

ROMO1: A Distinct Mitochondrial Protein with Dual Roles in Dynamics and Function



{kind=link}
Abstract
1. Introduction
2. Structural and Molecular Characteristics
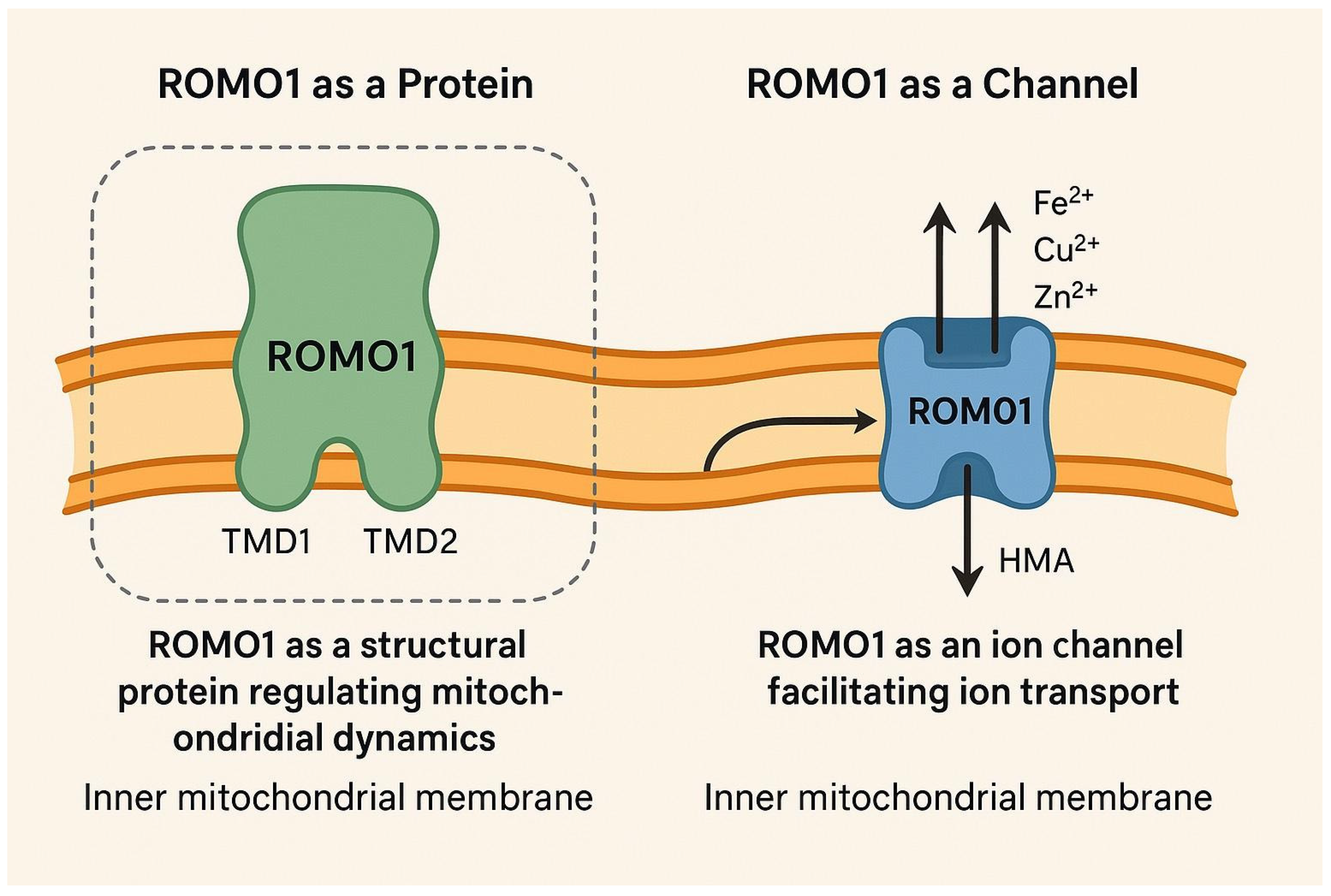
2.1. ROMO1 as a Protein
2.2. ROMO1 as a Channel
3. Physiological Roles
3.1. ROS Production and Regulation
3.2. Mitochondrial Dynamics
4. Pathophysiological Implications
4.1. Cancer
4.2. Neurodegenerative Diseases
4.3. Cardiovascular Diseases
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, G.Y.; You, D.G.; Lee, H.R.; Hwang, S.W.; Lee, C.J.; Yoo, Y.D. Romo1 is a mitochondrial nonselective cation channel with viroporin-like characteristics. J. Cell Biol. 2018, 217, 2059–2071. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chung, J.S.; Lee, S.B.; Park, S.H.; Kang, S.T.; Na, A.R.; Chang, T.S.; Kim, H.J.; Yoo, Y.D. Mitochondrial reactive oxygen species originating from Romo1 exert an important role in normal cell cycle progression by regulating p27(Kip1) expression. Free Radic. Res. 2009, 43, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Alfadda, A.A.; Sallam, R.M. Reactive oxygen species in health and disease. J. Biomed. Biotechnol. 2012, 2012, 936486. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kim, J.J.; Lee, S.B.; Park, J.K.; Yoo, Y.D. TNF-alpha-induced ROS production triggering apoptosis is directly linked to Romo1 and Bcl-X(L). Cell Death Differ. 2010, 17, 1420–1434. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.A.; Chung, J.S.; Cho, S.H.; Kim, H.J.; Yoo, Y.D. Romo1 expression contributes to oxidative stress-induced death of lung epithelial cells. Biochem. Biophys. Res. Commun. 2013, 439, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Norton, M.; Ng, A.C.; Baird, S.; Dumoulin, A.; Shutt, T.; Mah, N.; Andrade-Navarro, M.A.; McBride, H.M.; Screaton, R.A. ROMO1 is an essential redox-dependent regulator of mitochondrial dynamics. Sci. Signal. 2014, 7, ra10. [Google Scholar] [CrossRef] [PubMed]
- Tsoneva, E.; Vasileva-Slaveva, M.B.; Kostov, S.G.; Yordanov, A.D. ROMO1—A potential immunohistochemical prognostic marker for cancer development. Oncologie 2023, 25, 753–758. [Google Scholar] [CrossRef]
- Kim, H.J.; Jo, M.J.; Kim, B.R.; Kim, J.L.; Jeong, Y.A.; Na, Y.J.; Park, S.H.; Lee, S.-Y.; Lee, D.-H.; Lee, H.S.; et al. Reactive oxygen species modulator-1 (ROMO1) predicts unfavorable prognosis in colorectal cancer patients. PLoS ONE 2017, 12, e0176834. [Google Scholar] [CrossRef]
- Kim, T.; Cha, Y.J.; Park, J.H.; Kim, A.; Choi, Y.J.; Park, H.J. Reactive oxygen species modulator 1 expression predicts lymph node metastasis and survival in early-stage non-small cell lung cancer. PLoS ONE 2020, 15, e0239670. [Google Scholar] [CrossRef]
- Chung, J.S.; Park, S.; Park, S.H.; Park, E.R.; Cha, P.H.; Kim, B.Y.; Chung, Y.M.; Woo, S.R.; Han, C.J.; Kim, S.B.; et al. Overexpression of Romo1 promotes production of reactive oxygen species and invasiveness of hepatic tumor cells. Gastroenterology 2012, 143, 1084–1094.e7. [Google Scholar] [CrossRef]
- Bansal, S.; Biswas, G.; Avadhani, N.G. Mitochondria-targeted heme oxygenase-1 induces oxidative stress and mitochondrial dysfunction in macrophages, kidney fibroblasts and in chronic alcohol hepatotoxicity. Redox Biol. 2013, 2, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552 Pt 2, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Cowan, K.; Anichtchik, O.; Luo, S. Mitochondrial integrity in neurodegeneration. CNS Neurosci. Ther. 2019, 25, 825–836. [Google Scholar] [CrossRef]
- Chung, Y.M.; Lee, S.B.; Kim, H.J.; Park, S.H.; Kim, J.J.; Chung, J.S.; Yoo, Y.D. Replicative senescence induced by Romo1-derived reactive oxygen species. J. Biol. Chem. 2008, 283, 33763–33771. [Google Scholar] [CrossRef]
- Lynch, D.S.; Loh, S.H.Y.; Harley, J.; Noyce, A.J.; Martins, L.M.; Wood, N.W.; Houlden, H.; Plun-Favreau, H. Nonsyndromic Parkinson disease in a family with autosomal dominant optic atrophy due to OPA1 mutations. Neurol. Genet. 2017, 3, e188. [Google Scholar] [CrossRef]
- Martens, M.D.; Holody, C.D.; Wells, L.; Silver, H.L.; Morales-Llamas, D.Y.; Du, W.W.; Reeks, C.; Khairy, M.; Chen, H.; Ferdaoussi, M.; et al. Reactive Oxygen Species Modulator 1 Plays an Obligate Role in Cardiomyocyte Hypertrophy. Circ. Res. 2024, 134, 114–116. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Sun, M.H.; Lee, S.H.; Cui, X.S. ROMO1 is required for mitochondrial metabolism during preimplantation embryo development in pigs. Cell Div. 2021, 16, 7. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, F.; Wang, H.; Chen, H.; Guo, J.; Dang, X.; Ru, Y.; Wang, H. Mechanism of Ferroptosis and Its Role in Spinal Cord Injury. Front. Neurol. 2022, 13, 926780. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lee, S.B.; Kim, J.J.; Kim, T.W.; Kim, B.S.; Lee, M.S.; Yoo, Y.D. Serum deprivation-induced reactive oxygen species production is mediated by Romo1. Apoptosis 2010, 15, 204–218. [Google Scholar] [CrossRef] [PubMed]
- Na, A.R.; Chung, Y.M.; Lee, S.B.; Park, S.H.; Lee, M.S.; Yoo, Y.D. A critical role for Romo1-derived ROS in cell proliferation. Biochem. Biophys. Res. Commun. 2008, 369, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Sandamalika, W.M.G.; Udayantha, H.M.V.; Liyanage, D.S.; Lim, C.; Kim, G.; Kwon, H.; Lee, J. Identification of reactive oxygen species modulator 1 (Romo 1) from black rockfish (Sebastes schlegelii) and deciphering its molecular characteristics, immune responses, oxidative stress modulation, and wound healing properties. Fish Shellfish Immunol. 2022, 125, 266–275. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yordanov, A.; Tsoneva, E. ROMO1: A Distinct Mitochondrial Protein with Dual Roles in Dynamics and Function. Antioxidants 2025, 14, 540. https://doi.org/10.3390/antiox14050540
Yordanov A, Tsoneva E. ROMO1: A Distinct Mitochondrial Protein with Dual Roles in Dynamics and Function. Antioxidants. 2025; 14(5):540. https://doi.org/10.3390/antiox14050540
Chicago/Turabian StyleYordanov, Angel, and Eva Tsoneva. 2025. "ROMO1: A Distinct Mitochondrial Protein with Dual Roles in Dynamics and Function" Antioxidants 14, no. 5: 540. https://doi.org/10.3390/antiox14050540
APA StyleYordanov, A., & Tsoneva, E. (2025). ROMO1: A Distinct Mitochondrial Protein with Dual Roles in Dynamics and Function. Antioxidants, 14(5), 540. https://doi.org/10.3390/antiox14050540