
Inactivated Cells and Metabolites of Saccharomyces boulardii Alleviate Inflammation Damage in Caco-2 Monolayer Cells and Mice with Ulcerative Colitis


Abstract
1. Introduction
2. Materials and Methods
2.1. Main Reagents and Materials
2.2. S. Boulardii Metabolite Analysis
2.3. Preparation of Postbiotics
2.4. Caco-2 Cell Culture and Viability Assay
2.5. Measurement of Antioxidant Capacity
2.6. Measurement of Transepithelial Electrical Resistance (TEER)
2.7. Measurement of Alkaline Phosphatase (ALP) Activity
2.8. Animals and Animal Experiment Design
2.9. Histopathological Analysis
2.10. Measurement of Pro-Inflammatory Cytokine
2.11. Measurement of mRNA Expression
2.12. Measurement of Protein Expression
2.13. Analysis of Gut Microbiota
2.14. Statistical Analysis
3. Results
3.1. Main Components of S. Boulardii Metabolites
3.2. S. Boulardii Postbiotics Have Antioxidant Capacity
3.3. S. Boulardii Postbiotics Improve the TEER and ALP Activity of the Caco-2 Cell Monolayer
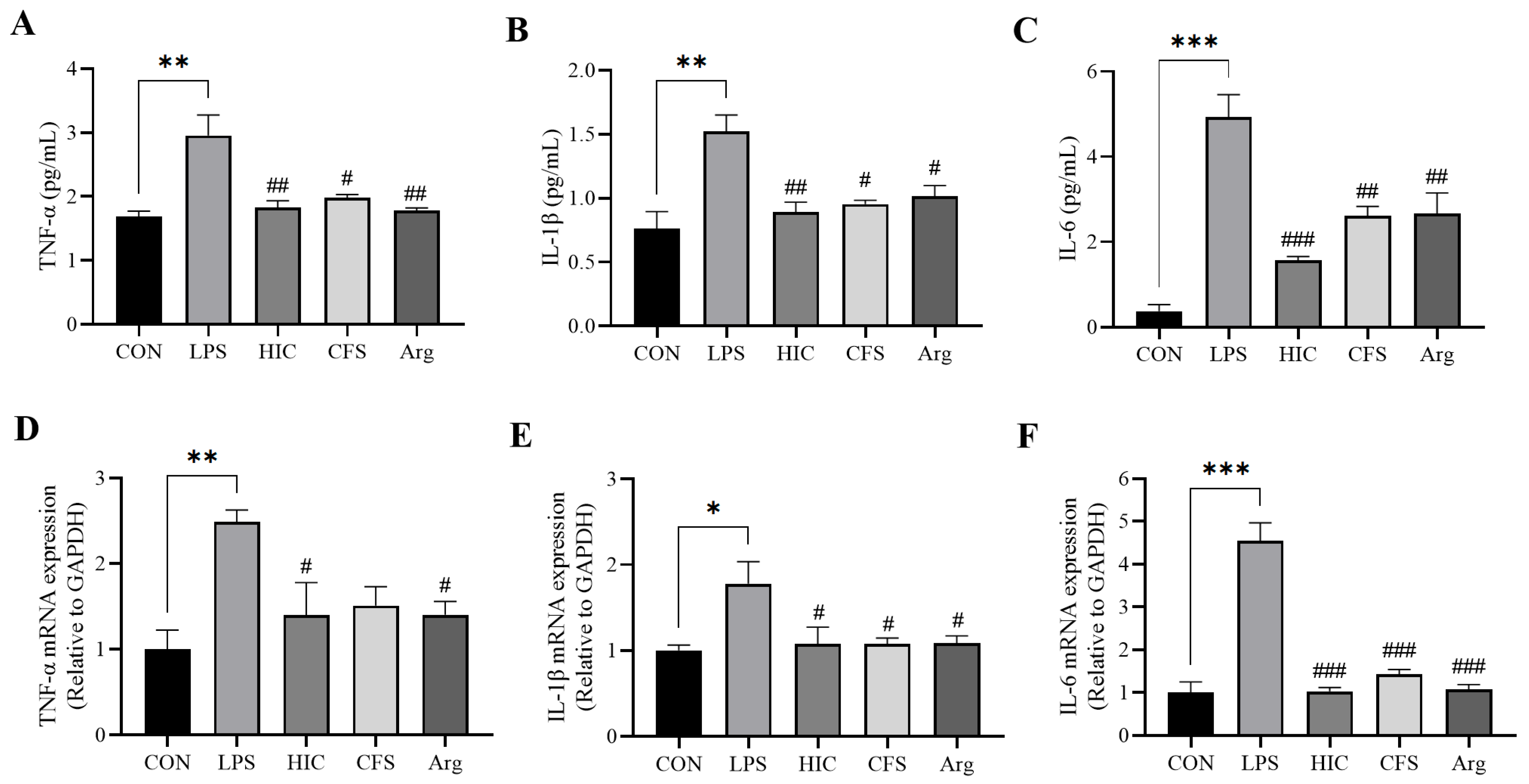
3.4. S. Boulardii Postbiotics Inhibit the Secretion of Pro-Inflammatory Cytokines by Caco-2 Cell Monolayers
3.5. S. Boulardii Postbiotics Protect TJPs of Caco-2 Cell Monolayers
3.6. S. Boulardii Postbiotics Alleviate the Conventional Pathological Indicators in Colitis Mice
3.7. S. Boulardii Postbiotics Improve the Colon Condition in Colitis Mice
3.8. S. Boulardii Postbiotics Inhibit Pro-Inflammatory Cytokines in Colitis Mice
3.9. S. Boulardii Postbiotics Protect TJPs in Colitis Mice
3.10. Effect of S. Boulardii Postbiotics on the Overall Structure of the Gut Microbiota in Colitis Mice
3.11. Effect of S. Boulardii Postbiotics on the Gut Microbiota at the Phylum and Genus Levels in Colitis Mice
3.12. Functional Prediction of Gut Microbiota Based on 16S rRNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| HIC | heat-inactivated cells |
| CFS | cell-free supernatant |
| Arg | L-arginine |
| SASP | sulfasalazine |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| ABTS | 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| LPS | lipopolysaccharide |
| DSS | dextran sulphate sodium |
| TNF-α | tumor necrosis factor alpha |
| IL-1β | interleukin 1 beta |
| IL-6 | interleukin 6 |
| ZO-1 | zonula occludens 1 |
| TJPs | tight junction proteins |
References
- Wang, R.; Li, Z.; Liu, S.; Zhang, D. Global, Regional and National Burden of Inflammatory Bowel Disease in 204 Countries and Territories from 1990 to 2019: A Systematic Analysis Based on the Global Burden of Disease Study 2019. BMJ Open 2023, 13, e065186. [Google Scholar] [CrossRef] [PubMed]
- Gros, B.; Kaplan, G.G. Ulcerative Colitis in Adults: A Review. JAMA 2023, 330, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, F.; Cannatelli, R.; Monico, M.C.; Maconi, G.; Ardizzone, S. An Update on Current Pharmacotherapeutic Options for the Treatment of Ulcerative Colitis. J. Clin. Med. 2022, 11, 2302. [Google Scholar] [CrossRef] [PubMed]
- Xia, P.; Hou, T.; Ma, M.; Li, S.; Jin, H.; Luo, X.; Li, J.; Geng, F.; Li, B. Konjac Oligosaccharides Attenuate DSS-Induced Ulcerative Colitis in Mice: Mechanistic Insights. Food Funct. 2022, 13, 5626–5639. [Google Scholar] [CrossRef]
- Hu, Y.; Jin, X.; Gao, F.; Lin, T.; Zhu, H.; Hou, X.; Yin, Y.; Kan, S.; Chen, D. Selenium-Enriched Bifidobacterium Longum DD98 Effectively Ameliorates Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice. Front. Microbiol. 2022, 13, 955112. [Google Scholar] [CrossRef]
- He, D.; Zeng, W.; Wang, Y.; Xing, Y.; Xiong, K.; Su, N.; Zhang, C.; Lu, Y.; Xing, X. Isolation and Characterization of Novel Peptides from Fermented Products of Lactobacillus for Ulcerative Colitis Prevention and Treatment. Food Sci. Hum. Wellness 2022, 11, 1464–1474. [Google Scholar] [CrossRef]
- Lazo-Velez, M.A.; Serna-Saldivar, S.O.; Rosales-Medina, M.F.; Tinoco-Alvear, M.; Briones-Garcia, M. Application of Saccharomyces cerevisiae Var. Boulardii in Food Processing: A Review. J. Appl. Microbiol. 2018, 125, 943–951. [Google Scholar] [CrossRef]
- de Souza, H.F.; Carosia, M.F.; Pinheiro, C.; de Carvalho, M.V.; Fernandes de Oliveira, C.A.; Kamimura, E.S. On Probiotic Yeasts in Food Development: Saccharomyces boulardii, a Trend. Food Sci. Technol. 2022, 42, e92321. [Google Scholar] [CrossRef]
- Chan, M.Z.A.; Liu, S.-Q. Fortifying Foods with Synbiotic and Postbiotic Preparations of the Probiotic Yeast, Saccharomyces boulardii. Curr. Opin. Food Sci. 2022, 43, 216–224. [Google Scholar] [CrossRef]
- Ansari, F.; Alian Samakkhah, S.; Bahadori, A.; Jafari, S.M.; Ziaee, M.; Khodayari, M.T.; Pourjafar, H. Health-Promoting Properties of Saccharomyces cerevisiae var. Boulardii as a Probiotic; Characteristics, Isolation, and Applications in Dairy Products. Crit. Rev. Food Sci. Nutr. 2023, 63, 457–485. [Google Scholar] [CrossRef]
- Gao, H.; Li, Y.; Xu, J.; Zuo, X.; Yue, T.; Xu, H.; Sun, J.; Wang, M.; Ye, T.; Yu, Y.; et al. Saccharomyces boulardii Protects against Murine Experimental Colitis by Reshaping the Gut Microbiome and Its Metabolic Profile. Front. Microbiol. 2023, 14, 1204122. [Google Scholar] [CrossRef]
- Li, B.; Zhang, H.; Shi, L.; Li, R.; Luo, Y.; Deng, Y.; Li, S.; Li, R.; Liu, Z. Saccharomyces boulardii Alleviates DSS-Induced Intestinal Barrier Dysfunction and Inflammation in Humanized Mice. Food Funct. 2022, 13, 102–112. [Google Scholar] [CrossRef]
- Liang, D.; Wu, F.; Zhou, D.; Tan, B.; Chen, T. Commercial Probiotic Products in Public Health: Current Status and Potential Limitations. Crit. Rev. Food Sci. Nutr. 2024, 64, 6455–6476. [Google Scholar] [CrossRef]
- Walhe, R.; Alim, H.; Kumari, S. From Probiotics to Postbiotics: Key to Microbiome and Health. In Microbiome-Gut-Brain Axis: Implications on Health; Sayyed, R.Z., Khan, M., Eds.; Springer Nature: Singapore, 2022; pp. 367–381. [Google Scholar]
- Harat, S.G.; Pourjafar, H. Health Benefits and Safety of Postbiotics Derived from Different Probiotic Species. Curr. Pharm. Des. 2025, 31, 116–127. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Xu, X.; Wu, J.; Jin, Y.; Huang, K.; Zhang, Y.; Liang, Z. Both Saccharomyces boulardii and Its Postbiotics Alleviate Dextran Sulfate Sodium-Induced Colitis in Mice, Association with Modulating Inflammation and Intestinal Microbiota. Nutrients 2023, 15, 1484. [Google Scholar] [CrossRef]
- Jin, Y.; Wu, J.; Huang, K.; Liang, Z. Heat-Killed Saccharomyces boulardii Alleviates Dextran Sulfate Sodium-Induced Ulcerative Colitis by Restoring the Intestinal Barrier, Reducing Inflammation, and Modulating the Gut Microbiota. Nutrients 2024, 16, 702. [Google Scholar] [CrossRef]
- Datta, S.; Timson, D.J.; Annapure, U.S. Antioxidant Properties and Global Metabolite Screening of the Probiotic Yeast Saccharomyces cerevisiae Var. boulardii. J. Sci. Food Agric. 2017, 97, 3039–3049. [Google Scholar] [CrossRef]
- Zhang, W.; Bao, C.; Wang, J.; Zang, J.; Cao, Y. Administration of Saccharomyces boulardii Mafic-1701 Improves Feed Conversion Ratio, Promotes Antioxidant Capacity, Alleviates Intestinal Inflammation and Modulates Gut Microbiota in Weaned Piglets. J. Anim. Sci. Biotechnol. 2020, 11, 112. [Google Scholar] [CrossRef]
- Fu, J.; Liu, J.; Wen, X.; Zhang, G.; Cai, J.; Qiao, Z.; An, Z.; Zheng, J.; Li, L. Unique Probiotic Properties and Bioactive Metabolites of Saccharomyces boulardii. Probiotics Antimicrob. Proteins 2023, 15, 967–982. [Google Scholar] [CrossRef]
- Yang, Y.J.; Kim, M.J.; Lee, H.J.; Lee, W.-Y.; Yang, J.-H.; Kim, H.H.; Shim, M.S.; Heo, J.W.; Son, J.D.; Kim, W.H.; et al. Ziziphus Jujuba Miller Ethanol Extract Restores Disrupted Intestinal Barrier Function via Tight Junction Recovery and Reduces Inflammation. Antioxidants 2024, 13, 575. [Google Scholar] [CrossRef]
- Yu, M.; Wang, Q.; Ma, Y.; Li, L.; Yu, K.; Zhang, Z.; Chen, G.; Li, X.; Xiao, W.; Xu, P.; et al. Aryl Hydrocarbon Receptor Activation Modulates Intestinal Epithelial Barrier Function by Maintaining Tight Junction Integrity. Int. J. Biol. Sci. 2018, 14, 69–77. [Google Scholar] [CrossRef]
- Gong, S.; Zheng, J.; Zhang, J.; Wang, Y.; Xie, Z.; Wang, Y.; Han, J. Taxifolin Ameliorates Lipopolysaccharide-Induced Intestinal Epithelial Barrier Dysfunction via Attenuating NF-Kappa B/MLCK Pathway in a Caco-2 Cell Monolayer Model. Food Res. Int. 2022, 158, 111502. [Google Scholar] [CrossRef]
- Yang, Q.; Xing, M.; Wang, K.; Wei, Q.; Zhao, J.; Wang, Y.; Ji, K.; Song, S. Application of Fucoidan in Caco-2 Model Establishment. Pharmaceuticals 2022, 15, 418. [Google Scholar] [CrossRef]
- Markov, P.A.; Volkova, M.V.; Khasanshina, Z.R.; Martinson, E.A.; Popov, S.V. Anti-inflammatory activity of high and low methoxylated apple pectins, in vivo and in vitro. Vopr. Pitan. 2021, 90, 92–100. [Google Scholar]
- Qu, C.; Yuan, Z.-W.; Yu, X.-T.; Huang, Y.-F.; Yang, G.-H.; Chen, J.-N.; Lai, X.-P.; Su, Z.-R.; Zeng, H.-F.; Xie, Y.; et al. Patchouli Alcohol Ameliorates Dextran Sodium Sulfate-Induced Experimental Colitis and Suppresses Tryptophan Catabolism. Pharmacol. Res. 2017, 121, 70–82. [Google Scholar] [CrossRef]
- Xuan-qing, C.H.E.N.; Xiang-yu, L.V.; Shi-jia, L.I.U. Baitouweng Decoction Alleviates Dextran Sulfate Sodium-Induced Ulcerative Colitis by Regulating Intestinal Microbiota and the IL-6/STAT3 Signaling Pathway. J. Ethnopharmacol. 2021, 265, 113357. [Google Scholar] [CrossRef]
- Laroui, H.; Ingersoll, S.A.; Liu, H.C.; Baker, M.T.; Ayyadurai, S.; Charania, M.A.; Laroui, F.; Yan, Y.; Sitaraman, S.V.; Merlin, D. Dextran Sodium Sulfate (DSS) Induces Colitis in Mice by Forming Nano-Lipocomplexes with Medium-Chain-Length Fatty Acids in the Colon. PLoS ONE 2012, 7, e32084. [Google Scholar] [CrossRef]
- Zeng, W.; He, D.; Xing, Y.; Liu, J.; Su, N.; Zhang, C.; Wang, Y.; Xing, X. Internal Connections between Dietary Intake and Gut Microbiota Homeostasis in Disease Progression of Ulcerative Colitis: A Review. Food Sci. Hum. Wellness 2021, 10, 119–130. [Google Scholar] [CrossRef]
- Quaglio, A.E.V.; Grillo, T.G.; Oliveira, E.C.S.D.; Stasi, L.C.D.; Sassaki, L.Y. Gut Microbiota, Inflammatory Bowel Disease and Colorectal Cancer. World J. Gastroenterol. 2022, 28, 4053–4060. [Google Scholar] [CrossRef]
- Le Berre, C.; Honap, S.; Peyrin-Biroulet, L. Ulcerative Colitis. Lancet 2023, 402, 571–584. [Google Scholar] [CrossRef]
- Lan, J.; Dou, X.; Li, J.; Yang, Y.; Xue, C.; Wang, C.; Gao, N.; Shan, A. L-Arginine Ameliorates Lipopolysaccharide-Induced Intestinal Inflammation through Inhibiting the TLR4/NF-κB and MAPK Pathways and Stimulating β-Defensin Expression in Vivo and in Vitro. J. Agric. Food Chem. 2020, 68, 2648–2663. [Google Scholar] [CrossRef]
- Mehandru, S.; Colombel, J.-F. The Intestinal Barrier, an Arbitrator Turned Provocateur in IBD. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 83–84. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal Mucosal Barrier Function in Health and Disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Lopez-Escalera, S.; Wellejus, A. Evaluation of Caco-2 and Human Intestinal Epithelial Cells as in Vitro Models of Colonic and Small Intestinal Integrity. Biochem. Biophys. Rep. 2022, 31, 101314. [Google Scholar] [CrossRef]
- Santos, G.M.; Ismael, S.; Morais, J.; Araújo, J.R.; Faria, A.; Calhau, C.; Marques, C. Intestinal Alkaline Phosphatase: A Review of This Enzyme Role in the Intestinal Barrier Function. Microorganisms 2022, 10, 746. [Google Scholar] [CrossRef]
- Fukui, K.; Hada, T.; Imanishi, H.; Iwasaki, A.; Hirano, K.; Higashino, K. A Fetal Intestinal-Type Alkaline Phosphatase Produced in Caco-2 Cells. Tumor Biol. 1996, 17, 320–324. [Google Scholar] [CrossRef]
- Matsumoto, H.; Erickson, R.; Gum, J.; Yoshioka, M.; Gum, E.; Kim, Y. Biosynthesis of Alkaline-Phosphatase During Differentiation of the Human Colon Cancer Cell-Line Caco-2. Gastroenterology 1990, 98, 1199–1207. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, J.; Fujita, Y.; Shinno-Hashimoto, H.; Shan, J.; Wan, X.; Qu, Y.; Chang, L.; Wang, X.; Hashimoto, K. Effects of Spleen Nerve Denervation on Depression-like Phenotype, Systemic Inflammation, and Abnormal Composition of Gut Microbiota in Mice after Administration of Lipopolysaccharide: A Role of Brain-Spleen Axis. J. Affect. Disord. 2022, 317, 156–165. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, T.; Liao, L.; Fan, X.; Chang, L.; Hashimoto, K. Brain-Spleen Axis in Health and Diseases: A Review and Future Perspective. Brain Res. Bull. 2022, 182, 130–140. [Google Scholar] [CrossRef]
- Youn, H.-Y.; Kim, H.-J.; Kim, H.; Seo, K.-H. A Comparative Evaluation of the Kefir Yeast Kluyveromyces marxianus A4 and Sulfasalazine in Ulcerative Colitis: Anti-Inflammatory Impact and Gut Microbiota Modulation. Food Funct. 2024, 15, 6717–6730. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, S.; Ono, K.; Mikami, Y.; Naganuma, M.; Fukuda, T.; Minami, K.; Masaoka, T.; Terada, S.; Yoshida, T.; Saigusa, K.; et al. 5-Aminosalicylic Acid Intolerance Is Associated with a Risk of Adverse Clinical Outcomes and Dysbiosis in Patients with Ulcerative Colitis. Intest. Res. 2020, 18, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Sasahira, M.; Go, T.T.; Yo, S.; Ninomiya, T.; Osawa, M.; Handa, O.; Umegami, E.; Inoue, R.; Shiotani, A. Characteristics of Mucosa-Associated Microbiota in Ulcerative Colitis Patients with 5-Aminosalicylic Acid Intolerance. Biomedicines 2024, 12, 2125. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Han, R.; Yuan, Y.; Xing, Y.; Zhang, W.; Sun, Z.; Liu, Y.; Li, J.; Mao, T. The Role of Akkermansia Muciniphila in Inflammatory Bowel Disease: Current Knowledge and Perspectives. Front. Immunol. 2023, 13, 1089600. [Google Scholar] [CrossRef]
- Jadiya, S.; Upmanyu, N.; Sathiyanarayanan, A.; Jain, V.; Dubey, R.; Buwade, P. Formulation and Development of Novel Sulfasalazine Bilayer Tablets for The Treatment of Arthritis Associated With IBD: In-Vitro and In-Vivo Investigations. J. Pharm. Sci. 2024, 113, 1919–1926. [Google Scholar] [CrossRef]
- Ye, W.; Ding, Y.; Li, M.; Tian, Z.; Wang, S.; Liu, Z. Safety Assessment of Sulfasalazine: A Pharmacovigilance Study Based on FAERS Database. Front. Pharmacol. 2024, 15, 1452300. [Google Scholar] [CrossRef]
- Baske, M.M.; Timmerman, K.C.; Garmo, L.G.; Freitas, M.N.; Mccollum, K.A.; Ren, T.Y. Fecal Microbiota Transplant on Escherichia-Shigella Gut Composition and Its Potential Role in the Treatment of Generalized Anxiety Disorder: A Systematic Review. J. Affect. Disord. 2024, 354, 309–317. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria with Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, L.; Wang, X.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Roles of Intestinal Parabacteroides in Human Health and Diseases. FEMS Microbiol. Lett. 2022, 369, fnac072. [Google Scholar] [CrossRef]
- Wei, W.; Wong, C.C.; Jia, Z.; Liu, W.; Liu, C.; Ji, F.; Pan, Y.; Wang, F.; Wang, G.; Zhao, L.; et al. Parabacteroides Distasonis Uses Dietary Inulin to Suppress NASH via Its Metabolite Pentadecanoic Acid. Nat. Microbiol. 2023, 8, 1534–1548. [Google Scholar] [CrossRef]
- Zou, J.; Shen, Y.; Chen, M.; Zhang, Z.; Xiao, S.; Liu, C.; Wan, Y.; Yang, L.; Jiang, S.; Shang, E.; et al. Lizhong Decoction Ameliorates Ulcerative Colitis in Mice via Modulating Gut Microbiota and Its Metabolites. Appl. Microbiol. Biotechnol. 2020, 104, 5999–6012. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Jiang, C.; Lu, H.; Zhou, T.; Hu, W.; Tan, C.P.; Feng, Y.; Shen, G.; Xiang, X.; Chen, L. Camellia Oil Alleviates DSS-Induced Colitis in Mice by Regulating the Abundance of Intestinal Flora and Suppressing the NF-κB Signaling Pathway. J. Funct. Food. 2023, 108, 105777. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance Name | Molecular Weight | RT [min] | Relative Quantitative | |
|---|---|---|---|---|
| 1 | L-Arginine | 174 | 8.112 | 543.15 |
| 2 | L-Lysine | 146 | 1.286 | 371.75 |
| 3 | L-Tyrosine | 181 | 2.155 | 356.99 |
| 4 | L-Methionine | 149 | 1.943 | 209.13 |
| 5 | L-Phenylalanine | 165 | 4.803 | 142.42 |
| 6 | L-Tryptophan | 204 | 0.577 | 10.23 |
| 7 | L-Asparagine | 132 | 1.34 | 2.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Y.; Niu, Z.; Feng, M.; Che, H.; Liang, Z. Inactivated Cells and Metabolites of Saccharomyces boulardii Alleviate Inflammation Damage in Caco-2 Monolayer Cells and Mice with Ulcerative Colitis. Antioxidants 2025, 14, 737. https://doi.org/10.3390/antiox14060737
Jin Y, Niu Z, Feng M, Che H, Liang Z. Inactivated Cells and Metabolites of Saccharomyces boulardii Alleviate Inflammation Damage in Caco-2 Monolayer Cells and Mice with Ulcerative Colitis. Antioxidants. 2025; 14(6):737. https://doi.org/10.3390/antiox14060737
Chicago/Turabian StyleJin, Yuxin, Zehui Niu, Menglin Feng, Huilian Che, and Zhihong Liang. 2025. "Inactivated Cells and Metabolites of Saccharomyces boulardii Alleviate Inflammation Damage in Caco-2 Monolayer Cells and Mice with Ulcerative Colitis" Antioxidants 14, no. 6: 737. https://doi.org/10.3390/antiox14060737
APA StyleJin, Y., Niu, Z., Feng, M., Che, H., & Liang, Z. (2025). Inactivated Cells and Metabolites of Saccharomyces boulardii Alleviate Inflammation Damage in Caco-2 Monolayer Cells and Mice with Ulcerative Colitis. Antioxidants, 14(6), 737. https://doi.org/10.3390/antiox14060737