
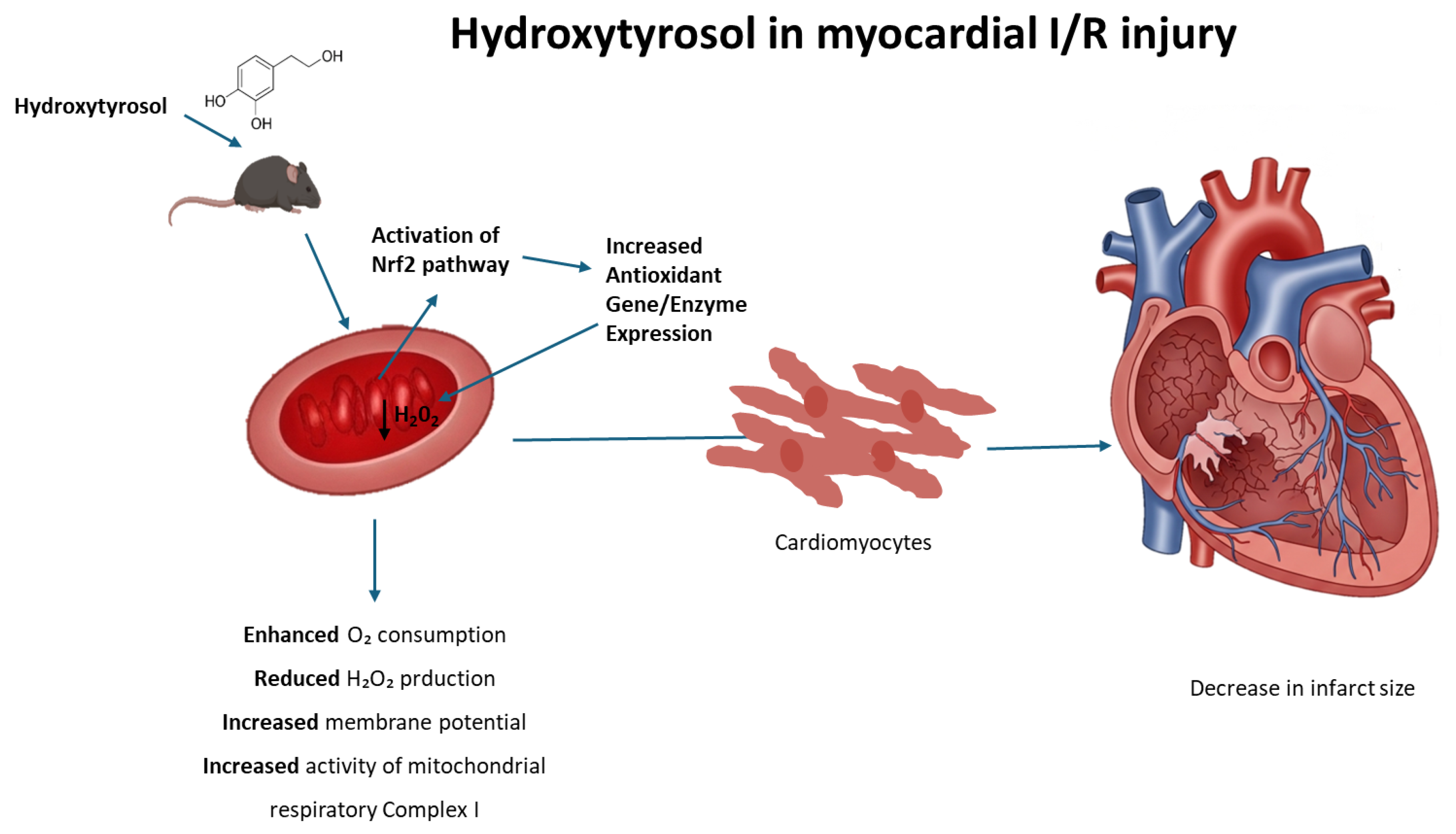
Unveiling the Cardioprotective Potential of Hydroxytyrosol: Insights from an Acute Myocardial Infarction Model

 ,
,  , , ,
, , ,  , and
, and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Care
2.3. Treatment of Mice with HT
- Control group: Untreated mice.
- I/R group: Mice subjected to I/R.
- HT group: Mice treated with HT at the specified dose.
- HT + I/R group: Mice treated with HT at the specified dose and subjected to I/R.
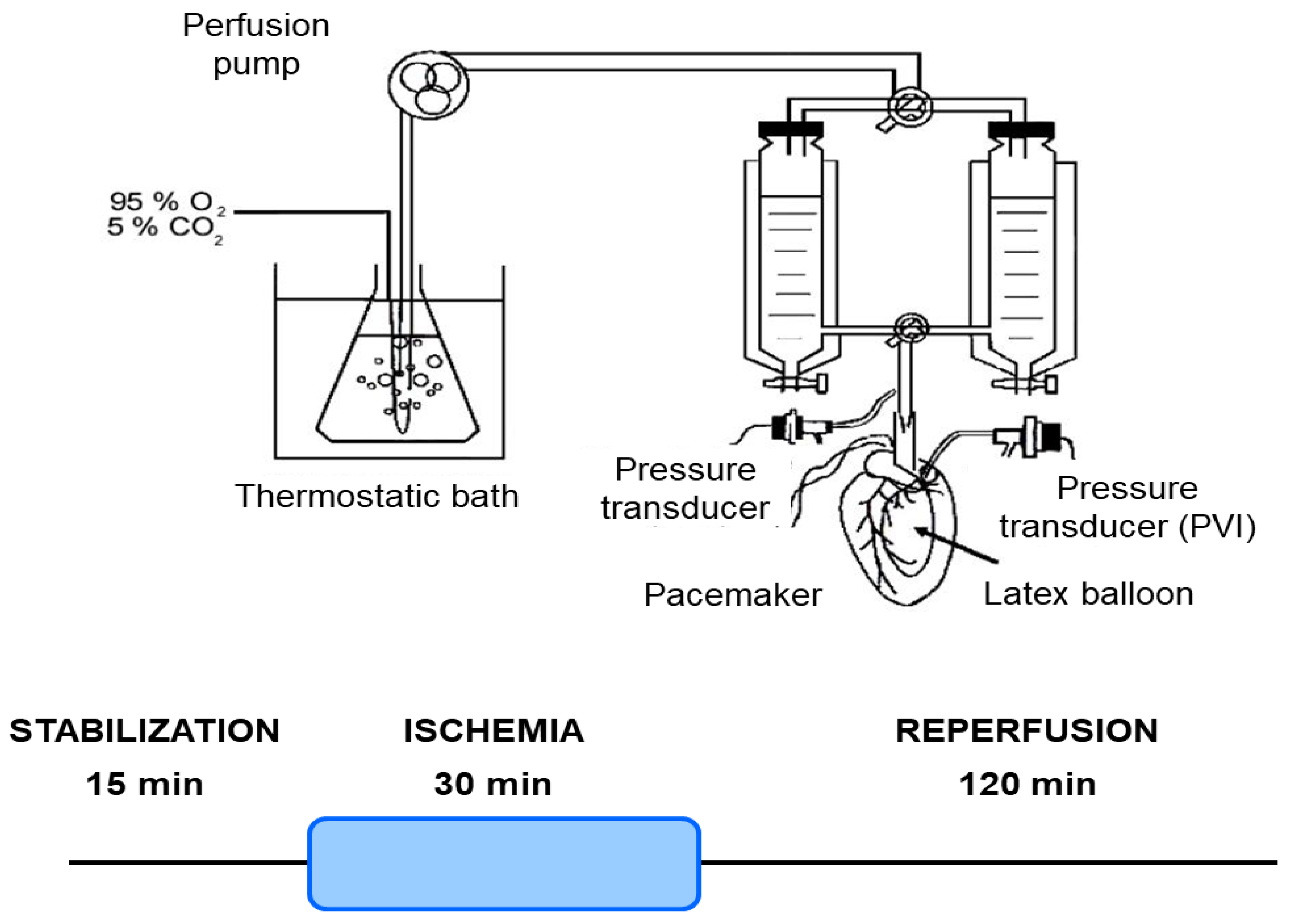
2.4. Isolated Mouse Hearts
2.5. Ischemia/Reperfusion Protocol
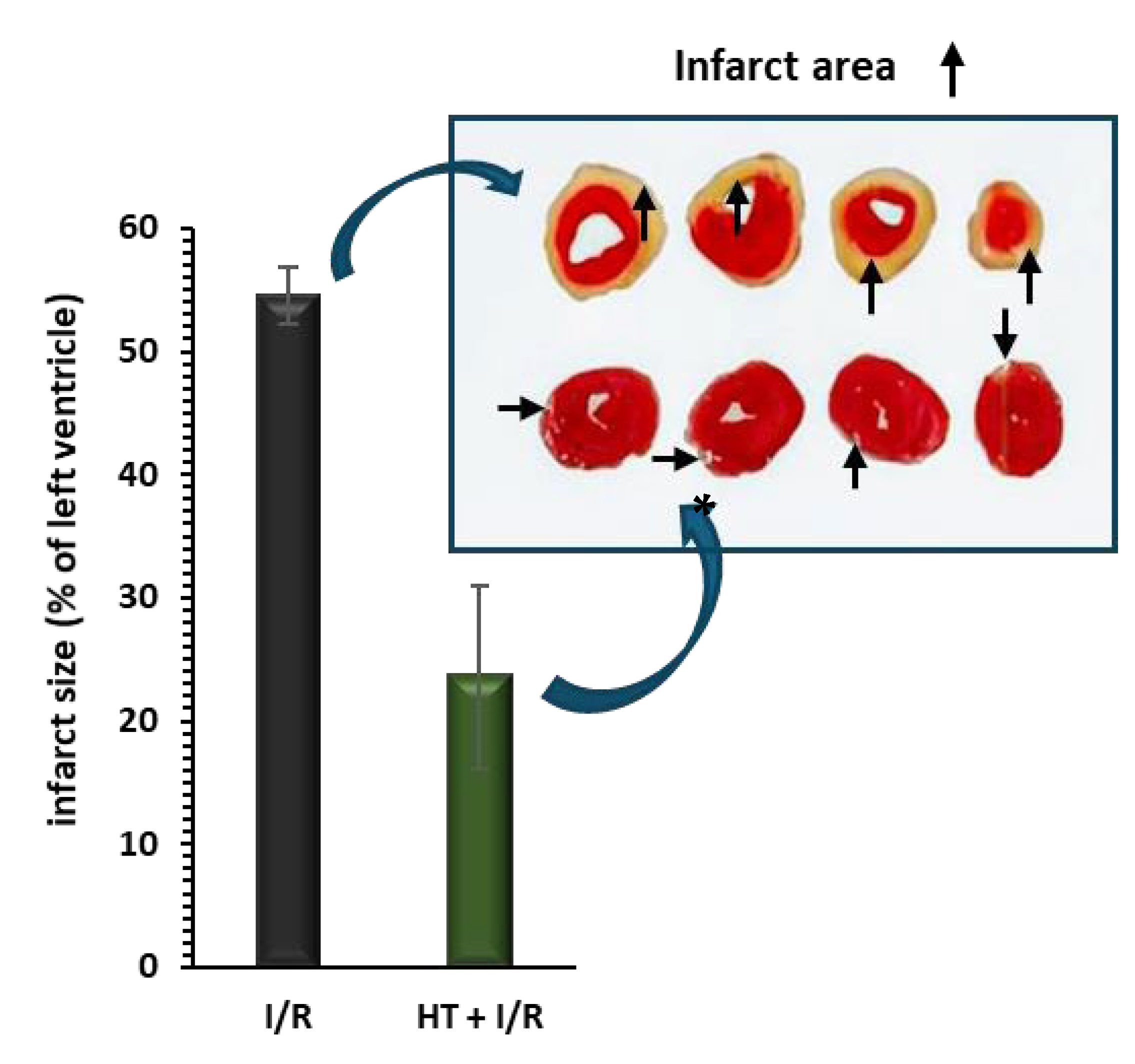
2.6. Infarct Size Measurement
2.7. Heart Mitochondrial Function
2.7.1. Heart Mitochondria Isolation and Mitochondrial Membrane Preparation
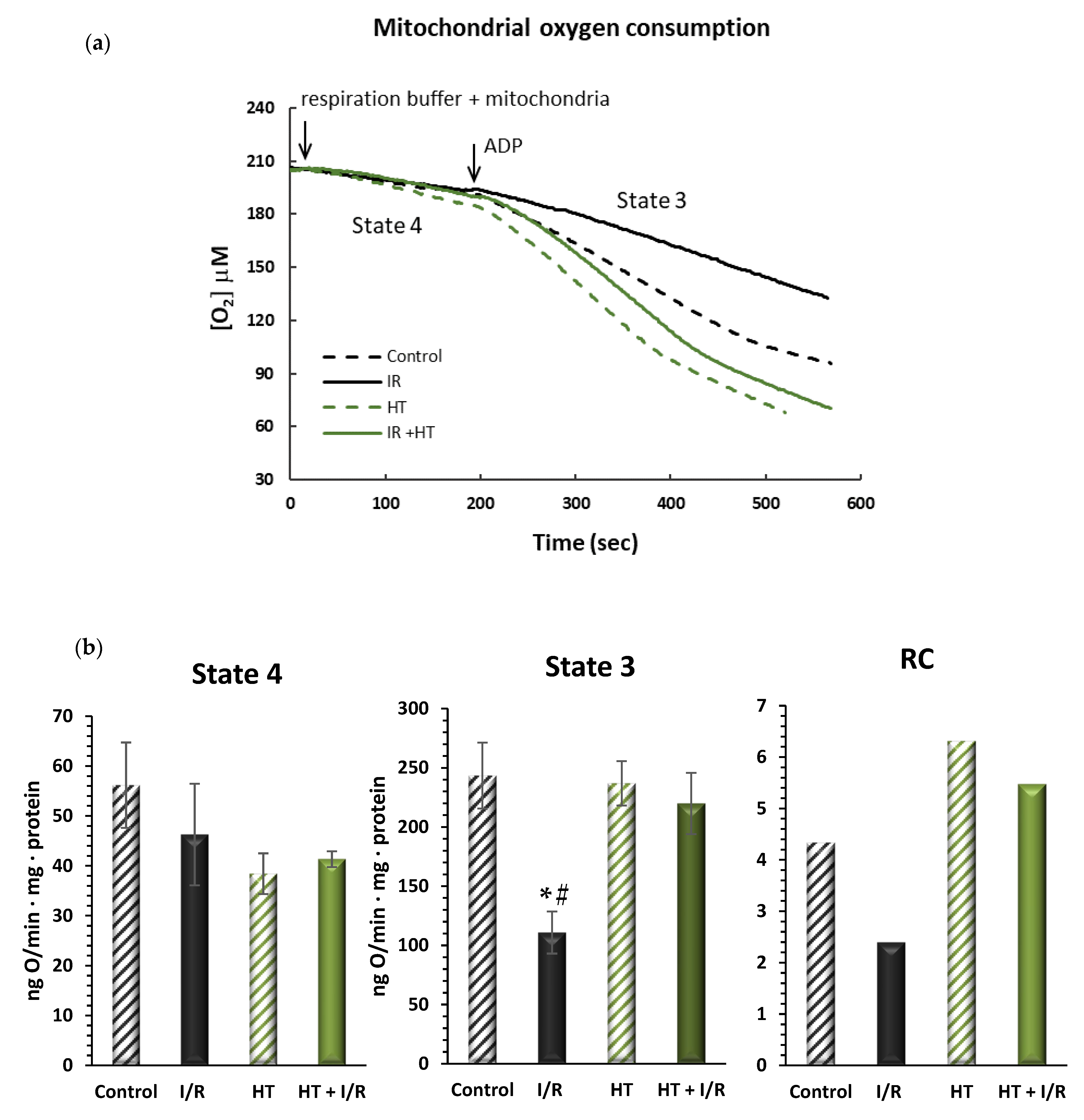
2.7.2. Mitochondrial Oxygen Consumption
2.7.3. Hydrogen Peroxide Production
2.7.4. Mitochondrial Inner Membrane Potential
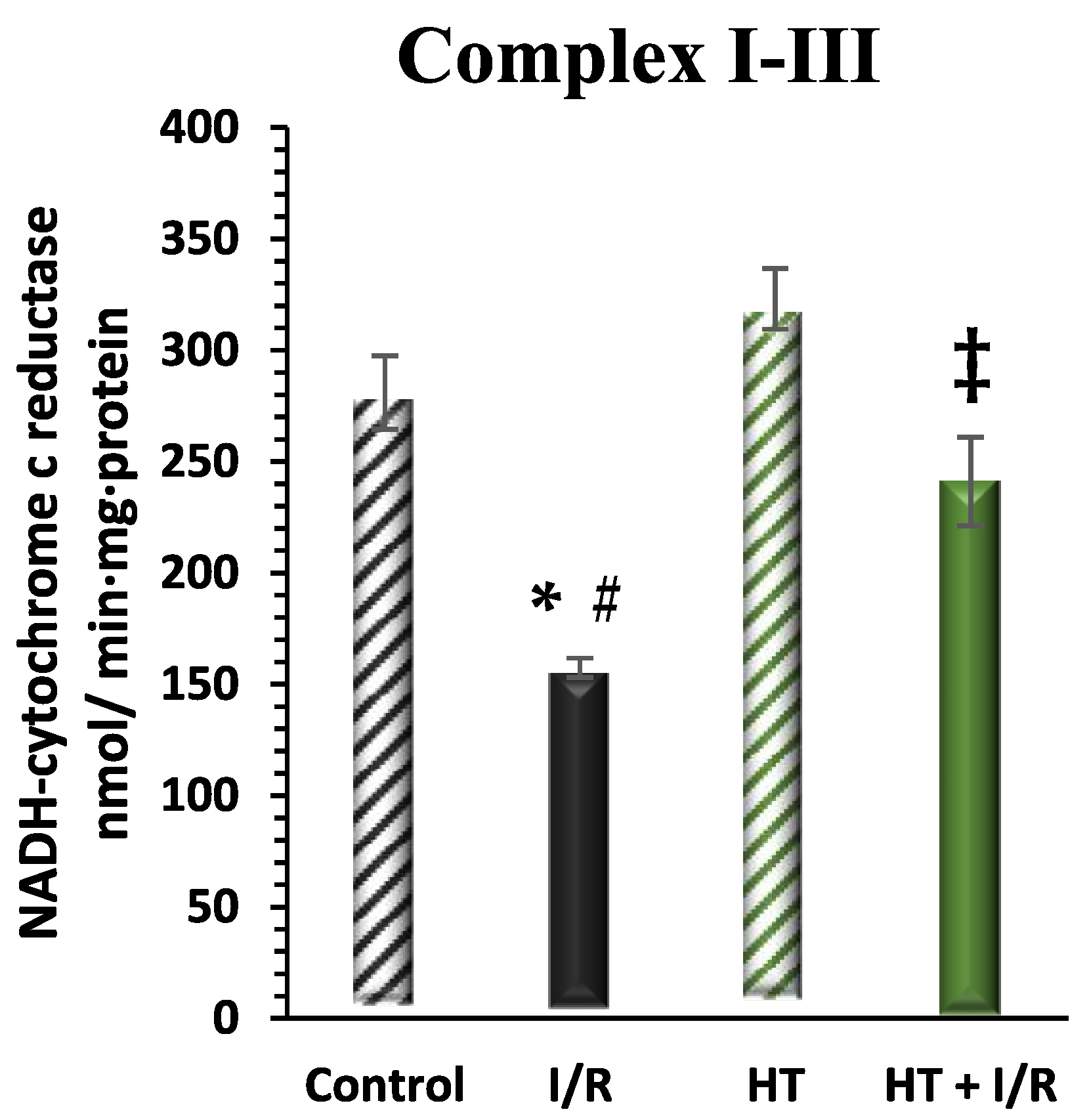
2.7.5. Mitochondrial Electron Transfer Activities
2.8. Statistical Analysis
3. Results
3.1. Infarct Size
3.2. Heart Mitochondrial Function
3.2.1. Mitochondrial Oxygen Consumption
3.2.2. Mitochondrial Hydrogen Peroxide Production
3.2.3. Mitochondrial Inner Membrane Potential
3.2.4. Mitochondrial Respiratory Complex Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- AL-Asmari1, K.M.; Al-Attar, A.M.; Abu Zeid, I.M. Potential health benefits and components of oliveoil: An overview. Biosci. Res. 2020, 17, 2673–2687. [Google Scholar]
- Guasch-Ferré, M.; Hu, F.B.; Martínez-González, M.A.; Fitó, M.; Bulló, M.; Estruch, R.; Ros, E.; Corella, D.; Recondo, J.; Gómez-Gracia, E.; et al. Olive oil intake and risk of cardiovascular disease and mortality in the PREDIMED Study. BMC Med. 2024, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, M.; Perel, P.; Taylor, S.; Kabudula, C.; Bixby, H.; Gaziano, T.A.; McGhie, D.V.; Mwangi, J.; Pervan, B.; Narula, J.; et al. The Heart of the World. Glob. Heart 2024, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Neumann, F.-J.; Sousa-Uva, M.; Ahlsson, A.; Alfonso, F.; Banning, A.P.; Benedetto, U.; Byrne, R.A.; Collet, J.-P.; Falk, V.; Head, S.J.; et al. ESC/EACTS Guidelines on Myocardial Revascularization. Eur. Heart J. 2018, 40, 87–165. [Google Scholar] [CrossRef]
- García-Niño, W.R.; Zazueta, C.; Buelna-Chontal, M.; Silva-Palacios, A. Mitochondrial Quality Control in Cardiac-Conditioning Strategies against Ischemia-Reperfusion Injury. Life 2021, 11, 1123. [Google Scholar] [CrossRef]
- Garcia-Dorado, D.; Rodríguez-Sinovas, A.; Ruiz-Meana, M.; Inserte, J. Protection Against Myocardial Ischemia-Reperfusion Injury in Clinical Practice. Rev. Española De Cardiol. (Engl. Ed. ) 2014, 67, 394–404. [Google Scholar] [CrossRef]
- Donato, M.; Evelson, P.; Gelpi, R.J. Protecting the Heart from Ischemia/Reperfusion Injury. Curr. Opin. Cardiol. 2017, 32, 784–790. [Google Scholar] [CrossRef]
- Daiber, A.; Andreadou, I.; Oelze, M.; Davidson, S.M.; Hausenloy, D.J. Discovery of New Therapeutic Redox Targets for Cardioprotection against Ischemia/Reperfusion Injury and Heart Failure. Free. Radic. Biol. Med. 2020, 163, 325–343. [Google Scholar] [CrossRef]
- Isaakidis, A.; Maghariki, J.E.; Carvalho-Barros, S.; Gomes, A.M.; Correia, M. Is There More to Olive Oil than Healthy Lipids? Nutrients 2023, 15, 3625. [Google Scholar] [CrossRef]
- Nunes, M.A.; Pimentel, F.B.; Costa, A.S.G.; Alves, R.C.; Oliveira, M.B.P.P. Olive By-Products for Functional and Food Applications: Challenging Opportunities to Face EnvI/Ronmental Constraints. Innov. Food Sci. Emerg. Technol. 2016, 35, 139–148. [Google Scholar] [CrossRef]
- Mallamaci, R.; Budriesi, R.; Clodoveo, M.L.; Biotti, G.; Micucci, M.; Ragusa, A.; Curci, F.; Muraglia, M.; Corbo, F.; Franchini, C. Olive Tree in CI/Rcular Economy as a Source of Secondary Metabolites Active for Human and Animal Health Beyond Oxidative Stress and Inflammation. Molecules 2021, 26, 1072. [Google Scholar] [CrossRef] [PubMed]
- Marković, A.K.; Torić, J.; Barbarić, M.; Brala, C.J. Hydroxytyrosol, Tyrosol and Derivatives and TheI/R Potential Effects on Human Health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef]
- Rigacci, S.; Stefani, M. Nutraceutical Properties of Olive Oil Polyphenols. An Itinerary from Cultured Cells through Animal Models to Humans. Int. J. Mol. Sci. 2016, 17, 843. [Google Scholar] [CrossRef] [PubMed]
- Fabiani, R. Anti-Cancer Properties of Olive Oil SecoI/Ridoid Phenols: A Systematic Review of in Vivo Studies. Food Funct. 2016, 7, 4145–4159. [Google Scholar] [CrossRef]
- Bitler, C.M.; Viale, T.M.; Damaj, B.; Crea, R. Hydrolyzed Olive Vegetation Water in Mice Has Anti-Inflammatory Activity. J. Nutr. 2005, 135, 1475–1479. [Google Scholar] [CrossRef]
- Ciriminna, R.; Meneguzzo, F.; Fidalgo, A.; Ilharco, L.M.; Pagliaro, M. Extraction, Benefits and Valorization of Olive Polyphenols. Eur. J. Lipid Sci. Technol. 2015, 118, 503–511. [Google Scholar] [CrossRef]
- The European Food Safety Authority. Scientific Opinion on the Substantiation of a Health Claim Related to Polyphenols in Olive and Maintenance of Normal Blood HDL Cholesterol Concentrations (ID 1639, Further Assessment) Pursuant to Article 13 of Regulation (EC) No 1924/2006. EFSA J. 2012, 10, 2848. [Google Scholar] [CrossRef]
- Zrelli, H.; Matsuoka, M.; Kitazaki, S.; Araki, M.; Kusunoki, M.; Zarrouk, M.; Miyazaki, H. Hydroxytyrosol Induces Proliferation and Cytoprotection against Oxidative Injury in Vascular Endothelial Cells: Role of Nrf2 Activation and HO-1 Induction. J. Agric. Food Chem. 2011, 59, 4473–4482. [Google Scholar] [CrossRef]
- Lamy, S.; Saad, A.B.; Zgheib, A.; Annabi, B. Olive Oil Compounds Inhibit the Paracrine Regulation of TNF-α-Induced Endothelial Cell Migration through Reduced Glioblastoma Cell Cyclooxygenase-2 Expression. J. Nutr. Biochem. 2015, 27, 136–145. [Google Scholar] [CrossRef]
- Calabriso, N.; Gnoni, A.; Stanca, E.; Cavallo, A.; Damiano, F.; Siculella, L.; Carluccio, M.A. Hydroxytyrosol Ameliorates Endothelial Function under Inflammatory Conditions by Preventing Mitochondrial Dysfunction. Oxidative Med. Cell. Longev. 2018, 2018, 9086947. [Google Scholar] [CrossRef]
- Evans, S.; Weinheimer, C.J.; Kovacs, A.; Williams, J.W.; Randolph, G.J.; Jiang, W.; Barger, P.M.; Mann, D.L. Ischemia Reperfusion Injury Provokes Adverse Left Ventricular Remodeling in Dysferlin-Deficient Hearts through a Pathway That Involves TIRAP Dependent Signaling. Sci. Rep. 2020, 10, 14129. [Google Scholar] [CrossRef] [PubMed]
- Chai, X.; Liang, Z.; Zhang, J.; Ding, J.; Zhang, Q.; Lv, S.; Deng, Y.; Zhang, R.; Lu, D. Chlorogenic Acid Protects against Myocardial Ischemia–Reperfusion Injury in Mice by Inhibiting Lnc Neat1/NLRP3 Inflammasome-Mediated Pyroptosis. Sci. Rep. 2023, 13, 17803. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Liu, L.; Pan, H.; Ma, Y.; Wang, D.; Kang, K.; Wang, J.; Sun, B.; Sun, X.; Jiang, H. Protective Effects of Hydroxytyrosol on Liver Ischemia/Reperfusion Injury in Mice. Mol. Nutr. Food Res. 2013, 57, 1218–1227. [Google Scholar] [CrossRef]
- Fan, L.; Zhao, L.; Zhu, Y.; Li, L.; Yang, X.; Ma, P.; Liu, J.; Zhao, Q.; Li, X. Hydroxytyrosol Ameliorates Stress-Induced Liver Injury through Activating Autophagy via HDAC1/2 Inhibition. Food Funct. 2024, 15, 5103–5117. [Google Scholar] [CrossRef]
- Pei, Y.-H.; Chen, J.; Xie, L.; Cai, X.-M.; Yang, R.-H.; Wang, X.; Gong, J.-B. Hydroxytyrosol Protects against Myocardial Ischemia/Reperfusion Injury through a PI3K/Akt-Dependent Mechanism. Mediat. Inflamm. 2016, 2016, 1232103. [Google Scholar] [CrossRef]
- Fernández-Bolaños, J.; Rodríguez-Gutiérrez, G.; Lama-Muñoz, A.; Rubio-Senent, F.; Fernández-Bolaños, J.M.; Maya, I.; López, O.; Marset, A. Method for Obtaining Hydroxytyrosol Extract, Mixture of Hydroxytyrosol and 3,4-Dihydroyphenylglycol Extract, and Hydroxytyrosol Acetate Extract, from By-Products of the Olive Tree, and the Purification Thereof. WO Patent 2013/007850, 17 January 2013. [Google Scholar]
- D’Annunzio, V.; Perez, V.; Mazo, T.; Muñoz, M.C.; Dominici, F.P.; Carreras, M.C.; Poderoso, J.J.; Sadoshima, J.; Gelpi, R.J. Loss of Myocardial Protection against Myocardial Infarction in Middle-Aged Transgenic Mice Overexpressing Cardiac Thioredoxin-1. Oncotarget 2016, 7, 11889–11898. [Google Scholar] [CrossRef]
- Perez, V.; Zaobornyj, T.; Vico, T.; Vanasco, V.; Marchini, T.; Godoy, E.; Alvarez, S.; Evelson, P.; Donato, M.; Gelpi, R.J.; et al. Middle-Age Abolishes Cardioprotection Conferred by Thioredoxin-1 in Mice. Arch. Biochem. Biophys. 2024, 753, 109880. [Google Scholar] [CrossRef]
- Boveris, A.; Arnaiz, S.L.; Bustamante, J.; Alvarez, S.; Valdez, L.; Boveris, A.D.; Navarro, A. Pharmacological Regulation of Mitochondrial Nitric Oxide Synthase. In Methods in Enzymology on CD-ROM/Methods in Enzymology; Academic Press: New York, NY, USA, 2002; pp. 328–339. [Google Scholar] [CrossRef]
- Boveris, A.; Costa, L.E.; Cadenas, E.; Poderoso, J.J. Regulation of Mitochondrial Respiration by Adenosine Diphosphate, Oxygen, and Nitric Oxide. In Methods in Enzymology on CD-ROM/Methods in Enzymology; Academic Press: New York, NY, USA, 1999; pp. 188–198. [Google Scholar] [CrossRef]
- Boveris, A. [57] Determination of the Production of Superoxide Radicals and Hydrogen Peroxide in Mitochondria. In Methods in Enzymology on CD-ROM/Methods in Enzymology; Academic Press: New York, NY, USA, 1984; pp. 429–435. [Google Scholar] [CrossRef]
- Scaduto, R.C.; Grotyohann, L.W. Measurement of Mitochondrial Membrane Potential Using Fluorescent Rhodamine Derivatives. Biophys. J. 1999, 76, 469–477. [Google Scholar] [CrossRef]
- Navarro, A.; Torrejón, R.; Bández, M.J.; López-Cepero, J.M.; Boveris, A. Mitochondrial Function and Mitochondria-Induced Apoptosis in an Overstimulated Rat Ovarian Cycle. AJP Endocrinol. Metab. 2005, 289, E1101–E1109. [Google Scholar] [CrossRef]
- Visioli, F.; De la Lastra, C.A.; Andres-Lacueva, C.; Aviram, M.; Calhau, C.; Cassano, A.; D’Archivio, M.; Faria, A.; Favé, G.; Fogliano, V.; et al. Polyphenols and Human Health: A Prospectus. Crit. Rev. Food Sci. Nutr. 2011, 51, 524–546. [Google Scholar] [CrossRef]
- Gibellini, L.; Bianchini, E.; De Biasi, S.; Nasi, M.; Cossarizza, A.; Pinti, M. Natural Compounds Modulating Mitochondrial Functions. Evid.-Based Complement. Altern. Med. 2015, 2015, 527209. [Google Scholar] [CrossRef] [PubMed]
- Dudylina, A.L.; Ivanova, M.V.; Shumaev, K.B.; Ruuge, E.K. Superoxide Formation in Cardiac Mitochondria and Effect of Phenolic Antioxidants. Cell Biochem. Biophys. 2018, 77, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.A.; Maalik, A.; Murtaza, G. Inhibitory Mechanism against Oxidative Stress of Caffeic Acid. J. Food Drug Anal. 2016, 24, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Landoni, G.; Guarracino, F.; Cariello, C.; Franco, A.; Baldassarri, R.; Borghi, G.; Covello, R.D.; Gerli, C.; Crivellari, M.; Zangrillo, A. Volatile Compared with Total Intravenous Anaesthesia in Patients Undergoing High-Risk Cardiac Surgery: A Randomized Multicentre Study. Br. J. Anaesth. 2014, 113, 955–963. [Google Scholar] [CrossRef]
- Hofland, J.; Ouattara, A.; Fellahi, J.-L.; Gruenewald, M.; Hazebroucq, J.; Ecoffey, C.; Joseph, P.; Heringlake, M.; Steib, A.; Coburn, M.; et al. Effect of Xenon Anesthesia Compared to Sevoflurane and Total Intravenous Anesthesia for Coronary Artery Bypass Graft Surgery on Postoperative Cardiac Troponin Release. Anesthesiology 2017, 127, 918–933. [Google Scholar] [CrossRef]
- Ferdinandy, P.; Andreadou, I.; Baxter, G.F.; Bøtker, H.E.; Davidson, S.M.; Dobrev, D.; Gersh, B.J.; Heusch, G.; Lecour, S.; Ruiz-Meana, M.; et al. Interaction of Cardiovascular Nonmodifiable Risk Factors, Comorbidities and Comedications With Ischemia/Reperfusion Injury and Cardioprotection by Pharmacological Treatments and Ischemic Conditioning. Pharmacol. Rev. 2022, 75, 159–216. [Google Scholar] [CrossRef]
- Penna, C.; Andreadou, I.; Aragno, M.; Beauloye, C.; Bertrand, L.; Lazou, A.; Falcão-Pires, I.; Bell, R.; Zuurbier, C.J.; Pagliaro, P.; et al. Effect of Hyperglycaemia and Diabetes on Acute Myocardial Ischaemia–Reperfusion Injury and Cardioprotection by Ischaemic Conditioning Protocols. Br. J. Pharmacol. 2020, 177, 5312–5335. [Google Scholar] [CrossRef]
- Wang, Q.; Zuurbier, C.J.; Huhn, R.; Torregroza, C.; Hollmann, M.W.; Preckel, B.; Van Den Brom, C.E.; Weber, N.C. Pharmacological Cardioprotection against Ischemia Reperfusion Injury—The Search for a Clinical Effective Therapy. Cells 2023, 12, 1432. [Google Scholar] [CrossRef]
- Meng, Z.; Zhang, Z.; Zhao, J.; Liu, C.; Yao, P.; Zhang, L.; Xie, D.; Lau, W.B.; Tsukuda, J.; Christopher, T.A.; et al. Nitrative Modification of Caveolin-3: A Novel Mechanism of Cardiac Insulin Resistance and a Potential Therapeutic Target Against Ischemic Heart Failure in Prediabetic Animals. Circulation 2023, 147, 1162–1179. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Yellon, D.M. Myocardial Ischemia-Reperfusion Injury: A Neglected Therapeutic Target. J. Clin. Investig. 2013, 123, 92–100. [Google Scholar] [CrossRef]
- Heusch, G. Critical Issues for the Translation of Cardioprotection. Circ. Res. 2017, 120, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez, B.; Heusch, G.; Ovize, M.; Van de Werf, F. Evolving Therapies for Myocardial Ischemia/Reperfusion Injury. J. Am. Coll. Cardiol. 2015, 65, 1454–1471. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Huang, Z.; Liu, S.; Li, X.; Jia, P.; Guo, Y.; Wu, N.; Jia, D. Hydroxytyrosol Protects against Myocardial Ischemia Reperfusion Injury by Inhibiting Mitochondrial Permeability Transition Pore Opening. Exp. Ther. Med. 2018, 17, 671–678. [Google Scholar] [CrossRef]
- Bruckbauer, A.; Zemel, M.B. Synergistic Effects of Polyphenols and Methylxanthines with Leucine on AMPK/SI/Rtuin-Mediated Metabolism in Muscle Cells and Adipocytes. PLoS ONE 2014, 9, e89166. [Google Scholar] [CrossRef]
- Torres, A.; Noriega, L.G.; Delgadillo-Puga, C.; Tovar, A.R.; Navarro-Ocaña, A. Caffeoylquinic Acid Derivatives of Purple Sweet Potato as Modulators of Mitochondrial Function in Mouse Primary Hepatocytes. Molecules 2021, 26, 319. [Google Scholar] [CrossRef]
- Ozbek, N.; Bali, E.B.; Karasu, C. Quercetin and Hydroxytyrosol Attenuates Xanthine/Xanthine Oxidase-Induced Toxicity in H9c2 Cardiomyocytes by Regulation of Oxidative Stress and Stress-Sensitive Signaling Pathways. Gen. Physiol. Biophys. 2015, 34, 407–414. [Google Scholar]
- Lagoa, R.; Graziani, I.; Lopez-Sanchez, C.; Garcia-Martinez, V.; Gutierrez-Merino, C. Complex I and Cytochrome c Are Molecular Targets of Flavonoids That Inhibit Hydrogen Peroxide Production by Mitochondria. Biochim. Et Biophys. Acta (BBA)-Bioenerg. 2011, 1807, 1562–1572. [Google Scholar] [CrossRef]
- Xing, S.; Yang, X.; Li, W.; Bian, F.; Wu, D.; Chi, J.; Xu, G.; Zhang, Y.; Jin, S. Salidroside Stimulates Mitochondrial Biogenesis and Protects against H2O2-Induced Endothelial Dysfunction. Oxidative Med. Cell. Longev. 2014, 2014, 904834. [Google Scholar] [CrossRef]
- Dong, Y.; Yu, M.; Wu, Y.; Xia, T.; Wang, L.; Song, K.; Zhang, C.; Lu, K.; Rahimnejad, S. Hydroxytyrosol Promotes the Mitochondrial Function through Activating Mitophagy. Antioxidants 2022, 11, 893. [Google Scholar] [CrossRef]
- Hao, J.; Shen, W.; Yu, G.; Jia, H.; Li, X.; Feng, Z.; Wang, Y.; Weber, P.; Wertz, K.; Sharman, E. Hydroxytyrosol Promotes Mitochondrial Biogenesis and Mitochondrial Function in 3T3-L1 Adipocytes. J. Nutr. Biochem. 2009, 21, 634–644. [Google Scholar] [CrossRef]
- Adlam, V.J.; Harrison, J.C.; Porteous, C.M.; James, A.M.; Smith, R.A.J.; Murphy, M.P.; Sammut, I.A. Targeting an Antioxidant to Mitochondria Decreases Cardiac Ischemia-reperfusion Injury. FASEB J. 2005, 19, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Mata, A.; Cadenas, S. The Antioxidant Transcription Factor Nrf2 in Cardiac Ischemia–Reperfusion Injury. Int. J. Mol. Sci. 2021, 22, 11939. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-M.; Calkins, M.J.; Chan, K.; Kan, Y.W.; Johnson, J.A. Identification of the NF-E2-Related Factor-2-Dependent Genes Conferring Protection against Oxidative Stress in Primary Cortical Astrocytes Using Oligonucleotide Microarray Analysis. J. Biol. Chem. 2003, 278, 12029–12038. [Google Scholar] [CrossRef]
- Cheng, L.; Jin, Z.; Zhao, R.; Ren, K.; Deng, C.; Yu, S. Resveratrol Attenuates Inflammation and Oxidative Stress Induced by Myocardial Ischemia-Reperfusion Injury: Role of Nrf2/ARE Pathway. Int. J. Clin. Exp. Med. 2015, 8, 10420–10428. [Google Scholar]
- Sahu, B.D.; Putcha, U.K.; Kuncha, M.; Rachamalla, S.S.; Sistla, R. Carnosic Acid Promotes Myocardial Antioxidant Response and Prevents Isoproterenol-Induced Myocardial Oxidative Stress and Apoptosis in Mice. Mol. Cell. Biochem. 2014, 394, 163–176. [Google Scholar] [CrossRef]
- Liu, X.; Gao, J.; Yan, Y.; Georgiou, E.A.; Lou, J.; Feng, M.; Zhang, X.; Gao, F.; Liu, J.; Kostakis, I.K.; et al. Mitochondria-Targeted Triphenylphosphonium-Hydroxytyrosol Prevents Lipotoxicity-Induced Endothelial Injury by Enhancing Mitochondrial Function and Redox Balance via Promoting FoxO1 and Nrf2 Nuclear Translocation and Suppressing Inflammation via Inhibiting P38/NF-кB Pathway. Antioxidants 2023, 12, 175. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bermúdez-Oria, A.; Godoy, E.; Pérez, V.; Musci Ferrari, C.; Donato, M.; Fernández-Bolaños, J.; Zaobornyj, T.; D’Annunzio, V. Unveiling the Cardioprotective Potential of Hydroxytyrosol: Insights from an Acute Myocardial Infarction Model. Antioxidants 2025, 14, 803. https://doi.org/10.3390/antiox14070803
Bermúdez-Oria A, Godoy E, Pérez V, Musci Ferrari C, Donato M, Fernández-Bolaños J, Zaobornyj T, D’Annunzio V. Unveiling the Cardioprotective Potential of Hydroxytyrosol: Insights from an Acute Myocardial Infarction Model. Antioxidants. 2025; 14(7):803. https://doi.org/10.3390/antiox14070803
Chicago/Turabian StyleBermúdez-Oria, Alejandra, Eugenia Godoy, Virginia Pérez, Camila Musci Ferrari, Martin Donato, Juan Fernández-Bolaños, Tamara Zaobornyj, and Verónica D’Annunzio. 2025. "Unveiling the Cardioprotective Potential of Hydroxytyrosol: Insights from an Acute Myocardial Infarction Model" Antioxidants 14, no. 7: 803. https://doi.org/10.3390/antiox14070803
APA StyleBermúdez-Oria, A., Godoy, E., Pérez, V., Musci Ferrari, C., Donato, M., Fernández-Bolaños, J., Zaobornyj, T., & D’Annunzio, V. (2025). Unveiling the Cardioprotective Potential of Hydroxytyrosol: Insights from an Acute Myocardial Infarction Model. Antioxidants, 14(7), 803. https://doi.org/10.3390/antiox14070803