
Oxidation-Induced Mixed Disulfide and Cataract Formation: A Review



Abstract
1. Introduction
2. Senile Cataract
3. Glutathione and Protein–Thiol Mixed Disulfides in the Lens
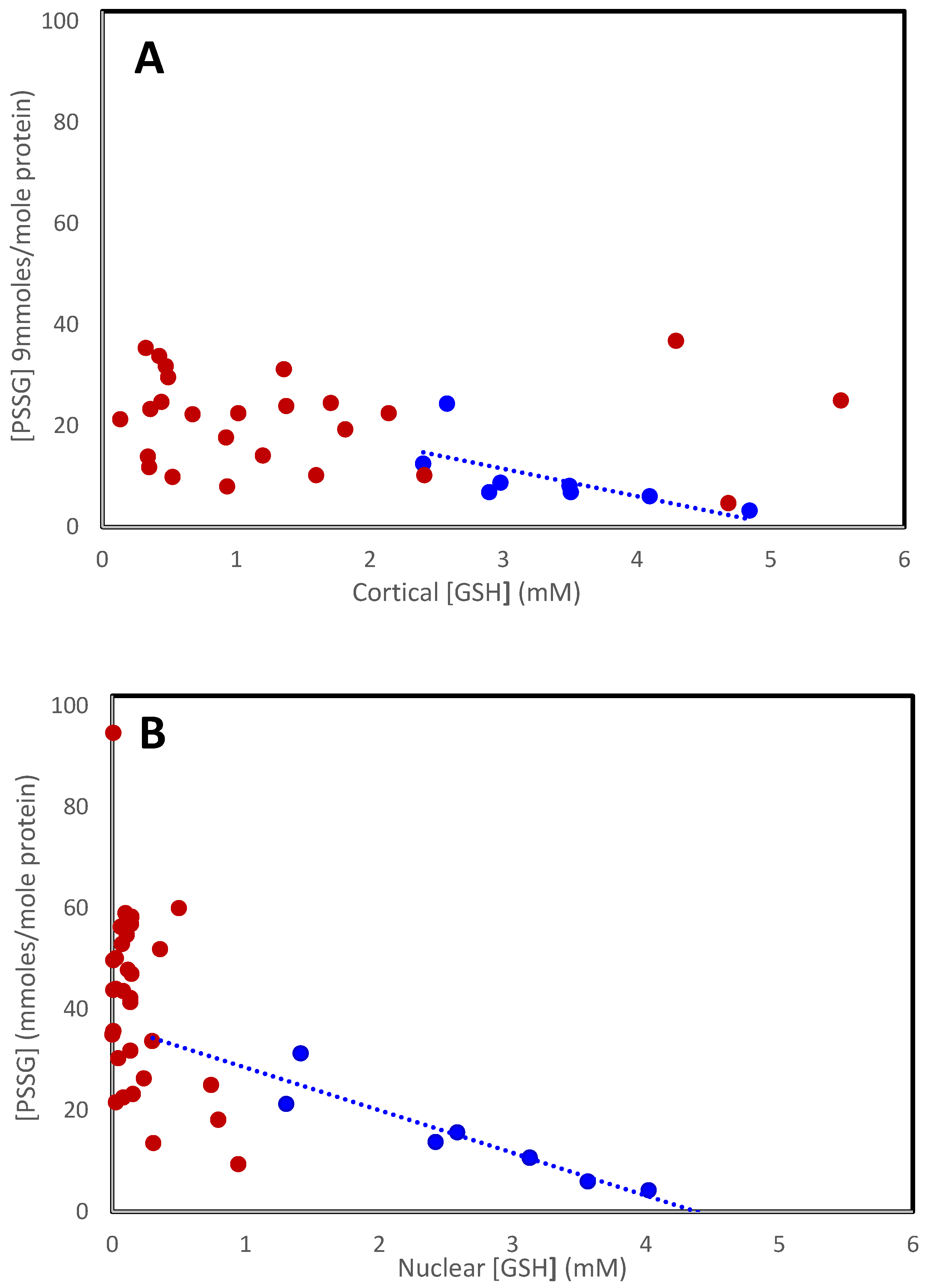
3.1. Mixed Disulfides in the Normal Human Lens with Age

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Lens | Cataractous Lens | |||||||
|---|---|---|---|---|---|---|---|---|
| Age | Cortex | Nucleus | Whole | Type | Cortex | Nucleus | Whole | |
| Lou and Dickerson [12] | 19–21 | 16 | ||||||
| <10 | 3 | 4 | 5 | |||||
| 11–80 | 3–24 | 4–60 | 40 | |||||
| Truscott and Augusteyn [13] | 45–62 | 3.5 | 4.4 | I | 10 | 23 | ||
| II | 23 | 49 | ||||||
| III | 25 | 44 | ||||||
| IV | 26 | 48 | ||||||
| Lou, Huang and Zigler [33] | na | 16 | Mixed | 44 | ||||
| na | 18 | Mixed | 32 | |||||
| 61 | 25 | IV | 49 | |||||
| 98 | 16 | IV | 53 | |||||
| Lou, Dickerson, Tung, Wolfe, Chylack [39] | 87–89 | 14 | I | 18 | ||||
| II | 25 | |||||||
| III | 52 | |||||||
| IV | 57 | |||||||
| Normal Lens | Cataractous Lens | |||||||
|---|---|---|---|---|---|---|---|---|
| Age | Cortex | Nucleus | Whole | Type | Cortex | Nucleus | Whole | |
| Lou and Dickerson [12] | 19–21 | 2.5 | ||||||
| <10 | 1.1 | 1.3 | 1.1 | |||||
| 11–80 | 1.4–5.6 | 1.8–9.4 | 6.3 | |||||
| Lou, Huang and Zigler [33] | na | 4.6 | Mixed | 11.1 | ||||
| 61 | 3.2 | IV | 58 | |||||
| 98 | 8.5 | IV | 26 | |||||
| Lou, Dickerson, Tung, Wolfe, Chylack [39] | 87–89 | 5 | I | 8 | ||||
| II | 8.8 | |||||||
| III | 17.5 | |||||||
| IV | 25.6 | |||||||
3.2. A Possible Function of Mixed Disulfides in the Lens
3.3. Mixed Disulfides and Cataracts
4. The Nature of the Oxidant Responsible for Cataracts
5. The Possible Role of Mixed Disulfides in Triggering Cataract Formation
6. Glutathionylation of Lens Protein Causes Conformational Changes
7. Model Studies on the Formation of PSSG in Lens Proteins During Oxidative Stress
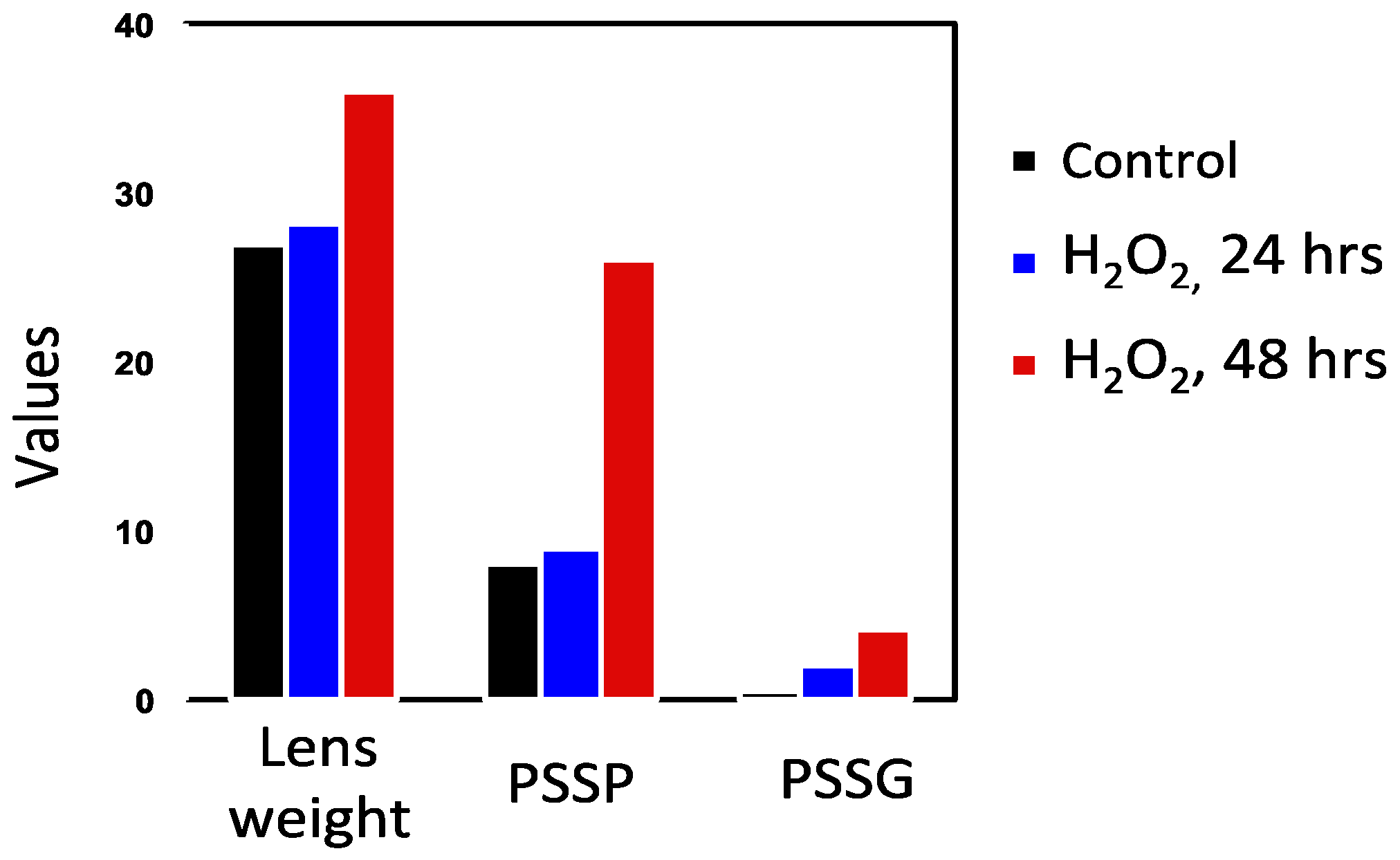
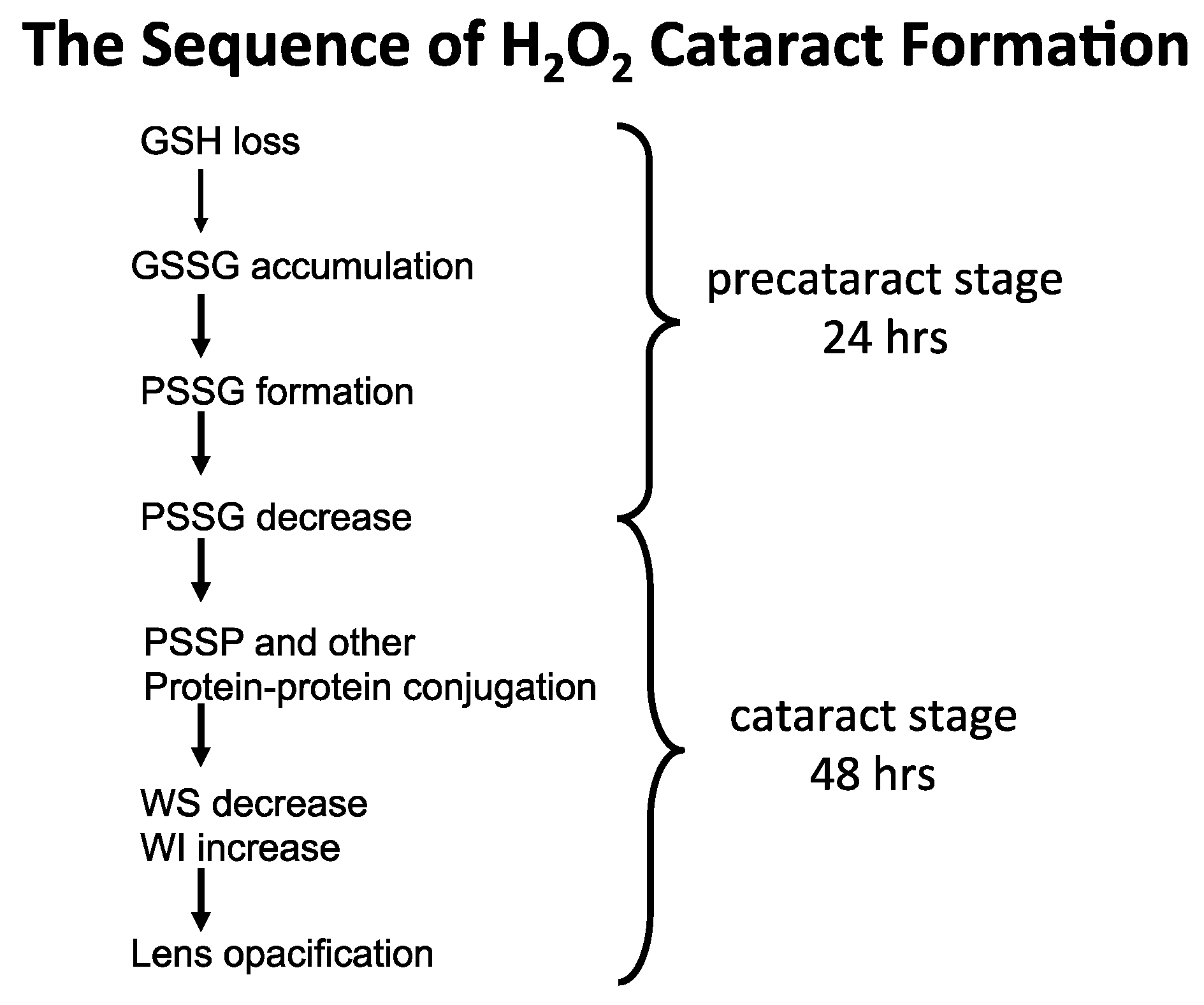
7.1. Rat Lens Culture: H2O2 Exposure and Post-Oxidation Recovery
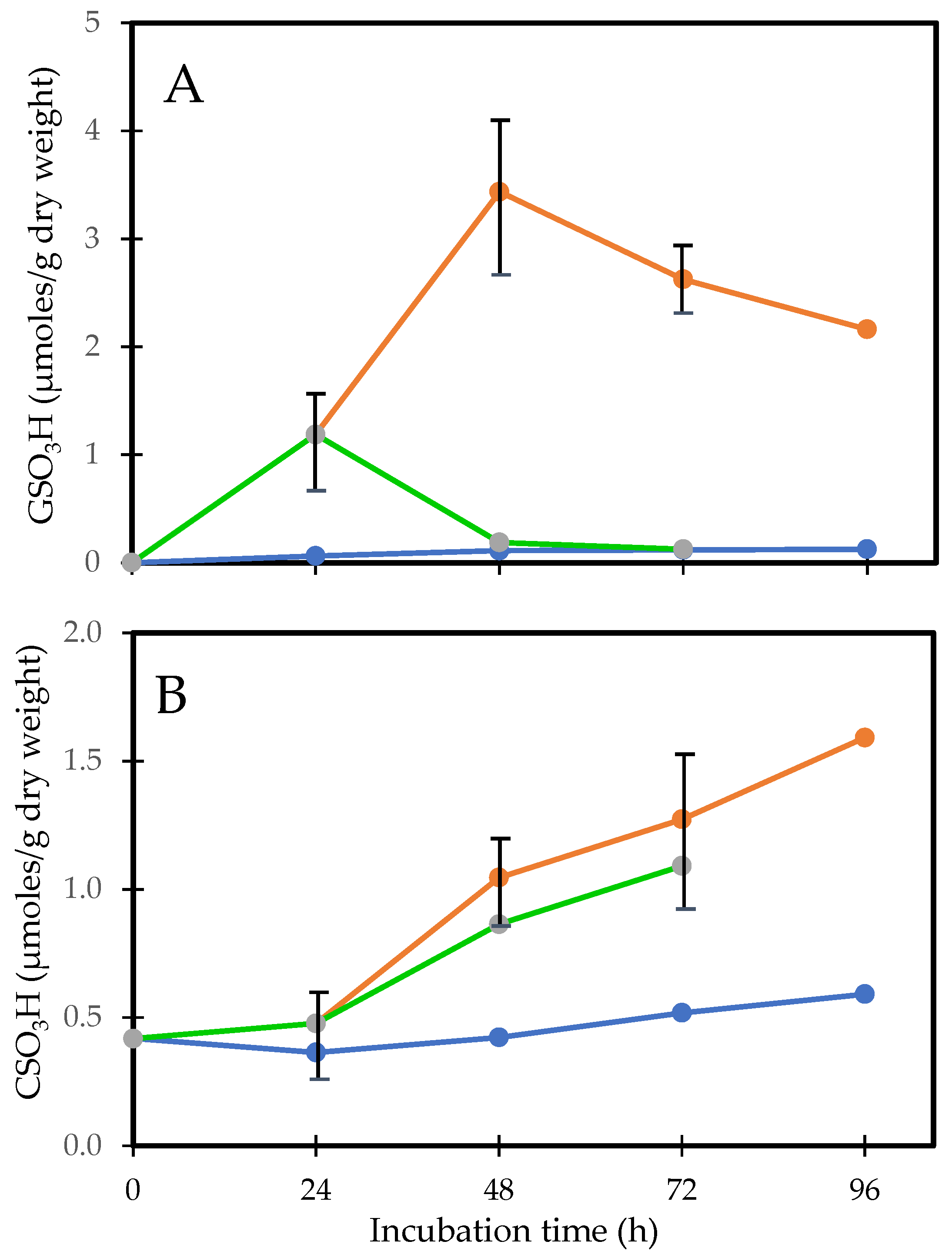
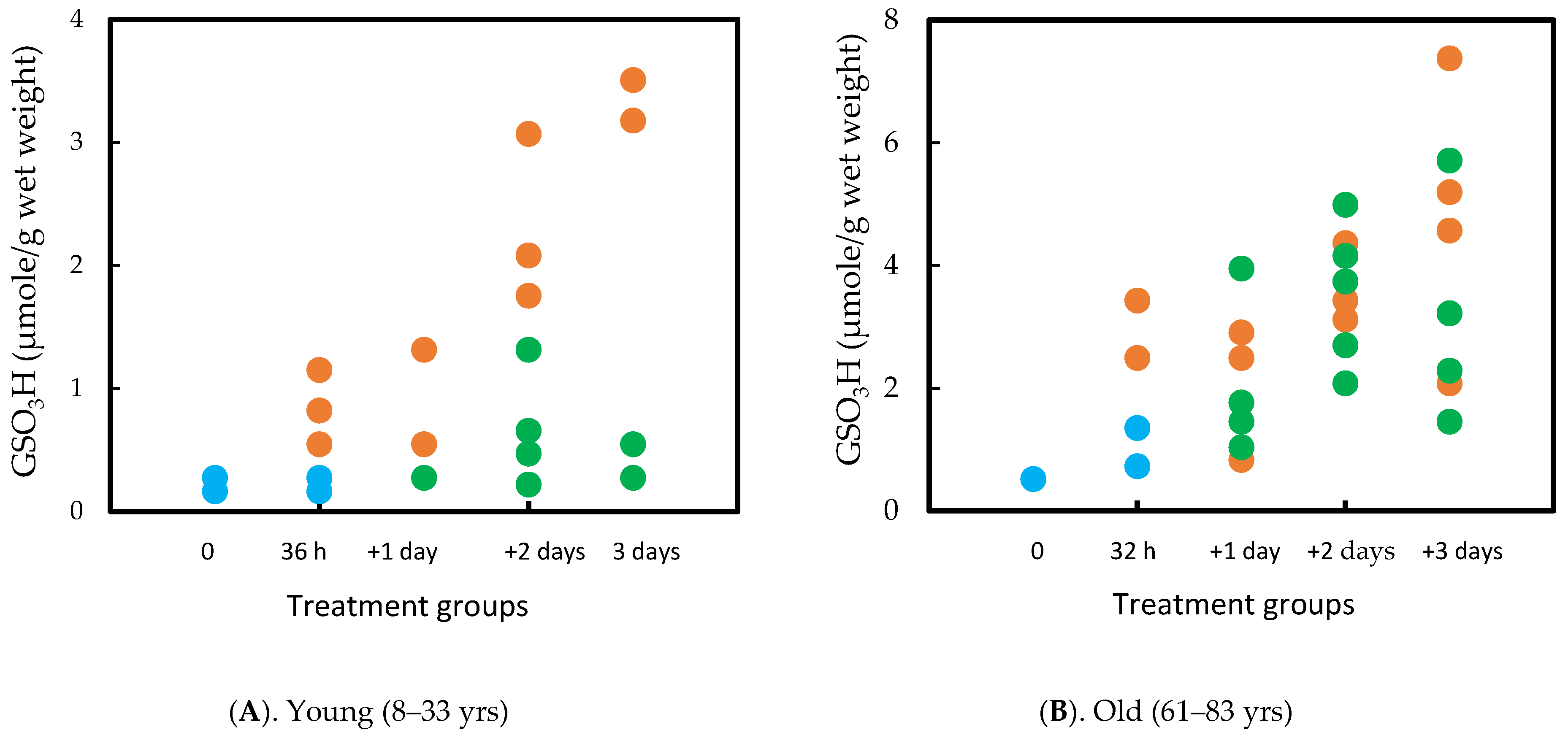
7.2. Human Lens Culture Studies: H2O2 Exposure and Post-Oxidation Recovery
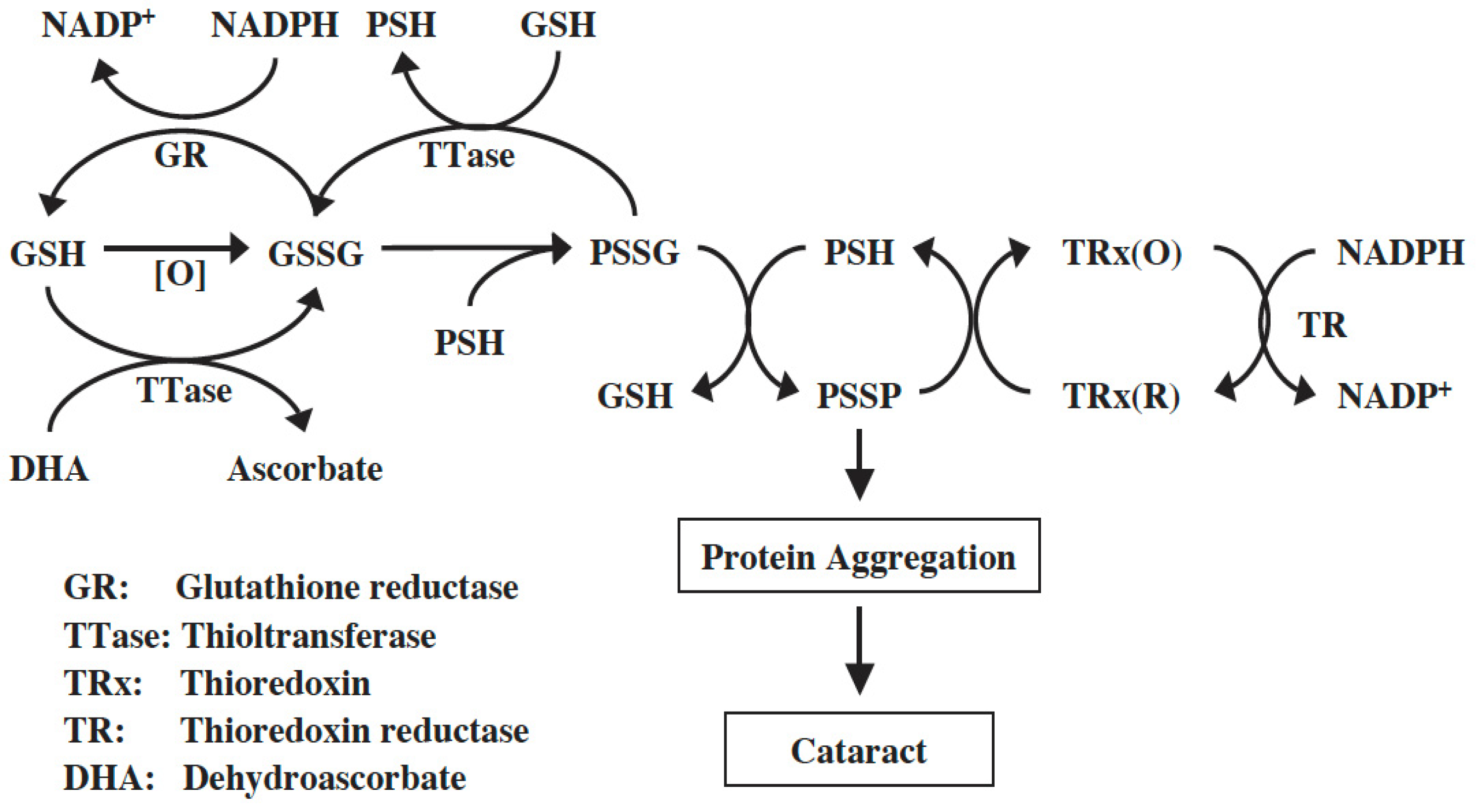
8. The Intrinsic SH/-S-S- Regulating Systems in the Lens
8.1. The GSH-Dependent Thioltransferase
8.2. The NADPH-Dependent Thioredoxin–Thioredoxin Reductase (TRx-TR) System
9. Loss of Thiol Repair Systems in Aging and Cataractous Lenses
9.1. Thiol Repair Systems in Normal Mouse and Human Lens as a Function of Age
9.2. Thiol Repair Systems in Cataractous Lenses
10. Effect of TTase Deletions on Cell Function and Lens Transparency
11. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Augusteyn, R.C. On the growth and internal structure of the human lens. Exp. Eye Res. 2010, 90, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.A.; Augusteyn, R.C. Ontogeny of human lens crystallins. Exp. Eye Res. 1985, 40, 393–410. [Google Scholar] [CrossRef]
- Truscott, R.J. Age-related nuclear cataract: A lens transport problem. Ophthalmic Res. 2000, 32, 185–194. [Google Scholar] [PubMed]
- Delaye, M.; Tardieu, A. Short-range order of crystallin proteins accounts for eye lens transparency. Nature 1983, 302, 415–417. [Google Scholar] [CrossRef]
- Augusteyn, R.C.; Hum, T.P.; Putilin, T.; Thomson, J.A. The location of sulphydryl groups in α-crystallin. Biochim. Biophys. Acta 1987, 915, 132–139. [Google Scholar] [CrossRef]
- Hum, T.P.; Augusteyn, R.C. The state of sulphydryl groups in proteins isolated from normal and cataractous human lenses. Curr. Eye Res. 1987, 6, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Truscott, R.J.; Friedrich, M.G. The etiology of human age-related cataract. Proteins don’t last forever. Biochim. Biophys. Acta 2016, 1860, 192–198. [Google Scholar] [CrossRef]
- Pirie, A. Color and solubility of the proteins of human cataracts. Investig. Ophthalmol. Vis. Sci. 1968, 7, 634–650. [Google Scholar]
- Drinkwater, J.J.; Davis, W.A.; Davis, T.M.E. A systematic review of risk factors for cataract in type 2 diabetes. Diabetes Metab. Res. Rev. 2019, 35, e3073. [Google Scholar] [CrossRef]
- Mrugacz, M.; Pony-Uram, M.; Bryl, A.; Zorena, K. Current Approach to the Pathogenesis of Diabetic Cataracts. Int. J. Mol. Sci. 2023, 24, 6317. [Google Scholar] [CrossRef]
- Fukui, H.N. The effect of hydrogen peroxide on the rubidium transport of the rat lens. Exp. Eye Res. 1976, 23, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Lou, M.F.; Dickerson, J.E., Jr. Protein-thiol mixed disulfides in human lens. Exp. Eye Res. 1992, 55, 889–896. [Google Scholar]
- Truscott, R.J.W.; Augusteyn, R.C. The state of sulphydryl groups in normal and cataractous human lenses. Exp. Eye Res. 1977, 25, 139–148. [Google Scholar] [PubMed]
- Truscott, R.J.W.; Augusteyn, R.C. Oxidative changes in human lens proteins during senile nuclear cataract formation. Biochim. Biophys. Acta 1977, 492, 43–52. [Google Scholar] [CrossRef]
- Truscott, R.J.W.; Augusteyn, R.C. Changes in human lens proteins during nuclear cataract formation. Exp. Eye Res. 1977, 24, 159–170. [Google Scholar]
- Reis, W. Uber die Cysteinreaktion der normalen und pathologisch veranderten Linsen. Arch. Ophthalmol. 1912, 60, 588–598. [Google Scholar] [CrossRef]
- Goldschmidt, M. Beitrag zur Physiologie und Pathologie der Krystallinse. Graefes. Arch. Ophthalmol. 1917, 93, 447–476. [Google Scholar]
- Dische, Z.; Zil, H. Studies on the oxidation of cysteine to cystine in lens proteins during cataract formation. Am. J. Ophthalmol. 1951, 34, 104–113. [Google Scholar] [CrossRef]
- Augusteyn, R.C. Protein modification in cataract: Possible oxidative mechanisms. In Mechanisms of Cataract Formation in the Human Lens; Duncan, G., Ed.; Academic Press: London, UK, 1981; pp. 71–115. [Google Scholar]
- Lou, M.F.; Dickerson, J.E.; Garadi, R. The role of protein-thiol mixed disulfides in cataractogenesis. Exp. Eye Res. 1990, 50, 819–826. [Google Scholar]
- Herman, H.; Moses, S.G. Content and state of glutathione in the tissues of the eye. J. Biol. Chem. 1945, 158, 33–45. [Google Scholar]
- Kinoshita, J.H. Selected Topics in Ophthalmic Biochemistry. Arch. Ophthalmol. 1964, 72, 554–572. [Google Scholar] [PubMed]
- Sinha, D.P.; Sinha, K.P. Observations on glutathione and ascorbic acid content in human cataractous lens. J. Indian Med. Assoc. 1966, 46, 646–649. [Google Scholar]
- Mach, H. Untersuchungen von Linseneiweiss und Mikroelektrophorese von wasserlijslirhem Eiweiss im Altersstar. Klin. Mbl. Augenheilk 1966, 143, 689–710. [Google Scholar]
- Consul, B.; Nagpal, P. Quantitative study of variations in the levels of glutathione and ascorbic acid in human lenses with senile cataract. Ear Nose Throat 1968, 47, 336–2339. [Google Scholar]
- Mostafapour, M.K.; Reddy, V.N. Interactions of glutathione disulfide with lens crystallins. Curr. Eye Res. 1982, 2, 591–596. [Google Scholar]
- Harding, J.J. Free and Protein-Bound Glutathione in Normal and Cataractous Human Lenses. Biochem. J. 1970, 117, 957–960. [Google Scholar]
- Srivastava, S.K.; Beutler, E. Cleavage of lens protein-GSH mixed disulfide by Glutathione reductase. Exp. Eye Res. 1973, 17, 33–42. [Google Scholar]
- Reddy, V.N.; Han, R.F. Protein-bound glutathione in mammalian lenses and in galactose cataract. Doc. Ophthalmol. 1976, 8, 153–160. [Google Scholar]
- Augusteyn, R.C. Growth of the eye lens; I. Weight accumulation in multiple species. Mol. Vis. 2014, 20, 410–426. [Google Scholar]
- Thomson, J.A.; Augusteyn, R.C. On the structure of alpha-crystallin: The minimum molecular weight. Curr. Eye Res. 1988, 7, 563–569. [Google Scholar]
- Lou, M.F.; McKellar, R.; Chyan, O. Quantitation of lens protein mixed disulfides by ion-exchange chromatography. Exp. Eye Res. 1986, 42, 607–616. [Google Scholar] [PubMed]
- Lou, M.F.; Huang, Q.L.; Zigler, S. Effect of opacification and pigmentation on human lens protein thiol/disulfide and solubility. Curr. Eye Res. 1989, 8, 883–890. [Google Scholar] [PubMed]
- Rathbun, W.B.; Murray, D.L. Age-related cysteine uptake as rate-limiting in glutathione synthesis and glutathione half-life in the cultured human lens. Exp. Eye Res. 1991, 53, 205–212. [Google Scholar] [PubMed]
- Li, B.; Kim, J.Y.; Martis, R.M.; Donaldson, P.J.; Lim, J.C. Characterisation of glutathione export from human donor lenses. Transl. Vis. Sci. Technol. 2020, 9, 37. [Google Scholar]
- Kinoshita, J.H.; Masurat, T. Studies on the glutathione in bovine lens. AMA. Arch. Ophthalmol. 1957, 57, 266–274. [Google Scholar]
- Fan, X.; Monnier, V.M.; Whitson, J. Lens glutathione homeostasis: Discrepancies and gaps in knowledge standing in the way of novel therapeutic approaches. Exp. Eye Res. 2017, 156, 103–111. [Google Scholar]
- Truscott, R.J.W. Age-related nuclear cataract—oxidation is the key. Exp. Eye Res. 2005, 80, 709–725. [Google Scholar]
- Lou, M.F.; Dickerson, J.E.; Tung, W.H.; Wolfe, J.K.; Chylack, L.T. Correlation of nuclear color and opalescence with protein S-thiolation in human lenses. Exp. Eye Res. 1999, 68, 547–555. [Google Scholar]
- Giblin, F.J. Glutathione: A vital lens antioxidant. J. Ocul. Pharmacol. Ther. 2000, 16, 121–135. [Google Scholar]
- Dickerson, J.E.; Lou, M.F. Free cysteine levels in normal human lenses. Exp. Eye Res. 1997, 65, 451–454. [Google Scholar]
- Dickerson, J.E.; Lou, M.F. A new mixed disulfide species in human cataractous and aged lenses. Biochim. Biophys. Acta 1993, 1157, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Poole, L.B. The Basics of Thiols and Cysteines in Redox Biology and Chemistry. Free Radic. Biol. Med. 2015, 80, 148–157. [Google Scholar] [CrossRef]
- Lou, M.F. Thiol regulation in the lens. J. Ocul. Pharmacol. Ther. 2000, 16, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Benedek, G.B.; Pande, J.; Thurston, G.M.; Clark, J.I. Theoretical and experimental basis for the inhibition of cataract. Prog. Retin. Eye Res. 1999, 18, 391–402. [Google Scholar]
- Giblin, F.J.; David, L.L.; Wilmarth, P.A.; Leverenz, V.R.; Simpanya, M.F. Shotgun proteomic analysis of S-thiolation sites of guinea pig lens nuclear crystallins following oxidative stress in vivo. Mol. Vis. 2013, 19, 267–280. [Google Scholar] [PubMed]
- Wang, Z.; Schey, K.L. Quantification of thioether-linked glutathione modifications in human lens proteins. Exp. Eye Res. 2018, 175, 83–89. [Google Scholar] [CrossRef]
- Lapko, V.N.; Cerny, R.L.; Smith, D.L.; Smith, J.B. Modifications of human βA1/βA3-crystallins include S-methylation, glutathiolation, and truncation. Protein Sci. 2005, 14, 45–54. [Google Scholar]
- Bechtel, T.J.; Weerapana, E. From structure to redox: The diverse functional roles of disulfides and implications in disease. Proteomics 2017, 17, 1600391. [Google Scholar] [CrossRef]
- Cui, X.-L.; Lou, M.F. The effect and recovery of long-term H2O2 exposure on lens morphology and biochemistry. Exp. Eye Res. 1993, 57, 157–167. [Google Scholar] [CrossRef]
- Latta, K.; Augusteyn, R.C. The purification and properties of human lens glutathione reductase. Exp. Eye Res. 1984, 39, 343–354. [Google Scholar] [CrossRef]
- Augusteyn, R.C. On the possible role of glutathione in maintaining human lens protein sulphydryls. Exp. Eye Res. 1979, 28, 665–671. [Google Scholar] [PubMed]
- Fecondo, J.V.; Augusteyn, R.C. Superoxide dismutase, catalase and glutathione peroxidase in the normal and cataractous human lens. Exp. Eye Res. 1983, 36, 15–24. [Google Scholar] [PubMed]
- Zhang, W.; Libondi, T.; Lou, M.F. Analysis of the free and protein-bound cysteine and glutathione in LOCS III classified human cataractous lenses. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2381. [Google Scholar]
- Liang, J.N.; Pelletier, M.R. Destabilization of lens protein conformation by glutathione mixed disulfide. Exp. Eye Res. 1988, 47, 17–25. [Google Scholar]
- Kono, M.; Sen, C.; Chakrabarti, B. Changes in the conformation stability of gamma-crystallins upon glutathione reaction. Biochemistry 1990, 29, 464–470. [Google Scholar]
- Harding, J.J. Conformational changes in human lens proteins in cataract. Biochem. J. 1972, 129, 97–100. [Google Scholar]
- Kurzel, B.S.; Crock, R.B.; Staton, G.W.; Wolbarscht, M.; Yamanishi, B.S.; Borkman, R.F. Tryptophan excited states and cataracts in the human lens. Nature 1973, 241, 132–133. [Google Scholar]
- McNamara, M.K.; Augusteyn, R.C. Conformational changes in soluble lens proteins during the development of senile nuclear cataract. Curr. Eye Res. 1984, 3, 571–583. [Google Scholar]
- McNamara, M.K.; Augusteyn, R.C. The effects of hydrogen peroxide on lens proteins: A possible model for nuclear cataract. Exp. Eye Res. 1984, 38, 45–56. [Google Scholar]
- Zigman, S.; Poxhia, T.; McDaniel, T.; Lou, M.F.; Yu, N.-T. Effect of chronic near-ultraviolet radiation on the gray squirrel lens in vivo. Investig. Ophthalmol. Vis. Sci. 1991, 32, 1723–1732. [Google Scholar]
- Giblin, F.J.; Leverenz, V.R.; Padgaonkar, V.A.; Unakar, N.J.; Dang, L.; Lin, L.-R.; Lou, M.F.; Reddy, V.N.; Borchman, D.; Dillon, J.P. UVA light in vivo reaches the nucleus of the guinea pig lens and produces deleterious oxidative effects. Exp. Eye Res. 2002, 75, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Giblin, F.; Padgaonkar, V.A.; Leverenz, V.R.; Lin, L.-R.; Lou, M.F.; Unakar, N.J.; Dang, L.; Dickerson, J.E., Jr.; Reddy, V.E. Nuclear light scattering, disulfide formation and membrane damage in lenses of older guinea pigs treated with hyperbaric oxygen. Exp. Eye Res. 1995, 60, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Giblin, F.; McCready, J.; Schrimscher, L.; Reddy, V. The role of glutathione metabolism in the detoxification of H2O2 in rabbit lens. Investig. Ophthalmol. Vis. Sci. 1987, 22, 330–335. [Google Scholar]
- Zhang, W.Z.; Augusteyn, R.C. Ageing of glutathione reductase in the lens. Exp. Eye Res. 1994, 59, 91–96. [Google Scholar] [CrossRef]
- Spector, A.; Garner, W.H. Hydrogen peroxide and human cataract. Exp. Eye Res. 1981, 33, 673–681. [Google Scholar] [CrossRef]
- Spector, A. Oxidative stress-induced cataract: Mechanism of action. FASEB J. 1995, 9, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Hanson, S.R.A.; Chen, A.A.; Smith, J.B.; Lou, M.F. Thiolation of the γB-crystallins in intact bovine lens exposed to hydrogen peroxide. J. Biol. Chem. 1999, 274, 4735–4742. [Google Scholar] [CrossRef]
- Smith, J.B.; Jiang, X.; Abraham, E.C. Identification of hydrogen peroxide oxidation sites of αA- and αB-crystallins. Free Radic. Res. 1997, 26, 103–111. [Google Scholar] [CrossRef]
- Taylor, H.R. Ultraviolet radiation and the eye: An epidemiologic study. Trans. Am. Ophthalmol. Soc. 1989, 87, 802–853. [Google Scholar]
- Brigelius, R. Mixed disulphides: Biological functions and increase in oxidative stress. In Oxidative Stress; Sies, H., Ed.; Academic Press: New York, NY, USA, 1985; pp. 43–72. [Google Scholar]
- Lou, M.F. Redox regulation in the lens. Prog. Retin. Eye Res. 2003, 22, 657–682. [Google Scholar] [CrossRef]
- Blundell, T.; Lindley, P.; Miller, L.; Moss, D.; Slingsby, C.; Tickle, I.; Turnell, B.; Wistow, G. The molecular structure and stability of the eye lens: X-ray analysis of gamma-crystallin II. Nature 1981, 289, 771–777. [Google Scholar]
- Lou, M.F.; Xu, G.-T.; Cui, X.-L. Further studies on the dynamic changes of glutathione and protein-thiol mixed disulfides in H2O2, induced cataract in rat lenses: Distributions and effect of aging. Curr. Eye Res. 1995, 14, 951–958. [Google Scholar] [PubMed]
- Lou, M.F. Protein-thiol mixed disulfides and thioltransferase in the lens—A review. In Advances in Ocular Toxicology; Green, K., Edelhauser, H.F., Hackett, R.B., Hull, D.S., Potter, D.E., Tripathi, R.C., Eds.; Plenum: New York, NY, USA, 1997; pp. 27–46. [Google Scholar]
- Lou, M.F.; Xu, G.-T. Recovery of oxidative damage in human lenses. Investig. Ophthalmol. Vis. Sci. 1994, 35, 1569. [Google Scholar]
- Raghavachari, N.; Lou, M.F. Evidences for the presence of thioltransferase in the lens. Exp. Eye Res. 1996, 63, 433–441. [Google Scholar]
- Gan, Z.R.; Wells, W. The primary structure of pig liver thioltransferase. J. Biol. Chem. 1987, 262, 6699–6703. [Google Scholar]
- Mieyal, J.J.; Starke, D.W.; Gravina, S.A.; Dothey, C.; Chung, J.W. Thioltransferase in human red blood cells: Purification and properties. Biochemistry 1991, 30, 6088–6097. [Google Scholar]
- Gladyshav, V.; Sun, C.A.; Liu, A.-M.; Lou, M.F. Identification and characterization of a new mammalian mitochondrial glutaredoxin (thioltransferase). J. Biol. Chem. 2001, 276, 30374–30380. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin and glutaredoxin systems. J. Biol. Chem. 1989, 264, 13963–13966. [Google Scholar]
- Hirota, K.; Matsui, M.; Iwata, S.; Nishiyama, A.; Mori, K.; Yodoi, J. Ap-1 transcriptional activity is regulated by a direct association between thioredoxin and Ref-1. Proc. Natl. Acad. Sci. USA 1997, 94, 3633–3638. [Google Scholar]
- Powis, G.; Montfort, W.R. Properties and biological activities of thioredoxins. Ann. Rev. Pharmacol. Toxicol. 2001, 41, 261–295. [Google Scholar]
- Matsui, M.; Oshima, M.; Oshima, H.; Takaku, K.; Maruyama, T.; Yodoi, J. Early mmbryonic lethality caused by targeted disruption of the mouse thioredoxin gene. Dev. Biol. 1996, 178, 179–185. [Google Scholar] [PubMed]
- Reddy, P.G.; Bhuyan, D.K.; Bhuyan, K.C. Thioredoxin gene expression and its protein distribution in the human lens with age. Investig. Ophthalmol. Vis Sci. 1999, 40, S170. [Google Scholar]
- Reddy, P.G.; Bhuyan, D.K.; Bhuyan, K.C. Lens-specific regulation of the thioredoxin-1 gene, but not thioredoxin-2 upon in vivo photochemical oxidative stress in the Emory mouse. Biochem. Biophys. Res. Commun. 1999, 265, 345–349. [Google Scholar] [PubMed]
- Yegorova, S.; Liu, A.-M.; Lou, M.F. Cloning, expression and characterization of human lens thioredoxin. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3263–3271. [Google Scholar]
- Xing, K.-Y.; Lou, M.F. Effect of age on the thioltransferase (glutaredoxin and thioredoxin systems in the human lens. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6598–6604. [Google Scholar]
- Zhang, J.; Yan, H.; Lofgren, S.; Tian, X.; Lou, M.F. Ultraviolet radiation-induced cataract in mice: The effect of age and the potential biochemical mechanism. Investig. Ophthalmol. Vis Sci. 2012, 53, 7276–7285. [Google Scholar]
- Xing, K.Y.; Lou, M.F. Effect of H2O2 on human lens epithelial cells and the possible mechanism for oxidative damage repair by thioltransferase. Exp. Eye Res. 2002, 74, 113–122. [Google Scholar]
- Moon, S.; Fernando, M.R.; Lou, M.F. Induction of thioltransferase and thioredoxin/thioredoxin reductase systems in cultured porcine lenses under oxidative stress. Investig. Ophthalmol. Vis Sci. 2005, 46, 3783–3789. [Google Scholar]
- Wei, M.; Xing, K.-Y.; Fan, Y.-C.; Lebondi, T.; Lou, M.F. Loss of the thiol repair systems in the human cataractous lenses. Investig. Ophthalmol. Vis. Sci. 2015, 56, 598–605. [Google Scholar]
- Spector, A.; Roy, D. Disulfide-linked high molecular weight protein associated with human cataract. Proc. Natl. Acad. Sci. USA 1978, 75, 3244–3248. [Google Scholar]
- Grimaud, R.; Ezatry, B.; Mitchell, J.K. Repair of oxidized proteins: Identification of a new methionine sulfoxide reductase. J. Biol. Chem. 2001, 276, 48915–48920. [Google Scholar] [CrossRef] [PubMed]
- Boschi-Muller, S.; Branlant, G. Methionine sulfoxide reductase: Chemistry, substrate binding, recycling process and oxidase activity. Bioorg. Chem. 2014, 57, 222–230. [Google Scholar] [CrossRef]
- Spector, A.; Scotto, R.; Weissbach, H.; Brot, N. Lens methionine sulfoxide reductase. Biochim. Biophys. Res. Commun. 1982, 108, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Kantarow, M.; Hawse, J.R.; Cowell, T.L.; Pizarro, G.O.; Reddy, V.N.; Hejtmancik, H. Methionine sulfoxide reductase A is important for lens cell viability and resistance to oxidative stress. Proc. Natl. Acad. Sci. USA 2004, 101, 9654–9659. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, M.A.; Pizarro, G.O.; Sagher, D.; DeAmicis, C.; Brot, N.; Hejtmancik, H.; Kantarow, M. Methionine sulphoxide reductases B1, B2, and B3 are present in the human lens and confer oxidative stress resistance to lens cells. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2107–2112. [Google Scholar] [CrossRef]
- Lou, M.F.; Xing, K.; Fernando, M.R.; Moon, Y.; Ho, Y.-S. Thioltransferase knockout mouse: A new cataract model. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1026. [Google Scholar]
- Zhang, J.; Xing, K.; Ho, Y.-S.; Lou, M.F. Age-dependent biochemical changes in the lenses of thioltransferase (TTase, or Grx1) knockout mice. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1548. [Google Scholar]
- Chylack, L.T., Jr.; Leske, M.C.; McCarthy, D.; Khu, P.; Kashiwagi, T.; Sperduto, R. Lens opacities classification system II (LOCS II). Arch. Ophthalmol. 1989, 107, 991–997. [Google Scholar] [CrossRef]
- Meyer, L.M.; Löfgren, S.; Ho, Y.; Lou, M.; Wegener, A.; Holz, F.; Söderberg, P.G. Absence of Glutaredoxin1 (Grx1) Increases Lens Susceptibility to Oxidative Stress Induced by UVR-B. Exp. Eye Res. 2009, 89, 833–839. [Google Scholar] [CrossRef]
- Kronschläger, M.; Galichanin, K.; Ekström, J.; Lou, M.F.; Söderberg, P.G. Protective effect of the thioltransferase gene on in vivo UVR-300 nm-induced cataract. Investig. Ophthalmol. Vis. Sci. 2012, 53, 248–252. [Google Scholar] [CrossRef]
- Li, C.; Chen, X.; Zhang, S.; Liang, C.; Ma, X.; Zhang, R.; Yan, H. Glutaredoxin 1 protects lens epithelial cells from epithelial-mesenchymal transition by preventing casein kinase 1α S-glutathionylation during posterior capsular opacification. Redox Biol. 2023, 62, 102676. [Google Scholar] [CrossRef]
- Li, C.; Yan, W.; Yan, H. Oxidative Stress, Glutaredoxins, and Their Therapeutic Potential in Posterior Capsular Opacification. Antioxidants 2024, 13, 2010. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lou, M.F.; Augusteyn, R.C. Oxidation-Induced Mixed Disulfide and Cataract Formation: A Review. Antioxidants 2025, 14, 425. https://doi.org/10.3390/antiox14040425
Lou MF, Augusteyn RC. Oxidation-Induced Mixed Disulfide and Cataract Formation: A Review. Antioxidants. 2025; 14(4):425. https://doi.org/10.3390/antiox14040425
Chicago/Turabian StyleLou, Marjorie F., and Robert C. Augusteyn. 2025. "Oxidation-Induced Mixed Disulfide and Cataract Formation: A Review" Antioxidants 14, no. 4: 425. https://doi.org/10.3390/antiox14040425
APA StyleLou, M. F., & Augusteyn, R. C. (2025). Oxidation-Induced Mixed Disulfide and Cataract Formation: A Review. Antioxidants, 14(4), 425. https://doi.org/10.3390/antiox14040425