
Targeted Cancer Therapy with Gold–Iron Oxide Nanourchins: Inducing Oxidative Stress, Paraptosis, and Sensitizing Tumor Cells to Cisplatin


 , , ,
, , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Nanourchins Synthesis
2.3. MTT Assay
2.4. Oil Red O Staining
2.5. Cell Rox Staining for Oxidative Stress Detection
2.6. Bodipy Staining for Lipidic Peroxidation Evaluation
2.7. JC-1 Assay for Mitochondrial Membrane Potential Analysis
2.8. Seahorse Metabolic Flux Analyses
2.9. Western Blotting
2.10. Autophagy Assessment Through CYTO-ID ENZO Detection Kit
2.11. Cell Cycle Analysis
2.12. Invasion Assay Through Geltrex
2.13. In Vitro Tube Formation Assay
2.14. Evaluation of Apoptosis Through Cytofluorimetric Annexin V/PI Double Staining
2.15. Statistics
3. Results
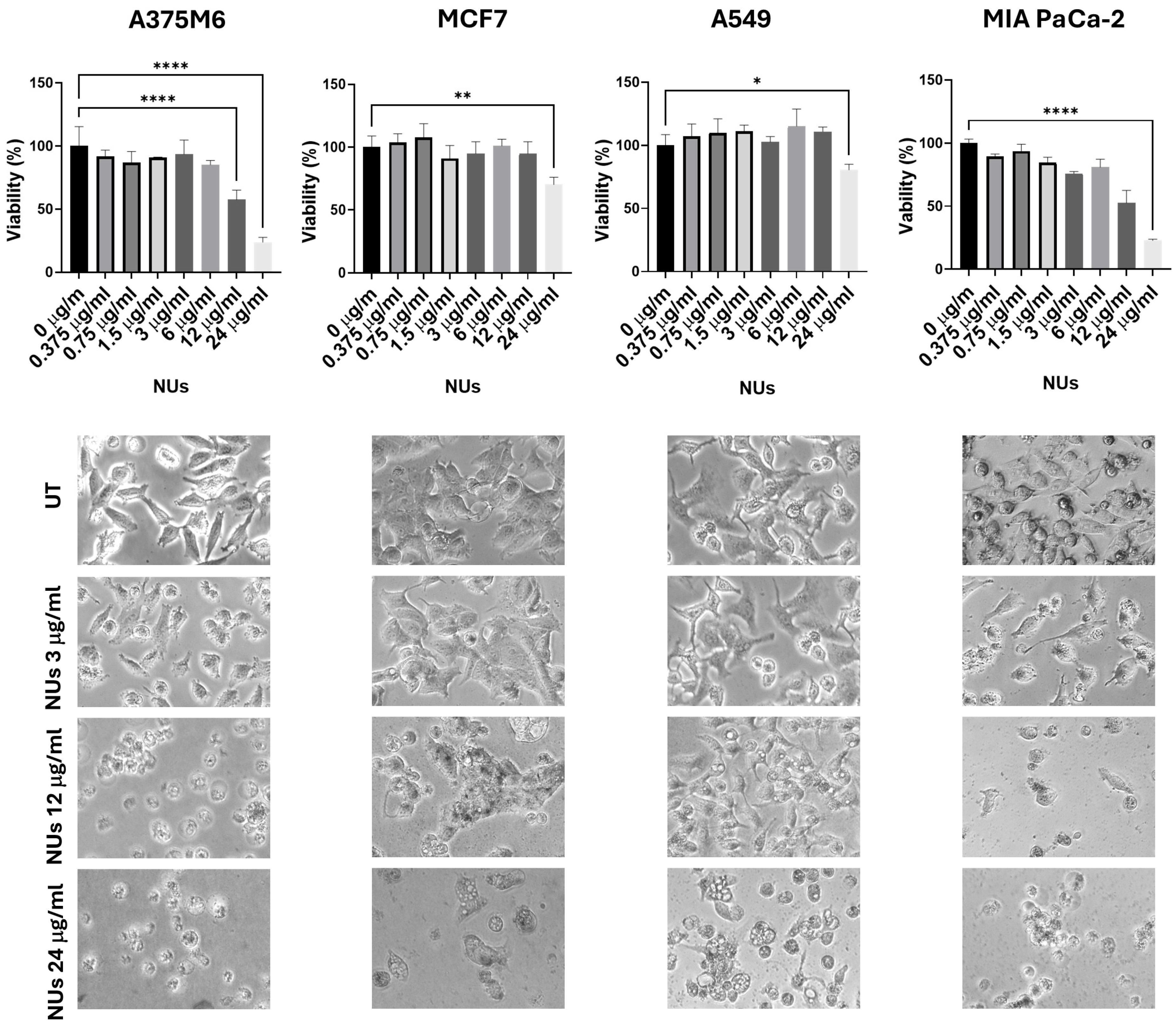
3.1. NUs Efficacy on Different Tumor Cell Types
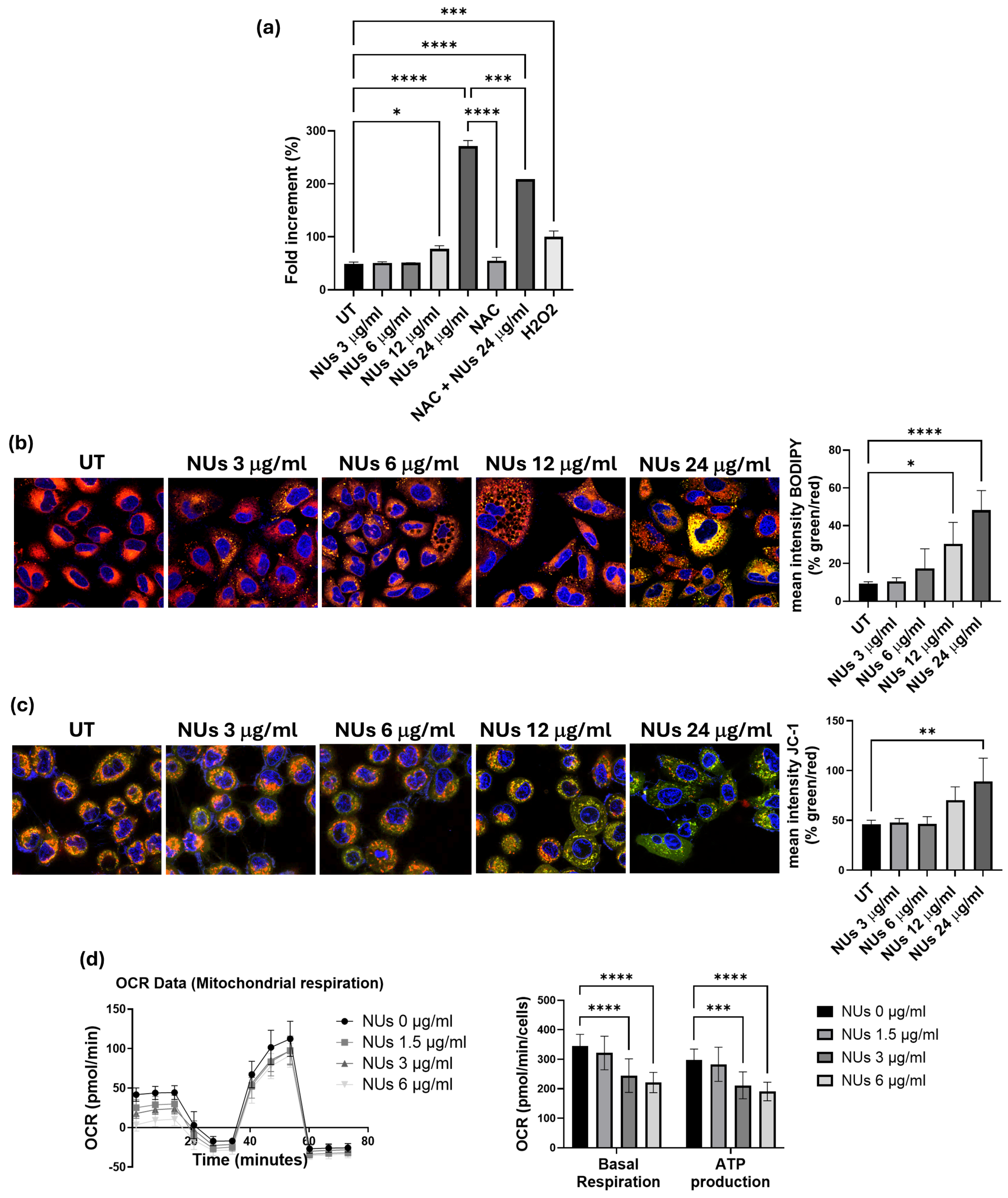
3.2. Pro-Oxidant Activity of NUs on A549 Cells
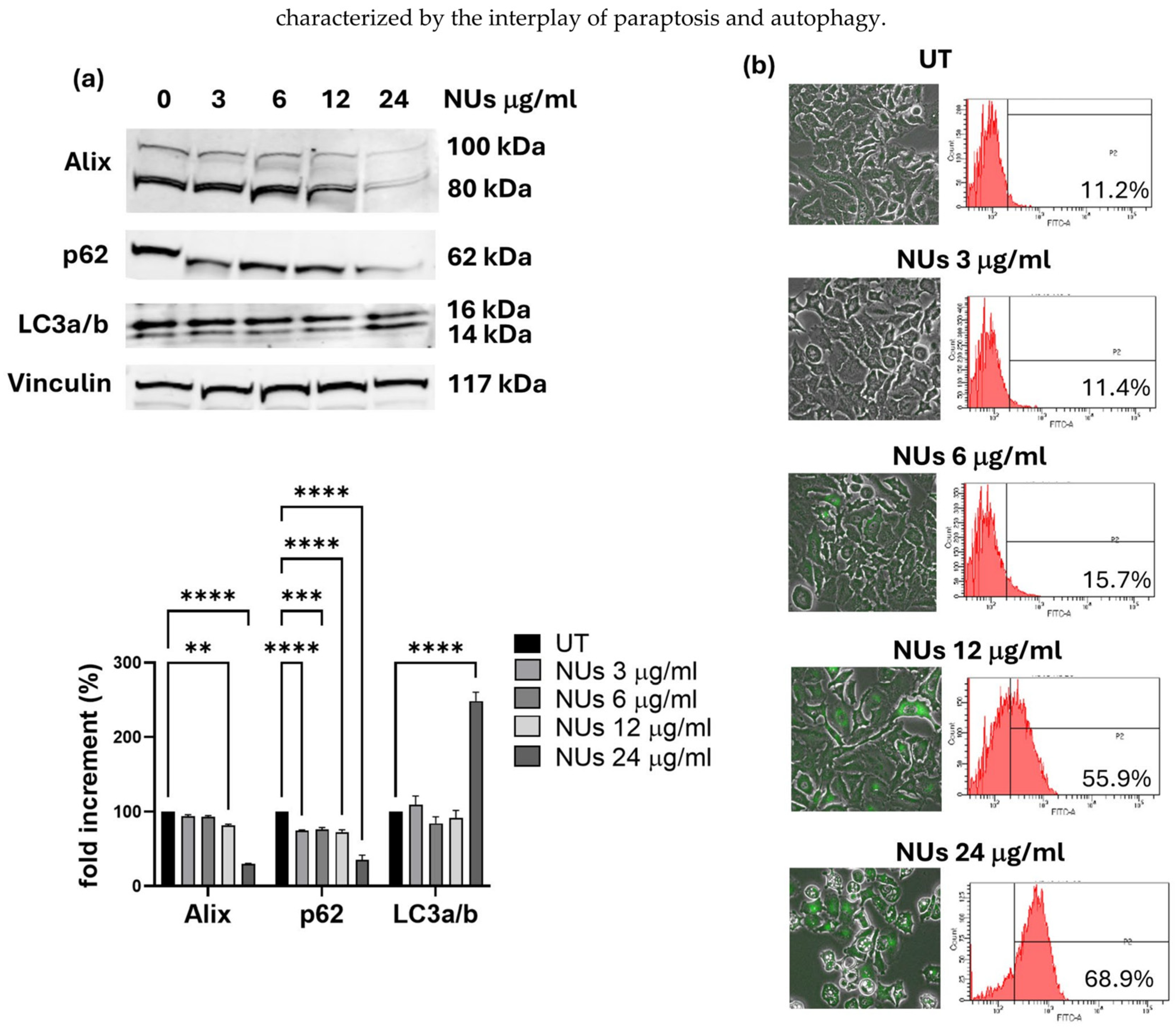
3.3. Autophagy-Related Cell Death Induced by Nus in a459 Cells
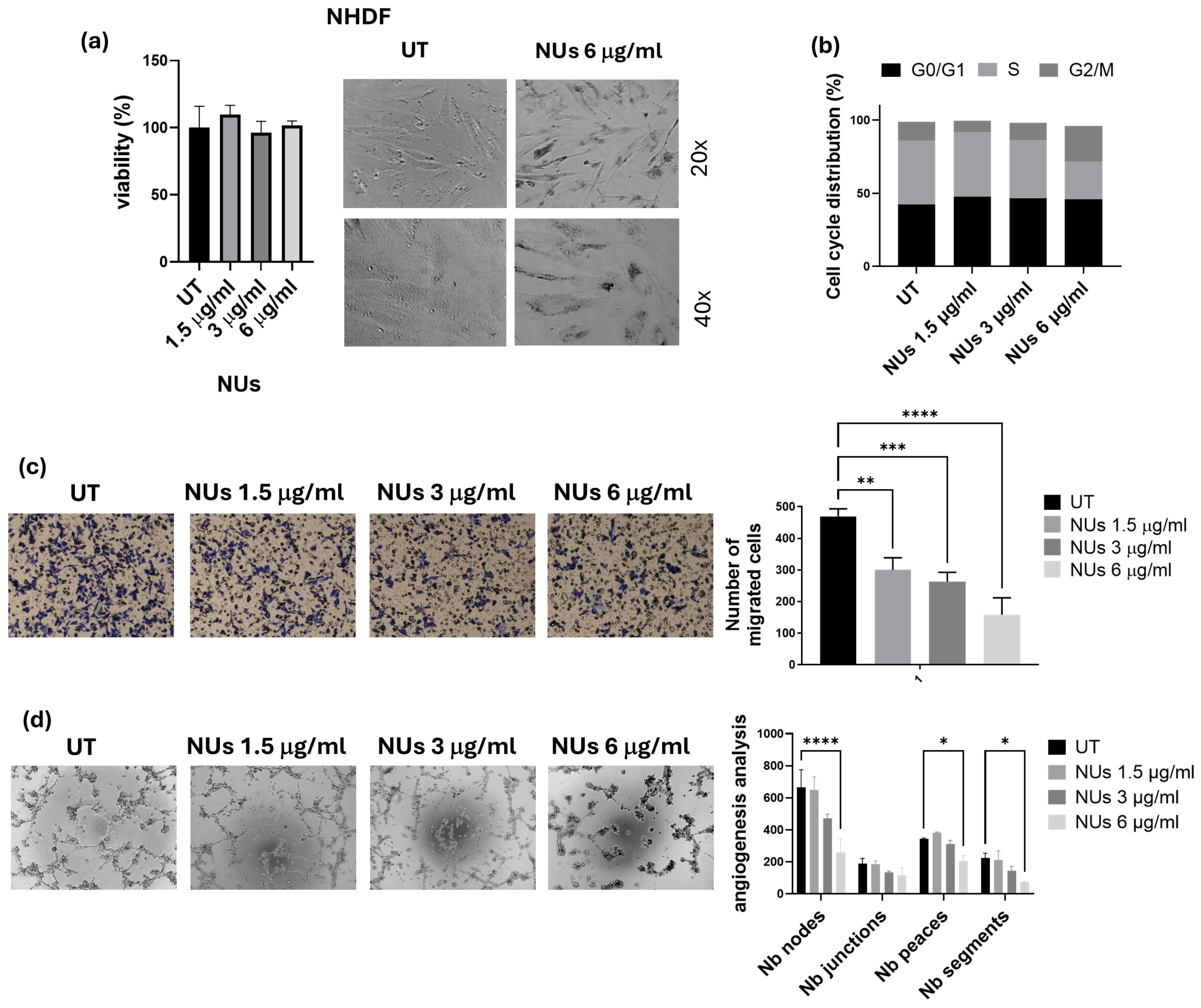
3.4. NUs Biological Effects on A549 Cells
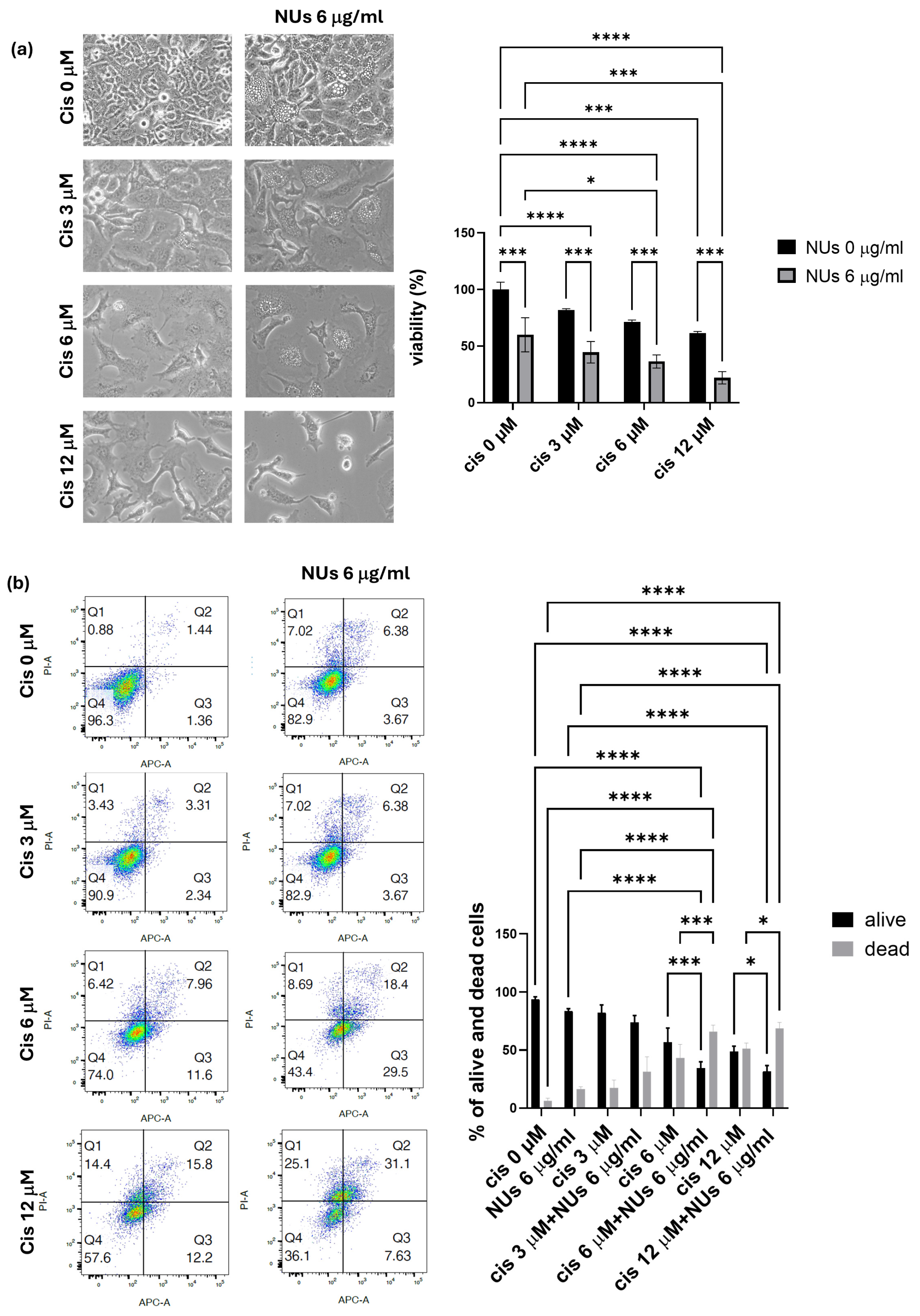
3.5. NUs Potentiation of Cisplatin Toxicity on A459 Cells
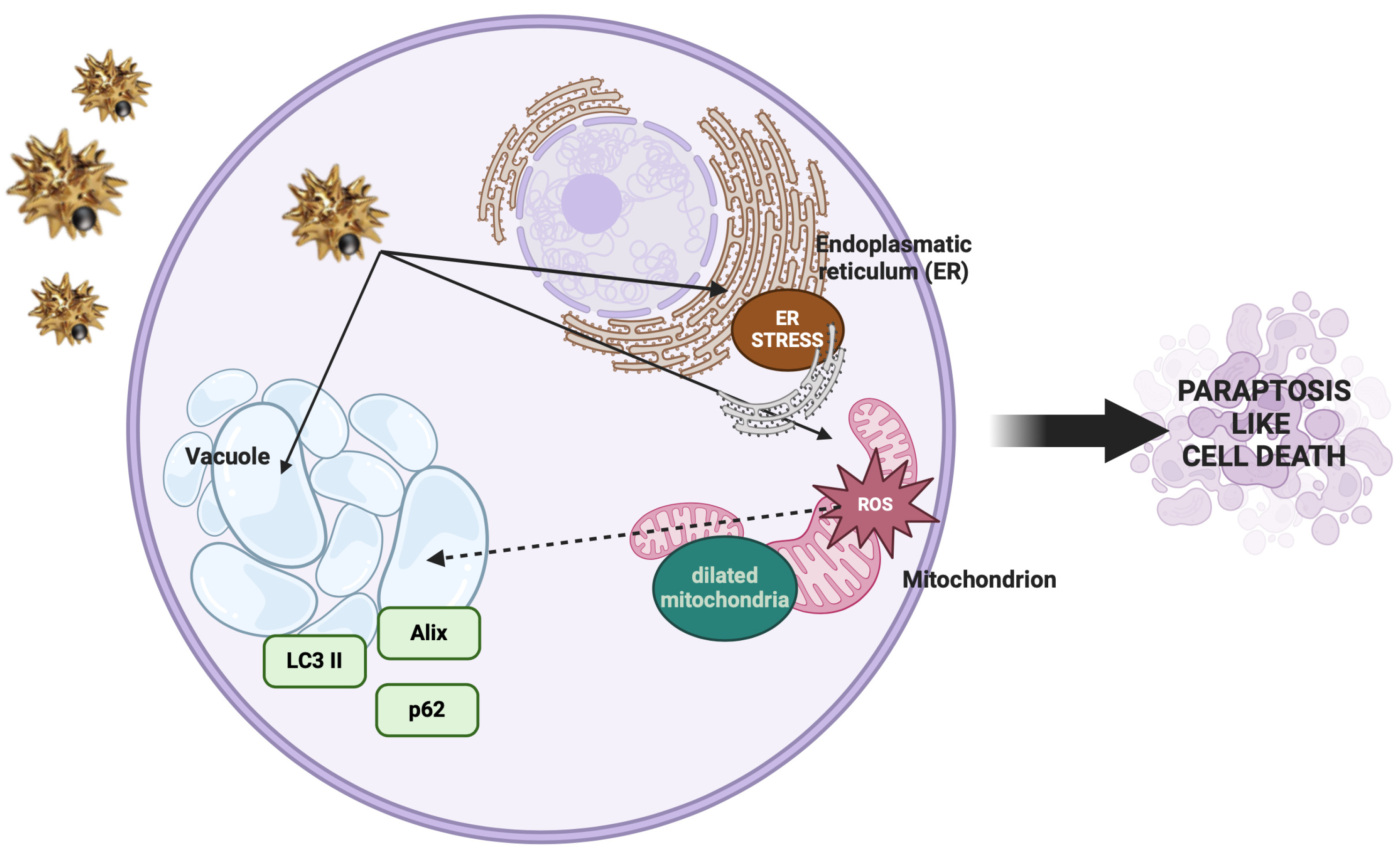
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| NUs | Nanourchins |
| NSCLC | Non-small-cell lung cancer |
| SCLC | Small-cell lung cancer |
| NPs | Nanoparticles |
| MMP | Mitochondrial membrane potential |
| ROS | Reactive oxygen species |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Hausman, D.M. What Is Cancer? Perspect. Biol. Med. 2019, 62, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Gridelli, C.; Rossi, A.; Carbone, D.P.; Guarize, J.; Karachaliou, N.; Mok, T.; Petrella, F.; Spaggiari, L.; Rosell, R. Non-Small-Cell Lung Cancer. Nat. Rev. Dis. Primers 2015, 1, 15009. [Google Scholar] [CrossRef]
- Smith, W.; Khuri, F.R. The Care of the Lung Cancer Patient in the 21st Century: A New Age. Semin. Oncol. 2004, 31, 11–15. [Google Scholar] [CrossRef]
- Maslyar, D.; Jahan, T.M.; Jablons, D.M. Mechanisms of and Potential Treatment Strategies for Metastatic Disease in Non-Small Cell Lung Cancer. Semin. Thorac. Cardiovasc. Surg. 2004, 16, 40–50. [Google Scholar]
- Han, M.; Zhao, Y.; Tan, C.; Xiong, Y.; Wang, W.; Wu, F.; Fei, Y.; Wang, L.; Liang, Z. Cathepsin L Upregulation-Induced EMT Phenotype Is Associated with the Acquisition of Cisplatin or Paclitaxel Resistance in A549 Cells. Acta Pharmacol. Sin. 2016, 37, 1606–1622. [Google Scholar] [CrossRef]
- Wu, H.; Zhu, H.; Li, X.; Liu, Z.; Zheng, W.; Chen, T.; Yu, B.; Wong, K.-H. Induction of Apoptosis and Cell Cycle Arrest in A549 Human Lung Adenocarcinoma Cells by Surface-Capping Selenium Nanoparticles: An Effect Enhanced by Polysaccharide–Protein Complexes from Polyporus Rhinocerus. J. Agric. Food Chem. 2013, 61, 9859–9866. [Google Scholar] [CrossRef]
- Pei, Z.; Chen, S.; Ding, L.; Liu, J.; Cui, X.; Li, F.; Qiu, F. Current Perspectives and Trend of Nanomedicine in Cancer: A Review and Bibliometric Analysis. J. Control. Release 2022, 352, 211–241. [Google Scholar] [CrossRef]
- Tiwari, J.N.; Tiwari, R.N.; Kim, K.S. Zero-Dimensional, One-Dimensional, Two-Dimensional and Three-Dimensional Nanostructured Materials for Advanced Electrochemical Energy Devices. Prog. Mater. Sci. 2012, 57, 724–803. [Google Scholar] [CrossRef]
- Pelaz, B.; Del Pino, P.; Maffre, P.; Hartmann, R.; Gallego, M.; Rivera-Fernández, S.; De La Fuente, J.M.; Nienhaus, G.U.; Parak, W.J. Surface Functionalization of Nanoparticles with Polyethylene Glycol: Effects on Protein Adsorption and Cellular Uptake. ACS Nano 2015, 9, 6996–7008. [Google Scholar] [CrossRef]
- Yaqoob, S.B.; Adnan, R.; Rameez Khan, R.M.; Rashid, M. Gold, Silver, and Palladium Nanoparticles: A Chemical Tool for Biomedical Applications. Front. Chem. 2020, 8, 376. [Google Scholar] [CrossRef] [PubMed]
- Burlec, A.F.; Corciova, A.; Boev, M.; Batir-Marin, D.; Mircea, C.; Cioanca, O.; Danila, G.; Danila, M.; Bucur, A.F.; Hancianu, M. Current Overview of Metal Nanoparticles’ Synthesis, Characterization, and Biomedical Applications, with a Focus on Silver and Gold Nanoparticles. Pharmaceuticals 2023, 16, 1410. [Google Scholar] [CrossRef] [PubMed]
- Kodiha, M.; Wang, Y.M.; Hutter, E.; Maysinger, D.; Stochaj, U. Off to the Organelles—Killing Cancer Cells with Targeted Gold Nanoparticles. Theranostics 2015, 5, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.; Shiau, A.; Chen, Y.; Chang, C.; Chen, H.H.; Wu, C. Increased Apoptotic Potential and Dose-Enhancing Effect of Gold Nanoparticles in Combination with Single-Dose Clinical Electron Beams on Tumor-Bearing Mice. Cancer Sci. 2008, 99, 1479–1484. [Google Scholar] [CrossRef]
- Khan, J.A.; Pillai, B.; Das, T.K.; Singh, Y.; Maiti, S. Molecular Effects of Uptake of Gold Nanoparticles in HeLa Cells. ChemBioChem 2007, 8, 1237–1240. [Google Scholar] [CrossRef]
- Cho, W.-S.; Cho, M.; Jeong, J.; Choi, M.; Cho, H.-Y.; Han, B.S.; Kim, S.H.; Kim, H.O.; Lim, Y.T.; Chung, B.H.; et al. Acute Toxicity and Pharmacokinetics of 13 Nm-Sized PEG-Coated Gold Nanoparticles. Toxicol. Appl. Pharmacol. 2009, 236, 16–24. [Google Scholar] [CrossRef]
- Sun, L.; Liu, D.; Wang, Z. Functional Gold Nanoparticle−Peptide Complexes as Cell-Targeting Agents. Langmuir 2008, 24, 10293–10297. [Google Scholar] [CrossRef]
- Samhadaneh, D.M.; Alqarni, K.A.; Smart, A.; Kuang, M.; Moujaber, O.; Maysinger, D.; Stochaj, U. Gold Nanourchins Induce Cellular Stress, Impair Proteostasis and Damage RNA. Nanomed. Nanotechnol. Biol. Med. 2019, 22, 102083. [Google Scholar] [CrossRef]
- Candreva, A.; Morrone, E.; La Deda, M. Gold Sea Urchin-Shaped Nanoparticles: Synthesis and Characterization of Energy Transducer Candidates. Plasmonics 2023, 18, 291–298. [Google Scholar] [CrossRef]
- Joshi, A.; Agrawal, A.; Bhattacharya, S. Formulation and Clinical Advancement of Nanourchins: A Novel Multibranched Nanoparticulate Drug-Delivery System. Nanomedicine 2022, 17, 1477–1499. [Google Scholar] [CrossRef]
- Ruzzolini, J.; Peppicelli, S.; Andreucci, E.; Bianchini, F.; Margheri, F.; Laurenzana, A.; Fibbi, G.; Pimpinelli, N.; Calorini, L. Everolimus Selectively Targets Vemurafenib Resistant BRAFV600E Melanoma Cells Adapted to Low pH. Cancer Lett. 2017, 408, 43–54. [Google Scholar] [CrossRef]
- Biagioni, A.; Chillà, A.; Del Rosso, M.; Fibbi, G.; Scavone, F.; Andreucci, E.; Peppicelli, S.; Bianchini, F.; Calorini, L.; Li Santi, A.; et al. CRISPR/Cas9 uPAR Gene Knockout Results in Tumor Growth Inhibition, EGFR Downregulation and Induction of Stemness Markers in Melanoma and Colon Carcinoma Cell Lines. Front. Oncol. 2021, 11, 663225. [Google Scholar] [CrossRef]
- Peri, S.; Ruzzolini, J.; Urciuoli, S.; Versienti, G.; Biagioni, A.; Andreucci, E.; Peppicelli, S.; Bianchini, F.; Bottari, A.; Calorini, L.; et al. An Oleocanthal-Enriched EVO Oil Extract Induces the ROS Production in Gastric Cancer Cells and Potentiates the Effect of Chemotherapy. Antioxidants 2022, 11, 1762. [Google Scholar] [CrossRef] [PubMed]
- Ruzzolini, J.; Peppicelli, S.; Bianchini, F.; Andreucci, E.; Urciuoli, S.; Romani, A.; Tortora, K.; Caderni, G.; Nediani, C.; Calorini, L. Cancer Glycolytic Dependence as a New Target of Olive Leaf Extract. Cancers 2020, 12, 317. [Google Scholar] [CrossRef]
- Peppicelli, S.; Ruzzolini, J.; Bianchini, F.; Andreucci, E.; Nediani, C.; Laurenzana, A.; Margheri, F.; Fibbi, G.; Calorini, L. Anoikis Resistance as a Further Trait of Acidic-Adapted Melanoma Cells. J. Oncol. 2019, 2019, 8340926. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, G.; Berndt, S.; Ferratge, S.; Rasband, W.; Cuendet, M.; Uzan, G.; Albanese, P. Angiogenesis Analyzer for ImageJ—A Comparative Morphometric Analysis of “Endothelial Tube Formation Assay” and “Fibrin Bead Assay”. Sci. Rep. 2020, 10, 11568. [Google Scholar] [CrossRef]
- Kim, D.-Y.; Moon, S.-H.; Han, J.-H.; Kim, M.-J.; Oh, S.-J.; Bharti, D.; Lee, S.-H.; Park, J.-K.; Rho, G.-J.; Jeon, B.-G. Terminal Differentiation into Adipocyte and Growth Inhibition by PPARγ Activation in Human A549 Lung Adenocarcinoma Cells. Anim. Cells Syst. 2020, 24, 329–340. [Google Scholar] [CrossRef]
- Sanati, M.; Afshari, A.R.; Kesharwani, P.; Sukhorukov, V.N.; Sahebkar, A. Recent Trends in the Application of Nanoparticles in Cancer Therapy: The Involvement of Oxidative Stress. J. Control. Release 2022, 348, 287–304. [Google Scholar] [CrossRef]
- Yang, Y.; Karakhanova, S.; Hartwig, W.; D’Haese, J.G.; Philippov, P.P.; Werner, J.; Bazhin, A.V. Mitochondria and Mitochondrial ROS in Cancer: Novel Targets for Anticancer Therapy. J. Cell. Physiol. 2016, 231, 2570–2581. [Google Scholar] [CrossRef]
- Burke, P.J. Mitochondria, Bioenergetics and Apoptosis in Cancer. Trends Cancer 2017, 3, 857–870. [Google Scholar] [CrossRef]
- Yoon, M.J.; Kang, Y.J.; Lee, J.A.; Kim, I.Y.; Kim, M.A.; Lee, Y.S.; Park, J.H.; Lee, B.Y.; Kim, I.A.; Kim, H.S.; et al. Stronger Proteasomal Inhibition and Higher CHOP Induction Are Responsible for More Effective Induction of Paraptosis by Dimethoxycurcumin than Curcumin. Cell Death Dis. 2014, 5, e1112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, F.; Ng, T.B. Interrelationship among Paraptosis, Apoptosis and Autophagy in Lung Cancer A549 Cells Induced by BEAP, an Antitumor Protein Isolated from the Edible Porcini Mushroom Boletus Edulis. Int. J. Biol. Macromol. 2021, 188, 313–322. [Google Scholar] [CrossRef]
- Boehnke, N.; Straehla, J.P.; Safford, H.C.; Kocak, M.; Rees, M.G.; Ronan, M.; Rosenberg, D.; Adelmann, C.H.; Chivukula, R.R.; Nabar, N.; et al. Massively Parallel Pooled Screening Reveals Genomic Determinants of Nanoparticle Delivery. Science 2022, 377, eabm5551. [Google Scholar] [CrossRef] [PubMed]
- Bangham, A.D.; Horne, R.W. Negative Staining of Phospholipids and Their Structural Modification by Surface-Active Agents as Observed in the Electron Microscope. J. Mol. Biol. 1964, 8, 660–668. [Google Scholar] [CrossRef]
- Mariani, D.; Ghasemishahrestani, Z.; Freitas, W.; Pezzuto, P.; Costa-da-Silva, A.C.; Tanuri, A.; Kanashiro, M.M.; Fernandes, C.; Horn, A.; Pereira, M.D. Antitumoral Synergism between a Copper(II) Complex and Cisplatin Improves In Vitro and In Vivo Anticancer Activity against Melanoma, Lung and Breast Cancer Cells. Biochim. Biophys. Acta BBA Gen. Subj. 2021, 1865, 129963. [Google Scholar] [CrossRef]
- Yilmazer, A. Cancer Cell Lines Involving Cancer Stem Cell Populations Respond to Oxidative Stress. Biotechnol. Rep. 2018, 17, 24–30. [Google Scholar] [CrossRef]
- Das, S.; Das, J.; Samadder, A.; Boujedaini, N.; Khuda-Bukhsh, A.R. Apigenin-Induced Apoptosis in A375 and A549 Cells Through Selective Action and Dysfunction of Mitochondria. Exp. Biol. Med. 2012, 237, 1433–1448. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Feng, L.; Yue, Q.; Wu, W.; Guan, S.; Jiang, B.; Yang, M.; Liu, X.; Guo, D. Paraptosis Accompanied by Autophagy and Apoptosis Was Induced by Celastrol, a Natural Compound with Influence on Proteasome, ER Stress and Hsp90. J. Cell. Physiol. 2012, 227, 2196–2206. [Google Scholar] [CrossRef]
- Sperandio, S.; Poksay, K.; De Belle, I.; Lafuente, M.J.; Liu, B.; Nasir, J.; Bredesen, D.E. Paraptosis: Mediation by MAP Kinases and Inhibition by AIP-1/Alix. Cell Death Differ. 2004, 11, 1066–1075. [Google Scholar] [CrossRef]
- Sperandio, S.; De Belle, I.; Bredesen, D.E. An Alternative, Nonapoptotic Form of Programmed Cell Death. Proc. Natl. Acad. Sci. USA 2000, 97, 14376–14381. [Google Scholar] [CrossRef]
- Klaunig, J.E. Oxidative Stress and Cancer. Curr. Pharm. Des. 2019, 24, 4771–4778. [Google Scholar] [CrossRef]
- Ghosh, K.; De, S.; Das, S.; Mukherjee, S.; Sengupta Bandyopadhyay, S. Withaferin A Induces ROS-Mediated Paraptosis in Human Breast Cancer Cell-Lines MCF-7 and MDA-MB-231. PLoS ONE 2016, 11, e0168488. [Google Scholar] [CrossRef]
- Chen, X.; Chen, X.; Zhang, X.; Wang, L.; Cao, P.; Rajamanickam, V.; Wu, C.; Zhou, H.; Cai, Y.; Liang, G.; et al. Curcuminoid B63 Induces ROS-Mediated Paraptosis-like Cell Death by Targeting TrxR1 in Gastric Cells. Redox Biol. 2019, 21, 101061. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, X.; Liang, J.; Fang, H.; Huang, H.; Zhao, Y.; Chen, X.; Ma, S. Chalcomoracin Inhibits Cell Proliferation and Increases Sensitivity to Radiotherapy in Human Non-Small Cell Lung Cancer Cells via Inducing Endoplasmic Reticulum Stress-Mediated Paraptosis. Acta Pharmacol. Sin. 2020, 41, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Jeong, H.; Yu, S.-W. Autophagy as a Decisive Process for Cell Death. Exp. Mol. Med. 2020, 52, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kim, W.; Kim, D.R. Autophagy in Cell Survival and Death. Int. J. Mol. Sci. 2023, 24, 4744. [Google Scholar] [CrossRef]
- Liu, W.-J.; Pan, P.; Sun, Y.; Wang, J.; Zhou, H.; Xie, X.; Duan, Z.; Dong, H.; Chen, W.; Zhang, L.; et al. Deferoxamine Counteracts Cisplatin Resistance in A549 Lung Adenocarcinoma Cells by Increasing Vulnerability to Glutamine Deprivation-Induced Cell Death. Front. Oncol. 2022, 11, 794735. [Google Scholar] [CrossRef]
- Ye, X.; Zheng, C.; Chen, J.; Gao, Y.; Murray, C.B. Using Binary Surfactant Mixtures To Simultaneously Improve the Dimensional Tunability and Monodispersity in the Seeded Growth of Gold Nanorods. Nano Lett. 2013, 13, 765–771. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruzzolini, J.; Anceschi, C.; Albino, M.; Balica, E.; Muzzi, B.; Sangregorio, C.; Frediani, E.; Formica, N.; Margheri, F.; Chillà, A.; et al. Targeted Cancer Therapy with Gold–Iron Oxide Nanourchins: Inducing Oxidative Stress, Paraptosis, and Sensitizing Tumor Cells to Cisplatin. Antioxidants 2025, 14, 422. https://doi.org/10.3390/antiox14040422
Ruzzolini J, Anceschi C, Albino M, Balica E, Muzzi B, Sangregorio C, Frediani E, Formica N, Margheri F, Chillà A, et al. Targeted Cancer Therapy with Gold–Iron Oxide Nanourchins: Inducing Oxidative Stress, Paraptosis, and Sensitizing Tumor Cells to Cisplatin. Antioxidants. 2025; 14(4):422. https://doi.org/10.3390/antiox14040422
Chicago/Turabian StyleRuzzolini, Jessica, Cecilia Anceschi, Martin Albino, Elena Balica, Beatrice Muzzi, Claudio Sangregorio, Elena Frediani, Noemi Formica, Francesca Margheri, Anastasia Chillà, and et al. 2025. "Targeted Cancer Therapy with Gold–Iron Oxide Nanourchins: Inducing Oxidative Stress, Paraptosis, and Sensitizing Tumor Cells to Cisplatin" Antioxidants 14, no. 4: 422. https://doi.org/10.3390/antiox14040422
APA StyleRuzzolini, J., Anceschi, C., Albino, M., Balica, E., Muzzi, B., Sangregorio, C., Frediani, E., Formica, N., Margheri, F., Chillà, A., Fibbi, G., & Laurenzana, A. (2025). Targeted Cancer Therapy with Gold–Iron Oxide Nanourchins: Inducing Oxidative Stress, Paraptosis, and Sensitizing Tumor Cells to Cisplatin. Antioxidants, 14(4), 422. https://doi.org/10.3390/antiox14040422