
Zinc and Oxidative Stress: Current Mechanisms

{kind=link}
Abstract
:1. Introduction
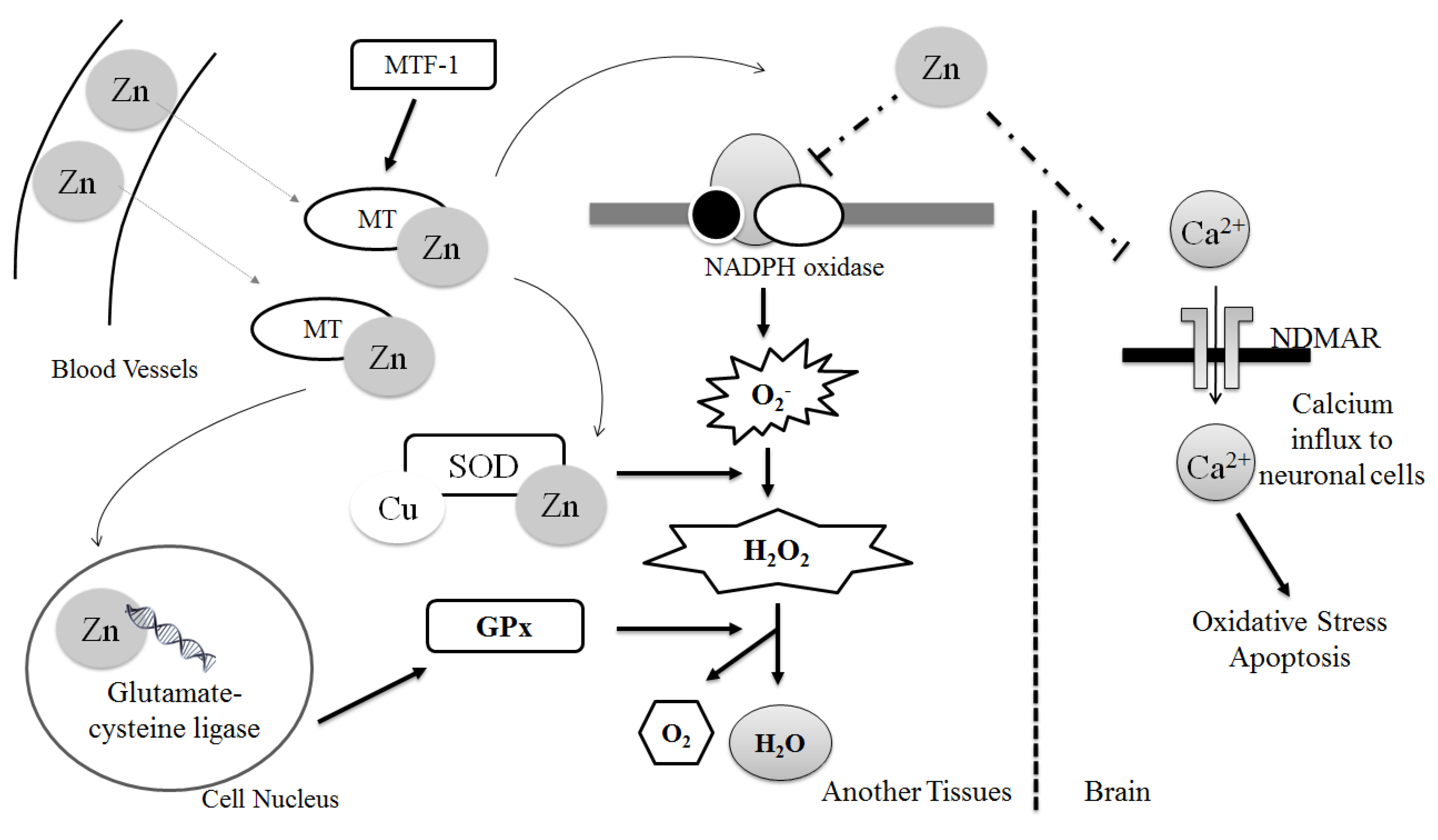
2. Biochemical Aspects
3. Chronic Diseases
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Butterfield, D.A.; Domenico, F.B. Elevated risk of type 2 diabetes for development of Alzheimer disease: A key role for oxidative stress in brain. Biochim. Biophys. Acta 2014, 1824, 1693–1706. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Ruiz, M.A. Oxidative-stress-induced epigenetic changes in chronic diabetic complications. Can. J. Physiol. Pharmacol. 2013, 91, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Roshanravan, N.; Alizadeh, M. Effect of zinc supplementation on insulin resistance, energy and macronutrients intakes in pregnant women with impaired glucose tolerance. Iran J. Public Health 2015, 44, 211–217. [Google Scholar] [PubMed]
- Jurowski, K.; Szewczyk, B. Biological consequences of zinc deficiency in the pathomechanisms of selected diseases. J. Biol. Inorg. Chem. 2014, 19, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.; Chu, A. Zinc transporter gene expression and glycemic control in post-menopausal women with type 2 diabetes mellitus. J. Trace Elem. Med. Biol. 2014, 28, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Fung, E.B.; Gildengorin, G. Zinc status affects glucose homeostasis and insulin secretion in patients with thalassemia. Nutrients 2015, 7, 4296–4307. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.; Rosenkranz, E. Disturbed zinc homeostasis in diabetic patients by in vitro and in vivo analysis of insulinomimetic activity of zinc. J. Nutr. Biochem. 2012, 23, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Ruz, M.; Carrasco, F. Zinc as a potential coadjuvant in therapy for type 2 diabetes. Food Nutr. Bull. 2013, 34, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Chasapis, C.T.; Loutsidou, A.C. Zinc and human health: An update. Arch. Toxicol. 2012, 86, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Cruz, K.J.C; Oliveira, A.R.S. Antioxidant role of zinc in diabetes mellitus. World J. Diabetes 2015, 6, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Lima, V.B.S.; Sampaio, F.A. Parameters of glycemic control and their relationship with zinc concentrations in blood and with superoxide dismutase enzyme activity in type 2 diabetes patients. Arq. Bras. Endocrinol. Metab. 2011, 55, 701–707. [Google Scholar]
- Cruz, J.B.F.; Soares, H.F. Uma revisão sobre o zinco. Ensaios Ciência Ciências Biológicas Agrárias Saúde 2011, 15, 207–222. [Google Scholar]
- Homma, K.; Fujisawa, T. SOD1 as a molecular switch for initiating the homeostatic ER stress response under zinc deficiency. Mol. Cell. 2013, 52, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. The oxidative stress of zinc deficiency. Metallomics 2011, 3, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Ha, K.N.; Chen, Y. Increased glutathione synthesis through an ARE-Nrf2-dependent pathway by zinc in the RPE: Implication for protection against oxidative stress. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2709–2715. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.; Samman, S. Zinc and redox signaling: Perturbations associated with cardiovascular disease and diabetes mellitus. Antioxid. Redox. Signal. 2010, 13, 1549–1573. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Krezel, A. Cellular zinc and redox buffering capacity of metallothionein/thionein in health and disease. Mol. Med. 2007, 13, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Özcelik, D.; Nazıroglu, M. Zinc supplementation attenuates metallothionein and oxidative stress changes in kidney of streptozotocin-induced diabetic rats. Biol. Trace Elem. Res. 2012, 150, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Zhang, Q. Zinc treatment prevents type 1 diabetes-induced hepatic oxidative damage, endoplasmic reticulum stress, and cell death, and even prevents possible steatohepatitis in the OVE26 mouse model: Important role of metallothionein. Toxicol. Lett. 2015, 233, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Coyle, P.; Philcox, J.C. Methallothionein: The Multipurpose Protein. Cell Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Günther, V.; Davis, A.L. A conserved cysteine cluster, essential for transcriptional activity, mediates homodimerization of human metal-responsive transcription factor-1 (MTF-1). Biochim. Biophys. Acta 2012, 1823, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Günther, V.; Lindert, U. The taste of heavy metals: Gene regulation by MTF-1. Biochim. Biophys. Acta 2012, 1823, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Andrews, G.K. Regulation of metallothionein gene expression by oxidative stress and metal ions. Biochem. Pharmacol. 2000, 59, 95–104. [Google Scholar] [CrossRef]
- Sekler, I.; Sensi, S.L. Mechanism and regulation of cellular zinc transport. Mol. Med. 2007, 13, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, P.; Benedetti, G. Zinc and its role in immunity and inflammation. Autoimmunity Rev. 2015, 14, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Grzywacz, A.; Gdula-Argasińska, J. Metal responsive transcription factor 1 (MTF-1) regulates zinc dependent cellular processes at the molecular level. Acta Biochim. Pol. 2015, 62, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Oteiza, P.I. Zinc and the modulation of redox homeostasis. Free Radic. Biol. Med. 2012, 53, 1748–1759. [Google Scholar] [CrossRef] [PubMed]
- Bryan, S.; Baregzay, B. Redox-inflammatory synergy in the metabolic syndrome. Can. J. Physiol. Pharmacol. 2013, 91, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S.; Bao, B. Zinc-suppressed inflammatory cytokines by induction of A20-mediated inhibition of nuclear factor-κB. Nutrition 2011, 27, 816–823. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Prasad, A.S. Zinc decreases C-reactive protein, lipid peroxidation, and inflammatory cytokines in elderly subjects: A potential implication of zinc as an atheroprotective agent. Am. J. Clin. Nutr. 2010, 91, 1634–1641. [Google Scholar] [CrossRef] [PubMed]
- Biagiotti, S.; Menotta, M. Dexamethasone improves redox state in ataxia telangiectasia cells by promoting an NRF2-mediated antioxidant response. FEBS J. 2016. [Google Scholar] [CrossRef] [PubMed]
- Jenner, A.; Ren, M. Zinc supplementation inhibits lipid peroxidation and the development of atherosclerosis in rabbits fed a high cholesterol diet. Free Radic. Biol. Med. 2007, 42, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Wang, Y. Zinc protects against diabetes-induced pathogenic changes in the aorta: Roles of metallothionein and nuclear factor (erythroid-derived 2)-like 2. Cardiovasc. Diabetol. 2013, 12, 54. [Google Scholar] [CrossRef] [PubMed]
- Weglicki, W.B.; Chmielinska, J.J. Cardiovascular and intestinal responses to oxidative and nitrosative stress during prolonged magnesium deficiency. Am. J. Med. Sci. 2011, 342, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Ranasinghe, P.; Pigera, S. Zinc and diabetes mellitus: Understanding molecular mechanisms and clinical implications. Daru 2015, 23, 44. [Google Scholar] [CrossRef] [PubMed]
- Vardatsikos, G.; Pandey, N.R. Insulino-mimetic and anti-diabetic effects of zinc. J. Inorg. Biochem. 2013, 120, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Xiang, L. ZnT7 can protect MC3T3-E1 cells from oxidative stress-induced apoptosis via PI3K/Akt and MAPK/ERK signaling pathways. Cell Signal. 2013, 25, 1126–1135. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Li, Q. Dysregulation of hepatic zinc transporters in a mouse model of alcoholic liver disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, T.B.; Chang, S.M. Zinc transporter ZIP14 functions in hepatic zinc, iron and glucose homeostasis during the innate immune response (endotoxemia). PLoS ONE 2012, 7, e48679. [Google Scholar]
- Kirschke, C.P.; Huang, L. ZnT7, a novel mammalian zinc transporter, accumulates zinc in the Golgi apparatus. J. Biol. Chem. 2003, 278, 4096–4102. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Morgan, H.E. Structure-function analysis of HKE4, a member of the new LIV-1 subfamily of zinc transporters. Biochem. J. 2004, 377, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhong, W. Zinc deficiency mediates alcohol-induced apoptotic cell death in the liver of rats through activating ER and mitochondrial cell death pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Sandstead, H.H. Zinc requirements and the risks and benefits of zinc supplementation. J. Trace Elem. Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Clegg, M.S.; Hanna, L.A. Zinc deficiency-induced cell death. IUBMB Life 2005, 57, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Metallothionein redox biology in the cytoprotective and cytotoxic functions of zinc. Exp. Gerontol. 2008, 43, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.M.; Oliveira, A.R.S. Influence of cortisol on zinc metabolism in morbidly obese women. Nutr. Hosp. 2014, 29, 57–63. [Google Scholar]
- Sulibuska, J.; Bogdanski, P. Changes in mineral status are associated with improvements in insulin sensitivity in obese patients following L-arginine supplementation. Eur. J. Nutr. 2014, 53, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.A.; Saad, E.A. Pro-inflammatory adipocytokines, oxidative stress, insulin, Zn and Cu: Interrelations with obesity in Egyptian non-diabetic obese children and adolescents. Adv. Med. Sci. 2015, 60, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, M.N.; Wiebe, H.B. Trace elements in hemodialysis patients: A systematic review and meta-analysis. BMC Med. 2009, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, R.C.N.; Araújo, C.G.B. Nutritional status of zinc and activity superoxide dismutase in chronic renal patients undergoing hemodialysis. Nutr. Hosp. 2011, 26, 1456–1461. [Google Scholar]
- Guo, C.H.; Wang, C.L. Effects of zinc supplementation on plasma copper/zinc ratios, oxidative stress, and immunological status in hemodialysis patients. Int. J. Med. Sci. 2013, 10, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Sensi, S.L.; Paoletti, P. The neurophysiology and pathology of brain zinc. J. Neurosci. 2011, 31, 16076–16085. [Google Scholar] [CrossRef] [PubMed]
- Vergnano, A.M.; Rebola, N. Zinc dynamics and action at excitatory synapses. Neuron 2014, 82, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.R. Magnesium Status and Its Relationship with C-Reactive Protein in Obese Women. Biol. Trace Elem. Res. 2015, 168, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Prabasheela, B.; Singh, A.K. Association between Antioxidant Enzymes and Breast Cancer. Rec. Res. Sci. Tech. 2011, 3, 93–95. [Google Scholar]
- Bostanci, Z.; Mack, R.P. Paradoxical zinc toxicity and oxidative stress in the mammary gland during marginal dietary zinc deficiency. Reprod. Toxicol. 2015, 54, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Kelleher, S.L. Cellular mechanisms of zinc dysregulation: A perspective on zinc homeostasis as an etiological factor in the development and progression of breast cancer. Nutrients 2012, 4, 875–903. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Koh, J.Y. The role of NADPH oxidase and neuronal nitric oxide synthase in zinc-induced poly (ADP-ribose) polymerase activation and cell death in cortical culture. Exp. Neurol. 2002, 177, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y.; Segawa, S. Microglial zinc uptake via zinc transporters induces ATP release and the activation of microglia. Glia 2011, 59, 1933–1945. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Ohshima, C. Oxidative stress upregulates zinc uptake activity via Zrt/Irt-like protein 1 (ZIP1) in cultured mouse astrocytes. Life Sci. 2016, 151, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.M.; Liu, Z. The zinc ion chelating agent TPEN attenuates neuronal death/apoptosis caused by hypoxia/ischemia via mediating the pathophysiological cascade including excitotoxicity, oxidative stress, and inflammation. CNS Neurosci. Ther. 2015, 21, 708–717. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marreiro, D.D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; De Oliveira, A.R.S. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants 2017, 6, 24. https://doi.org/10.3390/antiox6020024
Marreiro DDN, Cruz KJC, Morais JBS, Beserra JB, Severo JS, De Oliveira ARS. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants. 2017; 6(2):24. https://doi.org/10.3390/antiox6020024
Chicago/Turabian StyleMarreiro, Dilina Do Nascimento, Kyria Jayanne Clímaco Cruz, Jennifer Beatriz Silva Morais, Jéssica Batista Beserra, Juliana Soares Severo, and Ana Raquel Soares De Oliveira. 2017. "Zinc and Oxidative Stress: Current Mechanisms" Antioxidants 6, no. 2: 24. https://doi.org/10.3390/antiox6020024
APA StyleMarreiro, D. D. N., Cruz, K. J. C., Morais, J. B. S., Beserra, J. B., Severo, J. S., & De Oliveira, A. R. S. (2017). Zinc and Oxidative Stress: Current Mechanisms. Antioxidants, 6(2), 24. https://doi.org/10.3390/antiox6020024