
Glucosinolate Metabolites and Brain Health: An Updated Review on Their Potential Benefits in Neurodegenerative, Neurodevelopmental, and Psychiatric Disorders


Abstract
1. Introduction
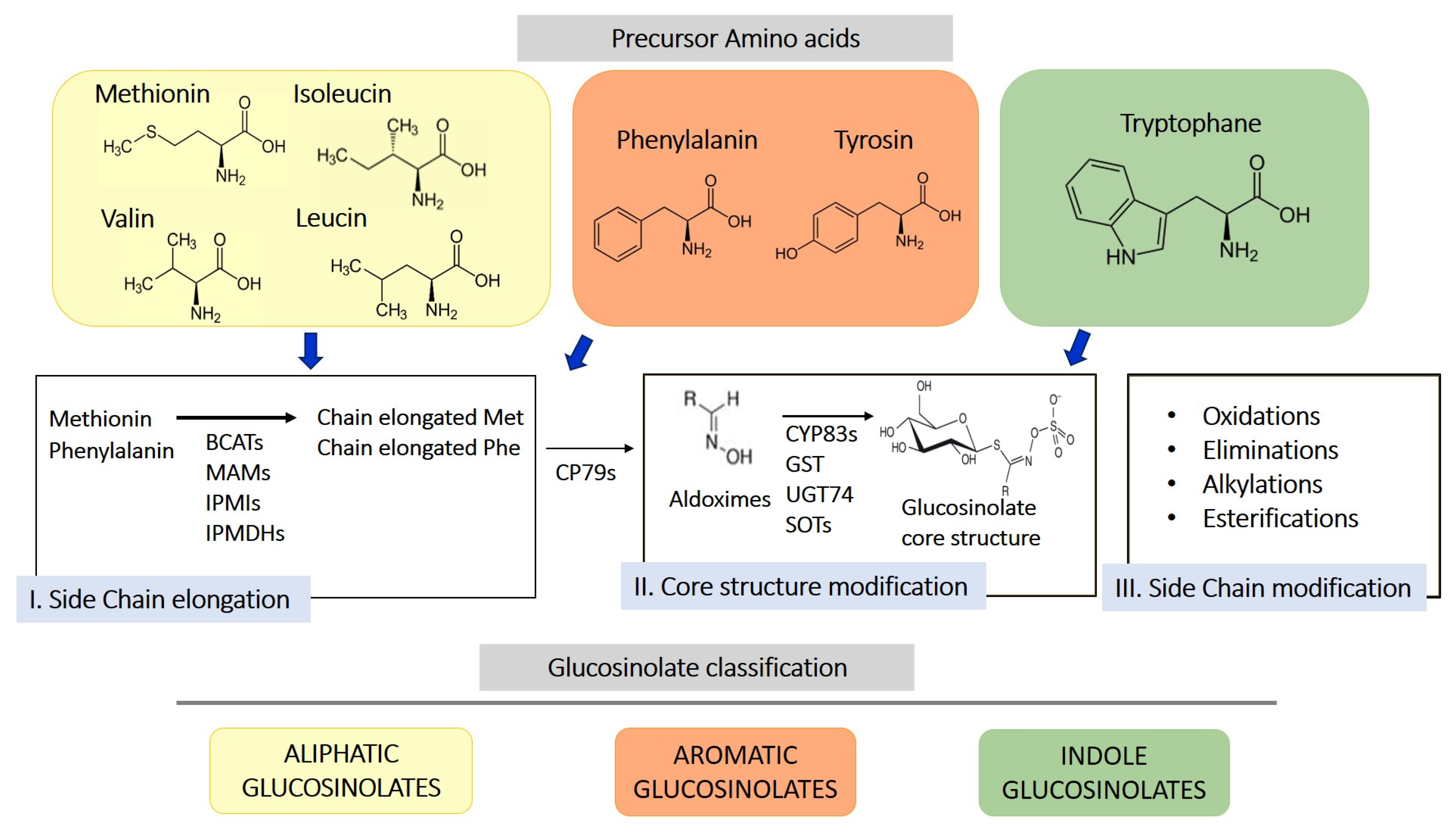
2. Origin, Structure, and Biosynthesis of Glucosinolates and Their Derivatives
3. Metabolism and Bioavailability of Isothiocyanates
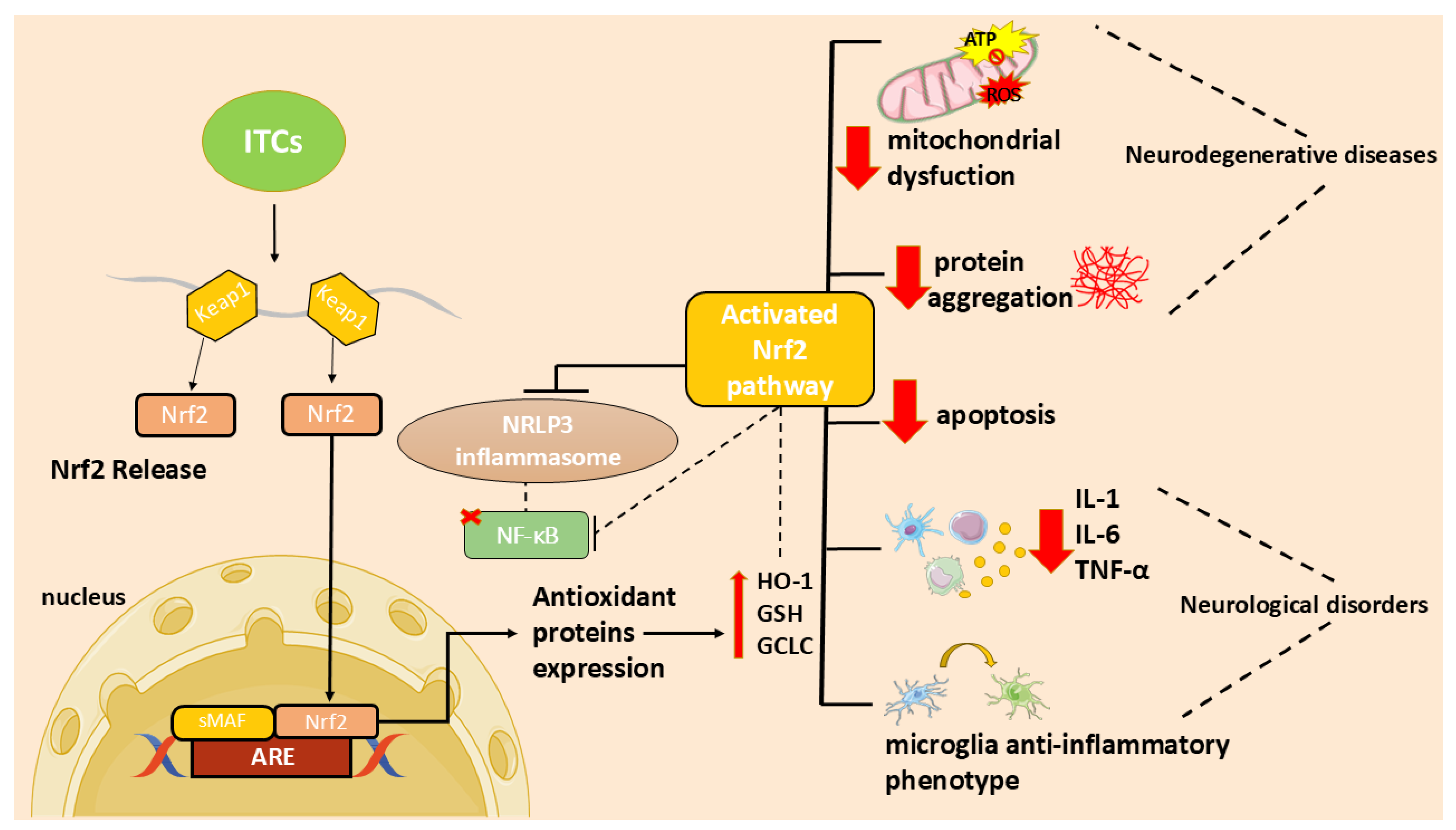
4. Neuroinflammation and Oxidative Stress
Preclinical Studies on Glucosinolate or Isothiocyanates in Neuroinflammation and Oxidative Stress
5. Neurodegenerative Disorders
5.1. Alzheimer’s Disease (AD)
Preclinical Studies on Glucosinolates or Isothiocyanates in AD
5.2. Parkinson’s Disease (PD)
5.2.1. Preclinical Studies on Glucosinolate or Isothiocyanates in PD
5.2.2. Clinical Studies on Glucosinolates or Isothiocyanates in PD
5.3. Multiple Sclerosis (MS)
Preclinical Studies on Glucosinolates or Isothiocyanates in MS
6. Neurodevelopmental Disorders, Psychiatric Conditions, and Epilepsy
6.1. Autism Spectrum Disorder (ASD)
6.1.1. Preclinical Studies on Glucosinolate or Isothiocyanates in ASD
6.1.2. Clinical Studies on Glucosinolate or Isothiocyanates and ASD
6.2. Schizophrenia
6.2.1. Preclinical Studies on Glucosinolates or Isothiocyanates in Schizophrenia
6.2.2. Clinical Studies on Glucosinolate or Isothiocyanates and Schizophrenia
6.3. Depression and Anxiety
6.3.1. Preclinical Studies on Glucosinolate or Isothiocyanates and Depression/Anxiety
6.3.2. Clinical Study on Sulforaphane in Depression/Anxiety
6.4. Epilepsy
Preclinical Studies on Glucosinolate or Isothiocyanates in Epilepsy
7. Adverse Effects
8. Clinical Challenges and Future Perspectives
9. Methods
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| PD | Parkinson’s disease |
| MS | Multiple sclerosis |
| ASD | Autism spectrum disorder |
| GLS | Glucosinolate |
| ITCs | Isothiocyanates |
| Nrf2 | Nuclear factor 2-related erythroid-derived factor |
| ARE | Antioxidant response element |
| NF-κB | Nuclear factor-κB |
| SFN | Sulforaphane |
| MYR | Myrosinase |
| GRA | Glucoraphanin |
| SIN | Sinigrin |
| AITC | Allyl isothiocyanate |
| GST | Gluconasturtiin |
| PEITC | Phenethyl isothiocyanate |
| GER | Glucoerucin |
| ER | Erucin |
| GTL | Glucotropaeolin |
| BITC | Benzyl isothiocyanate |
| GMG | Glucomoringin |
| MOR | Moringin |
| GSH | Glutathione |
| GST | Glutathione S-transferases |
| GTP | Gamma-glutamyl transpeptidase |
| CG | Cysteinylglycinase |
| NAT | N-acetyl transferase |
| NAC-ITC | N-acetylcysteine isothiocyanate |
| CNS | Central nervous system |
| ROS | Reactive oxygen species |
| LPS | Lipopolysaccharide |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| COX | Cyclooxygenase |
| NO | Nitric oxide |
| PGE2 | Prostaglandin E2 |
| MGO | Methylglyoxal |
| CPF | Chlorpyrifos |
| HO-1 | Heme oxygenase-1 |
| BDNF | Brain-derived neurotrophic factor |
| OKA | Okadaic Acid |
| TNF | Tumor necrosis factor |
| TBI | Traumatic brain injury |
| MHE | Minimal hepatic encephalopathy |
| GMG-ITC | Glucomoringin-isothiocyanate |
| MMP | Metalloproteinase |
| Aβ | Amyloid β |
| RB | Romanesco broccoli |
| PB | Purple broccoli |
| ESI-MS | Electrospray ionization mass spectrometry |
| BACE1 | β-site amyloid precursor protein cleaving enzyme 1 |
| NQO1 | NAD(P)H quinone oxidoreductase |
| SNP | Sodium nitroprusside |
| HSP | Heat shock protein |
| AQP4 | Aquaporin-4 |
| HDACs | Histone deacetylases |
| CVI | Cognitive vascular impartment |
| SCOP | Scopolamine |
| BRO | Broccoli extract |
| 6-MSITC | 6-methylsulfinyl hexyl isothiocyanate |
| ADAM17 | Metalloproteinase 17 |
| AChE | Acetylcholinesterase |
| LB | Lewy bodies |
| α-syn | α-synuclein |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| C/EBPβ | CCAAT/enhancer-binding protein β |
| MeCP2 | Methyl-binding protein CpG2 |
| RTT | Rett syndrome |
| 6-OHDA | 6-hydroxydopamine |
| TH | Tyrosine hydroxylase |
| BST | Broccoli seed tea |
| BBB | Blood–brain barrier |
| MIF | Migration inhibitory factor |
| EAE | Experimental autoimmune encephalitis |
| Trx1/TrxR1 | Thioredoxin 1/thioredoxin reductase 1 |
| EVs | Extracellular vesicles |
| PBMCs | Peripheral blood mononuclear cells |
| MIA | Maternal immune activation |
| mPFC | Medial prefrontal cortex |
| ABC | Aberrant Behavior Checklist |
| ADOS-2 | Autism Diagnostic Observation Schedule-2 |
| SRS-2 | Social Responsiveness Scale-2 |
| OARS-4 | OSU Autism Rating Scale-DSM-IV |
| iMGLC | hiPSC-derived microglia-like cells |
| 4-HNE | 4-hydroxynonenal |
| OCLT | One Card Learning Task |
| MCCB | MATRICS Consensus Cognitive Battery |
| HPA | Hypothalamic–pituitary–adrenal |
| SSRIs | Selective serotonin reuptake inhibitors |
| TST | Tail suspension test |
| FST | Forced swim test |
| SPT | Sucrose preference test |
| OFT | Open field test |
| ACTH | Adrenocorticotropic hormone |
| DG | Dentate gyrus |
| PFC | Prefrontal cortex |
| NAc | Nucleus accumbens |
| AβOs | β-amyloid oligomers |
| PITC | Phenyl isothiocyanate |
| CSDS | Chronic social defeat stress |
| SIT | Social interaction test |
| TAC | Total antioxidant capacity |
| PCI | Percutaneous coronary intervention |
| TLE | Temporal lobe epilepsy |
| SE | Status epilepticus |
| CABG | Coronary artery bypass grafting |
| HAM-D | Hamilton Rating Scale for Depression |
| PAHs | Polycyclic aromatic hydrocarbons |
References
- Teleanu, D.M.; Niculescu, A.G.; Lungu, I.I.; Radu, C.I.; Vladacenco, O.; Roza, E.; Costachescu, B.; Grumezescu, A.M.; Teleanu, R.I. An Overview of Oxidative Stress, Neuroinflammation, and Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef] [PubMed]
- GBD Mental Disorders Collaborators. Global, regional, and national burden of 12 mental disorders in 204 countries and territories, 1990-2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet Psychiatry 2022, 9, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Doernberg, E.; Hollander, E. Neurodevelopmental Disorders (ASD and ADHD): DSM-5, ICD-10, and ICD-11. CNS Spectr. 2016, 21, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Curpan, A.S.; Luca, A.C.; Ciobica, A. Potential Novel Therapies for Neurodevelopmental Diseases Targeting Oxidative Stress. Oxidative Med. Cell. Longev. 2021, 2021, 6640206. [Google Scholar] [CrossRef]
- Jiang, Q.; Liu, J.; Huang, S.; Wang, X.Y.; Chen, X.; Liu, G.H.; Ye, K.; Song, W.; Masters, C.L.; Wang, J.; et al. Antiageing strategy for neurodegenerative diseases: From mechanisms to clinical advances. Signal Transduct. Target. Ther. 2025, 10, 76. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Rollin, P. Glucosinolates: Novel Sources and Biological Potential. In Glucosinolates; Mérillon, J.-M., Ramawat, K.G., Eds.; Springer International Publishing: Cham, Germany, 2016; pp. 1–58. [Google Scholar]
- Giacoppo, S.; Galuppo, M.; Montaut, S.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. An overview on neuroprotective effects of isothiocyanates for the treatment of neurodegenerative diseases. Fitoterapia 2015, 106, 12–21. [Google Scholar] [CrossRef]
- Barba, F.J.; Nikmaram, N.; Roohinejad, S.; Khelfa, A.; Zhu, Z.; Koubaa, M. Bioavailability of Glucosinolates and Their Breakdown Products: Impact of Processing. Front. Nutr. 2016, 3, 24. [Google Scholar] [CrossRef]
- Liu, J.; Chandaka, G.K.; Zhang, R.; Parfenova, H. Acute antioxidant and cytoprotective effects of sulforaphane in brain endothelial cells and astrocytes during inflammation and excitotoxicity. Pharmacol. Res. Perspect. 2020, 8, e00630. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Agerbirk, N.; Olsen, C.E. Glucosinolate structures in evolution. Phytochemistry 2012, 77, 16–45. [Google Scholar] [CrossRef]
- Bird, K.A.; Ramos, A.A.; Kliebenstein, D.J. Phylogenetic and genomic mechanisms shaping glucosinolate innovation. Curr. Opin. Plant Biol. 2025, 85, 102705. [Google Scholar] [CrossRef]
- Garcia, G.; Treccarichi, S.; Arena, D.; Ben Ammar, H.; Maggioni, L.; Branca, F. Capturing the L. wild relatives diversity for improving nutraceutical traits of cole crops. Genet. Resour. Crop Evol. 2025. [Google Scholar] [CrossRef]
- Lo Scalzo, R.; Bianchi, G.; Picchi, V.; Campanelli, G.; Ficcadenti, N.; Treccarichi, S.; Arena, D.; Sestili, S.; Branca, F. Compositional traits of hybrid populations of Brassica oleracea L. var. italica (broccoli) and Brassica oleracea L. var. botrytis (cauliflower) during four organic breeding cycles. J. Food Compos. Anal. 2024, 131, 106209. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Yuan, J.M.; Kensler, T.W.; Fahey, J.W. Isothiocyanates: Translating the Power of Plants to People. Mol. Nutr. Food Res. 2018, 62, e1700965. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef]
- Williams, D.J.; Critchley, C.; Pun, S.; Chaliha, M.; O’Hare, T.J. Differing mechanisms of simple nitrile formation on glucosinolate degradation in Lepidium sativum and Nasturtium officinale seeds. Phytochemistry 2009, 70, 1401–1409. [Google Scholar] [CrossRef]
- Kim, S.H.; Ochar, K.; Iwar, K.; Lee, Y.J.; Kang, H.J.; Na, Y.W. Variations of Major Glucosinolates in Diverse Chinese Cabbage (Brassica rapa ssp. pekinensis) Germplasm as Analyzed by UPLC-ESI-MS/MS. Int. J. Mol. Sci. 2024, 25, 4829. [Google Scholar] [CrossRef]
- Grubb, C.D.; Abel, S. Glucosinolate metabolism and its control. Trends Plant Sci. 2006, 11, 89–100. [Google Scholar] [CrossRef]
- Holst, B.; Williamson, G. A critical review of the bioavailability of glucosinolates and related compounds. Nat. Prod. Rep. 2004, 21, 425–447. [Google Scholar] [CrossRef]
- Jaafaru, M.S.; Abd Karim, N.A.; Enas, M.E.; Rollin, P.; Mazzon, E.; Abdull Razis, A.F. Protective Effect of Glucosinolates Hydrolytic Products in Neurodegenerative Diseases (NDDs). Nutrients 2018, 10, 580. [Google Scholar] [CrossRef]
- Vanduchova, A.; Anzenbacher, P.; Anzenbacherova, E. Isothiocyanate from Broccoli, Sulforaphane, and Its Properties. J. Med. Food 2019, 22, 121–126. [Google Scholar] [CrossRef]
- Burcul, F.; Generalic Mekinic, I.; Radan, M.; Rollin, P.; Blazevic, I. Isothiocyanates: Cholinesterase inhibiting, antioxidant, and anti-inflammatory activity. J. Enzym. Inhib. Med. Chem. 2018, 33, 577–582. [Google Scholar] [CrossRef]
- Prabu, S.L.; Umamaheswari, A.; Puratchikody, A. Phytopharmacological potential of the natural gift Moringa oleifera Lam and its therapeutic application: An overview. Asian Pac. J. Trop. Med. 2019, 12, 485–498. [Google Scholar] [CrossRef]
- Yadav, K.; Dhankhar, J.; Preeti, K. Isothiocyanates—A Review of their Health Benefits and Potential Food Applications. Curr. Res. Nutr. Food Sci. 2022, 10, 476–502. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef]
- Zhang, Y.; Kolm, R.H.; Mannervik, B.; Talalay, P. Reversible conjugation of isothiocyanates with glutathione catalyzed by human glutathione transferases. Biochem. Biophys. Res. Commun. 1995, 206, 748–755. [Google Scholar] [CrossRef]
- Egner, P.A.; Kensler, T.W.; Chen, J.G.; Gange, S.J.; Groopman, J.D.; Friesen, M.D. Quantification of sulforaphane mercapturic acid pathway conjugates in human urine by high-performance liquid chromatography and isotope-dilution tandem mass spectrometry. Chem. Res. Toxicol. 2008, 21, 1991–1996. [Google Scholar] [CrossRef]
- Shapiro, T.A.; Fahey, J.W.; Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Stephenson, K.K.; Wade, K.L.; Ye, L.; Talalay, P. Safety, tolerance, and metabolism of broccoli sprout glucosinolates and isothiocyanates: A clinical phase I study. Nutr. Cancer 2006, 55, 53–62. [Google Scholar] [CrossRef]
- Getahun, S.M.; Chung, F.L. Conversion of glucosinolates to isothiocyanates in humans after ingestion of cooked watercress. Cancer Epidemiol. Biomark. Prev. 1999, 8, 447–451. [Google Scholar]
- Conaway, C.C.; Getahun, S.M.; Liebes, L.L.; Pusateri, D.J.; Topham, D.K.; Botero-Omary, M.; Chung, F.L. Disposition of glucosinolates and sulforaphane in humans after ingestion of steamed and fresh broccoli. Nutr. Cancer 2000, 38, 168–178. [Google Scholar] [CrossRef]
- Rungapamestry, V.; Duncan, A.J.; Fuller, Z.; Ratcliffe, B. Changes in glucosinolate concentrations, myrosinase activity, and production of metabolites of glucosinolates in cabbage (Brassica oleracea Var. capitata) cooked for different durations. J. Agric. Food Chem. 2006, 54, 7628–7634. [Google Scholar] [CrossRef]
- Rouzaud, G.; Young, S.A.; Duncan, A.J. Hydrolysis of glucosinolates to isothiocyanates after ingestion of raw or microwaved cabbage by human volunteers. Cancer Epidemiol. Biomark. Prev. 2004, 13, 125–131. [Google Scholar] [CrossRef]
- Shapiro, T.A.; Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Chemoprotective glucosinolates and isothiocyanates of broccoli sprouts: Metabolism and excretion in humans. Cancer Epidemiol. Biomark. Prev. 2001, 10, 501–508. [Google Scholar]
- Egner, P.A.; Chen, J.G.; Wang, J.B.; Wu, Y.; Sun, Y.; Lu, J.H.; Zhu, J.; Zhang, Y.H.; Chen, Y.S.; Friesen, M.D.; et al. Bioavailability of Sulforaphane from two broccoli sprout beverages: Results of a short-term, cross-over clinical trial in Qidong, China. Cancer Prev. Res. 2011, 4, 384–395. [Google Scholar] [CrossRef]
- Cramer, J.M.; Jeffery, E.H. Sulforaphane absorption and excretion following ingestion of a semi-purified broccoli powder rich in glucoraphanin and broccoli sprouts in healthy men. Nutr. Cancer 2011, 63, 196–201. [Google Scholar] [CrossRef]
- Tang, L.; Zirpoli, G.R.; Guru, K.; Moysich, K.B.; Zhang, Y.; Ambrosone, C.B.; McCann, S.E. Intake of cruciferous vegetables modifies bladder cancer survival. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1806–1811. [Google Scholar] [CrossRef]
- Rivest, S. Regulation of innate immune responses in the brain. Nat. Rev. Immunol. 2009, 9, 429–439. [Google Scholar] [CrossRef]
- Subedi, L.; Lee, J.H.; Yumnam, S.; Ji, E.; Kim, S.Y. Anti-Inflammatory Effect of Sulforaphane on LPS-Activated Microglia Potentially through JNK/AP-1/NF-kappaB Inhibition and Nrf2/HO-1 Activation. Cells 2019, 8, 194. [Google Scholar] [CrossRef]
- Qin, S.; Yang, C.; Huang, W.; Du, S.; Mai, H.; Xiao, J.; Lu, T. Sulforaphane attenuates microglia-mediated neuronal necroptosis through down-regulation of MAPK/NF-kappaB signaling pathways in LPS-activated BV-2 microglia. Pharmacol. Res. 2018, 133, 218–235. [Google Scholar] [CrossRef]
- Tufekci, K.U.; Ercan, I.; Isci, K.B.; Olcum, M.; Tastan, B.; Gonul, C.P.; Genc, K.; Genc, S. Sulforaphane inhibits NLRP3 inflammasome activation in microglia through Nrf2-mediated miRNA alteration. Immunol. Lett. 2021, 233, 20–30. [Google Scholar] [CrossRef]
- Subedi, L.; Lee, J.H.; Gaire, B.P.; Kim, S.Y. Sulforaphane Inhibits MGO-AGE-Mediated Neuroinflammation by Suppressing NF-kappaB, MAPK, and AGE-RAGE Signaling Pathways in Microglial Cells. Antioxidants 2020, 9, 792. [Google Scholar] [CrossRef]
- Townsend, B.E.; Johnson, R.W. Sulforaphane induces Nrf2 target genes and attenuates inflammatory gene expression in microglia from brain of young adult and aged mice. Exp. Gerontol. 2016, 73, 42–48. [Google Scholar] [CrossRef]
- Brasil, F.B.; de Almeida, F.J.S.; Luckachaki, M.D.; Dall’Oglio, E.L.; de Oliveira, M.R. The isothiocyanate sulforaphane prevents mitochondrial impairment and neuroinflammation in the human dopaminergic SH-SY5Y and in the mouse microglial BV2 cells: Role for heme oxygenase-1. Metab. Brain Dis. 2023, 38, 419–435. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, M.; Wu, J.; Hu, P.; Xu, X.; Zhang, Y.; Wang, D.; Chen, Z.; Huang, C. Sulforaphane triggers a functional elongation of microglial process via the Akt signal. J. Nutr. Biochem. 2019, 67, 51–62. [Google Scholar] [CrossRef]
- Wang, Z.C.; Chen, Q.; Wang, J.; Yu, L.S.; Chen, L.W. Sulforaphane mitigates LPS-induced neuroinflammation through modulation of Cezanne/NF-kappaB signalling. Life Sci. 2020, 262, 118519. [Google Scholar] [CrossRef]
- Gao, J.; Xiong, B.; Zhang, B.; Li, S.; Huang, N.; Zhan, G.; Jiang, R.; Yang, L.; Wu, Y.; Miao, L.; et al. Sulforaphane Alleviates Lipopolysaccharide-induced Spatial Learning and Memory Dysfunction in Mice: The Role of BDNF-mTOR Signaling Pathway. Neuroscience 2018, 388, 357–366. [Google Scholar] [CrossRef]
- Dwivedi, S.; Rajasekar, N.; Hanif, K.; Nath, C.; Shukla, R. Sulforaphane Ameliorates Okadaic Acid-Induced Memory Impairment in Rats by Activating the Nrf2/HO-1 Antioxidant Pathway. Mol. Neurobiol. 2016, 53, 5310–5323. [Google Scholar] [CrossRef]
- Castro-Sanchez, S.; Garcia-Yague, A.J.; Kugler, S.; Lastres-Becker, I. CX3CR1-deficient microglia shows impaired signalling of the transcription factor NRF2: Implications in tauopathies. Redox Biol. 2019, 22, 101118. [Google Scholar] [CrossRef]
- Kyyriainen, J.; Kajevu, N.; Banuelos, I.; Lara, L.; Lipponen, A.; Balosso, S.; Hamalainen, E.; Das Gupta, S.; Puhakka, N.; Natunen, T.; et al. Targeting Oxidative Stress with Antioxidant Duotherapy after Experimental Traumatic Brain Injury. Int. J. Mol. Sci. 2021, 22, 10555. [Google Scholar] [CrossRef]
- Townsend, B.E.; Johnson, R.W. Sulforaphane reduces lipopolysaccharide-induced proinflammatory markers in hippocampus and liver but does not improve sickness behavior. Nutr. Neurosci. 2017, 20, 195–202. [Google Scholar] [CrossRef]
- Canto, A.; Martinez-Gonzalez, J.; Miranda, M.; Olivar, T.; Almansa, I.; Hernandez-Rabaza, V. Sulforaphane Modulates the Inflammation and Delays Neurodegeneration on a Retinitis Pigmentosa Mice Model. Front. Pharmacol. 2022, 13, 811257. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Rabaza, V.; Cabrera-Pastor, A.; Taoro-Gonzalez, L.; Gonzalez-Usano, A.; Agusti, A.; Balzano, T.; Llansola, M.; Felipo, V. Neuroinflammation increases GABAergic tone and impairs cognitive and motor function in hyperammonemia by increasing GAT-3 membrane expression. Reversal by sulforaphane by promoting M2 polarization of microglia. J. Neuroinflamm. 2016, 13, 83. [Google Scholar] [CrossRef] [PubMed]
- Franke, M.; Bieber, M.; Kraft, P.; Weber, A.N.R.; Stoll, G.; Schuhmann, M.K. The NLRP3 inflammasome drives inflammation in ischemia/reperfusion injury after transient middle cerebral artery occlusion in mice. Brain Behav. Immun. 2021, 92, 223–233. [Google Scholar] [CrossRef]
- Subedi, L.; Cho, K.; Park, Y.U.; Choi, H.J.; Kim, S.Y. Sulforaphane-Enriched Broccoli Sprouts Pretreated by Pulsed Electric Fields Reduces Neuroinflammation and Ameliorates Scopolamine-Induced Amnesia in Mouse Brain through Its Antioxidant Ability via Nrf2-HO-1 Activation. Oxidative Med. Cell. Longev. 2019, 2019, 3549274. [Google Scholar] [CrossRef]
- Townsend, B.E.; Chen, Y.J.; Jeffery, E.H.; Johnson, R.W. Dietary broccoli mildly improves neuroinflammation in aged mice but does not reduce lipopolysaccharide-induced sickness behavior. Nutr. Res. 2014, 34, 990–999. [Google Scholar] [CrossRef]
- Lee, C.M.; Lee, D.S.; Jung, W.K.; Yoo, J.S.; Yim, M.J.; Choi, Y.H.; Park, S.; Seo, S.K.; Choi, J.S.; Lee, Y.M.; et al. Benzyl isothiocyanate inhibits inflammasome activation in E. coli LPS-stimulated BV2 cells. Int. J. Mol. Med. 2016, 38, 912–918. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Giacoppo, S.; Ficicchia, M.; Aliquo, A.; Bramanti, P.; Mazzon, E. Eruca sativa seed extract: A novel natural product able to counteract neuroinflammation. Mol. Med. Rep. 2018, 17, 6235–6244. [Google Scholar] [CrossRef]
- Jaafaru, M.S.; Nordin, N.; Rosli, R.; Shaari, K.; Bako, H.Y.; Saad, N.; Noor, N.M.; Abdull Razis, A.F. Neuroprotective effects of glucomoringin-isothiocyanate against H(2)O(2)-Induced cytotoxicity in neuroblastoma (SH-SY5Y) cells. Neurotoxicology 2019, 75, 89–104. [Google Scholar] [CrossRef]
- Subedi, L.; Venkatesan, R.; Kim, S.Y. Neuroprotective and Anti-Inflammatory Activities of Allyl Isothiocyanate through Attenuation of JNK/NF-kappaB/TNF-alpha Signaling. Int. J. Mol. Sci. 2017, 18, 1423. [Google Scholar] [CrossRef]
- Latronico, T.; Larocca, M.; Milella, S.; Fasano, A.; Rossano, R.; Liuzzi, G.M. Neuroprotective potential of isothiocyanates in an in vitro model of neuroinflammation. Inflammopharmacology 2021, 29, 561–571. [Google Scholar] [CrossRef]
- Egea, J.; Buendia, I.; Parada, E.; Navarro, E.; Rada, P.; Cuadrado, A.; Lopez, M.G.; Garcia, A.G.; Leon, R. Melatonin-sulforaphane hybrid ITH12674 induces neuroprotection in oxidative stress conditions by a ‘drug-prodrug’ mechanism of action. Br. J. Pharmacol. 2015, 172, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Michalska, P.; Buendia, I.; Duarte, P.; FernandezMendivil, C.; Negredo, P.; Cuadrado, A.; Lopez, M.G.; Leon, R. Melatonin-sulforaphane hybrid ITH12674 attenuates glial response in vivo by blocking LPS binding to MD2 and receptor oligomerization. Pharmacol. Res. 2020, 152, 104597. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, R.L.; Ellis, C.E. Alzheimer’s disease and Parkinson’s disease. N. Engl. J. Med. 2003, 348, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Mi, K.; Johnson, G.V. The role of tau phosphorylation in the pathogenesis of Alzheimer’s disease. Curr. Alzheimer Res. 2006, 3, 449–463. [Google Scholar] [CrossRef]
- Nijholt, D.A.; van Haastert, E.S.; Rozemuller, A.J.; Scheper, W.; Hoozemans, J.J. The unfolded protein response is associated with early tau pathology in the hippocampus of tauopathies. J. Pathol. 2012, 226, 693–702. [Google Scholar] [CrossRef]
- Salminen, A.; Ojala, J.; Kauppinen, A.; Kaarniranta, K.; Suuronen, T. Inflammation in Alzheimer’s disease: Amyloid-beta oligomers trigger innate immunity defence via pattern recognition receptors. Prog. Neurobiol. 2009, 87, 181–194. [Google Scholar] [CrossRef]
- Bui, T.T.; Nguyen, T.H. Natural product for the treatment of Alzheimer’s disease. J. Basic Clin. Physiol. Pharmacol. 2017, 28, 413–423. [Google Scholar] [CrossRef]
- Vu, G.H.; Nguyen, H.D. Molecular mechanisms of sulforaphane in Alzheimer’s disease: Insights from an in-silico study. Silico Pharmacol. 2024, 12, 96. [Google Scholar] [CrossRef]
- Ibrahim, R.M.; El-Shiekh, R.A.; Mohamed, O.G.; Al-Karmalawy, A.A.; Tripathi, A.; Abdel-Baki, P.M. LC/MS-Based Metabolomics Reveals Chemical Variations of Two Broccoli Varieties in Relation to Their Anticholinesterase Activity: In vitro and In silico Studies. Plant Foods Hum. Nutr. 2024, 79, 359–366. [Google Scholar] [CrossRef]
- Nagaveni, V.; Lakshmi, V.V.; Prabhakar, S. Sulforaphane interaction with amyloid beta 1-40 peptide studied by electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2014, 28, 2171–2180. [Google Scholar] [CrossRef]
- Youn, K.; Yoon, J.H.; Lee, N.; Lim, G.; Lee, J.; Sang, S.; Ho, C.T.; Jun, M. Discovery of Sulforaphane as a Potent BACE1 Inhibitor Based on Kinetics and Computational Studies. Nutrients 2020, 12, 3026. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Jimenez-Moreno, N.; Garcia-Yague, A.J.; Escoll, M.; de Ceballos, M.L.; Van Leuven, F.; Rabano, A.; Yamamoto, M.; Rojo, A.I.; Cuadrado, A. Transcription factor NFE2L2/NRF2 is a regulator of macroautophagy genes. Autophagy 2016, 12, 1902–1916. [Google Scholar] [CrossRef] [PubMed]
- Villavicencio-Tejo, F.; Olesen, M.A.; Aranguiz, A.; Quintanilla, R.A. Activation of the Nrf2 Pathway Prevents Mitochondrial Dysfunction Induced by Caspase-3 Cleaved Tau: Implications for Alzheimer’s Disease. Antioxidants 2022, 11, 515. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhang, J.; Chang, N. Epigenetic modification of Nrf2 by sulforaphane increases the antioxidative and anti-inflammatory capacity in a cellular model of Alzheimer’s disease. Eur. J. Pharmacol. 2018, 824, 1–10. [Google Scholar] [CrossRef]
- Denzer, I.; Munch, G.; Pischetsrieder, M.; Friedland, K. S-allyl-L-cysteine and isoliquiritigenin improve mitochondrial function in cellular models of oxidative and nitrosative stress. Food Chem. 2016, 194, 843–848. [Google Scholar] [CrossRef]
- Chilakala, R.R.; Manchikalapudi, A.L.; Kumar, A.; Sunkaria, A. Sulforaphane Attenuates Abeta Oligomers Mediated Decrease in Phagocytic Activity of Microglial Cells. Neuroscience 2020, 429, 225–234. [Google Scholar] [CrossRef]
- Lee, S.; Choi, B.R.; Kim, J.; LaFerla, F.M.; Park, J.H.Y.; Han, J.S.; Lee, K.W.; Kim, J. Sulforaphane Upregulates the Heat Shock Protein Co-Chaperone CHIP and Clears Amyloid-beta and Tau in a Mouse Model of Alzheimer’s Disease. Mol. Nutr. Food Res. 2018, 62, e1800240. [Google Scholar] [CrossRef]
- Masci, A.; Mattioli, R.; Costantino, P.; Baima, S.; Morelli, G.; Punzi, P.; Giordano, C.; Pinto, A.; Donini, L.M.; d’Erme, M.; et al. Neuroprotective Effect of Brassica oleracea Sprouts Crude Juice in a Cellular Model of Alzheimer’s Disease. Oxidative Med. Cell. Longev. 2015, 2015, 781938. [Google Scholar] [CrossRef]
- Yang, C.; Qin, S.; Zhang, J.; Wang, Y.; Li, H.; Lu, T. Sulforaphane Upregulates Cultured Mouse Astrocytic Aquaporin-4 Expression through p38 MAPK Pathway. J. Healthc. Eng. 2022, 2022, 1144124. [Google Scholar] [CrossRef]
- Jhang, K.A.; Park, J.S.; Kim, H.S.; Chong, Y.H. Sulforaphane rescues amyloid-beta peptide-mediated decrease in MerTK expression through its anti-inflammatory effect in human THP-1 macrophages. J. Neuroinflamm. 2018, 15, 75. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, J.; Yang, C.; Dong, B.; Fu, Y.; Wang, Y.; Gong, M.; Liu, T.; Qiu, P.; Xie, W.; et al. Sulforaphane attenuates microglia-mediated neuronal damage by down-regulating the ROS/autophagy/NLRP3 signal axis in fibrillar Abeta-activated microglia. Brain Res. 2023, 1801, 148206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, C.; Zhang, S.; Cao, T.; Peng, R.; Guo, W.; Yan, Y.; Xie, S.; Peng, X.; Lu, T.; et al. Sulforaphane reverses Abeta fiber-mediated M1 type microglia polarization and neuroinflammation-mediated necroptosis of neural stem cells by downregulating the MAPK/NF-kappaB signaling pathways. Nan Fang Yi Ke Da Xue Xue Bao 2023, 43, 2132–2138. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, S.; Choi, B.R.; Yang, H.; Hwang, Y.; Park, J.H.; LaFerla, F.M.; Han, J.S.; Lee, K.W.; Kim, J. Sulforaphane epigenetically enhances neuronal BDNF expression and TrkB signaling pathways. Mol. Nutr. Food Res. 2017, 61, 1600194. [Google Scholar] [CrossRef]
- Angeloni, C.; Malaguti, M.; Rizzo, B.; Barbalace, M.C.; Fabbri, D.; Hrelia, S. Neuroprotective effect of sulforaphane against methylglyoxal cytotoxicity. Chem. Res. Toxicol. 2015, 28, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.T.; Yang, H.Y.; Wang, W.; Wu, Q.Q.; Tian, Y.R.; Jia, J.P. Sulforaphane Inhibits the Generation of Amyloid-beta Oligomer and Promotes Spatial Learning and Memory in Alzheimer’s Disease (PS1V97L) Transgenic Mice. J. Alzheimers Dis. 2018, 62, 1803–1813. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, R.; Zhan, Z.; Li, X.; Zhou, F.; Xing, A.; Jiang, C.; Chen, Y.; An, L. Beneficial Effects of Sulforaphane Treatment in Alzheimer’s Disease May Be Mediated through Reduced HDAC1/3 and Increased P75NTR Expression. Front. Aging Neurosci. 2017, 9, 121. [Google Scholar] [CrossRef]
- Zhang, R.; Miao, Q.W.; Zhu, C.X.; Zhao, Y.; Liu, L.; Yang, J.; An, L. Sulforaphane ameliorates neurobehavioral deficits and protects the brain from amyloid beta deposits and peroxidation in mice with Alzheimer-like lesions. Am. J. Alzheimers Dis. Other Demen. 2015, 30, 183–191. [Google Scholar] [CrossRef]
- Yang, W.; Liu, Y.; Xu, Q.Q.; Xian, Y.F.; Lin, Z.X. Sulforaphene Ameliorates Neuroinflammation and Hyperphosphorylated Tau Protein via Regulating the PI3K/Akt/GSK-3beta Pathway in Experimental Models of Alzheimer’s Disease. Oxidative Med. Cell. Longev. 2020, 2020, 4754195. [Google Scholar] [CrossRef]
- An, Y.W.; Jhang, K.A.; Woo, S.Y.; Kang, J.L.; Chong, Y.H. Sulforaphane exerts its anti-inflammatory effect against amyloid-beta peptide via STAT-1 dephosphorylation and activation of Nrf2/HO-1 cascade in human THP-1 macrophages. Neurobiol. Aging 2016, 38, 1–10. [Google Scholar] [CrossRef]
- Khan, W.U.; Salman, M.; Ali, M.; Majid, H.; Yar, M.S.; Akhtar, M.; Parvez, S.; Najmi, A.K. Neuroprotective Effects of Sulforaphane in a rat model of Alzheimer’s Disease induced by Abeta (1-42) peptides. Neurochem. Int. 2024, 179, 105839. [Google Scholar] [CrossRef]
- Pu, D.; Zhao, Y.; Chen, J.; Sun, Y.; Lv, A.; Zhu, S.; Luo, C.; Zhao, K.; Xiao, Q. Protective Effects of Sulforaphane on Cognitive Impairments and AD-like Lesions in Diabetic Mice are Associated with the Upregulation of Nrf2 Transcription Activity. Neuroscience 2018, 381, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, L.; Li, X.; Hu, Q.; Mao, L.; Shao, Y.; Han, M.; Zhang, S.; Ejaz, I.; Mesbah, L.; et al. Sulforaphane suppresses Abeta accumulation and tau hyperphosphorylation in vascular cognitive impairment(VCI). J. Nutr. Biochem. 2025, 136, 109803. [Google Scholar] [CrossRef] [PubMed]
- Minuti, A.; Mazzon, E.; Iori, R.; Chiricosta, L.; Artimagnella, O. Bioactivated Glucoraphanin Improves Cell Survival, Upregulating Phospho-AKT, and Modulates Genes Involved in DNA Repair in an In Vitro Alzheimer’s Disease Model: A Network-Transcriptomic Analysis. Nutrients 2024, 16, 4202. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, J.; Fang, L.; Li, X.; Zhao, Y.; Shi, W.; An, L. Neuroprotective effects of sulforaphane on cholinergic neurons in mice with Alzheimer’s disease-like lesions. Int. J. Mol. Sci. 2014, 15, 14396–14410. [Google Scholar] [CrossRef]
- Wang, W.; Wei, C.; Quan, M.; Li, T.; Jia, J. Sulforaphane Reverses the Amyloid-beta Oligomers Induced Depressive-like Behavior. J. Alzheimers Dis. 2020, 78, 127–137. [Google Scholar] [CrossRef]
- Zhao, J.; Bi, W.; Xiao, S.; Lan, X.; Cheng, X.; Zhang, J.; Lu, D.; Wei, W.; Wang, Y.; Li, H.; et al. Neuroinflammation induced by lipopolysaccharide causes cognitive impairment in mice. Sci. Rep. 2019, 9, 5790. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, Y.K.; Yuk, D.Y.; Choi, D.Y.; Ban, S.B.; Oh, K.W.; Hong, J.T. Neuro-inflammation induced by lipopolysaccharide causes cognitive impairment through enhancement of beta-amyloid generation. J. Neuroinflamm. 2008, 5, 37. [Google Scholar] [CrossRef]
- Alzahrani, N.A.; Bahaidrah, K.A.; Mansouri, R.A.; Aldhahri, R.S.; Abd El-Aziz, G.S.; Alghamdi, B.S. Possible Prophylactic Effects of Sulforaphane on LPS-Induced Recognition Memory Impairment Mediated by Regulating Oxidative Stress and Neuroinflammatory Proteins in the Prefrontal Cortex Region of the Brain. Biomedicines 2024, 12, 1107. [Google Scholar] [CrossRef]
- Park, H.S.; Hwang, E.S.; Choi, G.Y.; Kim, H.B.; Park, K.S.; Sul, J.Y.; Hwang, Y.; Choi, G.W.; Kim, B.I.; Park, H.; et al. Sulforaphane enhances long-term potentiation and ameliorate scopolamine-induced memory impairment. Physiol. Behav. 2021, 238, 113467. [Google Scholar] [CrossRef]
- Rajesh, V.; Ilanthalir, S. Cognition Enhancing Activity of Sulforaphane Against Scopolamine Induced Cognitive Impairment in Zebra Fish (Danio rerio). Neurochem. Res. 2016, 41, 2538–2548. [Google Scholar] [CrossRef]
- Holloway, P.M.; Gillespie, S.; Becker, F.; Vital, S.A.; Nguyen, V.; Alexander, J.S.; Evans, P.C.; Gavins, F.N.E. Sulforaphane induces neurovascular protection against a systemic inflammatory challenge via both Nrf2-dependent and independent pathways. Vasc. Pharmacol. 2016, 85, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Hortal, M.D.; Romero-Marquez, J.M.; Lopez-Bascon, M.A.; Sanchez-Gonzalez, C.; Xiao, J.; Sumalla-Cano, S.; Battino, M.; Forbes-Hernandez, T.Y.; Quiles, J.L. In Vitro and In Vivo Insights into a Broccoli Byproduct as a Healthy Ingredient for the Management of Alzheimer’s Disease and Aging through Redox Biology. J. Agric. Food Chem. 2024, 72, 5197–5211. [Google Scholar] [CrossRef] [PubMed]
- Carnicero-Senabre, D.; Jimenez-Villegas, J.; Alvarez-Garrote, S.; Escoll, M.; Cuadrado, A.; Rojo, A.I. NRF2 activation by 6-MSITC increases the generation of neuroprotective, soluble alpha amyloid precursor protein by inducing the metalloprotease gene ADAM17. Free Radic. Biol. Med. 2025, 227, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Uruno, A.; Matsumaru, D.; Ryoke, R.; Saito, R.; Kadoguchi, S.; Saigusa, D.; Saito, T.; Saido, T.C.; Kawashima, R.; Yamamoto, M. Nrf2 Suppresses Oxidative Stress and Inflammation in App Knock-In Alzheimer’s Disease Model Mice. Mol. Cell. Biol. 2020, 40, e00467-19. [Google Scholar] [CrossRef]
- Morroni, F.; Sita, G.; Graziosi, A.; Turrini, E.; Fimognari, C.; Tarozzi, A.; Hrelia, P. Protective Effects of 6-(Methylsulfinyl)hexyl Isothiocyanate on Abeta(1-42)-Induced Cognitive Deficit, Oxidative Stress, Inflammation, and Apoptosis in Mice. Int. J. Mol. Sci. 2018, 19, 2083. [Google Scholar] [CrossRef]
- Azlan, U.K.; Khairul Annuar, N.A.; Mediani, A.; Aizat, W.M.; Damanhuri, H.A.; Tong, X.; Yanagisawa, D.; Tooyama, I.; Wan Ngah, W.Z.; Jantan, I.; et al. An insight into the neuroprotective and anti-neuroinflammatory effects and mechanisms of Moringa oleifera. Front. Pharmacol. 2022, 13, 1035220. [Google Scholar] [CrossRef]
- Silvestro, S.; Chiricosta, L.; Gugliandolo, A.; Iori, R.; Rollin, P.; Perenzoni, D.; Mattivi, F.; Bramanti, P.; Mazzon, E. The Moringin/alpha-CD Pretreatment Induces Neuroprotection in an In Vitro Model of Alzheimer’s Disease: A Transcriptomic Study. Curr. Issues Mol. Biol. 2021, 43, 197–214. [Google Scholar] [CrossRef]
- Chiricosta, L.; Gugliandolo, A.; Diomede, F.; Pizzicannella, J.; Trubiani, O.; Iori, R.; Tardiolo, G.; Guarnieri, S.; Bramanti, P.; Mazzon, E. Moringin Pretreatment Inhibits the Expression of Genes Involved in Mitophagy in the Stem Cell of the Human Periodontal Ligament. Molecules 2019, 24, 3217. [Google Scholar] [CrossRef]
- Sestito, S.; Daniele, S.; Pietrobono, D.; Citi, V.; Bellusci, L.; Chiellini, G.; Calderone, V.; Martini, C.; Rapposelli, S. Memantine prodrug as a new agent for Alzheimer’s Disease. Sci. Rep. 2019, 9, 4612. [Google Scholar] [CrossRef]
- Sestito, S.; Pruccoli, L.; Runfola, M.; Citi, V.; Martelli, A.; Saccomanni, G.; Calderone, V.; Tarozzi, A.; Rapposelli, S. Design and synthesis of H(2)S-donor hybrids: A new treatment for Alzheimer’s disease? Eur. J. Med. Chem. 2019, 184, 111745. [Google Scholar] [CrossRef]
- Francardo, V.; Recchia, A.; Popovic, N.; Andersson, D.; Nissbrandt, H.; Cenci, M.A. Impact of the lesion procedure on the profiles of motor impairment and molecular responsiveness to L-DOPA in the 6-hydroxydopamine mouse model of Parkinson’s disease. Neurobiol. Dis. 2011, 42, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.P.; Duda, J.E. Dietary modifications in Parkinson’s disease: A neuroprotective intervention? Med. Hypotheses 2015, 85, 1002–1005. [Google Scholar] [CrossRef] [PubMed]
- Vega-Galvez, A.; Gomez-Perez, L.S.; Zepeda, F.; Vidal, R.L.; Grunenwald, F.; Mejias, N.; Pasten, A.; Araya, M.; Ah-Hen, K.S. Assessment of Bio-Compounds Content, Antioxidant Activity, and Neuroprotective Effect of Red Cabbage (Brassica oleracea var. Capitata rubra) Processed by Convective Drying at Different Temperatures. Antioxidants 2023, 12, 1789. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Wang, X.; Gong, L.; Zhai, X.; Wang, K.; Qiu, X.; Zhang, H.; Tang, Z.; Jiang, H.; Wang, X. Screening of H(2)S donors with a red emission mitochondria-targetable fluorescent probe: Toward discovering a new therapeutic strategy for Parkinson’s disease. Biosens. Bioelectron. 2023, 237, 115521. [Google Scholar] [CrossRef]
- Bao, B.; Zhang, M.Q.; Chen, Z.Y.; Wu, X.B.; Xia, Z.B.; Chai, J.Y.; Yin, X.P. Sulforaphane prevents PC12 cells from oxidative damage via the Nrf2 pathway. Mol. Med. Rep. 2019, 19, 4890–4896. [Google Scholar] [CrossRef]
- Izumi, Y.; Kataoka, H.; Inose, Y.; Akaike, A.; Koyama, Y.; Kume, T. Neuroprotective effect of an Nrf2-ARE activator identified from a chemical library on dopaminergic neurons. Eur. J. Pharmacol. 2018, 818, 470–479. [Google Scholar] [CrossRef]
- Pu, Y.; Qu, Y.; Chang, L.; Wang, S.M.; Zhang, K.; Ushida, Y.; Suganuma, H.; Hashimoto, K. Dietary intake of glucoraphanin prevents the reduction of dopamine transporter in the mouse striatum after repeated administration of MPTP. Neuropsychopharmacol. Rep. 2019, 39, 247–251. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, B.; Wang, X.; Wu, L.; Yang, Y.; Cheng, X.; Hu, Z.; Cai, X.; Yang, J.; Sun, X.; et al. Sulforaphane protects against rotenone-induced neurotoxicity in vivo: Involvement of the mTOR, Nrf2, and autophagy pathways. Sci. Rep. 2016, 6, 32206. [Google Scholar] [CrossRef]
- Lin, Z.; Huang, L.; Cao, Q.; Luo, H.; Yao, W.; Zhang, J.C. Inhibition of abnormal C/EBPbeta/alpha-Syn signaling pathway through activation of Nrf2 ameliorates Parkinson’s disease-like pathology. Aging Cell 2023, 22, e13958. [Google Scholar] [CrossRef]
- Sarkar, A.; Singh, M.P. A Complex Interplay of DJ-1, LRRK2, and Nrf2 in the Regulation of Mitochondrial Function in Cypermethrin-Induced Parkinsonism. Mol. Neurobiol. 2024, 61, 953–970. [Google Scholar] [CrossRef]
- Asuncion, R.M.D.; Ramani, P.K. Rett Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Renieri, A.; Mari, F.; Mencarelli, M.A.; Scala, E.; Ariani, F.; Longo, I.; Meloni, I.; Cevenini, G.; Pini, G.; Hayek, G.; et al. Diagnostic criteria for the Zappella variant of Rett syndrome (the preserved speech variant). Brain Dev. 2009, 31, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Zou, Q.; Zhao, X.; Zhang, Y.; Qu, Y.; Wang, N.; Murayama, S.; Qi, Q.; Hashimoto, K.; Lin, S.; et al. Regulation of BDNF transcription by Nrf2 and MeCP2 ameliorates MPTP-induced neurotoxicity. Cell Death Discov. 2022, 8, 267. [Google Scholar] [CrossRef] [PubMed]
- Platz, S.; Piberger, A.L.; Budnowski, J.; Herz, C.; Schreiner, M.; Blaut, M.; Hartwig, A.; Lamy, E.; Hanske, L.; Rohn, S. Bioavailability and biotransformation of sulforaphane and erucin metabolites in different biological matrices determined by LC-MS-MS. Anal. Bioanal. Chem. 2015, 407, 1819–1829. [Google Scholar] [CrossRef]
- Morroni, F.; Sita, G.; Djemil, A.; D’Amico, M.; Pruccoli, L.; Cantelli-Forti, G.; Hrelia, P.; Tarozzi, A. Comparison of Adaptive Neuroprotective Mechanisms of Sulforaphane and its Interconversion Product Erucin in in Vitro and in Vivo Models of Parkinson’s Disease. J. Agric. Food Chem. 2018, 66, 856–865. [Google Scholar] [CrossRef]
- Balducci, M.; Perez, J.T.; Del Rio, C.T.; Perez, M.C.; Carranza, A.D.V.; Gomez Escribano, A.P.; Vazquez-Manrique, R.P.; Tarozzi, A. Erucin, a Natural Isothiocyanate, Prevents Polyglutamine-Induced Toxicity in Caenorhabditis elegans via aak-2/AMPK and daf-16/FOXO Signaling. Int. J. Mol. Sci. 2024, 25, 12220. [Google Scholar] [CrossRef]
- Giacoppo, S.; Rajan, T.S.; De Nicola, G.R.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. The Isothiocyanate Isolated from Moringa oleifera Shows Potent Anti-Inflammatory Activity in the Treatment of Murine Subacute Parkinson’s Disease. Rejuvenation Res. 2017, 20, 50–63. [Google Scholar] [CrossRef]
- Lee, J.A.; Son, H.J.; Park, K.D.; Han, S.H.; Shin, N.; Kim, J.H.; Kim, H.R.; Kim, D.J.; Hwang, O. A Novel Compound ITC-3 Activates the Nrf2 Signaling and Provides Neuroprotection in Parkinson’s Disease Models. Neurotox. Res. 2015, 28, 332–345. [Google Scholar] [CrossRef]
- Lee, J.A.; Son, H.J.; Kim, J.H.; Park, K.D.; Shin, N.; Kim, H.R.; Kim, E.M.; Kim, D.J.; Hwang, O. A novel synthetic isothiocyanate ITC-57 displays antioxidant, anti-inflammatory, and neuroprotective properties in a mouse Parkinson’s disease model. Free Radic. Res. 2016, 50, 1188–1199. [Google Scholar] [CrossRef]
- Chauhan, A.K.; Mittra, N.; Singh, B.K.; Singh, C. Inhibition of glutathione S-transferase-pi triggers c-jun N-terminal kinase-dependent neuronal death in Zn-induced Parkinsonism. Mol. Cell. Biochem. 2019, 452, 95–104. [Google Scholar] [CrossRef]
- Morroni, F.; Sita, G.; Tarozzi, A.; Cantelli-Forti, G.; Hrelia, P. Neuroprotection by 6-(methylsulfinyl)hexyl isothiocyanate in a 6-hydroxydopamine mouse model of Parkinson’s disease. Brain Res. 2014, 1589, 93–104. [Google Scholar] [CrossRef]
- Wright, A.F. The Redox Stress Test: A novel technique reveals oxidative stress in Parkinson’s disease. Med. Res. Arch. 2024, 12, 4955. [Google Scholar] [CrossRef]
- Reich, D.S.; Lucchinetti, C.F.; Calabresi, P.A. Multiple Sclerosis. N. Engl. J. Med. 2018, 378, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.; Chartier, G.; Quandt, J. Modulating inflammation and neuroprotection in multiple sclerosis. J. Neurosci. Res. 2018, 96, 927–950. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, B.; Ignarro, L.J.; Montestruque, S.; Smoll, A.; Merrill, J.E. Nitric oxide as a potential pathological mechanism in demyelination: Its differential effects on primary glial cells in vitro. Neuroscience 1994, 61, 575–585. [Google Scholar] [CrossRef]
- Lim, J.L.; van der Pol, S.M.; Baron, W.; McCord, J.M.; de Vries, H.E.; van Horssen, J. Protandim Protects Oligodendrocytes against an Oxidative Insult. Antioxidants 2016, 5, 30. [Google Scholar] [CrossRef]
- Spencer, E.S.; Dale, E.J.; Gommans, A.L.; Rutledge, M.T.; Vo, C.T.; Nakatani, Y.; Gamble, A.B.; Smith, R.A.; Wilbanks, S.M.; Hampton, M.B.; et al. Multiple binding modes of isothiocyanates that inhibit macrophage migration inhibitory factor. Eur. J. Med. Chem. 2015, 93, 501–510. [Google Scholar] [CrossRef]
- Ellwardt, E.; Zipp, F. Molecular mechanisms linking neuroinflammation and neurodegeneration in MS. Exp. Neurol. 2014, 262 Pt A, 8–17. [Google Scholar] [CrossRef]
- Yoo, I.H.; Kim, M.J.; Kim, J.; Sung, J.J.; Park, S.T.; Ahn, S.W. The Anti-Inflammatory Effect of Sulforaphane in Mice with Experimental Autoimmune Encephalomyelitis. J. Korean Med. Sci 2019, 34, e197. [Google Scholar] [CrossRef]
- Giacoppo, S.; Galuppo, M.; Iori, R.; De Nicola, G.R.; Bramanti, P.; Mazzon, E. The protective effects of bioactive (RS)-glucoraphanin on the permeability of the mice blood-brain barrier following experimental autoimmune encephalomyelitis. Eur. Rev. Med. Pharmacol Sci 2014, 18, 194–204. [Google Scholar]
- Galea, I.; Copple, I.M.; Howat, D.W.; Franklin, S. SFX-01 reduces residual disability after experimental autoimmune encephalomyelitis. Mult. Scler. Relat. Disord. 2019, 30, 257–261. [Google Scholar] [CrossRef]
- Galuppo, M.; Giacoppo, S.; De Nicola, G.R.; Iori, R.; Navarra, M.; Lombardo, G.E.; Bramanti, P.; Mazzon, E. Antiinflammatory activity of glucomoringin isothiocyanate in a mouse model of experimental autoimmune encephalomyelitis. Fitoterapia 2014, 95, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Soundara Rajan, T.; De Nicola, G.R.; Iori, R.; Bramanti, P.; Mazzon, E. Moringin activates Wnt canonical pathway by inhibiting GSK3beta in a mouse model of experimental autoimmune encephalomyelitis. Drug Des. Devel. Ther. 2016, 10, 3291–3304. [Google Scholar] [CrossRef] [PubMed]
- Giacoppo, S.; Iori, R.; Bramanti, P.; Mazzon, E. Topical moringin-cream relieves neuropathic pain by suppression of inflammatory pathway and voltage-gated ion channels in murine model of multiple sclerosis. Mol. Pain 2017, 13, 1744806917724318. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Gao, Q.; Zhao, P.; Gao, Y.; Xi, Y.; Wang, X.; Liang, Y.; Shi, H.; Ma, Y. Sulforaphane produces antidepressant- and anxiolytic-like effects in adult mice. Behav. Brain Res. 2016, 301, 55–62. [Google Scholar] [CrossRef]
- Panjwani, A.A.; Liu, H.; Fahey, J.W. Crucifers and related vegetables and supplements for neurologic disorders: What is the evidence? Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 451–457. [Google Scholar] [CrossRef]
- Hodges, H.; Fealko, C.; Soares, N. Autism spectrum disorder: Definition, epidemiology, causes, and clinical evaluation. Transl. Pediatr. 2020, 9, S55–S65. [Google Scholar] [CrossRef]
- Maenner, M.J.; Warren, Z.; Williams, A.R.; Amoakohene, E.; Bakian, A.V.; Bilder, D.A.; Durkin, M.S.; Fitzgerald, R.T.; Furnier, S.M.; Hughes, M.M.; et al. Prevalence and Characteristics of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2020. MMWR Surveill. Summ. 2023, 72, 1–14. [Google Scholar] [CrossRef]
- Liu, H.; Talalay, P.; Fahey, J.W. Biomarker-Guided Strategy for Treatment of Autism Spectrum Disorder (ASD). CNS Neurol. Disord. Drug Targets 2016, 15, 602–613. [Google Scholar] [CrossRef]
- Alshehri, S.; Ahmad, S.F.; Albekairi, N.A.; Alqarni, S.S.; Al-Harbi, N.O.; Al-Ayadhi, L.Y.; Attia, S.M.; Alfardan, A.S.; Bakheet, S.A.; Nadeem, A. Thioredoxin 1 and Thioredoxin Reductase 1 Redox System Is Dysregulated in Neutrophils of Subjects with Autism: In Vitro Effects of Environmental Toxicant, Methylmercury. Toxics 2023, 11, 739. [Google Scholar] [CrossRef]
- Shahlaei, M.; Saeidifar, M.; Zamanian, A. Sustained release of sulforaphane by bioactive extracellular vesicles for neuroprotective effect on chick model. J. Biomed. Mater. Res. B Appl. Biomater. 2022, 110, 2636–2648. [Google Scholar] [CrossRef]
- Liu, H.; Zimmerman, A.W.; Singh, K.; Connors, S.L.; Diggins, E.; Stephenson, K.K.; Dinkova-Kostova, A.T.; Fahey, J.W. Biomarker Exploration in Human Peripheral Blood Mononuclear Cells for Monitoring Sulforaphane Treatment Responses in Autism Spectrum Disorder. Sci. Rep. 2020, 10, 5822. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, A.; Ahmad, S.F.; Al-Ayadhi, L.Y.; Attia, S.M.; Al-Harbi, N.O.; Alzahrani, K.S.; Bakheet, S.A. Differential regulation of Nrf2 is linked to elevated inflammation and nitrative stress in monocytes of children with autism. Psychoneuroendocrinology 2020, 113, 104554. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, A.; Ahmad, S.F.; Al-Harbi, N.O.; Attia, S.M.; Bakheet, S.A.; Ibrahim, K.E.; Alqahtani, F.; Alqinyah, M. Nrf2 activator, sulforaphane ameliorates autism-like symptoms through suppression of Th17 related signaling and rectification of oxidant-antioxidant imbalance in periphery and brain of BTBR T+tf/J mice. Behav. Brain Res. 2019, 364, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; He, L.; Dai, S.; Zheng, H.; Cui, X.; Ou, J.; Zhang, X. Therapeutic efficacy of sulforaphane in autism spectrum disorders and its association with gut microbiota: Animal model and human longitudinal studies. Front. Nutr. 2023, 10, 1294057. [Google Scholar] [CrossRef]
- Fujita, Y.; Fujita, A.; Ishima, T.; Hirai, A.; Suzuki, S.; Suganuma, H.; Hashimoto, K. Dietary intake of glucoraphanin during pregnancy and lactation prevents the behavioral abnormalities in the offspring after maternal immune activation. Neuropsychopharmacol. Rep. 2020, 40, 268–274. [Google Scholar] [CrossRef]
- Singh, K.; Connors, S.L.; Macklin, E.A.; Smith, K.D.; Fahey, J.W.; Talalay, P.; Zimmerman, A.W. Sulforaphane treatment of autism spectrum disorder (ASD). Proc. Natl. Acad. Sci. USA 2014, 111, 15550–15555. [Google Scholar] [CrossRef]
- Lynch, R.; Diggins, E.L.; Connors, S.L.; Zimmerman, A.W.; Singh, K.; Liu, H.; Talalay, P.; Fahey, J.W. Sulforaphane from Broccoli Reduces Symptoms of Autism: A Follow-up Case Series from a Randomized Double-blind Study. Glob. Adv. Health Med. 2017, 6, 2164957X17735826. [Google Scholar] [CrossRef]
- Bent, S.; Lawton, B.; Warren, T.; Widjaja, F.; Dang, K.; Fahey, J.W.; Cornblatt, B.; Kinchen, J.M.; Delucchi, K.; Hendren, R.L. Identification of urinary metabolites that correlate with clinical improvements in children with autism treated with sulforaphane from broccoli. Mol. Autism 2018, 9, 35. [Google Scholar] [CrossRef]
- Momtazmanesh, S.; Amirimoghaddam-Yazdi, Z.; Moghaddam, H.S.; Mohammadi, M.R.; Akhondzadeh, S. Sulforaphane as an adjunctive treatment for irritability in children with autism spectrum disorder: A randomized, double-blind, placebo-controlled clinical trial. Psychiatry Clin. Neurosci. 2020, 74, 398–405. [Google Scholar] [CrossRef]
- Zimmerman, A.W.; Singh, K.; Connors, S.L.; Liu, H.; Panjwani, A.A.; Lee, L.C.; Diggins, E.; Foley, A.; Melnyk, S.; Singh, I.N.; et al. Randomized controlled trial of sulforaphane and metabolite discovery in children with Autism Spectrum Disorder. Mol. Autism 2021, 12, 38. [Google Scholar] [CrossRef]
- Magner, M.; Thorova, K.; Zupova, V.; Houska, M.; Svandova, I.; Novotna, P.; Triska, J.; Vrchotova, N.; Soural, I.; Jilek, L. Sulforaphane Treatment in Children with Autism: A Prospective Randomized Double-Blind Study. Nutrients 2023, 15, 718. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Smith, R.C.; Tobe, R.H.; Lin, J.; Arriaza, J.; Fahey, J.W.; Liu, R.; Zeng, Y.; Liu, Y.; Huang, L.; et al. Efficacy of Sulforaphane in Treatment of Children with Autism Spectrum Disorder: A Randomized Double-Blind Placebo-Controlled Multi-center Trial. J. Autism Dev. Disord. 2024, 54, 628–641. [Google Scholar] [CrossRef] [PubMed]
- Meral, G.; Aslan, E.S.; Burkay, N.; Alper Acar, E.G.; Karagoz, M.F.; Ozkaya, M.; Sahin, E.; Alp, M.Y. Importance of Using Epigenetic Nutrition and Supplements Based on Nutrigenetic Tests in Personalized Medicine. Cureus 2024, 16, e66959. [Google Scholar] [CrossRef] [PubMed]
- Buck, B.E.; Healey, K.M.; Gagen, E.C.; Roberts, D.L.; Penn, D.L. Social cognition in schizophrenia: Factor structure, clinical and functional correlates. J. Ment. Health 2016, 25, 330–337. [Google Scholar] [CrossRef]
- Kahn, R.S.; Keefe, R.S. Schizophrenia is a cognitive illness: Time for a change in focus. JAMA Psychiatry 2013, 70, 1107–1112. [Google Scholar] [CrossRef]
- Hashimoto, K. Targeting of NMDA receptors in new treatments for schizophrenia. Expert Opin. Ther. Targets 2014, 18, 1049–1063. [Google Scholar] [CrossRef]
- Ladermann, A.; Edwards, T.B.; Walch, G. Arm lengthening after reverse shoulder arthroplasty: A review. Int. Orthop. 2014, 38, 991–1000. [Google Scholar] [CrossRef]
- Koskuvi, M.; Porsti, E.; Hewitt, T.; Rasanen, N.; Wu, Y.C.; Trontti, K.; McQuade, A.; Kalyanaraman, S.; Ojansuu, I.; Vaurio, O.; et al. Genetic contribution to microglial activation in schizophrenia. Mol. Psychiatry 2024, 29, 2622–2633. [Google Scholar] [CrossRef]
- Shirai, Y.; Fujita, Y.; Hashimoto, R.; Ohi, K.; Yamamori, H.; Yasuda, Y.; Ishima, T.; Suganuma, H.; Ushida, Y.; Takeda, M.; et al. Dietary Intake of Sulforaphane-Rich Broccoli Sprout Extracts during Juvenile and Adolescence Can Prevent Phencyclidine-Induced Cognitive Deficits at Adulthood. PLoS ONE 2015, 10, e0127244. [Google Scholar] [CrossRef]
- Matsuura, A.; Ishima, T.; Fujita, Y.; Iwayama, Y.; Hasegawa, S.; Kawahara-Miki, R.; Maekawa, M.; Toyoshima, M.; Ushida, Y.; Suganuma, H.; et al. Dietary glucoraphanin prevents the onset of psychosis in the adult offspring after maternal immune activation. Sci. Rep. 2018, 8, 2158. [Google Scholar] [CrossRef]
- Isaacson, R.H.; Beier, J.I.; Khoo, N.K.; Freeman, B.A.; Freyberg, Z.; Arteel, G.E. Olanzapine-induced liver injury in mice: Aggravation by high-fat diet and protection with sulforaphane. J. Nutr. Biochem. 2020, 81, 108399. [Google Scholar] [CrossRef] [PubMed]
- Kaur Sodhi, R.; Kumar, H.; Singh, R.; Bansal, Y.; Singh, Y.; Kiran Kondepudi, K.; Bishnoi, M.; Kuhad, A. Allyl isothiocyanate, a TRPA1 agonist, protects against olanzapine-induced hypothalamic and hepatic metabolic aberrations in female mice. Biochem. Pharmacol. 2024, 222, 116074. [Google Scholar] [CrossRef] [PubMed]
- El-Shoura, E.A.M.; Abdelzaher, L.A.; Mahmoud, N.I.; Farghaly, O.A.; Sabry, M.; Girgis Shahataa, M.; Salem, E.A.; Saad, H.M.; Elhussieny, O.; Kozman, M.R.; et al. Combined sulforaphane and beta-sitosterol mitigate olanzapine-induced metabolic disorders in rats: Insights on FOXO, PI3K/AKT, JAK/STAT3, and MAPK signaling pathways. Int. Immunopharmacol. 2024, 140, 112904. [Google Scholar] [CrossRef]
- Shiina, A.; Kanahara, N.; Sasaki, T.; Oda, Y.; Hashimoto, T.; Hasegawa, T.; Yoshida, T.; Iyo, M.; Hashimoto, K. An Open Study of Sulforaphane-rich Broccoli Sprout Extract in Patients with Schizophrenia. Clin. Psychopharmacol. Neurosci. 2015, 13, 62–67. [Google Scholar] [CrossRef]
- Dickerson, F.; Origoni, A.; Katsafanas, E.; Squire, A.; Newman, T.; Fahey, J.; Xiao, J.C.; Stallings, C.; Goga, J.; Khushalani, S.; et al. Randomized controlled trial of an adjunctive sulforaphane nutraceutical in schizophrenia. Schizophr. Res. 2021, 231, 142–144. [Google Scholar] [CrossRef]
- Hei, G.; Smith, R.C.; Li, R.; Ou, J.; Song, X.; Zheng, Y.; He, Y.; Arriaza, J.; Fahey, J.W.; Cornblatt, B.; et al. Sulforaphane Effects on Cognition and Symptoms in First and Early Episode Schizophrenia: A Randomized Double-Blind Trial. Schizophr. Bull. Open 2022, 3, sgac024. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, W.; Lu, X.; Zhou, H.; Huang, J.; Xu, Z.; Liao, H.; Liang, J.; Liang, M.; Ye, C.; et al. The association of SOD and HsCRP with the efficacy of sulforaphane in schizophrenia patients with residual negative symptoms. Eur. Arch. Psychiatry Clin. Neurosci. 2024, 274, 1083–1092. [Google Scholar] [CrossRef]
- Maletic, V.; Raison, C.L. Neurobiology of depression, fibromyalgia and neuropathic pain. Front. Biosci. 2009, 14, 5291–5338. [Google Scholar] [CrossRef]
- Attal, N.; Lanteri-Minet, M.; Laurent, B.; Fermanian, J.; Bouhassira, D. The specific disease burden of neuropathic pain: Results of a French nationwide survey. Pain 2011, 152, 2836–2843. [Google Scholar] [CrossRef]
- Stachowicz, K.; Sowa-Kucma, M. The treatment of depression—Searching for new ideas. Front. Pharmacol. 2022, 13, 988648. [Google Scholar] [CrossRef]
- Whiteford, H.A.; Degenhardt, L.; Rehm, J.; Baxter, A.J.; Ferrari, A.J.; Erskine, H.E.; Charlson, F.J.; Norman, R.E.; Flaxman, A.D.; Johns, N.; et al. Global burden of disease attributable to mental and substance use disorders: Findings from the Global Burden of Disease Study 2010. Lancet 2013, 382, 1575–1586. [Google Scholar] [CrossRef] [PubMed]
- Walker, E.R.; McGee, R.E.; Druss, B.G. Mortality in mental disorders and global disease burden implications: A systematic review and meta-analysis. JAMA Psychiatry 2015, 72, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Raison, C.L.; Miller, A.H. Pathogen-Host Defense in the Evolution of Depression: Insights into Epidemiology, Genetics, Bioregional Differences and Female Preponderance. Neuropsychopharmacology 2017, 42, 5–27. [Google Scholar] [CrossRef] [PubMed]
- Juruena, M.F.; Eror, F.; Cleare, A.J.; Young, A.H. The Role of Early Life Stress in HPA Axis and Anxiety. Adv. Exp. Med. Biol. 2020, 1191, 141–153. [Google Scholar] [CrossRef]
- Gautam, S.; Jain, A.; Gautam, M.; Vahia, V.N.; Grover, S. Clinical Practice Guidelines for the management of Depression. Indian J. Psychiatry 2017, 59, S34–S50. [Google Scholar] [CrossRef]
- Sackeim, H.A. The definition and meaning of treatment-resistant depression. J. Clin. Psychiatry 2001, 62 (Suppl. S16), 10–17. [Google Scholar]
- Bigger, J.T.; Glassman, A.H. The American Heart Association science advisory on depression and coronary heart disease: An exploration of the issues raised. Cleve. Clin. J. Med. 2010, 77 (Suppl. S3), S12–S19. [Google Scholar] [CrossRef]
- Ferguson, J.M. SSRI Antidepressant Medications: Adverse Effects and Tolerability. Prim. Care Companion J. Clin. Psychiatry 2001, 3, 22–27. [Google Scholar] [CrossRef]
- Li, Y.; Lv, M.R.; Wei, Y.J.; Sun, L.; Zhang, J.X.; Zhang, H.G.; Li, B. Dietary patterns and depression risk: A meta-analysis. Psychiatry Res. 2017, 253, 373–382. [Google Scholar] [CrossRef]
- Yao, W.; Zhang, J.C.; Ishima, T.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Wu, J.; Suganuma, H.; et al. Role of Keap1-Nrf2 signaling in depression and dietary intake of glucoraphanin confers stress resilience in mice. Sci. Rep. 2016, 6, 30659. [Google Scholar] [CrossRef]
- Zhang, J.C.; Yao, W.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Wu, J.; Ushida, Y.; Suganuma, H.; et al. Prophylactic effects of sulforaphane on depression-like behavior and dendritic changes in mice after inflammation. J. Nutr. Biochem. 2017, 39, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Chamorro, P.; Redondo, A.; Riego, G.; Leanez, S.; Pol, O. Sulforaphane Inhibited the Nociceptive Responses, Anxiety- and Depressive-Like Behaviors Associated With Neuropathic Pain and Improved the Anti-allodynic Effects of Morphine in Mice. Front. Pharmacol. 2018, 9, 1332. [Google Scholar] [CrossRef] [PubMed]
- Tucci, P.; Bove, M.; Sikora, V.; Dimonte, S.; Morgese, M.G.; Schiavone, S.; Di Cesare Mannelli, L.; Ghelardini, C.; Trabace, L. Glucoraphanin Triggers Rapid Antidepressant Responses in a Rat Model of Beta Amyloid-Induced Depressive-like Behaviour. Pharmaceuticals 2022, 15, 1054. [Google Scholar] [CrossRef] [PubMed]
- Batalle, G.; Cabarga, L.; Pol, O. The Inhibitory Effects of Slow-Releasing Hydrogen Sulfide Donors in the Mechanical Allodynia, Grip Strength Deficits, and Depressive-Like Behaviors Associated with Chronic Osteoarthritis Pain. Antioxidants 2019, 9, 31. [Google Scholar] [CrossRef]
- Cabarga, L.; Batalle, G.; Pol, O. Treatment with slow-releasing hydrogen sulfide donors inhibits the nociceptive and depressive-like behaviours accompanying chronic neuropathic pain: Endogenous antioxidant system activation. J. Psychopharmacol. 2020, 34, 737–749. [Google Scholar] [CrossRef]
- Yao, W.; Lin, S.; Su, J.; Cao, Q.; Chen, Y.; Chen, J.; Zhang, Z.; Hashimoto, K.; Qi, Q.; Zhang, J.C. Activation of BDNF by transcription factor Nrf2 contributes to antidepressant-like actions in rodents. Transl. Psychiatry 2021, 11, 140. [Google Scholar] [CrossRef]
- Tang, R.; Cao, Q.Q.; Hu, S.W.; He, L.J.; Du, P.F.; Chen, G.; Fu, R.; Xiao, F.; Sun, Y.R.; Zhang, J.C.; et al. Sulforaphane activates anti-inflammatory microglia, modulating stress resilience associated with BDNF transcription. Acta Pharmacol. Sin. 2022, 43, 829–839. [Google Scholar] [CrossRef]
- He, L.; Zheng, Y.; Huang, L.; Ye, J.; Ye, Y.; Luo, H.; Chen, X.; Yao, W.; Chen, J.; Zhang, J.C. Nrf2 regulates the arginase 1(+) microglia phenotype through the initiation of TREM2 transcription, ameliorating depression-like behavior in mice. Transl. Psychiatry 2022, 12, 459. [Google Scholar] [CrossRef]
- Hernandez-Sanchez, L.Y.; Gonzalez-Trujano, M.E.; Moreno, D.A.; Vibrans, H.; Castillo-Juarez, I.; Dorazco-Gonzalez, A.; Soto-Hernandez, M. Pharmacological evaluation of the anxiolytic-like effects of an aqueous extract of the Raphanus sativus L. sprouts in mice. Biomed. Pharmacother. 2023, 162, 114579. [Google Scholar] [CrossRef]
- Panczyszyn-Trzewik, P.; Stachowicz, K.; Misztak, P.; Nowak, G.; Sowa-Kucma, M. Repeated Sulforaphane Treatment Reverses Depressive-like Behavior and Exerts Antioxidant Effects in the Olfactory Bulbectomy Model in Mice. Pharmaceuticals 2024, 17, 762. [Google Scholar] [CrossRef]
- Ghazizadeh-Hashemi, F.; Bagheri, S.; Ashraf-Ganjouei, A.; Moradi, K.; Shahmansouri, N.; Mehrpooya, M.; Noorbala, A.A.; Akhondzadeh, S. Efficacy and safety of sulforaphane for treatment of mild to moderate depression in patients with history of cardiac interventions: A randomized, double-blind, placebo-controlled clinical trial. Psychiatry Clin. Neurosci. 2021, 75, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.S.; Bonner, A.M. The Revised Definition and Classification of Epilepsy for Neurodiagnostic Technologists. Neurodiagnostic J. 2018, 58, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Dubey, V.; Dixit, A.B.; Tripathi, M.; Chandra, P.S.; Banerjee, J. Differential Levels of Tryptophan-Kynurenine Pathway Metabolites in the Hippocampus, Anterior Temporal Lobe, and Neocortex in an Animal Model of Temporal Lobe Epilepsy. Cells 2022, 11, 3560. [Google Scholar] [CrossRef]
- Janmohamed, M.; Brodie, M.J.; Kwan, P. Pharmacoresistance—Epidemiology, mechanisms, and impact on epilepsy treatment. Neuropharmacology 2020, 168, 107790. [Google Scholar] [CrossRef]
- Kubova, H.; Mares, P.; Suchomelova, L.; Brozek, G.; Druga, R.; Pitkanen, A. Status epilepticus in immature rats leads to behavioural and cognitive impairment and epileptogenesis. Eur. J. Neurosci. 2004, 19, 3255–3265. [Google Scholar] [CrossRef]
- Pearson-Smith, J.N.; Patel, M. Metabolic Dysfunction and Oxidative Stress in Epilepsy. Int. J. Mol. Sci. 2017, 18, 2365. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, X.; Jia, W.; Zhang, C.; Boczek, T.; Harding, M.; Liu, Y.; Li, M.; Zhang, S.; Lei, S.; et al. Circulating glutathione peroxidase and superoxide dismutase levels in patients with epilepsy: A meta-analysis. Seizure 2021, 91, 278–286. [Google Scholar] [CrossRef]
- El Badawy, S.A.; Ogaly, H.A.; Abd-Elsalam, R.M.; Azouz, A.A. Benzyl isothiocyanates modulate inflammation, oxidative stress, and apoptosis via Nrf2/HO-1 and NF-kappaB signaling pathways on indomethacin-induced gastric injury in rats. Food Funct. 2021, 12, 6001–6013. [Google Scholar] [CrossRef]
- Wang, W.; Wu, Y.; Zhang, G.; Fang, H.; Wang, H.; Zang, H.; Xie, T.; Wang, W. Activation of Nrf2-ARE signal pathway protects the brain from damage induced by epileptic seizure. Brain Res. 2014, 1544, 54–61. [Google Scholar] [CrossRef]
- Carrasco-Pozo, C.; Tan, K.N.; Borges, K. Sulforaphane is anticonvulsant and improves mitochondrial function. J. Neurochem. 2015, 135, 932–942. [Google Scholar] [CrossRef]
- Socala, K.; Nieoczym, D.; Kowalczuk-Vasilev, E.; Wyska, E.; Wlaz, P. Increased seizure susceptibility and other toxicity symptoms following acute sulforaphane treatment in mice. Toxicol. Appl. Pharmacol. 2017, 326, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Pauletti, A.; Terrone, G.; Shekh-Ahmad, T.; Salamone, A.; Ravizza, T.; Rizzi, M.; Pastore, A.; Pascente, R.; Liang, L.P.; Villa, B.R.; et al. Targeting oxidative stress improves disease outcomes in a rat model of acquired epilepsy. Brain A J. Neurol. 2019, 142, e39. [Google Scholar] [CrossRef] [PubMed]
- Sandouka, S.; Shekh-Ahmad, T. Induction of the Nrf2 Pathway by Sulforaphane Is Neuroprotective in a Rat Temporal Lobe Epilepsy Model. Antioxidants 2021, 10, 1702. [Google Scholar] [CrossRef] [PubMed]
- Danek, J.; Danacikova, S.; Kala, D.; Svoboda, J.; Kapoor, S.; Posusta, A.; Folbergrova, J.; Tauchmannova, K.; Mracek, T.; Otahal, J. Sulforaphane Ameliorates Metabolic Changes Associated With Status Epilepticus in Immature Rats. Front. Cell. Neurosci. 2022, 16, 855161. [Google Scholar] [CrossRef]
- Folbergrova, J.; Jesina, P.; Otahal, J. Protective Effect of Sulforaphane on Oxidative Stress and Mitochondrial Dysfunction Associated with Status Epilepticus in Immature Rats. Mol. Neurobiol. 2023, 60, 2024–2035. [Google Scholar] [CrossRef]
- Aghaie, F.; Rajabi, M.; Hosseini, A.; Moradifar, F.; Koneshlou, S.; Hosseini, A. Preventive Effects of Sinigrin Against the Memory Deterioration in the Pentylenetetrazole-Kindled Male Wistar Rats: Possible Modulation of NLRP3 Pathway. Neuromolecular Med. 2022, 24, 311–319. [Google Scholar] [CrossRef]
- Xiaoyu, C.; Hongzhen, Z.; Nan, P.; Tengwei, G.; Yanan, G.; Yan, G.; Haiyan, L.; Li, M.; Haiya, W.; Yujun, W.; et al. Benzyl isothiocyanate ameliorates cognitive function in mice of chronic temporal lobe epilepsy. Front. Neurol. 2024, 15, 1330102. [Google Scholar] [CrossRef]
- Canistro, D.; Barillari, J.; Melega, S.; Sapone, A.; Iori, R.; Speroni, E.; Paolini, M. Black cabbage seed extract affects rat Cyp-mediated biotransformation: Organ and sex related differences. Food Chem. Toxicol. 2012, 50, 2612–2621. [Google Scholar] [CrossRef]
- Paolini, M.; Perocco, P.; Canistro, D.; Valgimigli, L.; Pedulli, G.F.; Iori, R.; Croce, C.D.; Cantelli-Forti, G.; Legator, M.S.; Abdel-Rahman, S.Z. Induction of cytochrome P450, generation of oxidative stress and in vitro cell-transforming and DNA-damaging activities by glucoraphanin, the bioprecursor of the chemopreventive agent sulforaphane found in broccoli. Carcinogenesis 2004, 25, 61–67. [Google Scholar] [CrossRef]
- Nugon-Baudon, L.; Rabot, S.; Szylit, O.; Raibaud, P. Glucosinolates toxicity in growing rats: Interactions with the hepatic detoxification system. Xenobiotica 1990, 20, 223–230. [Google Scholar] [CrossRef]
- Galanty, A.; Grudzinska, M.; Pazdziora, W.; Sluzaly, P.; Pasko, P. Do Brassica Vegetables Affect Thyroid Function?—A Comprehensive Systematic Review. Int. J. Mol. Sci. 2024, 25, 3988. [Google Scholar] [CrossRef]
- Yagishita, Y.; Fahey, J.W.; Dinkova-Kostova, A.T.; Kensler, T.W. Broccoli or Sulforaphane: Is It the Source or Dose That Matters? Molecules 2019, 24, 3593. [Google Scholar] [CrossRef] [PubMed]
- Zanichelli, F.; Capasso, S.; Cipollaro, M.; Pagnotta, E.; Carteni, M.; Casale, F.; Iori, R.; Galderisi, U. Dose-dependent effects of R-sulforaphane isothiocyanate on the biology of human mesenchymal stem cells, at dietary amounts, it promotes cell proliferation and reduces senescence and apoptosis, while at anti-cancer drug doses, it has a cytotoxic effect. Age 2012, 34, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Zanichelli, F.; Capasso, S.; Di Bernardo, G.; Cipollaro, M.; Pagnotta, E.; Carteni, M.; Casale, F.; Iori, R.; Giordano, A.; Galderisi, U. Low concentrations of isothiocyanates protect mesenchymal stem cells from oxidative injuries, while high concentrations exacerbate DNA damage. Apoptosis 2012, 17, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Sestili, P.; Paolillo, M.; Lenzi, M.; Colombo, E.; Vallorani, L.; Casadei, L.; Martinelli, C.; Fimognari, C. Sulforaphane induces DNA single strand breaks in cultured human cells. Mutat. Res. 2010, 689, 65–73. [Google Scholar] [CrossRef]
- Xie, J.; Yang, M.R.; Hu, X.; Hong, Z.S.; Bai, Y.Y.; Sheng, J.; Tian, Y.; Shi, C.Y. Moringa oleifera Lam. Isothiocyanate Quinazolinone Derivatives Inhibit U251 Glioma Cell Proliferation through Cell Cycle Regulation and Apoptosis Induction. Int. J. Mol. Sci. 2023, 24, 11376. [Google Scholar] [CrossRef]
- Hac, A.; Brokowska, J.; Rintz, E.; Bartkowski, M.; Wegrzyn, G.; Herman-Antosiewicz, A. Mechanism of selective anticancer activity of isothiocyanates relies on differences in DNA damage repair between cancer and healthy cells. Eur. J. Nutr. 2020, 59, 1421–1432. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Compounds | In Vitro Model | Dose | Treatment Duration | Main Findings | Ref. |
|---|---|---|---|---|---|
| SFN | BV2 cells treated with LPS (100 ng/mL) for 6 h and 24 h | 5–10 μM | 1, 3, 6, 12, 24 h | Inhibition of nitrite production, iNOS, and COX-2 expression; modulation of the MAPK pathway. | [39] |
| SFN | BV-2 cells treated with LPS (0.5 mg/L) for 12 h | 5, 10, and 15 μM | 12 h | Inhibition of NF-kB and the release of inflammatory mediators. Inhibition of necroptosis mediated by JNK E p65. | [40] |
| SFN | N9 microglia cells treated with LPS (1 μg/mL) for 4 h | 5 μM | 1 h before LPS | Inhibition of NLRP3 inflammasome. | [41] |
| SFN | BV-2 cells treated with MGO-AGEs (1 mg/mL) | 1–20 μM | 24 h | Reduction in ROS. Inhibition of NF-κB activation and the release of pro-inflammatory cytokines. | [42] |
| SFN | Primary microglia isolated from young and adult Balb/c mouse and BV-2 microglia cell line treated with LPS (100 ng/mL) for 8 h | 2.5 μM | Primary microglia: pre-treatment for 1 h before LPS BV-2 cell line: 1, 6, 9, 24 h | Upregulation of ARE genes including NQO1, HMOX1, and GCLM in both cell lines. Reduction in pro-inflammatory markers in primary microglia from adult and aged mice. | [43] |
| SFN | SH-SY5Y cells treated with CPF (100 μM) for 24 h BV2 cells treated with LPS (500 ng/mL) for 24 h | 5 μM | 6–24 h | Downregulation of IL-1β, TNF-α, and PGE2. Increased HO-1 expression. | [44] |
| SFN | Primary microglia treated with LPS (1 μg/mL) for 24 h | 5, 10, and 30 μM | 1, 3, and 5 h | Induction of reversible elongations of microglia; increased expres-sion of Akt. | [45] |
| SFN | BV2 cells treated with LPS (1 μg/mL) for 24 h | 20 μM | 24 h | Inhibition of NF-κB pathway. | [46] |
| SFN | Astrocytoma cells transfected with Nrf2 short interference RNA (siRNA) and treated with OKA (20 nM) for 24 h | 10 μM | 24 h | Increased cell viability; restored Nrf2, HO-1, and GCLC expression. | [48] |
| SFN NAC | BV2 cells and primary cortical neuronal culture treated with LPS (200 ng/mL) + IFN-γ (20 ng/mL) for 48 h | SFN: NAC concentrations: 1 μM:10 μM or 5 μM:50 μM | 48 h | Increased viability; reduction in nitrite level and of TNF-α. | [50] |
| SFN | BV2 cells and primary microglia of aged mouse treated with LPS (100 ng/mL) | 2.5 μM | 3, 6, 9, and 24 h | Nrf2 activation and reduction in IL-6 and IL-1β. | [51] |
| SFN-Enriched Broccoli Sprouts | BV-2 cells treated with LPS (100 ng/mL) for 24 h | 100 μg/mL | 24 h | Inhibition of NF-κB and of the secretion of pro-inflammatory pro-teins (iNOS, COX-2, TNF- α, IL-6, IL-1β, PGE2). Increased expression of Nrf2 and HO-1 and reduction in apoptosis. | [55] |
| BITC | BV2 cell treated with LPS (1 μg/mL) for 3 h and 48 h | 1, 5 and 10 μM | 1 h before LPS | Reduced IL-1β protein levels. Inhibition of NLRP3 and IL-1β. | [57] |
| ESE | NSC-34 motor neurons treated with the medium of LPS-stimulated RAW 264.7 (1 μg/mL) for 24 h | 0.1, 0.2, 0.3 e 0.4 μg/mL | 24 h pre-treatment | Increased expression of IL-10. Reduction in apoptosis and TNF-α expression. | [58] |
| GMG-ITC | SH-SY5Y treated with H2O2 (300 μM) for 4 h | 1.25 μg/mL | 72 h | Reduction in ROS through Nrf2 activation; modulation of the MAPK pathway and reduction in NF-κB levels. | [59] |
| AITC | BV2 cells treated with LPS (100 ng/mL) for 24 h | 1, 5, 10, 20 μM | 30 min before LPS | Inhibition of COX-2, iNOS, TNF-α, IL-6, PGE2, and NO. | [60] |
| AITC, PEITC, SFN | Astrocytes from the neocortical tissues of 1-day-old Wistar rats treated with LPS (10 μg/mL) for 20 h | AITC: 5, 25, 50, 400 μM PEITC and SFN: 5, 25, 50 μM | 20 h | Inhibition of MMP1 and MMP3 release through modulation of the ERK pathway. | [61] |
| ITH12674 | Cortical neuron treated with TBH (30 μM) for 24 h | 0.1, 0.3, and 1 μM | 24 h | Reduction in ROS. Increased GSH levels. | [62] |
| ITH12674 | Primary glial cell treated with LPS (1 μg/mL) for 18 h | 10 μM | 18 h | Reduction in IL-1β, TNFα, and NF-κB. Increased expression of Nrf2 and HO-1. | [63] |
| Compounds | In Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN | C57BL6/J mice treated with LPS (100 μg/kg) for 5 days | 5 mg/kg | 5 days | Increased expression of Akt and elongation in microglia cells. | [45] |
| SFN | C57BL/6 mice treated with LPS (0.25 mg/kg) for 7 days | 20 mg/kg | 7 days | Improvement in learning and memory deficits. Modulation of the mTOR pathway, increasing BDNF expression. | [47] |
| SFN | Male Sprague–Dawley rats OKA-injected (200 ng) for 2 days | 5 mg/kg | 1 h prior to and 24 h after OKA administration | Improvement in memory function. | [48] |
| SFN | B6.129P-Cx3cr1tm1Litt/J) mice; Cx3cr1+/+ mutant for hTAUP301L | 50 mg/kg | 3 weeks | Inverted astrogliosis. | [49] |
| SFN NAC | Male Sprague–Dawley rats with induction of lateral FPI | NAC: 500 mg/kg; SFN: 5 mg/kg | 13 days | Reduction in pro-inflammatory biomarkers. | [50] |
| SFN | Balb/c mice treated with LPS (1 μg) | 50 mg/kg | 3 days | Reduction in pro-inflammatory mediators in hippocampus and liver. | [51] |
| SFN | C57BL/6J wild type; Pde6brd10 mice | 20 mg/kg | 7 days | Reduction in glial cell activation. Reduction in IL-1β and retinal degeneration. | [52] |
| SFN | Male Wistar rats treated with ammonium diet for 6 weeks | 0.5 mg/kg | 6 weeks | Reduction in astrocyte and microglia activation. Normalized extracellular GABA. Reduction in pro-inflammatory cytokines. | [53] |
| SFN | Male C57Bl/6N mice tMCAO model | 25 mg/kg body weight | 3, 7 and 23 h after tMCAO | Inhibition of inflammasomes causes anti-inflammatory effects in ischemic stroke. | [54] |
| Broccoli sprouts enriched with SFN | Male ICR mice treated with SCOP (1.2 mg/kg) | 200 mg/kg | 2 weeks | Nrf2 activation. Inhibition of caspase-3 and reduction in neuronal apoptotic process. | [55] |
| 10% Broccoli diet | Adult (4-month-old) and aged (18-month-old) BALB/c mice treated with LPS (0.33 mg/kg) for 24 h | AIN-93M; AIN-93M + 10% freeze-dried broccoli | 24 days | Reduction in astrocyte markers and oxidative stress. | [56] |
| ITH12674 | Sprague–Dawley rats and C57/BL6 j mice treated with LPS (0.5 mg/kg) | 1 mg/kg | 2 h before LPS | Improvement in locomotion and social interaction. Reduced expression of IL-1β, TNF-α, GFAP, IBA1, and CD68. | [63] |
| Compounds | Computational and Investigational Studies | Main Findings | Ref. | ||
|---|---|---|---|---|---|
| RB and PB | Docking study | Anti-cholinesterase activities (AChE and BuChE inhibitory activities). | [70] | ||
| SFN | In silico study using different database | Identification of 45 targets involved in the pathogenesis of AD. | [69] | ||
| SFN | LC/ESI-MS | Less inclination of Aβ to aggregate. | [71] | ||
| SFN | In silico docking simulation | BACE1 inhibitor. | [72] | ||
| Compounds | In Vitro Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN | HEK293T transfected with NFE2L2 and HT22 cells | 15 μM | 12 h | Modulation of autophagy. | [73] |
| SFN | CN1.4 cells transfected by GFP and GFP-tau (s) form | 10 μM | 24 h | Reduction in ROS. Increased antioxidant gene expression. | [74] |
| SFN | N2a/APPswe cells | 1.25 and 2.5 mΜ | 48 h | Epigenetic modification. | [75] |
| SFN | PC12 cells treated with SNP (350–400 μM) | 1 μM | 24 h | Increased cell viability in the model of nitrosative stress. | [76] |
| SFN | EOC-20 microglial cell line treated with Aβ (100 ng/mL, 500 ng/mL, and 1000 ng/mL) for 24 h | 5 µM | 24 h | Increased phagocytic activity. | [77] |
| SFN | Primary cortical neuronal cells derived from 3 × Tg-AD mouse | 10 µM | 6 h | Increased CHIP levels and decreased Aβ accumulation levels. | [78] |
| Broccoli Sprouts Juices | SH-SY5Y treated with Aβ25–35 (25 μM) for 24, 48, 72 h | 10 μL/mL | 24, 48, 72 h | Inhibition of apoptosis. | [79] |
| SFN | Astrocytes from neonatal C57BL/6J mouse | 0, 0.1, 0.25, 0.5, 2.5, 5, and 10 μM | 6, 12, and 24 h | Upregulation of AQP4 expression via p38 MAPK pathway. | [80] |
| SFN | THP-1 cells treated with Aβ1–42 (10 μM) for 8 h | 5 μM | pretreated for 30 min | Decreased MerTK expression through inhibiting NF-κB nuclear translocation. | [81] |
| SFN | Primary microglia were prepared from 1-day-old C57 mice treated with Aβ (50 μM) for 24 h; primary cortical neurons from 1-day-old C57 mice treated with Aβ (50 μM) for 24 h; microglial cell line (BV-2) treated with Aβ (50 μM) for 24 h | 1 μM | For 24 h | SFN inhibits NLRP3 inflammasome activation in Aβ-activated microglia and suppresses ROS accumulation. | [82] |
| SFN | BV-2 cells treated with fAβ25–35 (50 μmol/L) for 24 h | 10 μmol/L | 24 h | SFN reverses M1-type microglia polarization by downregulating the MAPK/NF-κB signaling pathway in Aβ25–35-activated BV-2 cells. | [83] |
| SFN | Primary cortical neuronal derived by ICT mouse | 10 or 20 μM | 3–6 h | Epigenetic modification of BDNF. | [84] |
| SFN | SH-SY5Y treated with MGO (0.5 mM) | 2.5 μM | 24 h | Decreased MAPK activation. Reduced oxidative stress and increased intracellular GSH levels. | [85] |
| SFN | Primary cortical neuron cultures treated with Aβ (10 μM) for 48 h | 0.1 µM | 48 h | Maintenance of neuronal dendritic integrity. | [86] |
| SFN | SH-SY5Y treated with Aβ25−35 (20 μM) for 24 h | 2 mΜ | 3 h | Increased expression of p75NTR and decreased expression of HDAC1 and HDAC3. | [87] |
| SFN | BV-2 cells treated with LPS (1 μg/mL) for 23 h | 0.5–32 μM | 24 h | Decreased levels of NO, TNF-α, and IL-6, as well as NF-κB activation. | [89] |
| SFN | THP-1 cells treated with Aβ (5, 10, 20 μM) for 24 h | 1, 2, 5 µM | 24 h | Inhibition of inflammasome. | [90] |
| SFN | SH-SY5Y treated with Aβ (10 µM) for 24 h | 1, 2.5, 5 μM | 24 h | Inhibition of apoptosis and Aβ-induced DNA damage. | [94] |
| 6-MSITC | SH-SY5Y transfected with pCDNA3.1-NRF2ΔETGE-V5 | 9 μM | 6–16 h | Increased protein and mRNA levels of ADAM17. | [104] |
| MOR | SH-SY5Y treated with Aβ1–42 (10 µM) for 24 h | 0.5 µM | 24 h | Modulated the expression of genes involved in autophagy and senescence. | [108] |
| MOR | hPDLSCs | 0.5 µM | 48 h | Downregulation of the genes involved in mitophagy. | [109] |
| Memit | H9-derived NSCs treated with a cocktail of LPS and TNF-α (50 ng/mL for 16 h) | 10 µM | 24 h | Reduction in ROS production. Reduction in apoptosis. | [110] |
| H2S Hybrid Compounds | BV-2 cells treated with LPS (5 μg/mL for 24 h); SH-SY5Y treated with H2O2 or t-BuOOH (50 Μm for 30 min) | 1 and 5 μM | 24 h | Anti-inflammatory action; reduction in ROS and NO levels. | [111] |
| Compounds | In Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN | 3 × Tg-AD mouse and non-transgenic mouse | 10 and 50 mg/kg | 8 weeks | Increased level of CHIP and HSP70 and reduced the accumulation of Aβ and tau. | [78] |
| SFN | 3 × Tg-AD mouse | 10 mg/kg/day and 50 mg/kg/day | 8 weeks | Epigenetic modification of BDNF. | [84] |
| SFN | PS1V97L transgenic mice | 5 mg/kg | 4 months | Improvements of cognitive deficits. | [86] |
| FN | APP/PS1 double-transgenic mice | 25 mg/kg | 5 months | Improved cognitive function. | [87] |
| SFN | C57BL/6 mice receiving aluminum-containing water (0.4 g/100 mL) and i.j. with 200 mg/kg d-galactose for 80 days | 25 mg/kg | 80 days | Ameliorated neurobehavioral deficits. Reduced Aβ deposits and peroxidation. | [88] |
| SFN | Adult male SD rats treated with STZ 5 μL injection (3 mg/kg) at day 1 and day 3 | 25 and 50 mg/kg | 6 weeks | Inhibition of tau protein phosphorylation by modulating the PI3K/Akt/GSK-3β pathway. | [89] |
| SFN | Wistar rats injected with Aβ (5 μg/μL) | 10–20 mg/kg | 28 days | Reduction in oxidative stress and neuroinflammation. | [91] |
| SFN | Diabetic mice (db/db) and C57BLKS/J | 1 mg/kg | 28 days | Mitigated cognitive decline and hippocampal AD-like lesions. Increased Nrf2 level. | [92] |
| SFN | Sprague–Dawley rats with vascular cognitive impairment induced through permanent occlusion of bilateral common carotid arteries | 10 mg/kg | After surgery twice a week for 42 days | Improved learning and memory function. | [93] |
| SFN | Kunming mice receiving aluminum-containing water (0.4 g/100 mL) and s.i. of 200 mg/kg d-galactose for 90 days | 25 mg/kg | 90 days | Reduced cholinergic neuron loss. | [95] |
| SFN | Sprague–Dawley rats with i.c.v injection of 10 μL AβOs (500 pmol for 3 days) | 5 mg/kg | 7 days | Reduction in neuroinflammation and oxidative stress. Effect on the seratoninergic system. Reduced memory impairment and depressive behavior. | [96] |
| SFN | 6 adult male Swiss mice (SWR/J) treated with LPS (0.75 mg/kg) for 3 weeks | 25 mg/kg | 3 weeks | Reduction oin oxidative stress and accumulation of Aβ. Activation of the AMPK signaling pathway and reduction in caspase-3. | [99] |
| SFN | Sprague–Dawley (SD) rats treated with SCOP (1.5 mg/kg, i.p.) from 11th to 19th days | 15 mg/kg | 14 days | Improvements in behavioral and memory tests. Increased levels of BDNF and CREB, resulting in improvements of the hippocampal synaptic activity. | [100] |
| SFN | Zebra fish (Danio rerio) treated with SCOP (400 μM/L) | 25 μM/L | 1 h | Improvement in memory function. | [101] |
| SFN | Male wild-type (WT) C57BL/6J mice and Nrf2-KO mice receiving an LPS (0.5 mg/kg) i.p. injection 4 h prior to imaging of the cerebral microcirculation | 5 mg/kg and 50 mg/kg | 24 h prior to LPS/vehicle injection | Reduction in recruitment of leukocytes at the brain level by downregulating E-cadherin and VCAM. | [102] |
| BRO | Caenorhabditis elegans | 0, 100, 500, 1000, 5000, 7500, and 10,000 μg/mL | 24 h | Reduction in oxidative stress. | [103] |
| 6-MSITC | AppNLGF mice; C57BL/6J strain WT | 0.4 mg/mL | 10 months | Improved cognitive function. | [105] |
| 6-MSITC | C57Bl/6 mice receiving stereotaxic i.c.v. injection of Aβ1–42 | 5 mg/kg | 10 days | Decreased apoptosis. Reduction in ROS. Restoration of GSH levels. Increased spatial learning memory. | [106] |
| Compounds | Computational and Investigational Studies | Main Findings | Ref. | ||
|---|---|---|---|---|---|
| SFN and ER | LC-MS-MS | The two compounds have similar biotransformation processes. | [125] | ||
| Compounds | In vitro model | Dose | Treatment Duration | Main findings | Ref. |
| Red cabbage extracts | SH-SY5Y treated with α-Syn for 24 h | 100 µg/mL | 24 h | Reduction in neurotoxicity. | [114] |
| SFN | PC-12 cells treated with MPP (500 µmoL/L) for 24 h | 2.5 µmol/L | 24 h | Antioxidant activity by Nrf2 pathway. | [116] |
| SFN | PC-12 cells knockdown for Nrf2 transfected with siRNA (200 pmoL) | 1–40 μM | 9; 24 h | Activation of the Nrf2-ARE pathway. | [117] |
| SFN | SH-SY5Y treated with Rotenone (0.5 and 1 μM) for 24 h | 10 μM | Pre-treatment for 2 h | Decreased oxidative stress, mTOR-dependent inhibition of neuronal apoptosis, and the restoration of normal autophagy. | [119] |
| SFN | SH-SY5Y, HEK293T, and primary neurons treated with MPP+ (1 mM) | 1 μM | 24 h | Reduction in C/EBPβ transcription; reduction of α-Syn expression and inflammation markers. | [120] |
| SFN | SH-SY5Y transfected with DJ1/LRRK2/scramble siRNA and treated with cypermethrin (15 μM) for 24 h | 5 µM | 30 min before the cypermethrinexposure | Increased levels of Nrf2, DJ-1, Prx3, and Trx2. Decreased apoptosis. | [121] |
| SFN | SH-SY5Y treated with MPP+ (1 mM) treated for 24 h | 1 μM | 24 h | Increased levels of Nrf2 and BDNF proteins. Decreased MeCP2 protein expression. | [124] |
| SFN or ER | SH-SY5Y treated with 6-OHDA (100 μM) for 24 h | 5 µM | 24 h | Modulated mTOR expression. Increased Nrf2 and GSH levels. | [126] |
| GMG or MOR | RAW 264.7 treated with LPS (10 μg/mL) for 24 h | GMG: 1 mg/mL; MOR obtained by GMG+MYR: 20 μL/mL | Pre-treatment for 2 h. After LPS for 15 min. | Reduction in pro-inflammatory cytokines including TNF-α and IL-1β. Reduction in TLR4 receptor activation. | [128] |
| ITC-3 | BV-2 cells and DAergic CATH.a neuronal cells treated with LPS (0.2 μg/mL) for 24 h | 1–20 µM | 24 h | Reduction in pro-inflammatory cytokines. Increased Nrf2 levels and the antioxidant enzymes NQO1, HO-1, and GCL. | [129] |
| ITC-57 | BV-2 cells treated with LPS (0.2 μg/mL) for 24 h andDAergic CATH.a neuronal cells exposed to BH4 or H2O2 (0.05–0.5 μM) for 24 h | 0.5–3 μM | 24 h | Reduction in inflammation. | [130] |
| Compounds | In Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN | C57BL/6N treated with MPTP (30 mg/kg) for 22 days | 100 μM/kg | 22 days | Reduction in dopaminergic neuron apoptosis and improvement of motor deficit. | [115] |
| SFN | Wistar rats treated with cypermethrin (1.5 mg/kg twice weekly) between days 5 and 19 | 10 mg/kg | twice weekly for 12 weeks, 1 h prior to cypermethrin treatment | Reduced oxidative stress and apoptotic neuronal cell death in the rat nigrostriatal tissue. | [121] |
| GRA | C57BL/6 mice treated with MPTP food pellet (10 mg/kg for 28 days) | 0.1% GRA food pellet | 28 days | Increased levels of Nrf2 and antioxidant enzymes. | [118] |
| SFN | C57BL/6 mice treated with rotenone (30 mg/kg) for 60 days | 50 mg/kg | 60 days | Reduction in oxidative stress and apoptosis. | [119] |
| GRA | Transgenic mice expressing A53T human α-Syn treated with intrathecal administrations of C/EBPβ-HDO (100 nM per 2 μL) on Day 1 and Day 5 and contemporary MPTP (30 mg/kg for 5 days) | 0.1% | 30 days | Reduced degeneration of dopaminergic neurons. | [120] |
| SFN | Male adult C57BL/6 mice treated with MPTP (30 mg/kg) for 5 days | 10 mg/kg | 10 days | Reduction in dopaminergic neurotoxicity and neuroinflammation. Increased BDNF levels | [124] |
| SFN or ER | C57BL/6 mice treated with 6-OHDA (4 μg/μL) for 4 weeks | 30 μmol/kg | 4 weeks. | Improvement of motor deficits. Decreased death of dopaminergic neurons. Reduction in DNA fragmentation. | [126] |
| ER | C. elegans | 100 µM and 200 µM | 4 h | Reduction in α-Syn aggregation and improvement in motor capacity. | [127] |
| GMG or MOR | C57Bl/6 mice treated with MPTP (20 mg/kg) for 14 days | MOR obtained by activation of GMG: 10 mg/kg of GMG +5 μL MYR/mouse (20 μL/mL injection.); GMG: 10 mg/kg | Pre-treatment daily for 1 week | Compared with GRA, MOR had greater effects by reducing dopaminergic neuroxicity; it also reduced the apoptotic process and improved motor coordination. | [128] |
| ITC-3 | C57Bl/6 mice treated with MPTP (4 injections with 2 h intervals, 20 mg/kg for 7 days) | 30 mg/kg | 7 days | Reduction in motor deficit; activates the pathway of Nrf2, which determines both anti-inflammatory and antioxidant effects. | [129] |
| ITC-57 | C57BL/6J mice treated with MPTP (4 injections with 2 h intervals, 20 mg/kg for 7 days) | 30 mg/kg | 7 days | Improvement in motor deficits. Inhibition of inflammatory processes and oxidative stress. | [130] |
| BITC | Male Wistar rats treated with ZnSO4 (20 mg/kg twice a week for 2–12 weeks) | 10 mg/kg/day 1 h before to ZnSO4 | 12 weeks | Reduced Zn-induced neurobehavioral anomalies. Restored the level of dopamine and its metabolites. | [131] |
| 6-MSITC | C57Bl/6 mice treated with 6-OHDA (4 μg/mL for 4 weeks) | 5 mg/kg | 4 weeks | Reduction in neuroinflammation and stress. Improvement of motor deficits. | [132] |
| Compounds | In Vitro Model | Dose | Treatment Duration | Main Findings | Ref. |
|---|---|---|---|---|---|
| SFN | OLN-93 cells treated with 200 µM tert-butyl hydrogen peroxide for 4 h | 5–10 μM | 24 h | Increased the expression of HO-1, NQO-1, and p62. Increased cell viability and oligodendrocyte progenitor cell differentiation. | [137] |
| Synthetic ITCs compounds | The Jurkat T-lymphocyte cell line transfected with Human MIF | Stock solutions (1 M) of ITCs the work concentration was not specified | 30 min | Inhibition activity of MIF. | [138] |
| Compounds | In Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN | C57BL/6 mice treated with MOG (200 ng) for 14 days | 50 mg/kg | 28 days | Inhibition of inflammatory and demyelination processes. | [140] |
| Bioactive GRA | Male adult C57BL/6 mice in EAE model | 10 mg/kg GRA + 5 µL/mouse of MYR enzyme | Pre-treatment for 7 days Post drug-treatment for further 15 days | Reduction in caspase 3 levels. Increased claudin-1, -3, and -5 as well as ZO-1, and reduction in the inflammatory process. | [141] |
| SFX-01 | Female SJL mice in EAE model | 10, 50 or 300 mg/kg | Prophylactic experiment for 19 days Therapeutic experiment for 40 days | Reduced residual disability after EAE onset;reduced apoptosis. | [142] |
| GMG-ITC | C57BL/6 mice in EAE model | 10 mg/kg GMG + 5 μL/mouse MYR | Pretreatment once a day for 7 days Therapeutic experiment for 21 days | Modulation of MAP kinase pathway; reduction in the inflammatory process; reduction of apoptotic process. | [143] |
| GMG-ITC | C57Bl/6 mice in EAE model | 10 mg/kg GMG + 5 μL/mouse MYR | 28 days | Modulation of Wnt-beta catenin pathway. Blocking of the release of inflammation mediators including IL-1β, IL-6, and COX-2. Increased Nrf2 levels. | [144] |
| MOR-based cream | C57Bl/6 mice in EAE model | 2% (w/w) | 21 days | Reduction in pro-inflammatory cytokines (TNF-α, IFN-γ) and increased IL-10 levels. Reduction in neuropathic pain by blocking ion channels. | [145] |
| Compounds | In Vitro Model | Dose | Treatment Duration | Main Findings | Ref. |
|---|---|---|---|---|---|
| SFN | Neutrophils from peripheral blood of patients with ASD | 5 µM | 30 min | Recovered Trx1/TrxR1 redox system; reduced ROS production induced by methylmercury. | [151] |
| SFN@EVs | hPC12 cells | 2.25 mM SFN-loaded 0.021 mM EVs | 24 h | Increased Nrf2 gene expression. Decreased IL-6 gene expression. | [152] |
| Compounds | Ex Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN | Human PBMCs LPS-treated human PBMCs Human PBMCs | 2 or 5 µM 5 µM 0.5 µM | 6 h Pre-treatment 30 min Every 24 h for 3 days | Increased expression of cytoprotective enzymes (NQO1, HO-1, AKR1C1). Decreased levels of pro-inflammatory markers (IL-6, IL-1β, COX-2 e, TNF-α) after stimulation with LPS. Increased expression of cytoprotective enzymes (NQO1 and AKR1C1). | [153] |
| SFN | LPS-treated monocytes isolated from blood of ASD subjects | 5 µM | 12–16 h | Increased Nrf2 expression. Reduced oxidative damage (iNOS, Nitrotyrosine). Upregulated antioxidant enzymes (SOD1, GPx1, GR). Reduction in NF-κB activation. Decreased pro-inflammatory cytokines levels (IL-6, IL-1β). | [154] |
| Compounds | In Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN@EVs | Brain of chick embryos exposed to sodium valproate | 2.25 mM SFN-loaded 0.021 mM EVs | 10 days | Increased survival rate compared to sodium valproate-exposed groups. Restoration of normal gray and white matter structure. Improved bipolar neuron orientation. | [152] |
| SFN | Male BTBR T + Itpr3tf/J (BTBR) C57BL/6 (C57) mice (8–10 weeks old) | 50 mg/kg/day ≈ 282 μmol/kg/day 50 mg/kg/day | 7 days 7 days | Reduction in self-grooming/marble burying behavior, as well as increased social interaction. Reduction in Th17 immune responses (STAT3, RORC, IL-17 A, and IL-23R expression in CD4+T cells). Reduction in oxidative stress parameters in neutrophils/cerebellum (NF- κ B, iNOS, and lipid peroxides). Overexpression of SOD, GPx, and GR. | [155] |
| SFN | MIA-induced ASD-like rats | 20 µg/kg | 28 days | Improving of the sniffing times. Changes in the gut microbiota composition. | [156] |
| GRA | MIA-induced pregnant and lactating ddY famale mice | 0.1% GF pellets | 16 days | Improvement in the time spent in chamber in juvenile offspring after MIA. Improvement in the retention test (NORT) in the adult offspring after MIA. Reduction in parvalbumin immunoreactivity in the mPFC in the adult offspring after MIA. | [157] |
| Study Type | Partecipants | Treatment | Daily Dose | Duration | Main Findings | Ref. |
|---|---|---|---|---|---|---|
| Single-Arm Clinical Trial | 17 Children (aged 4–7) with ASD (n = 6) or healthy controls (n = 11) | GRA group: Tablet containing broccoli seed and sprout extracts as a source of GRA and active MYR enzyme (Avmacol®) Healthy control group: Not treated | ≥30 μmoL of GRA per table | 12 weeks | Improvement in verbal or non-verbal communication (OARS-4), but no changes in social interaction and repetitive/ritualistic behaviors. No association between gut microbial diversity and treatment. Correlation of 35 gut microbiome abundance changes with ASD symptoms. | [156] |
| Phase II, double-blind, randomized, placebo-controlled trial | 44 young men (aged 13–27) with moderate to severe ASD (n = 29 SFN vs. n = 15 placebo) | SFN group: Capsules of SFN-rich broccoli sprout extracts Control group: Placebo capsules containing microcrystalline cellulose | 50–150 µmoL of SFN once daily ≈1.41 μmoL/kg/day SFN ≈0.25 mg/kg/day SFN | 18 weeks | Improvement in social interaction, abnormal behavior, and verbal communication (ABC, SRS-2, CGI-I). | [158] |
| Phase III, open-label trial | 15 young men (aged 5–22) with ASD | Tablet containing broccoli seed and sprout extracts as a source of GRA and active MYR enzyme (Avmacol®) | ≈5.5 μmoL/kg/day GRA ~ 2.2 μmoL/kg/day SFN | 12 weeks | Improvement in social responsiveness related to 77 urinary metabolites. | [160] |
| Double-blind, randomized, placebo-controlled | 60 young men (aged 4–12) with ASD (n = 30 SFN vs. n = 30 placebo) | SFN group: Risperidone + capsules containing SFN Control group: Risperidone + placebo caspules | 50–100 µmoL of SFN once daily ≈2.14 μmoL/kg/day SFN ≈0.38 mg/kg/day SFN | 10 weeks | Improvements in irritability and hyperactivity, but none in lethargy/social interaction, stereotypy, inappropriate speech (ABC), and frequent adverse events. | [161] |
| Phase II, double-blind, randomized, placebo-controlled trial | 45 children (aged 3–12) with moderate or greater severity of ASD (n = 22 GRA vs. n = 23 placebo) | GRA group: Tablet containing broccoli seed and sprout extracts as a source of GRA and active MYR enzyme (Avmacol®) Control group: Placebo tables containing microcrystalline cellulose and magnesium stearate. | ≈5.5 μmoL/kg/day GRA ~ 2.2 μmoL/kg/day SFN | 30 weeks | Improvement in aberrant behavior. GSSG increased, while free GSH/GSSG and total GSH/GSSG decreased. IL-6, Il-1β, and TNF-α decreased. | [162] |
| Double-blind, randomized, placebo-controlled trial | 28 children (aged 3–7) with ASD (n = 15 SFN vs. n = 13 placebo) | SFN group: A SFN-rich broccoli sprout/red radish sprout powder mix (BroccoPhane®) Control group: Powder containing spinach puree | 50 µmoL of SFN once daily ≈1.41 μmoL/kg/day SFN ≈0.25 mg/kg/day SFN | 36 weeks | No statistically significant differences between the SFN and placebo groups in the behavioral and cognitive symptom rating scales (ABC, SRS-2, and ADOS-2). | [163] |
| Double-blind, randomized, placebo-controlled multi-center trial | 108 Children (aged 3–15) with ASD ASD (n = 60 GRA vs. n = 48 placebo) | GRA group: Tablet containing broccoli seed and sprout extracts as a source of GRA and active MYR enzyme (Avmacol®) Control group: Placebo tablets. | ≥30 μmoL of GRA per table 2–8 tables/day dose based on body weight | 12 weeks | Significant improvements in autism features (CGI-I) and (OARS-4). No significant effects on behavior (SRS, ABC, and RBS-R). Improvements higher in participants older than 10 years of age and effects spanned range of intelligence. | [164] |
| Compounds | In Vitro Model | Dose | Treatment Duration | Main Findings | Ref. |
|---|---|---|---|---|---|
| D, L-SFN | iMGLC | 5 μM | 24 h | Inhibition of NF-κB pathway activation (downregulated FOS and IL-1β). Nrf2 pathway activation (upregulated GCLM and HMOX1). Enhanced microglia phagocytosis. | [170] |
| Compounds | In Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN GRA(Pellets containing Broccoli sprout extract powder) | For SFN: Male ICR mice (6 weeks old) treated with phencyclidine For GRA: Mice (4 weeks old) treated with phencyclidine | 30 mg/kg/day of SFN 0.1% ≈2.3 mmoL GRA per 1 kg-diet | Prophylactic treatment with SFN 10 days (once daily on days 1–5 and 8–12) Therapeutic treatment with SFN 14 days (once daily on days 15–28) Prophylactic treatment with GRA 28 days | Prophylactic treatment with SFN: Prevention of phencyclidine-induced cognitive deficits. Prevention of phencyclidine-induced reduction in the spine density in the mPFC and hippocampal CA1. Prevention of phencyclidine-induced oxidative stress (8-oxo-dG-positive cells reduction). Therapeutic treatment with SFN: Improvement in phencyclidine-induced cognitive deficits. Prophylactic treatment with GRA: Prevention of phencyclidine-induced cognitive deficits. Reduction in parvalbumina-positive cells in mPFC and CA1. Prevention of phencyclidine-induced increase in 8-oxo-dG-positive cells. | [171] |
| GRA (Pellets containing Broccoli sprout extract powder) | Juvenile offspring mice (4 weeks old) derived from poly(I:C)-treated pregnant ddY mice | 0.1% ≈2.3 mmoL GRA per 1 kg-diet | 28 days | Improvement in cognitive deficits in adult offspring after MIA. Prevention of parvalbumin immunoreactivity reduction in the prelimbic regions of mPFC in adult offspring after MIA. Normalization of centrosome-related gene expression in prefrontal cortex and hippocampus of adult offspring after MIA. Prevention of SfI1 mRNA overexpression in prefrontal cortex and hippocampus of adult offspring after MIA. | [172] |
| SFN | Female C57BL/6 J mice (8 weeks old) treated with OLZ | 90 mg/kg/day | 35 days | Decreased fat accumulation and inflammation in liver (ALT and AST). Decreased triglyceride accumulation. Restored NEFA levels. Increased hepatic glycogen storage. Decreased adducts of 4-HNE in liver. Nrf2 pathway activation (increased NQO1 mRNA gene expression). Reduced lipid peroxidation (4-HNE). | [173] |
| AITC | Female BALB/c mice (6–8 weeks old) treated with OLZ | 7.5 or 15 mg/kg/day | 42 days | Reduced body weight, as well as food and water intake. Reduced adiposity and liver weight. Improved energy expenditure. Reduced fasting blood glucose. Improved glucose tolerance. Decreased insulin resistance. Reduced total cholesterol, LDL cholesterol, and triglycerides in the serum. Increased HDL cholesterol in the serum. Reduced pro-inflammatory markers (NF-κB; IL-6, IFN-γ, TNF-α). Increased expression of orexigenic neuropeptides (AgRP and NPY). | [174] |
| SFN | Male Wistar rats treated with OLZ | 30 mg/kg/day | 21 days | Reduced body weight, BMI, and food intake. Improved blood pressure and lipidic profile (increased HDL and reduced LDL, VLDL, triglycerides, and total cholesterol). Improvement in liver function parameters (AST, ALT, TBIL). Reduced oxidative stress (increased SOD and catalises; reduced MDA and total nitrite). Reduced pro-inflammatory cytokine levels (NF-κB; IL-6, TNF-α). | [175] |
| Study Type | Participants | Treatment | Daily Dose | Duration of Intervention | Main Findings | Ref. |
|---|---|---|---|---|---|---|
| Open-label Single-arm clinical trial | 7 patients (aged 20–65) with schizophrenia | Tablets containing GRA | 30 mg/day GRA ≈68.5 μmoL/day GRA | 8 weeks | Improvement in visual recognition, attention, and short-term memory (OCLT Accuracy). No significant changes in other cognitive parameters (CogSate Battery), PANSS Total Score, and serum BDNF levels. | [176] |
| Double-blind, randomized, placebo-controlled trial | 58 patients (aged 18–65) with schizophrenia or schizoaffective disorders (n = 29 GRA vs. n = 29 placebo) | GRA group: Tablet containing broccoli seed extract as a source of GRA and active MYR enzyme from broccoli sprouts (Avmacol®) Control group: Placebo capsules containing no active ingredients. | Dose per tablet: 16 mg or 37 μmoL of GRA 6 tablets/day: 222 μmoL/day of GRA ≈ 96 mg/day of GRA 6 tablets/day: appro. 100 μmoL/day of SFN ≈ appro. 17.7 mg/day | 16 weeks | No statistically significant differences in PANSS total scores, and improvement in MCCB cognitive scores between GRA and placebo groups. | [177] |
| Double-blind, randomized, placebo-controlled trial | 172 patients (aged 18–50) with schizophrenia (n = 58 GRA High Dose, n = 56 GRA Low Dose, n = 58 placebo) | GRA High-Dose group: Tablet containing broccoli seed extract as a source of GRA and active MYR enzyme from broccoli sprouts (Avmacol®) GRA Low-Dose group: Tablet containing broccoli seed extract as a source of GRA and active MYR enzyme from broccoli sprouts (Avmacol®) Control group: 6 placebo tablets | GRA High-Dose group: 6 tablets/day ≈219 μmoL/day of GRA ≈99 μmoL/day of SFN SFN Low-Dose group: 4 tablets/day + 2 placebo tablets ≈146 μmoL/day of GRA ≈66 μmoL/day of SFN Control group: 6 placebo tablets | 22 weeks | Improvement in spatial working memory, reasoning–problem solving, and verbal learning (MCCB Domain Score). No statistically significant improvement in several cognitive domains (MCCB overall composite score). No effects on PANSS total scores. | [178] |
| Single arm clinical trial | 45 patients (aged 18–50) with schizophrenia | Tablets containing SFN (Nutramax®) | 90 mg/day of SFN ≈507.4 μmol/day of SFN | 24 weeks | Improvement in the PANSS negative subscale and PANSS total score. No change in the PANSS positive subscale. Increased HsCRP levels. Increase in SOD activity. | [179] |
| Compounds | In Vitro Model | Dose | Treatment Duration | Main Findings | Ref. |
|---|---|---|---|---|---|
| (R, S)-SFN | PC12 cells treated with NGF | 0.01, 0.1, 1 μM | 4 days | Increased number of PC12 cells with neurite growth in a dose-dependent manner. Enhanced neurite growth by Nrf2 pathway activation. | [192] |
| Compounds | In Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| DL-SFN | Male ICR mice | 1–10 mg/kg/day | 14 days(once daily) | Antidepressant-like effects: Reduced immobility time (FST, TST). Increased sucrose preference (SPT). Anxiolytic-like effects: Increased time in the central zone (OPT). Decreased latency to feeding (NSF). Reduced serum corticosterone and adrenocorticotropic hormone levels in stressed mice. Decreased serum IL-6 and TNF-α levels in stressed mice. | [146] |
| (R, S)-SFN | Male adult C57BL/6 mice (8 weeks old) CD1 mice (14 weeks old) | 10 mg/kg | 30 min | Prevented decrease in social interaction in stressed mice. Attenuated decreased sucrose preference in stressed mice. Increased BDNF levels in CA3, DG, and PFC, and decreased BDNF in NAc in stressed mice. Restored p-TrkB/TrkB ratio in CA3, DG, and PFC, and reduced p-TrkB/TrkB ratio in NAc in stressed mice. Increased levels of Nrf2 and decreased Keap1 in CA3, DG, and PFC | [192] |
| GRA (Pellet containing broccoli sprout extract powder) | Male C57BL/6 mice (5 weeks old) | 0.1% ≈ 2.2 mmol GRA per 1 kg-diet | 21 days | Prevented decreased social interaction in stressed mice.Attenuated decreased sucrose preference in stressed mice. | |
| SFN | Male adult C57BL/6 mice (8 weeks old) | 3.0, 10, 30 mg/kg | 1 day | Reduced serum TNF-α levels and microglia activation in the brain after LPS administration. Increased serum IL-10 levels. Antidepressant-like effects: Decreased TST and FST. Prevented LPS-induced reduction in BDNF, PSD-95, and GluA1 levels in the PFC, hippocampus, and NAs. Restored LPS-induced reductions in dendritic spine density in the PFC and hippocampus. Prevented excessive increases in dendritic spine density in the NAc. Antidepressant-like effects: Decreased immobility time in TST and FST after LPS administration. | [193] |
| GRA (Pellets containing broccoli sprout extract powder) | Male adult C57BL/6 mice (8 weeks old) | 0.1% ≈ 2.3 mmol GAR per 1 kg-diet | 21 days | Prevented LPS-induced reduction in BDNF, PSD-95, and GluA1 levels in the PFC and hippocampus. Restored LPS-induced reductions in dendritic spine density in the PFC and hippocampus. Prevented LPS-induced excessive increase in BDNF levels and dendritic spine density in the NAc. | |
| SFN | Male C57BL/6J mice | 10 mg/kg | 15 days (from days 14 to 28 post-surgery) | Reduced pain responses in neuropathic pain models through the Nrf2/HO-1 signaling activation and microglial activation inhibition. Alleviated mechanical allodynia and thermal hyperalgesia by reducing oxidative stress and modulating MAPK pathways. Decreased microglial activation and neuroinflammation by the downregulation of CD11b/c and suppression of JNK, ERK1/2, and p-38 phosphorylation. Restored neuronal function and neuroplasticity by the upregulation of Nrf2, HO-1, and related antioxidant enzymes. Enhanced the antiallodynic effects of morphine by preventing downregulation of µ-opioid receptors. | [194] |
| SFN | Adult male Sprague–Dawley rats (12 weeks) | 5 mg/kg | 7 days | Restored normal locomotor activity in OFT. Increased sucrose preference. Improved spatial learning and memory in the MWM. Decreased immobility time and increased jumping in the FST. Protected the serotonergic system by restoring TPH2 levels in the DRN. Normalized SERT levels in the striatum, preventing serotonin dysregulation. Reduced lipid peroxidation by decreasing MDA levels. Restored antioxidant defense by increasing GSH levels. Reduced pro-inflammatory cytokines, IL-1β and TNF-α, levels. | [96] |
| GRA | Male Wistar rats (8 weeks old) | 50 mg/kg | 30 min | Antidepressant effects: Reduced immobility and increased swimming frequency in FST. Restored 5-HT and NA levels. Decreased pro-inflammatory cytokines (NF-κB levels). Reduced ROS levels. | [195] |
| AITC PITC | Famale C57BL/6 mice with depression and anxiety associated with chronic osteoarthritis pain (6–8 weeks) | AITC: 4.4 μmoL/kg PITC: 13.3 μmoL/kg | 4 days for AITC 10 days for PITC | Antidepressant effects (significant reduction in immobility time). Increased antioxidant enzymes levels (HO-1, NQO1, GSTM1, GSTA1). | [196] |
| AITC PITC | Male C57BL/6 mice with depression associated with neuropathic pain (5–6 weeks) | AITC: 4.4 μmoL/kg PITC: 13.3 μmoL/kg | 4 days for AITC 13 days for PITC | Antidepressant-like effects: Reduced immobility time in TST in both nerve-damaged and nerve-undamaged mice. | [197] |
| SFN | Male adult exposed to LPS for 24 h: C57BL/6 mice (8 weeks old) CD1 mice (14 weeks old) | 10 mg/kg | 1 h | Antidepressant-like effects: Reduced immobility time in TST and FS by activation of Nrf2 and BDNF expression. Increased sEPSC frequency in the hippocampus by upregulation of BDNF and downregulation of its repressors (HDAC2, mSin3A, MeCP2). Increased Nrf2 and BDNF levels in mPFC and the hippocampus. | [198] |
| Nrf2 homozygous KO mice (Nrf2−/−) | 10 mg/kg | 24 h | No changes in BDNF, synaptic activity, or depression-like behaviors in Nrf2 KO mice. | ||
| SFN | Male adult exposed to social defeat stress: C57BL/6 mice (8 weeks old) CD1 mice (14 weeks old) Thy1-YFP mice | 10 mg/kg | 10 days | Increased BDNF expression in the mPFC. Promoted neural plasticity by restoring dendritic spine density. Reduced pro-inflammatory cytokines (TNF-α, IL-1β, IL-6). Increased anti-inflammatory cytokines (IL-10, IL-4). Microglial modulation: Inhibited pro-inflammatory phenotype (iNOS +).Activated anti-inflammatory phenotype (Arginase1 +). Increased social interaction (SIT). Reduced immobility time in the FST. Increased sucrose preference in the SPT. | [199] |
| SFN | Male adult exposed to social defeat stress: C57BL/6 mice (8 weeks old) CD1 mice (14 weeks old) Thy1-YFP mice | 10 mg/kg | 10 days | Activated anti-inflammatory phenotype (Arginase1 +) by increasing TREM2. Antidepressant-like effects: Increased social avoidance time. Increased SPT. Improved synaptic plasticity by BDNF-TrkB pathway modulation. Increased anti-inflammatory cytokines gene expression (IL-10, IL-4). Nrf2 pathway activation. | [200] |
| AERSS rich in sulforaphene (0.001 mg/g) SFN (0.029 mg/g), iberin (5.037 mg/g dry weight) | Swiss Webster mice | 3, 10, 30, 100 mg/kg 500 mg/kg | 30 min | Anxiolytic effect: Increased time spent in open arms (Elevated Plus Maze test). Reduction in vertical stretching behavior but no change in ambulatory activity. | [201] |
| (R, S)-SFN | Male C57BL/J6 mice subjected to OB (~8 weeks old) | 2.5, 5, 10 mg/kg | 14 days | Improvement in depressive-like behavior: Reduced hyperactivity and increased time spent in the central zone in the OFT. Reduced latency and increased time and number of grooming instances in the ST. Reduced ambulation and distance traveled in the LA. Enhanced SOD activity in the serum. Increased total antioxidant capacity (TAC) levels in the frontal cortex and serum. | [202] |
| Compounds | In Vitro Model | Dose | Treatment Duration | Main Findings | Ref. |
|---|---|---|---|---|---|
| SFN | Mixed cortical neurons and glial cells cultures from postnatal (P0–P1) Sprague–Dawley rat pups | 5 μM | 1 day | Decreased ROS during epileptiform activity. Increased GSH levels. Decreased seizure-like activity-induced neuronal death. | [215] |
| Compounds | In Vivo Model | Dose | Treatment Duration | Main Findings | Ref. |
| SFN | Adult male Wistar rats Nrf2−/− and Nrf2+/+ mice | 5 mg/kg/day in corn oil | 15 days | Suppression of amygdale kindling progression. Improvement in oxidative stress and cognitive impairment induced by epileptic seizure by Nrf2 pathway activation. | [211] |
| SFN | Adult male CD1 mice | 5 mg/kg/day in corn oil | 5 days | Increased seizure thresholds and reduced the incidence of status epilepticus. Enhanced antioxidant defenses and improved mitochondrial respiration and ATP production in the hippocampus. | [212] |
| D,L-SFN | Albino Swiss mice | 10–300 mg/kg/day in corn oil | 1 day | Decreased seizure thresholds, procolvulsant effects, and leukopenia in mice induced by high doses of SFN (≥200 mg/kg). SFN (100 mg/kg) enhanced carbamazepine efficacy. Toxic doses caused sedation (at 150–300 mg/kg), hypothermia (at 150–300 mg/kg), impairment of motor coordination (at 200–300 mg/kg), decrease in skeletal muscle strength (at 250–300 mg/kg), and death (at 200–300 mg/kg). | [213] |
| SFN or NAC+SFN | Adult male Sprague–Dawley rats | 5 mg/kg/day of SFN NAC (10 mg/ kg/day) + SFN (5 mg/kg/day) | SFN for 14 days or NAC + SFN for 7 days followed by SFN alone for 7 days or NAC + SFN for 4 days | Delayed seizure onset and frequency, as well as prevention of disease progression. Normalized hippocampal GSH/GSSG ratios. Prevented mitochondrial dysfunction, reduced oxidative damage, and attenuated HMGB1 translocation. Reduced oxidative stress during epileptogenesis. | [214] |
| SFN | Male and female Sprague–Dawley rats | 5 mg/kg/day | 5 days | Increased expression of Nrf2, HO-1, and NQO1. Restored GSH/GSSG ratio in cortex in the hippocampus. Increased total antioxidant capacity in plasma and the hippocampus. Decreased neuronal death in CA1 and CA3 hippocampal subfields. | [215] |
| SFN | 10-day-old male Wistar rats | 5 mg/kg/day | 2 day (on day 10 and day 11) | Nrf2 pathway activation. Improved brain metabolic adaptation after status epilepticus. Normalized glucose metabolism, restored cerebral blood flow, and enhanced vascular responsiveness to stimulation. Supported neuroprotection via improved metabolic resilience. | [216] |
| SFN | Immature 12-day-old male Wistar rats | 5 mg/kg/day | 2 days (48 h and 24 h before SE) | Reduced oxidative stress and mitochondrial dysfunction following status epilepticus in immature rats. Attenuated superoxide production, 3-NT and 4-HNE levels, and partially preserved mitochondrial complex I activity. Nrf2-mediated neuroprotection. | [217] |
| SIN | Adult male Wistar rats | 10 and 20 mg/kg every other day | 1 month | Delayed seizure onset and reduced kindling progression in PTZ-kindled rats. Improved memory performance and increased antioxidant enzyme levels (SOD, CAT). Reduced hippocampal expression of pro-inflammatory genes Il1b and Nlrp3. | [218] |
| BITC | C57BL/6J mice | 10 mg/kg/day in corn oil | 7 days | Improved cognitive and motor function in epileptic mice and reduced neuronal loss, particularly in the cortex. Enhanced the expression of Nrf2, HO-1, and NQO1, increased antioxidant enzyme activity (GSH-Px), and reduced MDA levels. | [219] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muscarà, C.; Gugliandolo, A.; Mazzon, E.; Calì, G. Glucosinolate Metabolites and Brain Health: An Updated Review on Their Potential Benefits in Neurodegenerative, Neurodevelopmental, and Psychiatric Disorders. Antioxidants 2025, 14, 818. https://doi.org/10.3390/antiox14070818
Muscarà C, Gugliandolo A, Mazzon E, Calì G. Glucosinolate Metabolites and Brain Health: An Updated Review on Their Potential Benefits in Neurodegenerative, Neurodevelopmental, and Psychiatric Disorders. Antioxidants. 2025; 14(7):818. https://doi.org/10.3390/antiox14070818
Chicago/Turabian StyleMuscarà, Claudia, Agnese Gugliandolo, Emanuela Mazzon, and Gabriella Calì. 2025. "Glucosinolate Metabolites and Brain Health: An Updated Review on Their Potential Benefits in Neurodegenerative, Neurodevelopmental, and Psychiatric Disorders" Antioxidants 14, no. 7: 818. https://doi.org/10.3390/antiox14070818
APA StyleMuscarà, C., Gugliandolo, A., Mazzon, E., & Calì, G. (2025). Glucosinolate Metabolites and Brain Health: An Updated Review on Their Potential Benefits in Neurodegenerative, Neurodevelopmental, and Psychiatric Disorders. Antioxidants, 14(7), 818. https://doi.org/10.3390/antiox14070818