finally leading to cell death [42].
2.2. Effects of GTCs on SOD Activity
Apart from the direct effect on ROS production, EGCG is also involved in the modulation of SODs activity. Many studies, reviewed in [43], suggest that manganese superoxide dismutase (MnSOD) participates in the maintenance of mitochondrial integrity in cells exposed to oxidative stress. Morrissey and colleagues have shown that EGCG treatment (20 µM for 72 h) of NRP-154, a tumorigenic rat prostate epithelial cell line, determines a reduction of the expression of MnSOD, but not of catalase, concomitantly with apoptosis induction [43]. On the contrary, in DU145 cells, MnSOD expression was not affected by the administration of 3 µM EGCG, while a significant reduction was observed after treatment with ionizing radiation (IR). In the same experimental model, the combination of EGCG and IR significantly reduced IR-induced apoptosis, together with an increase of MnSOD expression. Therefore EGCG might promote radiotherapy resistance, inducing MnSOD [44]. In contrast, in Lobund-Wistar rats, the chronic consumption of green tea decreases the tumor incidence of genitourinary tissues in association with an increase of MnSOD expression, without effects on DNA damage, lipid peroxidation, and lipofuscin deposition [45]. Treatment of TRAMP (Transgenic Adenocarcinoma of the Mouse Prostate) mice for 13 weeks with EGCG (administered by gavage at a dosage of 200 mg/kg body weight), starting at 7 weeks of age, was associated with a significant reduction of severity/focalness of the histopathological lesions observed in the ventral lobe of the murine prostate. These effects are unlikely due to a direct antioxidant effect of EGCG; indeed the authors observed only a slight reduction in plasma H2O2 concentration and no significant effect on SOD expression [46]. Later on, in the same experimental model, the authors found that EGCG reduced the DNA oxidative damage marker but had no effects on the protein and lipid peroxidation markers [47].
2.3. Effects of GTCs on -RNS Production
Nitric oxide is a free radical produced by three different isoforms of nitric oxide synthase (NOS). NO• actions are mediated through cGMP-dependent or cGMP-independent pathways. Moreover, NO• at low concentrations can interact with different cellular compounds like other free radicals, proteins, and DNA. NO• exerts a dual role in cancer cells, which is dose dependent. At low concentrations (less than 100 nM), NO• is associated with tumor cell proliferation and angiogenesis; at high concentrations (more than 500 nM), NO• exerts a cytotoxic effect [48]. The anticancer effect of EGCG, through the inhibition of NO• production, has been observed in many different tumors [48,49,50,51]. In TRAMP mice who received 0.06% EGCG in tap water ad libitum for seven weeks starting at weaning, a reduction of the High Grade Prostate Intraepithelial Neoplasia (HG-PIN) incidence was observed concomitantly with a significant down-regulation of iNOS [52].
3. Specific Molecular Mechanisms of Action of GTCs
Apart from their antioxidant/pro-oxidant activity, GTCs have a high affinity with many biomolecules, including phospholipid bilayers, proteins, and nucleic acids. It has been shown that GTCs interact with cell surface receptors or enter the cells and directly interact with specific intracellular molecular targets [53,54]. Once GTCs have interfered with the ligand-receptor binding, or more generally with receptor activation, the effects are reflected in intracellular signaling pathways that control cell proliferation/cell death through the activation/inhibition of regulatory proteins (kinases/phosphatases) or effector molecules. GTCs interfere with gene transcription both by direct regulation of transcription factors and by epigenetic mechanisms that produce modifications of chromatin accessibility. GTCs may regulate gene expression also at the post transcriptional level by a mechanism that involves the expression of particular miRNAs (microRNAs). Of particular note, in the prostate, an endocrine tissue characterized by huge protein synthesis activity, GTCs have been shown to interfere with molecules, cellular compartments, or mechanisms reputed to control protein homeostasis, such as molecular chaperons, the endoplasmic reticulum (ER), and the main protein degradation mechanisms.
3.1. Cell Surface Receptors
GTCs interfere directly or indirectly with the formation of the ligand-receptor complex of several cell surface receptors [55,56]. Three different mechanisms have been proposed to explain the effect of GTCs on the regulation of the activity of cell surface receptors and their signaling pathways. The first mechanism proposes that GTCs physically prevent the interaction of the ligand with its cognate receptor by completely covering the cell surface through a “sealing effect” [57] or binding to the ligand [58,59] or competing with the ligand for the receptor [60].
The second mechanism involves the suppression of the intrinsic activity of lipid rafts-associated signaling proteins by lipid rafts disruption [55,61,62]. The third one prevents the interaction between receptor and ligand, promoting receptor clearance through physical sequestration inside endosomal vesicles [63].
Receptor tyrosine kinases (RTKs) have been recently identified as targets of GTCs in cancer cell growth inhibition [55,64,65]. The binding of a growth factor or a cytokine to the extracellular domain of RTKs induces the dimerization and autophosphorylation of the RTKs on specific tyrosine residues and the activation of downstream intracellular signaling that includes Ras/ERK/MAPK and PI3K/Akt. Ras/ERK/MAPK and PI3K/Akt activation triggers a cascade of molecular events involving enzymes, proteins, and transcription factors that have been extensively reviewed in [66]. The activation of cell surface RTKs controls many key important processes in normal cells such as cell proliferation, differentiation, survival, and migration. These receptor-associated pathways have been found altered in tumor cells, including PCa [55,66,67,68,69]. The RTKs family is comprised of epidermal growth factor receptors (EGFRs), fibroblast growth factor receptors (FGFRs), insulin-like growth factor receptors (IGF-R), platelet-derived growth factor receptors (PDGFRs), vascular endothelial growth factor receptors (VEGFRs), and hepatocyte growth factor receptors (HGFRs), also known as Met. The most effective inhibition toward RTKs is exerted by EGCG, followed by ECG and EGC [67].
GTCs administration inhibited EGFR activation in several cancer cell lines, even at concentrations as low as 1 µg/mL [70,71,72,73]. Liang et al. demonstrated that EGCG directly prevents the interaction between ligand and EGFR necessary for the tyrosine kinase activity in human epidermoid carcinoma cells (IC50 = 0.5 ÷ 1 µg/mL) [59].
Similar effects of GTCs have been observed towards insulin-like growth factor receptor 1 (IGF-R1) [74]. IGF-R1 is overexpressed in PCa [67]. Interestingly Li et al. demonstrated that EGCG competitively binds to the ATP binding site of IGF-R1, inhibiting its kinase activity in vitro (IC50 = 14 µmol/L) and in vivo [60]. The blockage of ligand-receptor binding has been suggested as the mechanism behind the inhibitory effect of GTCs on PDGFR and HGFR in multiple cell lines at concentrations of 1 ÷ 10 µM [58,75,76,77]. Tachibana et al. proposed that EGCG is a ligand of the 67-kDa laminin receptor (67-LR) with a dissociation constant (Kd) value of 0.004 µM [78]. 67-LR expression is elevated in cancer cells, where it acts as a cancer-specific cell death receptor. EGCG pro-apoptotic effects in multiple myeloma cells are abrogated by 67-LR silencing, suggesting that EGCG induced cell death is achieved through a 67-LR mediated mechanism [79]. Proof of the direct binding between EGCG and 67-LR has been recently provided by Shukla and colleagues. These authors have shown that the delivery of radioactive gold nanoparticles (NPs) functionalized with EGCG in PCa cells was mediated via the 67-LR. When the 67-LR was blocked by laminin or by anti-67LR antibody, the uptake of NPs was prevented [80]. Recent data have confirmed that EGCG undergoes oligomer formation by binding to the 67-LR [81]. In particular it has been identified a sensing motif on 67-LR, located within the peptide LR161-170, which is critical for the binding of EGCG [82].
The second mechanism proposed to explain the ubiquitous effect of EGCG on multiple receptors involves the disruption of lipid organization in the plasma membrane [61]. Lipid rafts are high ordered membrane structures, rich in cholesterol and sphingolipids, that regulate protein-protein interaction, receptor activation, and therefore, more generally, cellular signaling. EGCG alters the lipid organization of the plasma membrane and causes the rearrangement of lipid rafts [83], preventing the activation of cell surface receptors. In multiple myeloma cells, EGCG promoted apoptosis through lipid rafts clustering that, in turn, mediated the activation of 67-LR and triggered pro-death signaling [84,85,86,87]. GTCs inhibited RTKs activation through lipid rafts clustering. This mechanism has been reported for EGFR, HGFR, IGF-R1, and VEGFR [61,62,88,89,90,91].
Finally Adachi et al. found that the EGFR is quickly internalized inside endosomal vesicles in human colon cancer cells treated with EGCG (1 µg/mL for 30 min). The internalization makes the receptor inaccessible to epidermal growth factor and abrogates RTKs signaling cascade [63].
3.2. Effects of GTCs on Regulation of Gene Expression
Although the mechanisms of the cancer chemopreventive effect of GTCs have not been completely elucidated, there is evidence that EGCG administration results in the modulation of gene expression [12,92]. It has been demonstrated that EGCG modulates activity and expression of various transcription factors including Sp1, NF-kB, AP-1, STAT1, STAT3, and nuclear factor erythroid 2-related factor 2 (Nrf2) [27]. Among these, Nrf2 and NF-kB are two redox-sensitive transcription factors responding to inflammatory/oxidative stress stimuli that control the expression of many genes involved in PCa onset and progression. It is now accepted that free radicals act as redox cellular messengers, triggering signaling pathways able to influence the redox homeostasis. One of the most important cellular responses to cope with oxidative stress is the activation of Nrf2, which in turn activates a plethora of enzymes. Keap1, the Nrf2 inhibitor, is sensitive to redox imbalance. In presence of oxidants or electrophilic compounds, Keap1 dissociates from Nrf2, which then translocates to the nucleus and binds to electrophile-responsive element (EpRE) sequences, driving the transcription of phase II enzymes such as glutathione peroxidase (GPX), glutathione S-transferase (GST), NADPH quinone oxidoreductase 1 (NQO-1), and UDP-glucuronosyltransferase (UGT) [93,94]. Nrf2 also acts through Keap1 independent mechanisms [95,96]. It is important to highlight that GTCs are able to activate multiple pathways, such as MAPKs, PI3K, PKC, and NF-kB, which may play a crucial role in the Nrf2-mediated response to oxidative stress [93,95]. EGCG, and more generally GTCs, exert their anticancer effect in many different tumor models through the induction of a beneficial moderate oxidative stress, which in turn activates the Nrf2/EpRE response [93,97]. A direct target of Nrf2 is the gene GSTP1, which is downregulated in PCa. The effects of GTCs on GSTP1 re-expression are discussed in the next paragraph. NF-kB is constitutively activated in PCa cells, prostate premalignant lesions (intraepithelial tumors, PIN), and PCa tissues [98]. EGCG has been demonstrated to inhibit NF-kB transcriptional activity in DU145 and LNCaP cells, promoting apoptosis via Bax up-regulation and Bcl2 down-regulation [99,100]. Siddiqui et al. reported that the constitutive activation of NF-kB, observed during PCa progression, was prevented in TRAMP mice receiving 0,1% GTCs in drinking water from weaning, in comparison to water-fed age matched controls [101]. One of the targets of NF-kB is represented by matrix metalloproteinases (MMPs), a family of proteolytic enzymes that degrade collagene and other extracellular matrix proteins contributing to PCa progression. GTCs have been reported to inhibit various MMPs in PCa cells [99,102] and in TRAMP mice [16]. In silico molecular docking analyses have recently shown a strong direct interaction between EGCG and MMP-9, suggesting that EGCG might inhibit MMP-9 also by an NF-kB independent mechanism [103].
Epigenetic Mechanisms
Epigenetics generally refers to changes in gene expression and chromatin organization that are independent of alterations in the DNA sequence. Epigenetic phenomena are modifiable by environmental factors that include dietary habits [104]. Cancer development involves a complex multistep process caused by and associated with many genetic insults that include genetic mutations as well epigenetic alterations [105]. Many tumor suppressor and receptor genes have been reported to be hypermethylated and transcriptionally silenced during the development of various human cancers, including PCa [106]. The major epigenetic mechanisms that control gene expression are DNA methylation, histone modifications, and expression of noncoding regulatory micro RNA (miRNAs). Extensive in vitro experiments reviewed by Henning et al. in [104] have been performed in a variety of cancer cell lines to evaluate the effect of GTCs or, more generally, GTPs on DNA methylation. There is evidence that GTCs cancer preventive effect can be ascribed to the epigenetic reactivation of silenced gene through the inhibition of DNA methyltransferases (DNMTs) activity. Fang et al. demonstrated that EGCG binds to DNMT and competitively inhibits the enzymatic activity (Ki of 6.89 µM), yielding the reactivation of methylation-silenced genes in PC3 cells [107]. Molecular modeling and docking studies supported the binding of EGCG to DNMT3B in HeLa cells [108]. EGCG (5 ÷ 20 µM) or Polyphenon E® (1 ÷ 10 µg/mL) treatments of different PCa cell lines have determined a dose- and time-dependent re-expression of GSTP1 enzyme concomitantly with the down-regulation of DNMT1 [41,106]. The re-expression of GSTP1, induced by treatment with EGCG or Polyphenon E®, may be, at least in part, responsible for free radical species quenching and the reduction of DNA damage associated with oxidative stress [41]. Two studies concluded that GTCs did not produce a significant effect on DNA methylation in PCa cell lines and in TRAMP mice [109,110]. Potential reasons for these discrepancies might be due to different methods of analysis used to evaluate DNA methylation, different cell lines used, or inefficacy of the treatment protocol. Nonetheless, Henning et al. fed SCID mice implanted with LAPC4 androgen-dependent PCa cells with brewed green tea (containing 0.075% GTPs) or water for 13 weeks. The authors found that tumor growth and the expression of DNMT1 were reduced in xenograft tumors excised from the green tea-fed mice in comparison to the controls [111]. These results support the concept that GTCs may produce epigenetic changes in vivo following long-term administration.
The direct inhibition of EGCG on DNMT1 is enhanced in the presence of catechol-O-methyltransferase (COMT). The mechanism proposed by Lee et al. to explain this additive effect is due to the alteration of the S-adenosyl-methionine (SAM) and S-adenosyl-homocysteine (SAH) intracellular ratio. Indeed, galloylated GTCs are methylated by COMT, which catalyzes the transfer of a methyl group from the donor molecule SAM that in turn forms SAH. By this mechanism, the SAM/SAH is reduced and inhibits DNMTs activity that depends on the intracellular availability of SAM as methyl donor [112]. Navarro-Perak et al. [113] suggested that EGCG inhibits DHFR (dihydrofolate reductase) and/or folic acid uptake, acting as an antifolate compound. Apart from SAM, folates are an important source of the 1-carbon unit used to methylate DNA. Low folates in cancer cells induced by EGCG administration prevent DNA hypermethylation of specific genes. The effective concentration of EGCG on human DHFR inhibition is around 33 nM, a value physiologically achievable in human plasma after ingestion of standardized preparation of GTCs [114,115,116,117]. One of the most important mechanisms to control gene expression depends on the chromatin condensation status, which is regulated by different processes, with histone acetylation/deacetylation being one of the most studied and finely regulated [105]. Histone acetylation often correlates with chromatin relaxation. Inhibition of histone deacetylases (HDACs) in cancer cells causes the re-expression of genes epigenetically silenced during carcinogenesis [118]. Polyphenon E® administration has been reported to cause inhibition of HDACs activity and expression in PCa cell lines [106,119]. Molecular modeling and docking studies in HeLa cells suggested that EGCG inhibits HDAC1 activity by direct binding of the enzyme [108].
miRNAs are functional small non-coding RNAs that control gene expression by inducing degradation or translational inhibition of their target mRNAs. It has been demonstrated that various miRNAs are often overexpressed in cancer cells, including PCa cells, and their altered expression is associated with cancer progression [120]. miRNA-21 is regulated by androgens and promotes disease progression toward a hormone-independent phenotype [121]. Siddiqui et al. demonstrated that EGCG treatment of nude mice subcutaneously implanted with 22Rv1 PCa cells inhibited tumor growth by down-regulating miRNA-21 and induced the up-regulation of miRNA-330, a tumor suppressor that induces apoptosis in PCa cells [122].
3.3. Effects of GTCs on Protein Homeostasis
Tumor cells are continuously exposed to stressful conditions (such as hypoxia, nutrient deprivation, acidosis, chemotherapeutic treatments, etc.) and are characterized by a high translational activity; therefore, they are much more prone to accumulate misfolded and/or unfolded proteins. This stress is even higher in PCa secretory epithelial cells, which are characterized by a high rate of protein synthesis. For the maintenance of protein homeostasis (proteostasis), eukaryotic cells have developed a complex protein quality control system, which comprises molecular chaperons, the ER-associated degradation (ERAD) mechanism, the unfolded protein response (UPR), and the two most important systems of protein degradation, i.e. the proteasome system and the autophagic mechanism. Proteostasis impairment and accumulation of misfolded proteins in the ER lumen induce ER stress, which in turn activates the UPR, a complex network of molecular signaling to adapt and respond to ER stress conditions, thereby promoting cell survival. However, under chronic stress when proteostasis cannot be restored, the UPR activates apoptosis signaling that commits cells to death [123,124].
GTCs have been proven to trigger anticancer effects by targeting HSPs and proteasome functions and by interfering with the autophagic flux.
3.3.1. Heat Shock Proteins
Heat shock proteins (HSPs) are a superfamily of molecular chaperones representing the first line of defense against protein misfolding and aggregation. They aid in the folding and refolding of proteins, and target denatured proteins to degradative systems. HSPs function both in physiological and pathological conditions. Different cellular pathways, associated with cancer onset and progression, may be involved in the activation of the heat shock factor-1 (HSF-1), the primary factor associated with HSPs gene activation [125,126]. The expression of different members of HSPs is modulated during PCa progression [127]. It is documented that EGCG inhibits carcinogenesis thanks to the down-regulation of both HSP90 and HSP70 and through the dissociation of their complexes with co-chaperones or client proteins [128]. In different cell lines that mimic progressive stages of PCa, Moses and colleagues demonstrated that EGCG inhibited cell proliferation and induced apoptosis by direct binding to HSP90, causing instability and altering the functions of HSP90-client proteins involved in tumor progression [129].
3.3.2. Proteasome Inhibition
The ubiquitin-mediated proteasomal degradation is essential for the regulation of several cellular processes. The proteasome is a multicatalytic protein complex present in all eukaryotic cells, and it is responsible for the degradation of the major part of the cellular proteins. The chymotrypsin-like activity of the proteasome is associated with the tumor cell survival [130] since most of the intracellular proteins involved in the onset and progression of tumor are degraded through the ubiquitin-proteasome pathway. GTCs are strong and selective inhibitors of the proteasomal chymotrypsin-like activity in vitro (IC50 = 86 ÷ 194 nM) and in intact PCa cells (1 ÷ 10 µM). EGCG, among all the catechins tested, showed the strongest inhibitory activity [130]. The ester bond contained in galloylated catechins is required for potent inhibition of the proteasomal activity [130,131]. Other structure-activity relationship studies evidenced that the A ring and the gallate ester/amide bond are involved in the mechanism of proteasome inhibition [132]. Similar effects have been described for two synthetic enantiomeric analogs of natural GTCs in PCa cells [133]. The observed accumulation of short-lived proteins, normally addressed to proteosomal degradation, such as p21, p27, Bax, and ikBα in PCa cells after EGCG or GTCs treatment indirectly confirms that GTCs exert an inhibitory effect on the proteasome activity [98,134].
3.3.3. Autophagy
The catabolic process through which long-lived proteins, damaged organelles, and other unnecessary intracellular materials are delivered to lysosomes for degradation is called autophagy. Normally autophagy is constitutively active at low levels to guarantee cellular homeostasis, but it can be strongly induced in stressful conditions when this process seems to act primarily as a cellular protective response [135]. The effects of EGCG on autophagy seem to be dependent on the concentration used, cell type, and stress conditions.
We have observed at a morphological level, by electron microscopy, that GTCs administration (0.3% in tap water) in TRAMP mice drastically reduced the condensing and secretory activities of the ER of prostate cells, causing an imbalance of the protein trafficking and an engulfment of ER, finally leading to ER stress. No effects were observed in age matched animals that were water fed. In agreement with morphological data, we found that the biogenesis of secreted proteins is altered in TRAMP-C2 cells treated with 20 µg/mL of GTCs [136]. We reported that autophagy was transiently activated in human prostate PNT1a cells, commonly used to mimic the initial stages of PCa, as a survival response to overcome Polyphenon E®-induced ER stress [42]. Treatment of PNT1a with 35 µg/mL of Polyphenon E® for 24 h committed cells to apoptosis, while 145 µg/mL Polyphenon E® committed metastatic prostate cancer PC3 cells to caspase-independent programmed cell death [42].
4. GTCs in Human PCa Chemoprevention: From Epidemiological Data to Interventional Studies
Three large cohort prospective studies, all conducted in the Japanese population, aimed to examine the association between dietary intake and PCa risk [137,138,139]. All these observational studies did not find any statistically significant correlation between green tea consumption and PCa risk. A fourth study examined the association between green tea consumption and risk of PCa stratified by disease stage [140] and corrected the results for potential confounder factors. The authors did not find a significant association between green tea consumption and the global risk of being diagnosed with PCa. Nonetheless, a dose-dependent inverse relation for the risk to develop advanced PCa for men who consumed five cups of green tea/day compared with those who consumed one cup/day was observed. Two case-control studies examined the relation between green tea intake and PCa in patients diagnosed with PCa (histologically confirmed) and PCa free matched hospital inpatients as controls [141,142]. Both these studies found an inverse relation between green tea consumption and PCa risk. Nonetheless these results should be interpreted very carefully due to many critical points that include the small sample size, the hospital based case-control design, the simultaneous consumption of green and black teas, differences in tea brewing methods, and possible interferences from other dietary factors. Although epidemiological evidences are still not conclusive, the anticancer efficacy of green tea is fully displayed in the TRAMP model when intervention is started early, at weaning, and the malignancy is not fully developed. In this chemopreventive scenario, concentrations of standardized green tea extract ranging between 0.1 ÷ 0.3% in drinking water are effective to strongly delay or stop PCa growth [12,15,92]. Adhami at al. demonstrated that green tea extracts are more effective when feeding (0.1% in drinking water) is started at very early stages of PCa development (presence of “in situ” histological lesions), while the chemopreventive potential decreases in animals bearing tumors of a more advanced stage, finally resulting in largely ineffective in animals that display moderately differentiated adenocarcinoma and occasionally metastasis [143]. To reinforce the concept that GTCs are ineffective as chemotherapeutic molecules, very limited or no effects have been observed in patients with advanced PCa [144,145,146]. At difference, we introduced for the first “Proof of Concept” that secondary chemoprevention of PCa with GTCs is feasible [17]. We enrolled 60 Italian patients (all having a Caucasian genetic background) diagnosed with HG-PIN, a premalignant lesion associated with a 30% increased risk to develop PCa within 1 year [147]. 30 volunteers were randomly assigned to receive, according to a double-blind procedure, 600 mg/day of a standardized GTCs formulation (total catechins 75.7%; EGC, 5.5%; EC, 12.2%; EGCG, 51.9%; ECG, 6.12%), divided in three capsules of 200 mg each, or identical placebo capsules for one year. All the patients, during the study, were subjected to regular prostate biopsy (6 and 12 months after the study began) to assess the differences in PCa rates between the two arms. After one year of treatment, only one tumor was diagnosed among the 30 GTCs-treated men, while nine cancers were found among the 30 placebo-treated men. No significant side or adverse effects were documented throughout the whole study. A randomized, placebo-controlled trial evaluated whether 400 mg of Polyphenon E® significantly reduce PCa rates in a cohort of men diagnosed with HG-PIN and/or atypical small acinar proliferation (ASAP) [148]. 74 men (36 GTCs and 38 placebo) completed the one year protocol. No significant reduction of PCa incidence was observed between the two arms. Possible explanations of the discrepancies between the results recorded by Kumar et al. and those previously published by us may reflect differences in the study design, which include enrolment criteria (more heterogeneous) and GTCs posology (a lower dosage). Pathologists usually formulate ASAP diagnosis when it is not possible to find alterations in the cells’ morphology to support “without doubts” a PCa diagnosis. For this reason, ASAP is often considered an under-diagnosed cancer [149]. Men bearing both ASAP and multifocal HG-PIN are very likely patients in whom, at the moment of the first biopsy, PCa has already developed, even if not clearly supported by histopathological diagnosis [150]. We might hypothesize that in the study by Kumar et al. were included men for which the narrow window of time for therapeutic intervention with GTCs was already expired at the enrolment. As a consequence of the small number of patients enrolled, unfortunately, the study did not have sufficient statistical power to detect a reduction of PCa incidence in the subgroup of men diagnosed only with HG-PIN receiving GTCs in comparison with HG-PIN patients receiving the placebo group [148].
5. GTCs Bioavailability and Metabolism in Humans
Flavanols bioavailability in vivo is strongly limited by their high molecular weight and by a bulky hydration shell that magnifies the effective molecular size. Moreover, extensive enteroluminal biotransformation and phase II metabolism of flavan-3-ols, which includes methylation, glucuronidation, and O-methylation may affect GTCs bioactivity in vivo and explain inconsistent results in comparison to effects observed in vitro, using supraphysiological doses of EGCG [151]. Chow et al. investigated the pharmacokinetic of GTCs following single administration of Polyphenon E® at increasing concentrations ranging from 200 mg to 800 mg or equivalent doses of pure EGCG [116,152]. The average EGCG peak plasma concentration (Cmax) was reached after 2–4 h from ingestion and declined rapidly, with very low/undetectable levels at 24 h after dosing. EGCG peak concentration increased with increasing doses in a nonlinear fashion, reaching a value above 1 µM at the highest concentration tested, possibly due to the existence of saturable mechanisms that cause a presystemic elimination of orally administered GTCs. EGC and EC were detected in plasma and urine samples after Polyphenon E® administration, predominantly as glucuronic acid/sulfate conjugates, while EGCG was detected prevalently in its unchanged form. The presence of other catechins in the formulation tested does not influence EGCG plasma concentration, at difference the presence of food in the stomach interferes with catechins absorption and reduces threefold the plasmatic concentration of free catechins (EGCG, EGC, EC) [116,152]. The same authors observed that the systemic availability of free EGCG increases by 60% after repeated administration (four weeks of treatment) of a high daily bolus dose (800 mg once daily) and that this concentration is safe and well tolerated in healthy human subjects [116]. Similar results of catechins serum levels were obtained in patients diagnosed with PCa and receiving 800 mg of Polyphenon E® for four to six weeks before scheduled prostatectomy surgery [146]. The authors instead found a very low to undetectable level of catechins in the prostate surgical samples collected from the same patients, possibly due to a combination of rapid systemic clearance (less than 24 h) and low bioaccumulation of polyphenols in the tissue [146]. Different GTCs concentrations ranging from 21 pmol/g to 107 pmol/g tissue were found in the prostate of men who consumed 1.42 L of green tea for five days before prostatectomy [153]. In human prostate glands of PCa patients who consumed six cups of green tea daily for up to six weeks before prostatectomy, EGCG is detectable mainly in the free form (nonglucuronidated/sulfated) [154]. Approximately half of the EGCG was present in the methylated form as 4″-MeEGCG. Results obtained in LNCaP cells indicated that methylation of EGCG may occur directly in prostate tissue catalized by cytosolic COMT. None of the nongallated GTCs, including EGC and EC or their metabolites, were found in the prostate tissue, indicating a lower bioavailability for these compounds in comparison with EGCG [154].
Catechins that are not absorbed in the small intestine, as well as conjugated catechins excreted in the bile, reach the large intestine, where they may be metabolized by colonic bacteria [151]. (−)-EC, (−)-EGC, and (−)-EGCG, are degradated by the human colon microbiota to 5-(3’,4’,5’-trihydroxyphenyl)-γ-valerolactone, 5-(3’,4’-dihydroxyphenyl)-γ-valerolactone, and 5-(3’,5’-dihydroxyphenyl)-γ-valerolactone. Some of these colonic metabolites may enter the circulation and undergo further modifications before being excreted with urine in quantities corresponding to about 40% of total ingested GTCs [155]. Colonic biotransformation represents, therefore, a significant component of the overall bioavailability equation and significantly influences GTCs bioactivity. To the best of our knowledge, no data are available on the bioactivity of these compounds on PCa cells.
6. Nanotechnology Strategies to Improve GTCs Bioactivity
The great potential of use of GTCs or EGCG as safe natural molecules to prevent PCa onset or eventually revert early PCa lesions is limited by the intrinsic chemical instability, low bioavailability, and quick in vivo clearance of these products. Drug formulation strategies aimed at improving GTCs/EGCG stability assist efficient and specific delivery at target cells and limit undesired metabolism and are therefore expected to produce a valuable benefit in terms of an improved pharmacodynamic profile. The concept of nanochemoprevention, i.e. the uses of nanotechnology for enhancing the outcome of chemoprevention, was first introduced by Siddiqui et al. [156]. NPs are nanocarriers made of biodegradable/biocompatible materials sized between 1 nm and 100 nm, used as vehicles to improve drug delivery. Different types of NPs have been used to improve the pharmacological profile of EGCG, including gold nanoparticles, polymeric nanoparticles, liposomes, carbohydrates, and inorganic nanoparticles [157,158].
Siddiqui et al., in their “Proof of Principle”, study demonstrated that EGCG encapsulated in polylactic acid (PLA)–polyethylene glycol (PEG) nanoparticles has an improved efficacy against human PCa cells both in vitro and in vivo [156]. The inhibition of PC3 proliferation by half of normal values, measured as the IC50, was found to be tenfold lower when nano-EGCG was used in comparison to non encapsulated EGCG. In vivo, a tenfold lower dose of nano-EGCG was required to achieve a similar extent of xenograft tumors to that obtained with EGCG.
Polymeric EGCG-encapsulated NPs, functionalised with a mimetic ligand of the prostate specific membrane antigen (PSMA), shows an improved selective EGCG delivery in PSMA expressing PCa cells, thus leading to a more marked inhibition of cell proliferation [159,160]. The same authors observed a significant increase of tumor growth inhibition in mouse xenografts treated with PSMA targeted NPs, in comparison to EGCG. This effect may be attributed to the selective binding of NPs to PSMA, which presumably would promote an active targeting through receptor-mediated endocytosis [160]. One disadvantage of using PLA-PEG NPs is their low stability in the acidic environment that precludes the oral route of administration. Chitosan NPs encapsulating EGCG (Chit-nanoEGCG) at difference are characterized by the slow release of EGCG in simulated gastric juice acidic pH and a faster release in simulated intestinal fluid. Treatment with Chit-nanoEGCG resulted in significant growth inhibition of subcutaneously implanted 22Rν1 PCa xenografts. In tumor tissues of mice treated with Chit-nanoEGCG, compared with animals treated with EGCG, there was a significant induction of apoptosis and reduction of cell proliferation [161].
EGCG was efficiently loaded in polysaccharides NPs made of Arabic gum and maltodextrin. Encapsulated EGCG retained its biological activity; indeed it reduced cell viability and induced apoptosis when administered to DU145 PCa cells. Colony formation assay showed that nanoencapsulated EGCG possesses an enhanced inhibitory effect on cell proliferation at physiologically achievable concentrations (1 ÷ 2 µM), compared with free EGCG [162].
EGCG encapsulated in solid lipid nanoparticles (EGCG-SLN) increases molecule stability in aqueous solution, improves efficient cell delivery, and yields a fourfold increase in cytotoxicity against DU145 human PCa, in comparison to cells that received pure EGCG at the same concentration [163].
EGCG intercalated into Ca/Al-NO3 layered double hydroxide (LDH) NPs produced a higher inhibition of proliferation and colony formation suppression in PC3 cells, compared to EGCG alone. Similarly, enhanced apoptosis was observed in PC3 treated with EGCG-LDH in comparison to non-encapsulated EGCG. The enhanced antitumor activity of EGCG-loaded LDH-NPs is well correlated to the mechanism of diffusion of the EGCG through the vehicle that allowed a sustained gradual release of the drug in solution [164].
EGCG can also be used as a “Trojan horse” to deliver anticancer therapy specifically in PCa cells. Recently, it has been demonstrated that EGCG functionalized radioactive biocompatible gold NPs, derived from the Au-198 isotope (EGCG-198AuNPs), selectively bind with high affinity to the 67-LR, a receptor that is overexpressed in PCa cells. Significant amounts of EGCG-198AuNPs were internalized through 67-LR receptor endocytosis in tumor cells. Pharmacokinetic studies in PC-3 xenograft SCID mice showed a high retention of EGCG-198AuNPs in tumors and an 80% reduction of tumor volumes, compared to controls [80]. EGCG and gelatin-doxorubicin conjugate (GLT-DOX)-coated gold NPs (GLT-DOX/EGCG AuNPs) were produced to improve the specific uptake of doxorubicin in PC3 cells using EGCG/67-LR receptor-mediated delivery. Apoptosis of GLT-DOX/EGCG AuNPs treated PC3 was improved in comparison with cells treated with DOX [165].
7. Conclusions
Many authors have suggested that the anticancer effects of GTCs might be achieved through their direct ability to scavenge harmful ROS/RNS, or alternatively, through their autoxidation, which, in turn, produces H2O2, a toxic compound. Both these effects, observed in vitro, have been obtained using a supraphysiological concentration of GTCs and are very unlikely to take place in vivo. Recent achievements in this field of research have clarified that GTCs can interact with molecular targets that are specifically deregulated in PCa, such as the 67-LR receptor, at nanomolar concentrations. Others have demonstrated that GTCs modify the expressions of genes relevant for PCa progression, acting on transcription factors and through epigenetic mechanisms, or activating complex adaptive signaling such as the UPR. Despite the fact that GTCs have been proven to effectively inhibit PCa progression in animal models, their chemopreventive action in human PCa is still debated. The lessons learnt from clinical trials so far indicate that the therapeutic window for GTCs intervention is very narrow and possibly corresponds to early signs of prostate tissue transformation such as HG-PIN. Moreover inadequate dosage and/or short intervention periods may compromise the therapeutic effect. It is now clear that individual differences in GTCs metabolism and absorption may negatively affect the already low bioavailability, consequently reducing the bioactivity. New opportunities to improve the therapeutic value of GTCs come from the field of the nanotechnologies, which, when applied to chemoprevention, lead to the development of nanochemoprevention strategies. More than encouraging results have shown that biopolymeric NPs help to selectively deliver GTCs in PCa cancer cells, improving drug stability and increasing in vivo bioactivity. Large, well-designed, interventional studies are urgently needed to evaluate the efficacy of GTCs charged NPs in PCa prevention.
Acknowledgments
Valeria Naponelli was supported by a Fondazione Umberto Veronesi fellowship.
Author Contributions
Valeria Naponelli, Ileana Ramazzina, Chiara Lenzi, Saverio Bettuzzi and Federica Rizzi critically reviewed the literature and wrote the paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ferlay, J.; Soerjomataram, I.; Ervik, M.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Globocan 2012 v1.0, Cancer Incidence and Mortality Worldwide: Iarc Cancerbase No. 11. Available online: http://globocan.iarc.fr (accessed on 1 February 2017).
- Wong, M.C.; Goggins, W.B.; Wang, H.H.; Fung, F.D.; Leung, C.; Wong, S.Y.; Ng, C.F.; Sung, J.J. Global incidence and mortality for prostate cancer: Analysis of temporal patterns and trends in 36 countries. Eur. Urol. 2016, 70, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-L.; Wang, S.-C.; Ho, C.-J.; Kao, Y.-L.; Hsieh, T.-Y.; Chen, W.-J.; Chen, C.-J.; Wu, P.-R.; Ko, J.-L.; Lee, H.; et al. Prostate cancer mortality-to-incidence ratios are associated with cancer care disparities in 35 countries. Sci. Rep. 2017, 7, 40003. [Google Scholar] [CrossRef] [PubMed]
- Baade, P.D.; Youlden, D.R.; Krnjacki, L.J. International epidemiology of prostate cancer: Geographical distribution and secular trends. Mol. Nutr. Food Res. 2009, 53, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Ross, R.K.; Bernstein, L.; Yatani, R.; Henderson, B.E.; Mack, T.M. Cancers of the prostate and breast among japanese and white immigrants in los angeles county. Br. J. Cancer 1991, 63, 963–966. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Demissie, K.; Lu, S.E.; Rhoads, G.G. Cancer incidence among korean-american immigrants in the united states and native koreans in south korea. Cancer Control 2007, 14, 78–85. [Google Scholar] [PubMed]
- Bettuzzi, S.; Rizzi, F.; Belloni, L. Clinical relevance of the inhibitory effect of green tea catechins (GtCs) on prostate cancer progression in combination with molecular profiling of catechin-resistant tumors: An integrated view. Pol. J. Vet. Sci. 2007, 10, 57–60. [Google Scholar] [PubMed]
- Johnson, J.J.; Bailey, H.H.; Mukhtar, H. Green tea polyphenols for prostate cancer chemoprevention: A translational perspective. Phytomedicine 2010, 17, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Sawada, N. Risk and preventive factors for prostate cancer in Japan: The Japan public health center-based prospective (JPHC) study. J. Epidemiol. 2017, 27, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.M.; Wang, P.; Heber, D. Chemopreventive effects of tea in prostate cancer: Green tea versus black tea. Mol. Nutr. Food Res. 2011, 55, 905–920. [Google Scholar] [CrossRef] [PubMed]
- Takehiko, Y.; Mujo, K. Chemistry and Applications of Green Tea; CRC Press: New York, NY, USA, 1997; p. 1e130. [Google Scholar]
- Caporali, A.; Davalli, P.; Astancolle, S.; D’Arca, D.; Brausi, M.; Bettuzzi, S.; Corti, A. The chemopreventive action of catechins in the TRAMP mouse model of prostate carcinogenesis is accompanied by clusterin over-expression. Carcinogenesis 2004, 25, 2217–2224. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Feyes, D.K.; Nieminen, A.L.; Agarwal, R.; Mukhtar, H. Green tea constituent epigallocatechin-3-gallate and induction of apoptosis and cell cycle arrest in human carcinoma cells. J. Natl. Cancer Inst. 1997, 89, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, D.S.; Clubbs, E.A.; Ferruzzi, M.; Bomser, J.A. Epigallocatechin-3-gallate (EGCG) inhibits PC-3 prostate cancer cell proliferation via MEK-independent ERK1/2 activation. Chem. Biol. Interact. 2008, 171, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Hastak, K.; Ahmad, N.; Lewin, J.S.; Mukhtar, H. Inhibition of prostate carcinogenesis in TRAMP mice by oral infusion of green tea polyphenols. Proc. Natl. Acad. Sci. USA 2001, 98, 10350–10355. [Google Scholar] [CrossRef] [PubMed]
- Adhami, V.M.; Siddiqui, I.A.; Ahmad, N.; Gupta, S.; Mukhtar, H. Oral consumption of green tea polyphenols inhibits insulin-like growth factor-i-induced signaling in an autochthonous mouse model of prostate cancer. Cancer Res. 2004, 64, 8715–8722. [Google Scholar] [CrossRef] [PubMed]
- Bettuzzi, S.; Brausi, M.; Rizzi, F.; Castagnetti, G.; Peracchia, G.; Corti, A. Chemoprevention of human prostate cancer by oral administration of green tea catechins in volunteers with high-grade prostate intraepithelial neoplasia: A preliminary report from a one-year proof-of-principle study. Cancer Res. 2006, 66, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Brausi, M.; Rizzi, F.; Bettuzzi, S. Chemoprevention of human prostate cancer by green tea catechins: Two years later. A follow-up update. Eur. Urol. 2008, 54, 472–473. [Google Scholar] [CrossRef] [PubMed]
- Davalli, P.; Rizzi, F.; Caporali, A.; Pellacani, D.; Davoli, S.; Bettuzzi, S.; Brausi, M.; D’Arca, D. Anticancer activity of green tea polyphenols in prostate gland. Oxid. Med. Cell. Longev. 2012, 2012, 984219. [Google Scholar] [CrossRef] [PubMed]
- Sena, L.A.; Chandel, N.S. Physiological roles of mitochondrial reactive oxygen species. Mol. Cell 2012, 48, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Elias, R.J. The antioxidant and pro-oxidant activities of green tea polyphenols: A role in cancer prevention. Arch. Biochem. Biophys. 2010, 501, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Ludovico, P.; Burhans, W.C. Reactive oxygen species, ageing and the hormesis police. FEMS Yeast Res. 2014, 14, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Oh, B.; Figtree, G.; Costa, D.; Eade, T.; Hruby, G.; Lim, S.; Elfiky, A.; Martine, N.; Rosenthal, D.; Clarke, S.; et al. Oxidative stress in prostate cancer patients: A systematic review of case control studies. Prostate Int. 2016, 4, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Khandrika, L.; Kumar, B.; Koul, S.; Maroni, P.; Koul, H.K. Oxidative stress in prostate cancer. Cancer Lett. 2009, 282, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Thapa, D.; Ghosh, R. Antioxidants for prostate cancer chemoprevention: Challenges and opportunities. Biochem. Pharmacol. 2012, 83, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Shankar, S.; Srivastava, R.K. Green tea catechin, epigallocatechin-3-gallate (EGCG): Mechanisms, perspectives and clinical applications. Biochem. Pharmacol. 2011, 82, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Forester, S.C.; Lambert, J.D. The role of antioxidant versus pro-oxidant effects of green tea polyphenols in cancer prevention. Mol. Nutr. Food. Res. 2011, 55, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Babich, H.; Schuck, A.G.; Weisburg, J.H.; Zuckerbraun, H.L. Research strategies in the study of the pro-oxidant nature of polyphenol nutraceuticals. J. Toxicol. 2011, 2011, 467305. [Google Scholar] [CrossRef] [PubMed]
- Elbling, L.; Weiss, R.M.; Teufelhofer, O.; Uhl, M.; Knasmueller, S.; Schulte-Hermann, R.; Berger, W.; Micksche, M. Green tea extract and (−)-epigallocatechin-3-gallate, the major tea catechin, exert oxidant but lack antioxidant activities. FASEB J. 2005, 19, 807–809. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, M.; Sonda, T.; Nagayama, K.; Tabata, M. Effects of PH and metal ions on antioxidative activities of catechins. Biosci. Biotechnol. Biochem. 2001, 65, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Nanjo, F.; Mori, M.; Goto, K.; Hara, Y. Radical scavenging activity of tea catechins and their related compounds. Biosci. Biotechnol. Biochem. 1999, 63, 1621–1623. [Google Scholar] [CrossRef] [PubMed]
- Long, L.H.; Clement, M.V.; Halliwell, B. Artifacts in cell culture: Rapid generation of hydrogen peroxide on addition of (−)-epigallocatechin, (−)-epigallocatechin gallate, (+)-catechin, and quercetin to commonly used cell culture media. Biochem. Biophys. Res. Commun. 2000, 273, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Long, L.H.; Kirkland, D.; Whitwell, J.; Halliwell, B. Different cytotoxic and clastogenic effects of epigallocatechin gallate in various cell-culture media due to variable rates of its oxidation in the culture medium. Mutat. Res. 2007, 634, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Twum-Ampofo, J.; Fu, D.X.; Passaniti, A.; Hussain, A.; Siddiqui, M.M. Metabolic targets for potential prostate cancer therapeutics. Curr. Opin. Oncol. 2016, 28, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.L.; He, G.Q.; Yu, H.N.; Yang, J.G.; Borthakur, D.; Zhang, L.C.; Shen, S.R.; Das, U.N. Free Zn2+ enhances inhibitory effects of EGCG on the growth of PC-3 cells. Mol. Nutr. Food Res. 2008, 52, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, H.; Sun, S.; Zhang, L.; Das, U.N.; Ruan, H.; He, G.; Shen, S. Mechanism of free Zn2+ enhancing inhibitory effects of EGCG on the growth of PC-3 cells: Interactions with mitochondria. Biol. Trace. Elem. Res. 2009, 131, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.G.; Yu, H.N.; Sun, S.L.; Zhang, L.C.; He, G.Q.; Das, U.N.; Ruan, H.; Shen, S.R. Epigallocatechin-3-gallate affects the growth of LNCaP cells via membrane fluidity and distribution of cellular zinc. J. Zhejiang Univ. Sci. B 2009, 10, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Stearns, M.E.; Amatangelo, M.D.; Varma, D.; Sell, C.; Goodyear, S.M. Combination therapy with epigallocatechin-3-gallate and doxorubicin in human prostate tumor modeling studies: Inhibition of metastatic tumor growth in severe combined immunodeficiency mice. Am. J. Pathol. 2010, 177, 3169–3179. [Google Scholar] [CrossRef] [PubMed]
- Chung, L.Y.; Cheung, T.C.; Kong, S.K.; Fung, K.P.; Choy, Y.M.; Chan, Z.Y.; Kwok, T.T. Induction of apoptosis by green tea catechins in human prostate cancer DU145 cells. Life Sci. 2001, 68, 1207–1214. [Google Scholar] [CrossRef]
- Kanwal, R.; Pandey, M.; Bhaskaran, N.; Maclennan, G.T.; Fu, P.; Ponsky, L.E.; Gupta, S. Protection against oxidative DNA damage and stress in human prostate by glutathione S-transferase P1. Mol. Carcinog. 2014, 53, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, F.; Naponelli, V.; Silva, A.; Modernelli, A.; Ramazzina, I.; Bonacini, M.; Tardito, S.; Gatti, R.; Uggeri, J.; Bettuzzi, S. Polyphenon e®, a standardized green tea extract, induces endoplasmic reticulum stress, leading to death of immortalized PNT1a cells by anoikis and tumorigenic PC3 by necroptosis. Carcinogenesis 2014, 35, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Candas, D.; Li, J.J. Mnsod in oxidative stress response-potential regulation via mitochondrial protein influx. Antioxid. Redox Signal 2014, 20, 1599–1617. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Holly, J.M.; Persad, R.; Bahl, A.; Perks, C.M. Green tea extract (epigallocatechin-3-gallate) reduces efficacy of radiotherapy on prostate cancer cells. Urology 2011, 78, 475.e15–475.e21. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.; Sheridan, J.; Mulcahy, H.; Tenniswood, M.; Morrissey, C. The effect of green tea on oxidative damage and tumour formation in lobund-wistar rats. Eur. J. Cancer Prev. 2008, 17, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Nyska, A.; Suttie, A.; Bakshi, S.; Lomnitski, L.; Grossman, S.; Bergman, M.; Ben-Shaul, V.; Crocket, P.; Haseman, J.K.; Moser, G.; et al. Slowing tumorigenic progression in TRAMP mice and prostatic carcinoma cell lines using natural anti-oxidant from spinach, NAO—A comparative study of three anti-oxidants. Toxicol. Pathol. 2003, 31, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Tam, N.N.; Nyska, A.; Maronpot, R.R.; Kissling, G.; Lomnitski, L.; Suttie, A.; Bakshi, S.; Bergman, M.; Grossman, S.; Ho, S.M. Differential attenuation of oxidative/nitrosative injuries in early prostatic neoplastic lesions in TRAMP mice by dietary antioxidants. Prostate 2006, 66, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Vahora, H.; Khan, M.A.; Alalami, U.; Hussain, A. The potential role of nitric oxide in halting cancer progression through chemoprevention. J. Cancer Prev. 2016, 21, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J.; Chun, K.S.; Cha, H.H.; Han, S.S.; Keum, Y.S.; Park, K.K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-kappa B activation. Mutat. Res. 2001, 480–481, 243–268. [Google Scholar] [CrossRef]
- Hayakawa, S.; Saito, K.; Miyoshi, N.; Ohishi, T.; Oishi, Y.; Miyoshi, M.; Nakamura, Y. Anti-cancer effects of green tea by either anti- or pro- oxidative mechanisms. Asian Pac. J. Cancer Prev. 2016, 17, 1649–1654. [Google Scholar] [CrossRef] [PubMed]
- Dhakshinamoorthy, S.; Porter, A.G. Nitric oxide-induced transcriptional up-regulation of protective genes by Nrf2 via the antioxidant response element counteracts apoptosis of neuroblastoma cells. J. Biol. Chem. 2004, 279, 20096–20107. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.E.; Patel, B.B.; Wang, J.; Eltoum, I.A.; Lamartiniere, C.A. Epigallocatechin-3-gallate suppresses early stage, but not late stage prostate cancer in tramp mice: Mechanisms of action. Prostate 2007, 67, 1576–1589. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M. Cellular targets for the beneficial actions of tea polyphenols. Am. J. Clin. Nutr. 2013, 98, 1642S–1650S. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Lu, H.; Meng, X.; Ryu, J.H.; Hara, Y.; Yang, C.S. Stability, cellular uptake, biotransformation, and efflux of tea polyphenol (−)-epigallocatechin-3-gallate in ht-29 human colon adenocarcinoma cells. Cancer Res. 2002, 62, 7241–7246. [Google Scholar] [PubMed]
- Shimizu, M.; Adachi, S.; Masuda, M.; Kozawa, O.; Moriwaki, H. Cancer chemoprevention with green tea catechins by targeting receptor tyrosine kinases. Mol. Nutr. Food Res. 2011, 55, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Kitano, K.; Nam, K.Y.; Kimura, S.; Fujiki, H.; Imanishi, Y. Sealing effects of (−)-epigallocatechin gallate on protein kinase C and protein phosphatase 2A. Biophys. Chem. 1997, 65, 157–164. [Google Scholar] [CrossRef]
- Kondo, T.; Ohta, T.; Igura, K.; Hara, Y.; Kaji, K. Tea catechins inhibit angiogenesis in vitro, measured by human endothelial cell growth, migration and tube formation, through inhibition of vegf receptor binding. Cancer Lett. 2002, 180, 139–144. [Google Scholar] [CrossRef]
- Liang, Y.C.; Lin-shiau, S.Y.; Chen, C.F.; Lin, J.K. Suppression of extracellular signals and cell proliferation through EGF receptor binding by (−)-epigallocatechin gallate in human A431 epidermoid carcinoma cells. J. Cell. Biochem. 1997, 67, 55–65. [Google Scholar] [CrossRef]
- Li, M.; He, Z.; Ermakova, S.; Zheng, D.; Tang, F.; Cho, Y.Y.; Zhu, F.; Ma, W.Y.; Sham, Y.; Rogozin, E.A.; et al. Direct inhibition of insulin-like growth factor-I receptor kinase activity by (−)-epigallocatechin-3-gallate regulates cell transformation. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Nagao, T.; Ingolfsson, H.I.; Maxfield, F.R.; Andersen, O.S.; Kopelovich, L.; Weinstein, I.B. The inhibitory effect of (-)-epigallocatechin gallate on activation of the epidermal growth factor receptor is associated with altered lipid order in ht29 colon cancer cells. Cancer Res. 2007, 67, 6493–6501. [Google Scholar] [CrossRef] [PubMed]
- Duhon, D.; Bigelow, R.L.; Coleman, D.T.; Steffan, J.J.; Yu, C.; Langston, W.; Kevil, C.G.; Cardelli, J.A. The polyphenol epigallocatechin-3-gallate affects lipid rafts to block activation of the c-Met receptor in prostate cancer cells. Mol. Carcinog. 2010, 49, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Nagao, T.; To, S.; Joe, A.K.; Shimizu, M.; Matsushima-Nishiwaki, R.; Kozawa, O.; Moriwaki, H.; Maxfield, F.R.; Weinstein, I.B. (−)-epigallocatechin gallate causes internalization of the epidermal growth factor receptor in human colon cancer cells. Carcinogenesis 2008, 29, 1986–1993. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Afaq, F.; Saleem, M.; Ahmad, N.; Mukhtar, H. Targeting multiple signaling pathways by green tea polyphenol (-)-epigallocatechin-3-gallate. Cancer Res. 2006, 66, 2500–2505. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Weinstein, I.B. Modulation of signal transduction by tea catechins and related phytochemicals. Mutat. Res. 2005, 591, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Regad, T. Targeting RTK signaling pathways in cancer. Cancers (Basel) 2015, 7, 1758–1784. [Google Scholar] [CrossRef] [PubMed]
- Larsen, C.A.; Dashwood, R.H.; Bisson, W.H. Tea catechins as inhibitors of receptor tyrosine kinases: Mechanistic insights and human relevance. Pharmacol. Res. 2010, 62, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Rohan, T. Role of the insulin-like growth factor family in cancer development and progression. J. Natl. Cancer Inst. 2000, 92, 1472–1489. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, S.A.; Atreya, H.S. Defining the pathway to insulin-like growth factor system targeting in cancer. Biochem. Pharmacol. 2010, 80, 1115–1124. [Google Scholar] [CrossRef] [PubMed]
- Pianetti, S.; Guo, S.; Kavanagh, K.T.; Sonenshein, G.E. Green tea polyphenol epigallocatechin-3 gallate inhibits HER-2/neu signaling, proliferation, and transformed phenotype of breast cancer cells. Cancer Res. 2002, 62, 652–655. [Google Scholar] [PubMed]
- Masuda, M.; Suzui, M.; Lim, J.T.; Weinstein, I.B. Epigallocatechin-3-gallate inhibits activation of HER-2/neu and downstream signaling pathways in human head and neck and breast carcinoma cells. Clin. Cancer Res. 2003, 9, 3486–3491. [Google Scholar] [PubMed]
- Shimizu, M.; Deguchi, A.; Lim, J.T.E.; Moriwaki, H.; Kopelovich, L.; Weinstein, I.B. (−)-epigallocatechin gallate and polyphenon E inhibit growth and activation of the epidermal growth factor receptor and human epidermal growth factor receptor-2 signaling pathways in human colon cancer cells. Clin. Cancer Res. 2005, 11, 2735–2746. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Deguchi, A.; Joe, A.K.; McKoy, J.F.; Moriwaki, H.; Weinstein, I.B. EGCG inhibits activation of HER3 and expression of cyclooxygenase-2 in human colon cancer cells. J. Exp. Ther. Oncol. 2005, 5, 69–78. [Google Scholar] [PubMed]
- Shimizu, M.; Shirakami, Y.; Sakai, H.; Tatebe, H.; Nakagawa, T.; Hara, Y.; Weinstein, I.B.; Moriwaki, H. EGCG inhibits activation of the insulin-like growth factor (igf)/igf-1 receptor axis in human hepatocellular carcinoma cells. Cancer Lett. 2008, 262, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Sakata, R.; Ueno, T.; Nakamura, T.; Sakamoto, M.; Torimura, T.; Sata, M. Green tea polyphenol epigallocatechin-3-gallate inhibits platelet-derived growth factor-induced proliferation of human hepatic stellate cell line li90. J. Hepatol. 2004, 40, 52–59. [Google Scholar] [CrossRef]
- Larsen, C.A.; Dashwood, R.H. Suppression of Met activation in human colon cancer cells treated with (−)-epigallocatechin-3-gallate: Minor role of hydrogen peroxide. Biochem. Biophys. Res. Commun. 2009, 389, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Larsen, C.A.; Bisson, W.H.; Dashwood, R.H. Tea catechins inhibit hepatocyte growth factor receptor (Met kinase) activity in human colon cancer cells: Kinetic and molecular docking studies. J. Med. Chem. 2009, 52, 6543–6545. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Koga, K.; Fujimura, Y.; Yamada, K. A receptor for green tea polyphenol EGCG. Nat. Struct. Mol. Biol. 2004, 11, 380–381. [Google Scholar] [CrossRef] [PubMed]
- Umeda, D.; Yano, S.; Yamada, K.; Tachibana, H. Green tea polyphenol epigallocatechin-3-gallate signaling pathway through 67-kDa laminin receptor. J. Biol. Chem. 2008, 283, 3050–3058. [Google Scholar] [CrossRef] [PubMed]
- Shukla, R.; Chanda, N.; Zambre, A.; Upendran, A.; Katti, K.; Kulkarni, R.R.; Nune, S.K.; Casteel, S.W.; Smith, C.J.; Vimal, J.; et al. Laminin receptor specific therapeutic gold nanoparticles (198AuNP-EGCG) show efficacy in treating prostate cancer. Proc. Natl. Acad. Sci. USA 2012, 109, 12426–12431. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Sumida, M.; Kumazoe, M.; Sugihara, K.; Suemasu, Y.; Yamada, S.; Yamashita, S.; Miyakawa, J.; Takahashi, T.; Tanaka, H.; et al. Oligomer formation of a tea polyphenol, EGCG, on its sensing molecule 67 kDa laminin receptor. Chem. Commun. 2017, 53, 1941–1944. [Google Scholar] [CrossRef] [PubMed]
- Antopolsky, M.; Fujimura, Y.; Sumida, M.; Sugihara, K.; Tsukamoto, S.; Yamada, K.; Tachibana, H. Green tea polyphenol EGCG sensing motif on the 67-kDa laminin receptor. PLoS ONE 2012, 7, e37942. [Google Scholar]
- Mocanu, M.M.; Ganea, C.; Georgescu, L.; Varadi, T.; Shrestha, D.; Baran, I.; Katona, E.; Nagy, P.; Szollosi, J. Epigallocatechin 3-O-gallate induces 67 kDa laminin receptor-mediated cell death accompanied by downregulation of ErbB proteins and altered lipid raft clustering in mammary and epidermoid carcinoma cells. J. Nat. Prod. 2014, 77, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Hirotsu, K.; Kumazoe, M.; Goto, Y.; Sugihara, K.; Suda, T.; Tsurudome, Y.; Suzuki, T.; Yamashita, S.; Kim, Y.; et al. Green tea polyphenol EGCG induces lipid-raft clustering and apoptotic cell death by activating protein kinase Cdelta and acid sphingomyelinase through a 67 kDa laminin receptor in multiple myeloma cells. Biochem. J. 2012, 443, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, Y.; Umeda, D.; Kiyohara, Y.; Sunada, Y.; Yamada, K.; Tachibana, H. The involvement of the 67 kda laminin receptor-mediated modulation of cytoskeleton in the degranulation inhibition induced by epigallocatechin-3-O-gallate. Biochem. Biophys. Res. Commun. 2006, 348, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, Y.; Yamada, K.; Tachibana, H. A lipid raft-associated 67kDa laminin receptor mediates suppressive effect of epigallocatechin-3-O-gallate on fcepsilonri expression. Biochem. Biophys. Res. Commun. 2005, 336, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, H.; Fujimura, Y.; Yamada, K. Tea polyphenol epigallocatechin-3-gallate associates with plasma membrane lipid rafts: Lipid rafts mediate anti-allergic action of the catechin. Biofactors 2004, 21, 383–385. [Google Scholar] [CrossRef] [PubMed]
- Patra, S.K. Dissecting lipid raft facilitated cell signaling pathways in cancer. Biochim. Biophys. Acta 2008, 1785, 182–206. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J.; Han, X.; Gross, R.W. Epidermal growth factor receptors are localized to lipid rafts that contain a balance of inner and outer leaflet lipids: A shotgun lipidomics study. J. Biol. Chem. 2005, 280, 26796–26804. [Google Scholar] [CrossRef] [PubMed]
- Remacle-Bonnet, M.; Garrouste, F.; Baillat, G.; Andre, F.; Marvaldi, J.; Pommier, G. Membrane rafts segregate pro- from anti-apoptotic insulin-like growth factor-I receptor signaling in colon carcinoma cells stimulated by members of the tumor necrosis factor superfamily. Am. J. Pathol. 2005, 167, 761–773. [Google Scholar] [CrossRef]
- Oshikawa, J.; Urao, N.; Kim, H.W.; Kaplan, N.; Razvi, M.; McKinney, R.; Poole, L.B.; Fukai, T.; Ushio-Fukai, M. Extracellular SOD-derived H2O2 promotes VEGF signaling in caveolae/lipid rafts and post-ischemic angiogenesis in mice. PLoS ONE 2010, 5, e10189. [Google Scholar] [CrossRef] [PubMed]
- Scaltriti, M.; Belloni, L.; Caporali, A.; Davalli, P.; Remondini, D.; Rizzi, F.; Astancolle, S.; Corti, A.; Bettuzzi, S. Molecular classification of green tea catechin-sensitive and green tea catechin-resistant prostate cancer in the TRAMP mice model by quantitative real-time PCR gene profiling. Carcinogenesis 2006, 27, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Hun Lee, J.; Shu, L.; Fuentes, F.; Su, Z.Y.; Tony Kong, A.N. Cancer chemoprevention by traditional Chinese herbal medicine and dietary phytochemicals: Targeting Nrf2-mediated oxidative stress/anti-inflammatory responses, epigenetics, and cancer stem cells. J. Tradit. Complement. Med. 2013, 3, 69–79. [Google Scholar] [PubMed]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2: INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kong, A.N. Molecular mechanisms of Nrf2-mediated antioxidant response. Mol. Carcinog. 2009, 48, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, M.K.; Lee, J.H.; Chai, E.Z.; Kanchi, M.M.; Kar, S.; Arfuso, F.; Dharmarajan, A.; Kumar, A.P.; Ramar, P.S.; Looi, C.Y.; et al. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin. Cancer Biol. 2016, 40–41, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Connors, S.K.; Chornokur, G.; Kumar, N.B. New insights into the mechanisms of green tea catechins in the chemoprevention of prostate cancer. Nutr. Cancer 2012, 64, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Vayalil, P.K.; Katiyar, S.K. Treatment of epigallocatechin-3-gallate inhibits matrix metalloproteinases-2 and -9 via inhibition of activation of mitogen-activated protein kinases, c-jun and NF-kappaB in human prostate carcinoma DU-145 cells. Prostate 2004, 59, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Hastak, K.; Gupta, S.; Ahmad, N.; Agarwal, M.K.; Agarwal, M.L.; Mukhtar, H. Role of p53 and NF-kappaB in epigallocatechin-3-gallate-induced apoptosis of LNCaP cells. Oncogene 2003, 22, 4851–4859. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.A.; Shukla, Y.; Adhami, V.M.; Sarfaraz, S.; Asim, M.; Hafeez, B.B.; Mukhtar, H. Suppression of NFkappaB and its regulated gene products by oral administration of green tea polyphenols in an autochthonous mouse prostate cancer model. Pharm. Res. 2008, 25, 2135–2142. [Google Scholar] [CrossRef] [PubMed]
- Sartor, L.; Pezzato, E.; Dona, M.; Dell’Aica, I.; Calabrese, F.; Morini, M.; Albini, A.; Garbisa, S. Prostate carcinoma and green tea: (−)epigallocatechin-3-gallate inhibits inflammation-triggered MMP-2 activation and invasion in murine TRAMP model. Int. J. Cancer 2004, 112, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Nandy, S.K.; Chowdhury, A.; Chakraborti, T.; Chakraborti, S. Inhibition of MMP-9 by green tea catechins and prediction of their interaction by molecular docking analysis. Biomedi. Pharmacother. 2016, 84, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.M.; Wang, P.; Carpenter, C.L.; Heber, D. Epigenetic effects of green tea polyphenols in cancer. Epigenomics 2013, 5, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.; Shukla, S.; Gupta, S. Promoter demethylation and chromatin remodeling by green tea polyphenols leads to re-expression of GSTP1 in human prostate cancer cells. Int. J. Cancer 2010, 126, 2520–2533. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.Z.; Wang, Y.; Ai, N.; Hou, Z.; Sun, Y.; Lu, H.; Welsh, W.; Yang, C.S. Tea polyphenol (−)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer Res. 2003, 63, 7563–7570. [Google Scholar] [PubMed]
- Khan, M.A.; Hussain, A.; Sundaram, M.K.; Alalami, U.; Gunasekera, D.; Ramesh, L.; Hamza, A.; Quraishi, U. (−)-epigallocatechin-3-gallate reverses the expression of various tumor-suppressor genes by inhibiting DNA methyltransferases and histone deacetylases in human cervical cancer cells. Oncol. Rep. 2015, 33, 1976–1984. [Google Scholar] [CrossRef] [PubMed]
- Chuang, J.C.; Yoo, C.B.; Kwan, J.M.; Li, T.W.; Liang, G.; Yang, A.S.; Jones, P.A. Comparison of biological effects of non-nucleoside DNA methylation inhibitors versus 5-aza-2′-deoxycytidine. Mol. Cancer Ther. 2005, 4, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Morey Kinney, S.R.; Zhang, W.; Pascual, M.; Greally, J.M.; Gillard, B.M.; Karasik, E.; Foster, B.A.; Karpf, A.R. Lack of evidence for green tea polyphenols as DNA methylation inhibitors in murine prostate. Cancer Prev. Res. 2009, 2, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.M.; Wang, P.; Said, J.; Magyar, C.; Castor, B.; Doan, N.; Tosity, C.; Moro, A.; Gao, K.; Li, L.; et al. Polyphenols in brewed green tea inhibit prostate tumor xenograft growth by localizing to the tumor and decreasing oxidative stress and angiogenesis. J. Nutr. Biochem. 2012, 23, 1537–1542. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Shim, J.Y.; Zhu, B.T. Mechanisms for the inhibition of DNA methyltransferases by tea catechins and bioflavonoids. Mol. Pharmacol. 2005, 68, 1018–1030. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Peran, E.; Cabezas-Herrera, J.; Campo, L.S.; Rodriguez-Lopez, J.N. Effects of folate cycle disruption by the green tea polyphenol epigallocatechin-3-gallate. Int. J. Biochem. Cell Biol. 2007, 39, 2215–2225. [Google Scholar] [CrossRef] [PubMed]
- Schantz, M.; Erk, T.; Richling, E. Metabolism of green tea catechins by the human small intestine. Biotechnol. J. 2010, 5, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Chen, L.; Lee, M.J.; Balentine, D.; Kuo, M.C.; Schantz, S.P. Blood and urine levels of tea catechins after ingestion of different amounts of green tea by human volunteers. Cancer Epidemiol. Biomarkers Prev. 1998, 7, 351–354. [Google Scholar] [PubMed]
- Chow, H.H.; Cai, Y.; Hakim, I.A.; Crowell, J.A.; Shahi, F.; Brooks, C.A.; Dorr, R.T.; Hara, Y.; Alberts, D.S. Pharmacokinetics and safety of green tea polyphenols after multiple-dose administration of epigallocatechin gallate and polyphenon E in healthy individuals. Clin. Cancer Res. 2003, 9, 3312–3319. [Google Scholar] [PubMed]
- Chow, H.H.; Hakim, I.A.; Vining, D.R.; Crowell, J.A.; Ranger-Moore, J.; Chew, W.M.; Celaya, C.A.; Rodney, S.R.; Hara, Y.; Alberts, D.S. Effects of dosing condition on the oral bioavailability of green tea catechins after single-dose administration of polyphenon E in healthy individuals. Clin. Cancer Res. 2005, 11, 4627–4633. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shang, Y.-P.; Chen, H.-Y.; Li, J. Histone deacetylases function as novel potential therapeutic targets for cancer. Hepatol. Res. 2017, 47, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.S.; Gupta, K.; Gupta, S. Green tea polyphenols increase p53 transcriptional activity and acetylation by suppressing class i histone deacetylases. Int. J. Oncol. 2012, 41, 353–361. [Google Scholar] [PubMed]
- Rivera-Barahona, A.; Pérez, B.; Richard, E.; Desviat, L.R. Role of mirnas in human disease and inborn errors of metabolism. J. Inherit. Metab. Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ribas, J.; Ni, X.; Haffner, M.; Wentzel, E.A.; Salmasi, A.H.; Chowdhury, W.H.; Kudrolli, T.A.; Yegnasubramanian, S.; Luo, J.; Rodriguez, R.; et al. miR-21: An androgen receptor-regulated microRNA that promotes hormone-dependent and hormone-independent prostate cancer growth. Cancer Res. 2009, 69, 7165–7169. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.A.; Asim, M.; Hafeez, B.B.; Adhami, V.M.; Tarapore, R.S.; Mukhtar, H. Green tea polyphenol EGCG blunts androgen receptor function in prostate cancer. FASEB J. 2010, 25, 1198–1207. [Google Scholar] [CrossRef] [PubMed]
- Schonthal, A.H. Targeting endoplasmic reticulum stress for cancer therapy. Front. Biosci. (Schol. Ed.) 2012, 4, 412–431. [Google Scholar] [CrossRef] [PubMed]
- Bruning, A.; Juckstock, J. Misfolded proteins: From little villains to little helpers in the fight against cancer. Front. Oncol. 2015, 5, 47. [Google Scholar] [PubMed]
- Ciocca, D.R.; Arrigo, A.P.; Calderwood, S.K. Heat shock proteins and heat shock factor 1 in carcinogenesis and tumor development: An update. Arch. Toxicol. 2013, 87, 19–48. [Google Scholar] [CrossRef] [PubMed]
- Niforou, K.; Cheimonidou, C.; Trougakos, I.P. Molecular chaperones and proteostasis regulation during redox imbalance. Redox Biol. 2014, 2, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Ciocca, D.R.; Fanelli, M.A.; Cuello-Carrion, F.D.; Castro, G.N. Heat shock proteins in prostate cancer: From tumorigenesis to the clinic. Int. J. Hyperth. 2010, 26, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, D.; Xu, J.; Shi, J.; Jiang, L.; Yao, N.; Ye, W. Discovery and development of natural heat shock protein 90 inhibitors in cancer treatment. Acta Pharm. Sin. B 2012, 2, 238–245. [Google Scholar] [CrossRef]
- Moses, M.A.; Henry, E.C.; Ricke, W.A.; Gasiewicz, T.A. The heat shock protein 90 inhibitor, (−)-epigallocatechin gallate, has anticancer activity in a novel human prostate cancer progression model. Cancer Prev. Res. (Phila) 2015, 8, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.; Smith, D.M.; Dou, Q.P. Ester bond-containing tea polyphenols potently inhibit proteasome activity in vitro and in vivo. J. Biol. Chem. 2001, 276, 13322–13330. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Landis-Piwowar, K.; Chan, T.H.; Dou, Q.P. Green tea polyphenols as proteasome inhibitors: Implication in chemoprevention. Curr. Cancer Drug Targets 2011, 11, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Kazi, A.; Wang, Z.; Kumar, N.; Falsetti, S.C.; Chan, T.H.; Dou, Q.P. Structure-activity relationships of synthetic analogs of (−)-epigallocatechin-3-gallate as proteasome inhibitors. Anticancer Res. 2004, 24, 943–954. [Google Scholar] [PubMed]
- Smith, D.M.; Wang, Z.; Kazi, A.; Li, L.H.; Chan, T.H.; Dou, Q.P. Synthetic analogs of green tea polyphenols as proteasome inhibitors. Mol. Med. 2002, 8, 382–392. [Google Scholar] [PubMed]
- Modernelli, A.; Naponelli, V.; Giovanna Troglio, M.; Bonacini, M.; Ramazzina, I.; Bettuzzi, S.; Rizzi, F. EGCG antagonizes bortezomib cytotoxicity in prostate cancer cells by an autophagic mechanism. Sci. Rep. 2015, 5, 15270. [Google Scholar] [CrossRef] [PubMed]
- Naponelli, V.; Modernelli, A.; Bettuzzi, S.; Rizzi, F. Roles of autophagy induced by natural compounds in prostate cancer. Biomed. Res. Int. 2015, 2015, 121826. [Google Scholar] [CrossRef] [PubMed]
- Davalli, P.; Rizzi, F.; Caldara, G.F.; Davoli, S.; Corti, A.; Silva, A.; Astancolle, S.; Vitale, M.; Bettuzzi, S.; Arcari, M.; et al. Chronic administration of green tea extract to TRAMP mice induces the collapse of Golgi apparatus in prostate secretory cells and results in alterations of protein post-translational processing. Int. J. Oncol. 2011, 39, 1521–1527. [Google Scholar] [PubMed]
- Kikuchi, N.; Ohmori, K.; Shimazu, T.; Nakaya, N.; Kuriyama, S.; Nishino, Y.; Tsubono, Y.; Tsuji, I. No association between green tea and prostate cancer risk in Japanese men: The Ohsaki Cohort Study. Br. J. Cancer 2006, 95, 371–373. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.E.; Sauvaget, C.; Roddam, A.W.; Appleby, P.; Nagano, J.; Suzuki, G.; Key, T.J.; Koyama, K. A prospective study of diet and prostate cancer in Japanese men. Cancer Causes Control 2004, 15, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Severson, R.K.; Nomura, A.M.; Grove, J.S.; Stemmermann, G.N. A prospective study of demographics, diet, and prostate cancer among men of Japanese ancestry in Hawaii. Cancer Res. 1989, 49, 1857–1860. [Google Scholar] [PubMed]
- Kurahashi, N.; Sasazuki, S.; Iwasaki, M.; Inoue, M. Green tea consumption and prostate cancer risk in Japanese men: A prospective study. Am. J. Epidemiol. 2007, 167, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Jian, L.; Xie, L.P.; Lee, A.H.; Binns, C.W. Protective effect of green tea against prostate cancer: A case-control study in southeast China. Int. J. Cancer 2004, 108, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, T.; Nagata, Y.; Mori, M.; Miyanaga, N.; Takashima, N.; Okumura, K.; Goto, K.; Naito, S.; Fujimoto, K.; Hirao, Y.; et al. A case-control study of diet and prostate cancer in Japan: Possible protective effect of traditional Japanese diet. Cancer Sci. 2004, 95, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Adhami, V.M.; Siddiqui, I.A.; Sarfaraz, S.; Khwaja, S.I.; Hafeez, B.B.; Ahmad, N.; Mukhtar, H. Effective prostate cancer chemopreventive intervention with green tea polyphenols in the TRAMP model depends on the stage of the disease. Clin. Cancer Res. 2009, 15, 1947–1953. [Google Scholar] [CrossRef] [PubMed]
- Jatoi, A.; Ellison, N.; Burch, P.A.; Sloan, J.A.; Dakhil, S.R.; Novotny, P.; Tan, W.; Fitch, T.R.; Rowland, K.M.; Young, C.Y.; et al. A phase II trial of green tea in the treatment of patients with androgen independent metastatic prostate carcinoma. Cancer 2003, 97, 1442–1446. [Google Scholar] [CrossRef] [PubMed]
- McLarty, J.; Bigelow, R.L.; Smith, M.; Elmajian, D.; Ankem, M.; Cardelli, J.A. Tea polyphenols decrease serum levels of prostate-specific antigen, hepatocyte growth factor, and vascular endothelial growth factor in prostate cancer patients and inhibit production of hepatocyte growth factor and vascular endothelial growth factor in vitro. Cancer Prev. Res. (Phila) 2009, 2, 673–682. [Google Scholar] [PubMed]
- Nguyen, M.M.; Ahmann, F.R.; Nagle, R.B.; Hsu, C.H.; Tangrea, J.A.; Parnes, H.L.; Sokoloff, M.H.; Gretzer, M.B.; Chow, H.H. Randomized, double-blind, placebo-controlled trial of polyphenon e in prostate cancer patients before prostatectomy: Evaluation of potential chemopreventive activities. Cancer Prev. Res. (Phila) 2012, 5, 290–298. [Google Scholar] [CrossRef] [PubMed]
- O’Dowd G, J.; Miller, M.C.; Orozco, R.; Veltri, R.W. Analysis of repeated biopsy results within 1 year after a noncancer diagnosis. Urology 2000, 55, 553–559. [Google Scholar] [CrossRef]
- Kumar, N.B.; Pow-Sang, J.; Spiess, P.E.; Park, J.; Salup, R.; Williams, C.R.; Parnes, H.; Schell, M.J. Randomized, placebo-controlled trial evaluating the safety of one-year administration of green tea catechins. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Venigalla, S.; Zhao, C.; Miyamoto, H. Histopathologic features of atypical glands on prostate biopsy: Nucleolar size is a predictor of subsequent detection of prostatic adenocarcinoma. Prostate 2013, 73, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, P.; Wolski, Z.; Butkiewicz, R.; Nussbeutel, J.; Drewa, T. Significance of atypical small acinar proliferation and extensive high-grade prostatic intraepithelial neoplasm in clinical practice. Cent. Eur. J. Urol. 2014, 67, 136–141. [Google Scholar]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism, and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef] [PubMed]
- Chow, H.H.; Cai, Y.; Alberts, D.S.; Hakim, I.; Dorr, R.; Shahi, F.; Crowell, J.A.; Yang, C.S.; Hara, Y. Phase i pharmacokinetic study of tea polyphenols following single-dose administration of epigallocatechin gallate and polyphenon E. Cancer Epidemiol. Biomarkers Prev. 2001, 10, 53–58. [Google Scholar] [PubMed]
- Henning, S.M.; Aronson, W.; Niu, Y.; Conde, F.; Lee, N.H.; Seeram, N.P.; Lee, R.P.; Lu, J.; Harris, D.M.; Moro, A.; et al. Tea polyphenols and theaflavins are present in prostate tissue of humans and mice after green and black tea consumption. J. Nutr. 2006, 136, 1839–1843. [Google Scholar] [PubMed]
- Wang, P.; Aronson, W.J.; Huang, M.; Zhang, Y.; Lee, R.P.; Heber, D.; Henning, S.M. Green tea polyphenols and metabolites in prostatectomy tissue: Implications for cancer prevention. Cancer Prev. Res. 2010, 3, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Roowi, S.; Stalmach, A.; Mullen, W.; Lean, M.E.J.; Edwards, C.A.; Crozier, A. Green tea flavan-3-ols: Colonic degradation and urinary excretion of catabolites by humans. J. Agric. Food Chem. 2010, 58, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.A.; Adhami, V.M.; Bharali, D.J.; Hafeez, B.B.; Asim, M.; Khwaja, S.I.; Ahmad, N.; Cui, H.; Mousa, S.A.; Mukhtar, H. Introducing nanochemoprevention as a novel approach for cancer control: Proof of principle with green tea polyphenol epigallocatechin-3-gallate. Cancer Res. 2009, 69, 1712–1716. [Google Scholar] [CrossRef] [PubMed]
- Granja, A.; Pinheiro, M.; Reis, S. Epigallocatechin gallate nanodelivery systems for cancer therapy. Nutrients 2016, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, N.; De, R.; Begun, J.; Popat, A. Cancer therapeutics with epigallocatechin-3-gallate encapsulated in biopolymeric nanoparticles. Int. J. Pharm. 2017, 518, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Sanna, V.; Pintus, G.; Roggio, A.M.; Punzoni, S.; Posadino, A.M.; Arca, A.; Marceddu, S.; Bandiera, P.; Uzzau, S.; Sechi, M. Targeted biocompatible nanoparticles for the delivery of (−)-epigallocatechin 3-gallate to prostate cancer cells. J. Med. Chem. 2011, 54, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Sanna, V.; Singh, C.K.; Jashari, R.; Adhami, V.M.; Chamcheu, J.C.; Rady, I.; Sechi, M.; Mukhtar, H.; Siddiqui, I.A. Targeted nanoparticles encapsulating (−)-epigallocatechin-3-gallate for prostate cancer prevention and therapy. Sci. Rep. 2017, 7, 41573. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Bharali, D.J.; Adhami, V.M.; Siddiqui, I.A.; Cui, H.; Shabana, S.M.; Mousa, S.A.; Mukhtar, H. Oral administration of naturally occurring chitosan-based nanoformulated green tea polyphenol EGCG effectively inhibits prostate cancer cell growth in a xenograft model. Carcinogenesis 2013, 35, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.; Generalov, R.; Pereira, M.D.C.; Peres, I.; Juzenas, P.; Coelho, M.A.N. Epigallocatechin gallate-loaded polysaccharide nanoparticles for prostate cancer chemoprevention. Nanomedicine 2011, 6, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Kulhari, H.; Pooja, D.; Gudem, S.; Bhargava, S.; Shukla, R.; Sistla, R. Encapsulation of biophenolic phytochemical EGCG within lipid nanoparticles enhances its stability and cytotoxicity against cancer. Chem. Phys. Lipids 2016, 198, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.R.M.R.; Shafiei, S.S.; Solati-Hashjin, M.; Samadikuchaksaraei, A.; Kalantarinejad, R.; Asadi-Eydivand, M.; Abu Osman, N.A. Epigallocatechin gallate/layered double hydroxide nanohybrids: Preparation, characterization, and in vitro anti-tumor study. PLoS ONE 2015, 10, e0136530. [Google Scholar]
- Tsai, L.-C.; Hsieh, H.-Y.; Lu, K.-Y.; Wang, S.-Y.; Mi, F.-L. EGCG/gelatin-doxorubicin gold nanoparticles enhance therapeutic efficacy of doxorubicin for prostate cancer treatment. Nanomedicine 2016, 11, 9–30. [Google Scholar] [CrossRef] [PubMed]
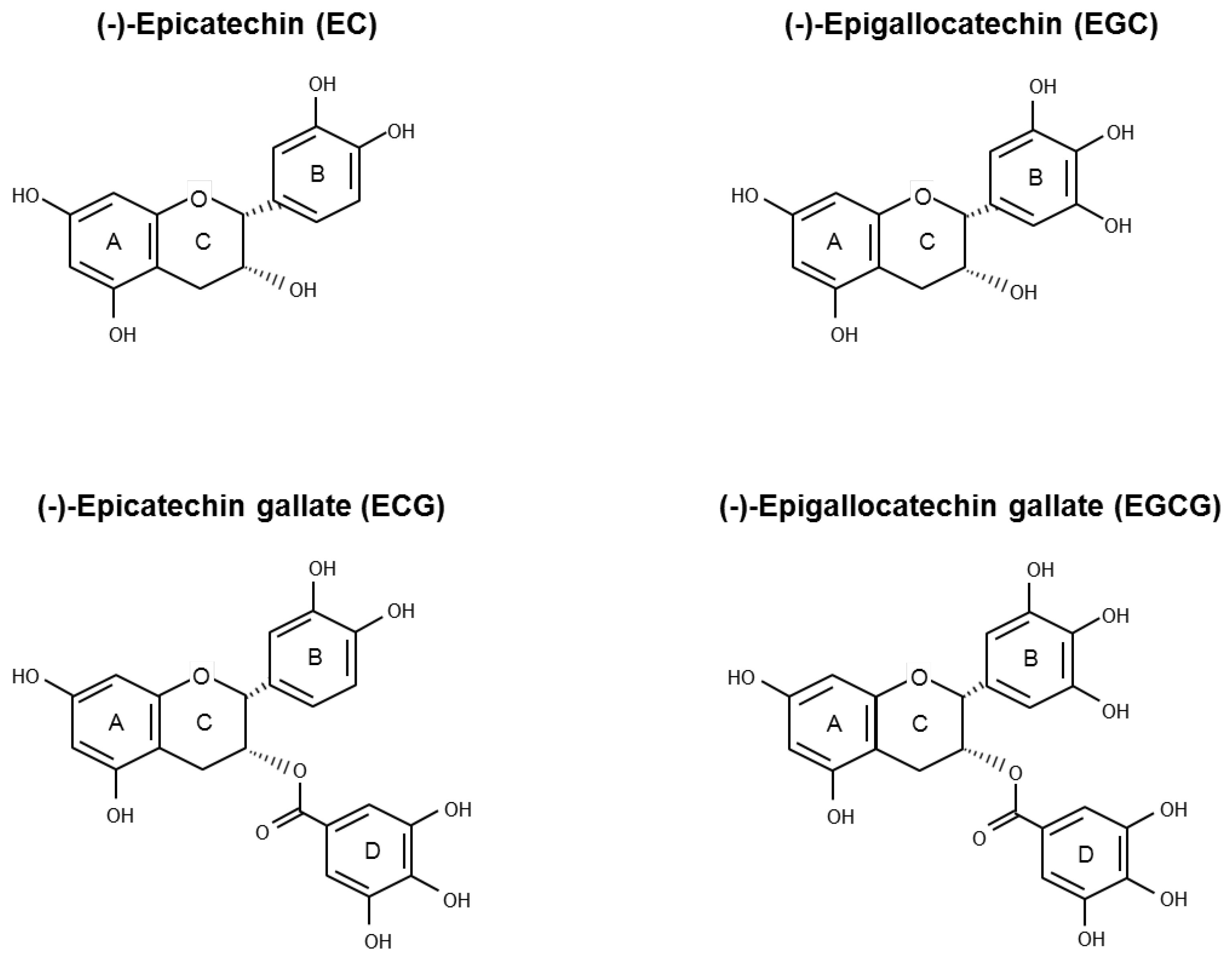
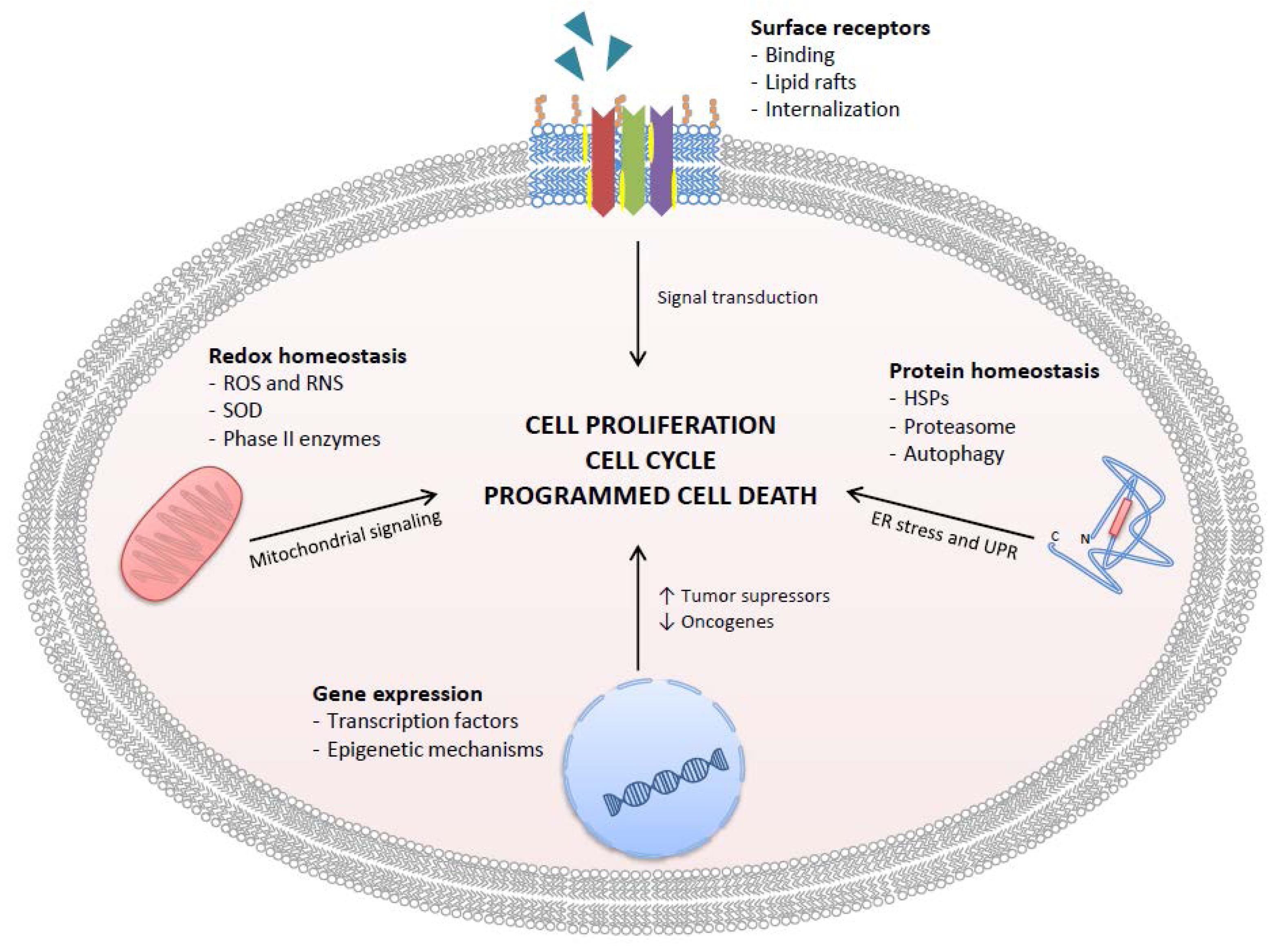
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).